
Abstract
We evaluated the behavior and skin temperature of dual-purpose cattle that grazed pastures having high (HC), low (LC), and no (NC) tree cover during the rainy and dry seasons in the hot and humid tropics of Mexico. We observed twenty-four adult cows (eight per treatment) over 24 days during each season, recording skin temperature and the time related to different daily activities. Across treatments, cows spent the same amount of time foraging during the rainy season (P > 0.05), but cows under HC spent less time during the dry season (P < 0.0001). During the rainy season, cows under HC showed more motivation to continue grazing than becoming restless or beginning rumination (P < 0.001) or roaming more than in other treatments (P < 0.001). During the dry season, cows under HC and LC also had less probability of initiating rest than NC (P < 0.001). Cows under HC had greater motivation to transition from grazing to roaming and less incentive to pass from rumination to rest than cows under LC and NC (P < 0.001). The frequency of water consumption was greater during the dry season (P < 0.001) and consistently high under NC (P < 0.0001). Skin temperature did not differ among treatments during the rainy season (P = 0.261), but during the dry season, it was greater under NC (P < 0.001). Tree cover improves cow behaviors by increasing the impetus to graze and perform daily activities, which contributes to reduced skin temperature during hotter seasons.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Livestock production in Mexico is particularly prominent in the dry and humid tropical zones (Enriquez-Quiroz et al. 2021), where livestock depends largely on pastures as sources of forage, generally grass monocultures that lack tree cover (Orantes-Zebadúa et al. 2014). In these regions, the environmental temperatures, the heat reflected from the soil, and the relative humidity frequently surpass the capacity of the mechanisms that cattle possess to dissipate body heat (Perez et al. 2008). The environmental temperature can negatively influence cattle welfare (Lees et al. 2019); higher than 35 °C affects the thermoregulatory systems of animals, causing increases in their body temperature and, consequently, heat stress. This process leads to a reduction in food consumption and the amount of time spent searching for food. This condition also compels animals to move their grazing times to nighttime hours, when the temperatures are lower (De Elia 2002), ultimately affecting indicators of animal production (Fisher et al. 2008) and reproduction (Blackshaw and Blackshaw 1994; Hall 2000; Wheelock et al. 2006; Lees et al. 2019).
At high temperatures, direct radiation leads animals to adopt physiological and behavioral measures to maintain homeothermy (Souza et al. 2010). Tree shade in grazing areas improves the microclimate (Betancourt et al. 2003; Perez et al. 2008), aiding livestock to regulate body temperature and reduce energy expenditure favoring choices for pasture activities that lead to the use of the present conditions for comfort (Pérez et al. 2008; Vieira-Junior et al. 2019).
Despite these benefits, knowledge regarding changes in the behavior and physiology of cattle under natural shade under warm weather conditions is scarce. Thus, the objective of the present investigation was to study the behavior of dual-purpose cows in tropical pastures with varied tree cover under high temperatures and relative humidity. A better understanding of this interaction would facilitate decision-making regarding the management of cows in tropical grasslands. The hypothesis was that cows grazing areas with more tree cover would maintain a lower body temperature during the day. Consequently, they would have more time for feeding, as their grazing schedules redistributhomogeneously throughout the day over hot periods when cows suffer thermal stress.
Materials and methods
Study site
The study site is in central Veracruz State, Mexico, which has a hot and humid climate. Its annual mean temperature is 23.5 °C, with 1991 mm of annual precipitation and a mean relative humidity of 79% (García 1981; SMN 2016). The soils are acidic, infertile, reddish-brown, and classified as Ultisols. An underground layer is semi-permeable to water, resulting in seasonal flooding within the root zone (INEGI 2008). The original vegetation was a seasonal semi-evergreen tropical forest dominated by trees reaching 40 m height (Cochrane 1984).
Seasons and climate conditions during the study
The study area presents three annual climatic seasons: rainy (July–October), windy (November–February), and dry (March–June). The experiment occurred during the rainy (October 26–November 18, 2010) and dry seasons (May 13–June 5, 2011). We obtained the records of air temperature, relative humidity, and solar radiation from an automatic weather station 400 m away from the experiment site. The weather station recorded the variables every hour. A single weather station does not allow for a differentiation between the environmental conditions of pasture treatments. However, the pastures share the same soil type, and the landscape is similar. During the rains, the average ambient temperature in the experimental area was 28.5 °C, with an average relative humidity of 48.7%, while solar radiation was 840 W m−2. During the drought, the average environmental temperature was 35.3 °C, with an average relative humidity of 43.6% and solar radiation of 1037.4 W m−2. The dry season exhibits extreme climatic conditions compared to the rainy season climate.
Animals, treatments, and pastures
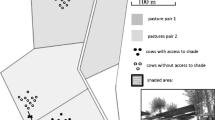
Twenty-four adult cows (Bos taurus L.) having 555.4 ± 77.6 kg of live weight and average milk production of 7.3 ± 4.2 L day−1, were randomly allocated among three treatments (eight cows per treatment): pastures having high tree cover (HC), low tree cover (LC) and no tree cover (NC), with two replicates per treatment (n = four cows). Each treatment contained a 4-ha pasture divided into four 1-ha paddocks, with two paddocks per replicate. Available biomass in the paddocks ranged between 5083 and 8853 kg DM ha−1 during the 2010 rainy season and between 1479 and 4980 kg DM ha−1 during the 2011 dry season across paddocks. These amounts allowed cows to use each paddock continuously for 12-day periods, with each group using the two assigned paddocks during the 24-day experiment periods each season. Only the HC pastures had bushy vegetation. The cover of this stratum was 3.06% and 1.55% during the rainy and dry seasons, respectively. The species that dominated this stratum during the rainy season were Mimosa pigra L. (32.6%), Scoparia dulcis L. (13.5%), Stemmadenia donnell-smithii (Rose ex Donn. Sm.) Woodson (14.9%), and Uncaria tomentosa (Willd. ex Schult.) DC (17.4%). During the dry season, this composition changed slightly but was dominated by the same species, with slight changes in the presence of the less abundant species.
We exposed the cows to the light of glare-reduction frosted lamps for 10 days before the experimental observations began so they got accustomed to the artificial light and the observers (Hubert et al. 2017). The observers stood between two treatment paddocks to record the behavior of all experimental units. We observed only four cows on the pasture from 9:00 to 12:00 because the others were at the milking shed. Otherwise, the cows remained in the field.
Tree canopy
Tree species greater than 3 m in height were identified and quantified for tree density and relative composition in each treatment. The index of tree canopy density was determined only for HC and LC at the beginning of each season using the LAI-2000 (LI-COR Biosciences, Nebraska, USA). Twenty-four readings between 12:00 and 14:00 h were systematically located along a transect in a replicate pasture chosen randomly from each treatment.
Cow haircoat temperature
During the two experiment periods (rainy and dry seasons), we recorded haircoat surface temperatures for all cows using an infrared thermometer (Raytek, Model MT4, Perfoparts S.A. de CV, Mexico, DF). The thermometer placement was 10–20 cm from the cow without disturbing its activity. This form of temperature recording is less invasive than other ways of taking body temperature, allowing more animals to be measured more rapidly with reduced stress. The readings were recorded every other day, during daylight hours when the temperatures were highest (12:00–14:00 h) (Pérez et al. 2008).
The Heat Load Index (HLI, Gaughan et al. 2008) integrates the combined effects of air temperature, relative humidity, and wind speed upon the capacity of the animal to dissipate heat energy to its surrounding atmosphere. Then, we used it to characterize the difference.
Cow behavior
Cows were observed every 10 min, 24 h a day (Gary et al. 1970), every other day during the two 24-days experiment periods. The times and hours that the cows dedicated to grazing (including browsing in the HC treatment), resting, roaming, ruminating, and drinking water were estimated, with all activities considered as mutually exclusive. Visual scanning was used as the method of animal observation, for which each scan was converted to 10 min periods after which the times were added by day and hours that each cow used for each activity (Penning and Rutter 2004). Behavior was recorded by four trained observers that rotated among treatments on a daily basis.
Statistical analysis
Times for grazing (including browsing in the HC treatment), resting, ruminating, roaming, drinking water, and skin temperature data were analyzed as a completely randomized design with two replicates using the procedure PROC MIXED in the Statistical Analysis System, version 4.3.0 (SAS Inc., 2010), and a compound symmetry covariance structure (Littell et al. 1998). The model effects included treatment, season, the interaction of treatment with the season, and cow nested in treatment as the random effect. The model effects were significant when P < 0.05. The least-squares-means option in PROC MIXED tested the differences between treatment means. All cow behaviors during the day and their distribution over a 24-h day were analyzed using descriptive statistics and graphics using Statistic, v. 6 (Statistica Inc., 2011).
Times spent in the different activities were analyzed using multi-state models, a process for analyzing phenomena occurring over time that can be characterized as an assembly of discreet states. Such data structures arise when we observe qualitative responses over time.
A multi-state model is an assembly of risk rates, \(\lambda_{{{\varvec{h}}i}} \left( t \right)\) in which h = 1, …, k indicates the type of transition and i = 1, …, n indicates the individuals. The risk rates describe the duration between transitions, between behavior, and is modeled in a multiplicative form as:
where \(\eta_{{{\varvec{h}}i}} \left( t \right)\) is an additive predictor that in this work consists of the following components:
-
A non-parametric base effect common to all observations denoted by \(g_{{\mathbf{0}}}^{{\left( {\varvec{h}} \right)}} \left( t \right)\) that depends on time.
-
Parametric effects from treatments HC, LC and NC denoted by \(\gamma_{HC}^{\left( h \right)} = 0\), \(\gamma_{LC}^{\left( h \right)}\) and \(\gamma_{NC}^{\left( h \right)}\), respectively. Note that HC is taken as a reference for comparison, such that \(\gamma_{LC}^{\left( h \right)}\) and \(\gamma_{NC}^{\left( h \right)}\) are interpreted as deviations concerning reference treatment effects.
-
A random effect \(b_{i}^{{\left( {\varvec{h}} \right)}}\) is included for implicit consideration of the correlation among observations within the same individual.
To model the base effect \(g_{{\mathbf{0}}}^{{\left( {\varvec{h}} \right)}} \left( t \right)\), penalized splines provided a parsimonious and flexible focus to represent smooth functions (Eilers and Marx 1996).
Cattle behaviors were categorized into four states: grazing (Gr), resting (Re), roaming (Ro), and ruminating (Ru); drinking was not included because of the great number of zeros that kept the data from being normally distributed. The transitions among these states were GrRe, GrRu, GrRo, ReGr, ReRu, RuGr, RuRe and RoGr, so that, for example, GrRe defines the transition from grazing to resting. The observations were performed every 10 min, but we assumed that the process is continuous over time. Our objective was to describe the behavioral dynamics of the animals. When there were insufficient observations for some transitions among states (behaviors) during the study, we assumed that there was no transition among the corresponding states. Thus, the estimated predictors were:
Predictor estimates were transformed using antilogarithms into probabilities of transition estimates.
To estimate the parametric and non-parametric effects, the method proposed by Kneib and Hennerfeind (2006) was applied, using the statistical software BayesX (2012) (http://www.stat.uni-muenchen.de/~bayesx/bayesx.html).
Results
Weather conditions during the experiment
During the rainy season, the average study site temperature was 28.6 ± 0.74 °C, with a mean relative humidity of 48.8 ± 3.6% and solar radiation of 840.0 ± 84.9 W m−2. During the dry season, the average temperature was 35.2 ± 0.8 °C, with a mean relative humidity of 43.6 ± 3.0%, and solar radiation of 1037.4 ± 86.3 W m−2. During the dry season, the climatic conditions were more extreme, resulting in a higher value representing the combined effects of air temperature and humidity associated with the level of thermal stress (RH/T index; Gaughan et al. 2008) (79.28) than during the rainy season (71.66).
Tree canopy characteristics
We identified nine tree species in the arboreal stratum of pastures across all treatments, of which the most abundant were Quercus rugosa Née, Croton draco Schltdl. & Cham., and Guazuma ulmifolia Lam. Tree density was 12.5 trees−ha in the HC treatment, 1.8 in the LC treatment, and 2.6 in the NC treatment. Tree density in these last two treatments was similar, but in the NC treatment the trees were aggregated near the entrance to the pasture and along the fence, while in the LC treatment they were dispersed within the pasture, with few along the fence, thus presenting a more uniform pattern of shade across the pasture.
The tree canopy density during the rainy season was 1.0 and 0.2 m−2 in the HC and LC treatments, respectively, while during the dry season, the density was 0.7 m−2 in the HC treatment and 0.3 m−2 in the LC treatment. We did not measure this variable under NC.
Cow skin temperature and behavior
The skin temperature of the cows did not differ among treatments in the rainy season (P = 0.261). During the dry season, however, skin temperature diminished as tree cover increased (P = 0.001) and was least in the HC treatment (Table 1).
Time dedicated by cattle to different activities
Time dedicated to grazing, ruminating, and roaming differed among cows in the three treatments as a function of season (P < 0.0001). During the rainy season, cows dedicated between 8.9 and 9.2 h per day to grazing with no differences between treatments (P > 0.05; Table 2), while during the dry season, cows in the LC and NC treatments dedicated the same amount of time to this activity (P = 0.699, Table 2). Yet, this value was higher than for cows in treatment HC (P < 0.0001).
During the rainy season, cows under LC and HC dedicated more time to ruminating than those under NC (P < 0.05, Table 2). The cows under HC roamed more during the rainy season (P < 0.0001, Table 2), and cows under tree cover (LC and HC) roamed similarly during the dry season.
HC y LC in resting and drinking differed (P < 0.0001). Cows under NC spent more time resting and drinking (P = 0.0001, Table 3). The time spent drinking water during the rainy season was 0.09 ± 0.03 h, and during the dry season was 0.30 ± 0.06 h (P < 0.0001). Cows grazing without tree cover drank more frequently (P = 0.0001; Table 3).
Cow activity schedules during the day
During the rainy season, all cows had three peaks of greater grazing activity over 24 h periods (Fig. 1). Although in general terms, these peaks had the same schedules, the cows under LC showed greater activity during the first peak and less during the first hours (11:00–14:00 h) of the second and third peaks; time they replaced during evening hours when the temperature declined (19:00–23:00 h; Fig. 1a). Overall, during the dry season cows grazed with greater uniformity between treatments throughout the day (Fig. 1a) than during the rainy season, and cows under HC tended to graze for more time than those in other treatments.

Distribution of daily times spent grazing (a), resting (b), and ruminating (c) by cows (Holstein × Cebu) grazing in pastures having high (HC), low (LC) and no tree cover (NC), during the rainy and dry seasons
The longest rest period was in the early morning and extended throughout the day, but with reduced frequency (Fig. 1b). Cows under NC rested more frequently throughout the day, especially during greater heat. Those under HC displayed a reduced rest frequency distributed more homogeneously throughout the day. Rumination time was distributed roughly similarly among the three treatments throughout the day, but it was more frequent under LC and NC during hours of greater heat (8:00–16:00 h; Fig. 1c).
Roaming frequency increased between 11:00 and 18:00 h for cows under HC. Roaming was distributed more or less homogeneously during the day but moved slightly toward 14:00–20:00 h, with somewhat greater frequency under HC (Fig. 2a). Drinking water occurred with a higher frequency during hotter hours in both seasons, except under HC, where the greater frequency of drinking water was observed during the afternoon (Fig. 2b).

Distribution of daily times spent roaming (a) and drinking water (b) by cows (Holstein x Cebu) grazing in pastures having high (HC), low (LC) and no tree cover (NC), during the rainy and dry seasons
Probabilities for transitions in behavior during the rainy season
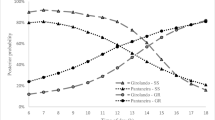
Tree cover influenced the probability that cows transitioned between activities (Fig. 3). Cows under HC showed less likelihood of passing from resting to grazing than under LC or NC (P < 0.001; Fig. 4a) and from grazing to ruminating (P < 0.001; Fig. 3b). However, cows showed greater motivation to pass from grazing to roaming throughout the experiment than in other treatments (P < 0.001; Fig. 3c, d). Although the effects of treatments LC and NC did not compare, their probabilities for all transitions were similar during this season.

Probabilities of transitioning from grazing to resting (a), ruminating (b) or roaming (c), and from roaming to grazing (d), by cows (Holstein × Cebu) grazing in pastures having high (HC), low (LC) and no tree cover (NC), during the rainy season

Probabilities of transitioning from resting to ruminating (a) or grazing (b), and from ruminating to resting (c) or grazing (d) by cows (Holstein × Cebu) grazing in pastures having high (HC), low (LC) and no tree cover (NC), during the rainy season
Cows under HC and LC had the same probability of passing from roaming to grazing (P > 0.05), while cows under NC were different (P = 0.031; Fig. 3d); an answer congruent with the behaviors that showed a reduced probability of transition from roaming to grazing.
Once resting, cows under HC showed a lower probability of initiating rumination than those under LC or NC (P < 0.001, Fig. 4a). However, cows under HC and LC showed equal chances of starting grazing (P = 0.201; Fig. 4b) but differed from cows under NC. The above does not imply a greater or lesser time for grazing (Table 3) (P < 0.001). Yet, the magnitude of the tendencies was similar among the three treatments (Fig. 4b). Similarly, the probability for cows under HC to pass to resting and to grazing after ruminating was lower than under LC (P < 0.01) or NC (P < 0.001, Fig. 4c, d).
Probabilities for transitioning in behavior during the dry season
Cows under HC and LC showed similar probabilities of passing from resting to grazing (P = 0.290). However, cows under NC were more motivated to do so than in the other treatments (P < 0.001; Fig. 5a). Regardless of treatment, all cows had the same probability of transitioning from grazing to ruminating during this season (P > 0.05; Fig. 5b). Yet, cows under HC were motivated to transition to roaming from grazing more strongly than those under LC (P = 0.001) and NC (P < 0.001; Fig. 5c). In all treatments, cows had the same motivation to continue grazing while roaming within the pastures (P > 0.05; Fig. 5d).

Probabilities of transitioning from grazing to resting (a), ruminating (b), or roaming (c), and from roaming to grazing (d) by cows (Holstein × Cebu) grazing in pastures having high (HC), low (LC) and no tree cover (NC), during the dry season
The probability of initiating rumination after resting was equal in all treatments (P > 0.05; Fig. 6a). However, cows under HC had the same motivation as those under LC (P = 0.834) to resume grazing from resting, but different and greater than those under NC (P = 0.007; Fig. 6b). As well, the probability of continuing to rest after terminating rumination was less under HC than under LC (P < 0.001) and NC (P < 0.001) (Fig. 6c). Finally, in cows that were ruminating, the motivation to reinitiate grazing was higher under LC (P < 0.001) and NC (P < 0.001) than under HC (Fig. 6d).

Probabilities of transitioning from resting to ruminating (a), or grazing (b), and from ruminating to resting (c), or grazing (d) for cows (Holstein × Cebu) grazing in pastures having high (HC), low (LC) and no tree cover (NC), during the dry season
Discussion
During both seasons of the study, climate conditions were harsh for cattle since the RHT indexes were at the danger level according to the Livestock Weather Safety Index (Johnson 1994; Valdivia-Cruz et al. 2021). Nevertheless, ambient temperatures and solar radiation were more benign during the rainy season. Tree canopy density was relatively low during both seasons in the HC and LC treatments. However, those levels provided adequate shade compared to the NC treatment, where grazing did not allow transiting within a sunny–shady pattern. Botanical composition and forage availability of the herbaceous and bushy vegetation ensured different grazing environments for each treatment and an adequate supply of dry matter for intake, avoiding shortages within the regional conditions where forage availability and quality fluctuate throughout the seasons (Jarillo-Rodriguez et al. 2011; Reis et al. 2021; Chebli et al. 2022). These pasture conditions ensured that potential behavioral differences among cows would be mainly attributed to treatment effects.
The skin temperature of the cows during the rainy season among the treatments was lower than during the dry season, because the body surface is more exposed and sensitive to changes in ambient temperatures (Lysyk 2008). Still, cows under HC and LC maintained their skin temperatures under 39 °C which suggests that shade helped cows to maintain surface (and likely body temperatures as well) at lower levels, but within a normal range during this season, as with other kinds of shade (Schütz et al. 2009).
Even though canopy density decreased during the dry season, the existing tree cover provided adequate shade across pastures. Canopy density reduced the passage of solar rays to the ground and to the skin surface of the cows, thus reducing extreme body temperature (Finch 1986). During the dry season, the skin temperature of the cows was 1.4 °C more under NC than under HC, although greater differences have been found in other studies where body temperatures were measured (Betancourt et al. 2003; Brown-Brandl et al. 2006; Kendall et al. 2006). We could not measure internal body temperatures during the study, although we suspect that the effect of shade on body temperature was positive.
Similar grazing times among treatments during the rainy season might indicate that more homogenous environmental conditions for cows override any shading effect. The most prolonged grazing periods occurred between 12:00 and 2:00 (principally in LC), between 6:00 and 9:00, and between 16 and 20 h. However, times dedicated to grazing by cows under LC and NC during the dry season could suggest shifting grazing bouts to nighttime when temperatures are higher improves comfort during grazing.
Grazing cattle modified the frequencies of some behaviors throughout the day between seasons due to differences in temperature, relative humidity, and solar radiation (Blackshaw and Blackshaw 1994; Schütz et al. 2009; Mishra 2021). In the present research, from the rainy to dry season, the daily temperatures increased by 6.6 °C, and relative humidity and solar radiation varied by 5.2% units and 197 W m−2 higher, respectively. In addition, tree cover in the HC and LC treatments decreased due to partial leaf senescence, so conditions during the dry season were more extreme than during the rainy season.
Nevertheless, the environmental conditions in the study site were extreme for the cows at all times (García and Wright 2007). Tree cover in the HC treatment promoted slight differences in the frequency and timing of all cow activities. However, the cows modify their behaviors (Schütz et al. 2009) and activate physiological mechanisms such as sweating and panting, as reported by Brown-Brandl et al. (2006), following tree cover to keep body temperatures below critical thresholds and to improve pasture use.
We observed that cows grazing under LC and HC had a lower probability of transitioning from feeding to resting or rumination. These cows had more transitions between grazing and roaming and vice versa, denoting a higher activity during the day than cows under NC during both seasons. As well, cows under HC dedicated more time to grazing (during the rainy season) and roaming (both seasons) between 12:00 and 18:00 h, probably due to the environmental comfort provided by shade, thus reducing water consumption and dedicating more time to rumination at night. The above-described situation contrasts with the behavior of cows under LC and NC, where grazing and roaming were observed mainly during the afternoon and night, increasing time dedicated to resting, ruminating, and drinking water during hours of the day when temperatures were higher. These cows also had a greater probability of passing from resting to ruminating (and from ruminating to resting).
In contrast, cows under HC tended to favor stable behavior performance. Heifers with less offered shadow grazed for more extended periods. In comparison, with a greater shade on offer, the animals rested and ruminated longer. Therefore, different tree densities promoted beneficial changes in the daytime grazing behavior of heifers (Carvalho de Oliveira et al. 2021).
Different rumination times among treatments during both seasons might be more related to pasture forage quality than tree cover effects. During the rainy season, forage cover and sward height across the NC pastures were more uniform and shorter, maintaining higher forage quality. During the following season, an unclear pattern in rumination suggests effects from pasture composition and forage quality (Jarillo-Rodríguez et al. 2011).
The HC showed a greater probability of roaming during both seasons and hours of greater heat, as well as less frequency of drinking. These might be indicators of greater comfort in pastures having shade, as has been found in other investigations using trees or other forms of artificial shade (Betancourt et al. 2003; Pérez et al. 2008; Fisher et al. 2008; Schütz et al. 2009).
Regardless of the season, drinking time also increased as tree cover decreased, a component of the behavioral mechanism to lower body heat (Blackshaw and Blackshaw 1994). Thus, livestock under shade have more comfort and need less water during the day. However, higher temperatures may have caused a considerable increase in water consumed during both seasons (not measured). Still, it could be a confounded response with the intake of fibrous forage having low water content (Pérez et al. 2005, 2008).
Conclusions
Cows grazing under tree cover improved their behavior and motivation to carry out and distribute their daily activities. Under this experiment's conditions, tree cover also reduced their body/skin temperatures in the dry season. However, the function of tree cover in alleviating thermal stress has a more significant impact during the dry season under hot weather conditions, when temperatures and relative humidity are more extreme toward the upper limit.
The behaviors of cows, their greater motivation to graze and roam, and the reduction in their skin temperature, mainly during the dry season, indicate that having trees in pastures and grasslands could lead to decision-making regarding the management of cows in tropical grasslands and, therefore more efficient use of pastures and improvements in animal production.
References
BayesX (2012) BayesX—Bayesian inference in structured additive regression models. Version 2.1 (07.05.2012). GNU Public License, Munich, Germany. http://www.stat.uni-muenchen.de/~bayesx/bayesxnews.html. Accessed Feb 2012
Betancourt K, Ibrahim M, Harvey C, Vargas B (2003) Efecto de la cobertura arbórea sobre el comportamiento animal en fincas ganaderas de doble propósito en Matiguás, Matagalpa, Nicaragua. Agrofor Am 10:47–51
Blackshaw JK, Blackshaw AW (1994) Heat stress in cattle and the effect of shade on production and behavior: a review. Aust J Exp Agric 34:285–295. https://doi.org/10.1071/EA9940285
Brown-Brandl TM, Eigenberg RA, Nienaber JA (2006) Heat stress risk factor of feedlot heifers. Livest Sci 105:57–68. https://doi.org/10.1016/j.livsci.2006.04.025
Carvalho de Oliveira C, Giolo de Almeida R, Karvatte-Junior N, Junqueira VSD, Bungenstab DJ, Villa AF (2021) Daytime ingestive behaviour of grazing heifers under tropical silvopastoral systems: responses to shade and grazing management. Appl Anim Behav Sci 240:1–7. https://doi.org/10.1016/j.applanim.2021.105360
Chebli Y, El Otmani S, Hornick JL, Keli A, Bindelle J, Cabaraux JF, Chentouf M (2022) Forage availability and quality, and feeding behaviour of indigenous goats grazing in a Mediterranean silvopastoral system. Ruminants 2:74–89. https://doi.org/10.3390/ruminants2010004
Cochrane TT (1984) Amazonia computerized overview of its climate, landscape, and soil resources. Interciencia 5(9):98–306
De Elia M (2002) Etología y comportamiento del bovino. Prod Anim, p 4. http://www.produccionanimal.comm. Accessed Sept 2010
Eilers CPH, Marx DB (1996) Flexible smoothing with B-splines and penalties. Stat Sci 11:89–121
Enríquez QJF, Esqueda EVA, Martínez MD (2021) Rehabilitación de praderas degradadas en el trópico de México. Rev Mex Cienc Pecu 12(3):243–260. https://doi.org/10.22319/rmcp.v12s3.5876
Finch VA (1986) Body temperature in beef cattle: its control and relevance to production in the tropics. J Anim Sci 62:531–542. https://doi.org/10.2527/jas1986.622531x
Fisher AD, Roberts N, Bluett SJ, Verkerk GA, Matthews LR (2008) Effects of shade provision on the behavior, body temperature and milk production of grazing dairy cows during a New Zealand summer. N Z J Agric Res 51:99–105. https://doi.org/10.1080/00288230809510439
García E (1981) Modificaciones al sistema de clasificación climática de Köppen, 3rd edn. Instituto de Geografía, UNAM, México
García A, Wright C (2007) Efectos del medio ambiente sobre los requerimientos nutricionales del ganado en pastoreo. Extensión extra ExEx4037-S. Sitio Argentino de Producción Animal. http://agbiopubs.sdstate.edu/articles/ExEx4037-S.pdf. Accessed Oct 2016
Gary LA, Sherritt GW, Hale EB (1970) Behaviour of Charolais cattle on pasture. J Anim Sci 30:203–206. https://doi.org/10.2527/jas1970.302203x
Gaughan JB, Mader TL, Holt SM, Lisle A (2008) A new heat load index form feedlot cattle. J Anim Sci 86:226–234. https://doi.org/10.2527/jas.2007-0305
Hall M (2000) Meet the challenges of heat stress feeding. Hoard’s Dairyman, Fort Atkinson
Hubert DJ, Huhnke RL, Harp SL (2017) Cattle handling safety in working facilities. Oklahoma State University. Id: BAE-1738. https://extension.okstate.edu/fact-sheets/cattle-handling-safety-in-working-facilities.html
INEGI (2008) Municipio Tlapacoyan, Veracruz, México. Medio Físico y Geográfico. http://www.tlapacoyanver.gob.mx. Accessed Aug 2010
Jarillo-Rodríguez J, Castillo-Gallegos E, Flores-Garrido AF, Valles-de la Mora B, Ramírez y Avilés L, Escobar-Hernández R, Ocaña-Zavaleta E (2011) Forage yield, quality and utilization efficiency on native pastures under different stocking rates and seasons of the year in the Mexican humid tropics. Trop Subtrop Agroecosyst 13:417–427
Johnson HD (1994) Animal physiology. In: Handbook of agricultural meteorology. Ed John Griffiths, New York
Kendall PE, Nielsen PP, Webster JR, Verkerk GA, Littlejohn RP, Matthews LR (2006) The effects of providing shade to lactating dairy cows in a temperate climate. Livest Sci 103:148–157. https://doi.org/10.1016/j.livsci.2006.02.004
Kneib T, Hennerfeind A (2006) Bayesian semiparametric multi-state models. Stat Model 8:169–198
Lees AM, Sejian V, Wallage AL, Steel CC, Mader TL, Lees JC, Gaughan JB (2019) The impact of heat load on cattle. Animals (basel) 9(6):322. https://doi.org/10.3390/ani9060322
Littell RC, Henry PR, Ammerman CB (1998) Statistical analysis of repeated measures data using SAS procedures. J Anim Sci 76:1216–1231. https://doi.org/10.2527/1998.7641216x
Lysyk TJ (2008) Effects of ambient temperature and cattle skin temperature on engorgement of Dermacentor andersoni. J Med Entomol 45:1000–1006. https://doi.org/10.1093/jmedent/45.6.1000
Mishra SR (2021) Behavioural, physiological, neuro-endocrine and molecular responses of cattle against heat stress: an updated review. Trop Anim Health Prod 53(3):400. https://doi.org/10.1007/s11250-021-02790-4
Orantes-Zebadúa MA, Platas-Rosado D, Córdova-Avalos V, De los Santos-Lara MC, Córdova-Avalos A (2014) Caracterización de la ganadería de doble propósito en una región de Chiapas, México. Ecosistemas Recur Agropecu 1:49–58
Penning PD, Rutter MS (2004) Ingestive behaviour. In: Penning PD (ed) Herb intake. Handbook, 2nd edn. British Grassland Society, Reading
Pérez OJ, Guillen RJ, Hernández SR, Martínez HPA (2005) Uso de las leguminosas arbustivas en los sistemas de producción animal en el trópico. R. Electron. Vet. REDVET‒ISSN 1695–7504. VI: 1–18
Pérez E, Soca M, Díaz L, Corzo M (2008) Comportamiento etológico de bovinos en sistemas silvopastoriles en Chiapas, México. Past Forr 31:165–170
Reis NS, Ferreira IC, Mazocco LA, Souza ACB, Pinho GAS, da Fonseca-Neto ÁM, Malaquias JV, Macena FA, Muller AG, Martins CF, Balbino LC, McManus CM (2021) Shade modifies behavioral and physiological responses of low to medium production dairy cows at pasture in an integrated crop-livestock-forest system. Animals (basel) 11(8):2411. https://doi.org/10.3390/ani11082411
SAS Inc (2010) Statistical analysis system, enterprise guide v. 4.3.0. SAS Institute Inc, Cary
Schütz KR, Rogers AR, Cox NR, Tucker CB (2009) Dairy cows prefer shade that offers greater protection against solar radiation in summer: shade use, behavior, and body temperature. Appl Anim Behav Sci 116:28–34. https://doi.org/10.10166/japplanim.2008.07.005
SMN (2016) Veracruz normales climatológicas 1951–2010. Comision Nacional Del Agua. http://smn.cna.gob.mx/es/informacion-climatologica-ver-estado?estado=ver. Accessed Nov 2016
Souza WD, Barbosa OR, Marques JDAJ, Gasparino E, Cecato U, Barbero LM (2010) Behavior of beef cattle in silvopastoral systems with eucalyptus. Rev Bras Zootec 39:677–684. https://doi.org/10.1590/S1516-35982010000300029
Statistic (2011) Statistic Inc., v. 6. Stat Soft Inc. 1984‒2003. Tulsa, Oklahoma, USA
Wheelock JB, Sanders SR, Shwartz G, Hernandez LL, Baker SH, McFadden JW, Odens LJ, Burgos R, Hartman SR, Johnson RM, Jones BE, Collier RJ, Rhoads RP, Van Baale MJ, Baumgard LH (2006) Effects of heat stress and rbST on production parameters and glucose homeostasis. J Dairy Sci 89(1):290–291
Valdivia-Cruz JC, Reyes-González JJ, Valdés-Paneque GR (2021) Effect of temperature and humidity index (THI) on the physiological responses of grazing dairy cows. Cuban J Agric Sci 55(1):21–29
Vieira Junior NA, Silva MAA, Caramori PH, Nitsche PR, Corrêa KAB, Alves DS (2019) Temperatura, conforto térmico e comportamento ingestivo animal em sistema silvipastoril. Semin Ciências Agrárias 40(1):403–416. https://doi.org/10.5433/1679-0359.2019v40n1p403
Acknowledgements
The authors thank the Centro de Enseñanza, Investigación y Extensión en Ganadería Tropical of the Universidad Nacional Autónoma de México for providing the facilities to carry out this experiment. We also thank the cattle management team at the same institution for field assistance in livestock handling and fencing.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pérez-Hernández, V.M., López-Ortiz, S., Pérez-Elizalde, S. et al. Effect of tree shade on behavior and haircoat temperature of grazing dual-purpose cows in a hot and humid tropical environment. Agroforest Syst 98, 165–178 (2024). https://doi.org/10.1007/s10457-023-00897-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-023-00897-1