
Impact of Soil Inoculation with Bacillus amyloliquefaciens FZB42 on the Phytoaccumulation of Germanium, Rare Earth Elements, and Potentially Toxic Elements

 ,
,  and
and
Abstract
:1. Introduction
2. Results
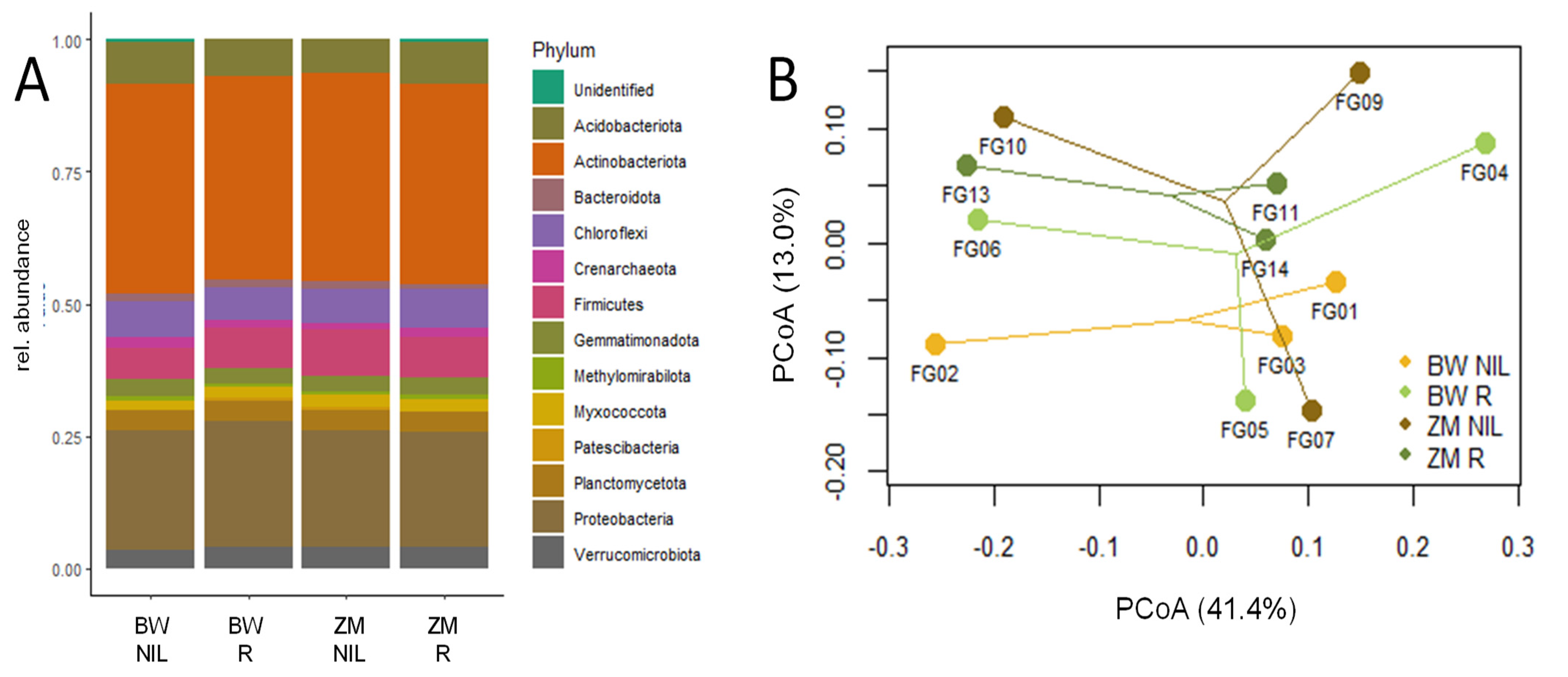
2.1. Effect of Inoculation on Soil Microbial Community Composition and B. amyloliquefaciens Abundance in Soil
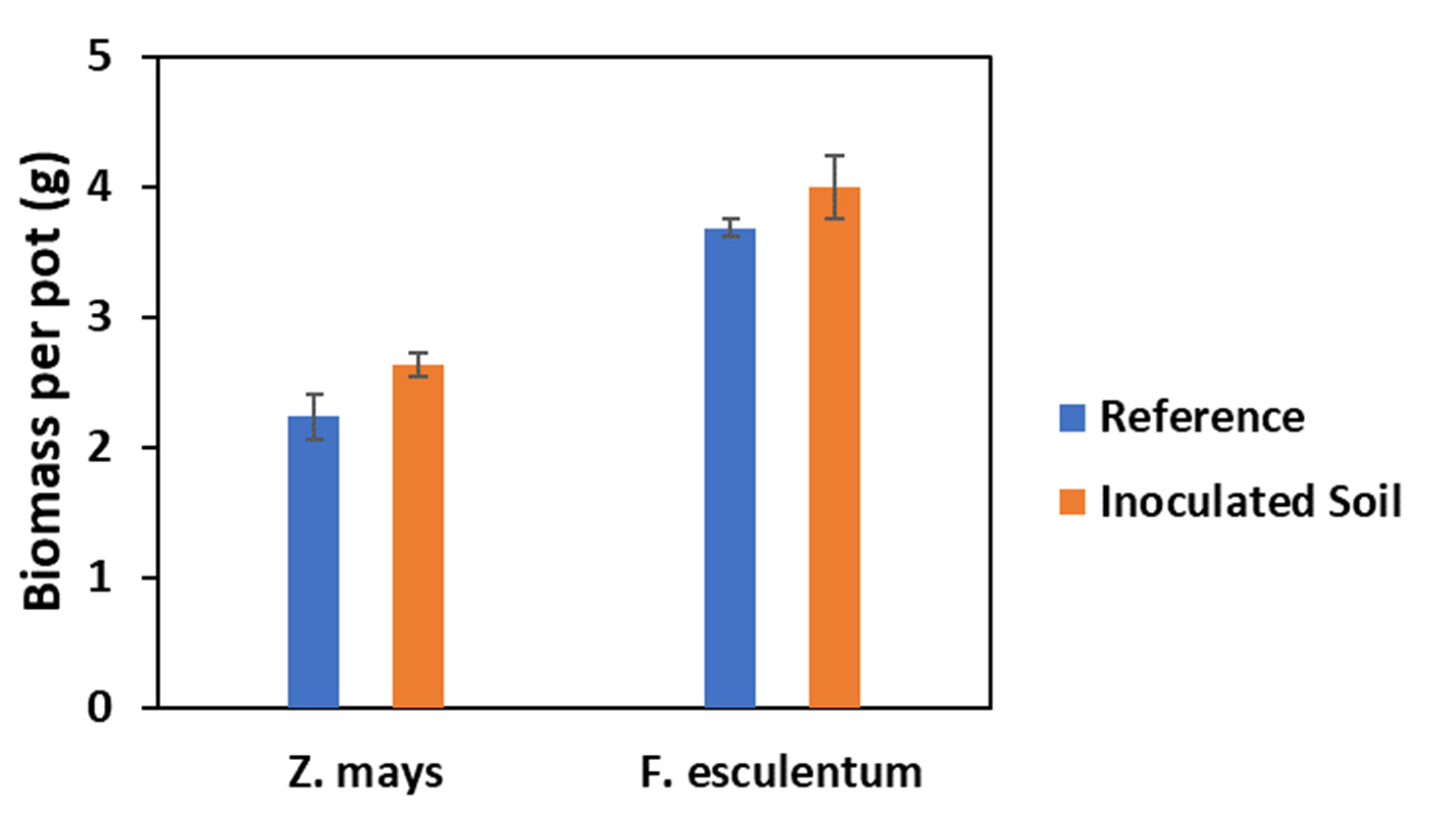
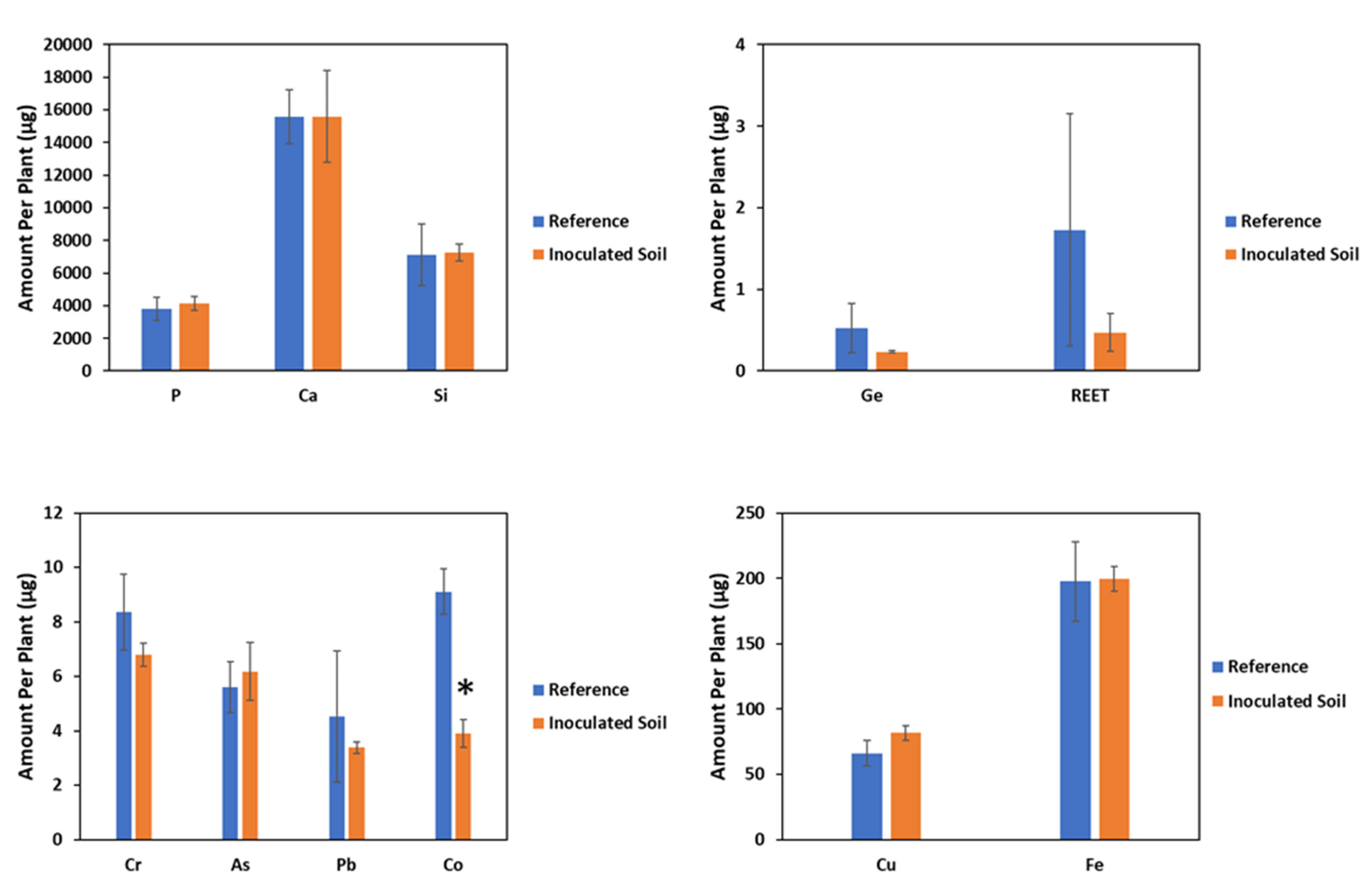
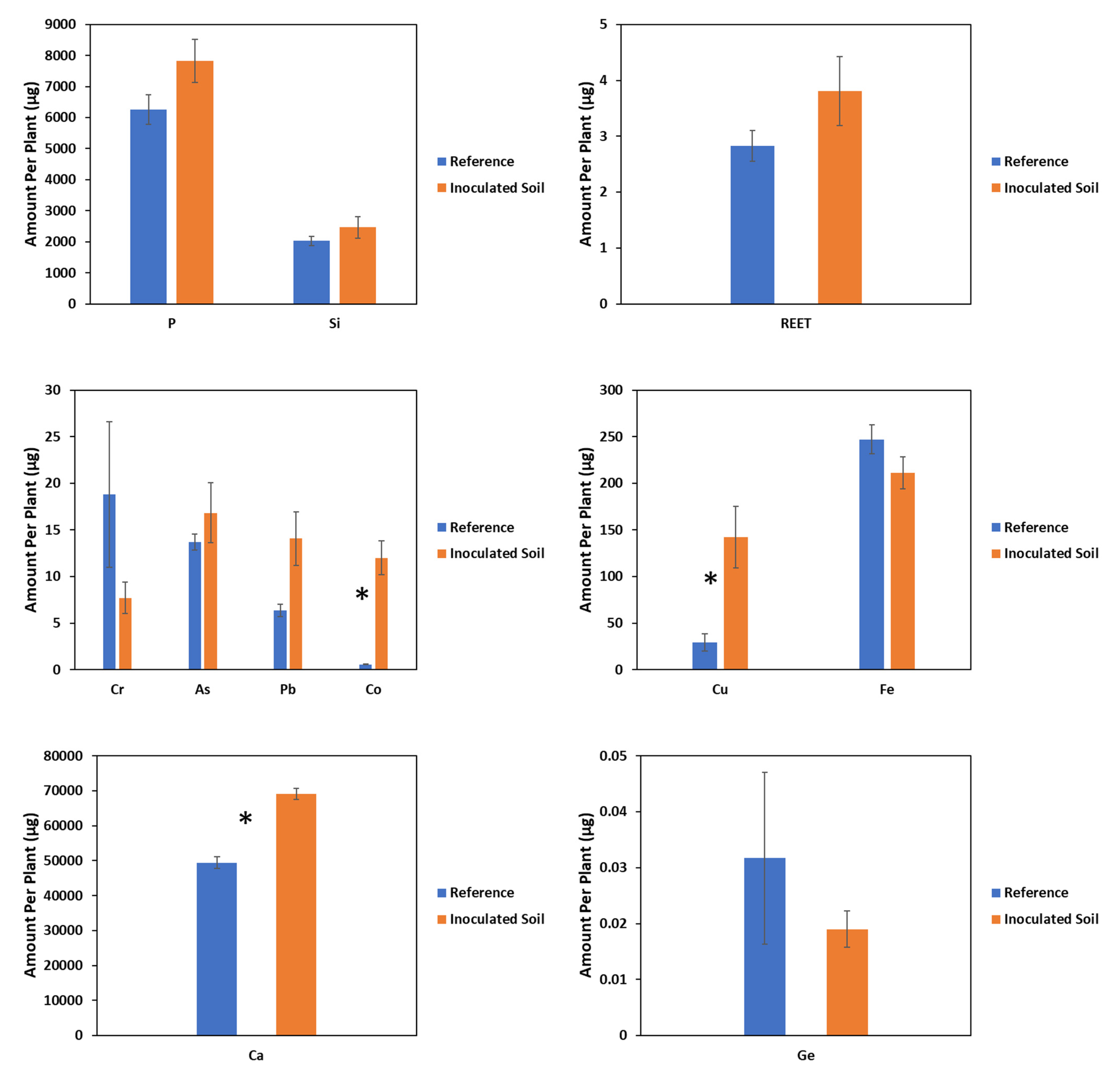
2.2. Effect of PGPR on Shoot Yield and Accumulation of Investigated Elements
3. Discussion
3.1. Effects of Inoculation on Root Colonization, Rhizosphere Bacterial Communities, Nutrient Supply, and Plant Growth
3.2. Effects of Inoculation on Shoot Yield
3.3. Effects of Inoculation on PTE and CRM Accumulation
4. Materials and Methods
4.1. Plant Growth Experiment and Soil Amendment
4.2. Sample Preparation and Analysis
4.2.1. Soil Samples (Before Inoculation)
4.2.2. Plant Samples
4.2.3. Soil DNA Extraction and Illumina Sequencing
4.2.4. Bioinformatics Workflow
4.2.5. Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Turan, M.; Topcuoğlu, B.; Kitir, N.; Alkaya, Ü.; Erçelik, F.; Nikerel, E.; Günes, A. Plant Growth Promoting Rhizobacteria’s (PGPRS) Enzyme Dynamics in Soil Remediation. In Soil Contamination-Current Consequences and Further Solutions; Larramendy, M.L., Soloneski, S., Eds.; IntechOpen: London, UK, 2016; pp. 209–231. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.-S.; Yamasaki, S.; Kimura, K. Rare Earth Element Content in Various Waste Ashes and the Potential Risk to Japanese Soils. Environ. Int. 2001, 27, 393–398. [Google Scholar] [CrossRef]
- Zhao, C.; Duan, D.; Li, Y.; Zhang, J. Rare Earth Elements in No. 2 Coal of Huangling Mine, Huanglong Coalfield, China. Energy Explor. Exploit. 2012, 30, 803–818. [Google Scholar] [CrossRef] [Green Version]
- Kaartinen, T.; Sormunen, K.; Rintala, J. Case Study on Sampling, Processing and Characterization of Landfilled Municipal Solid Waste in the View of Landfill Mining. J. Clean. Prod. 2013, 55, 56–66. [Google Scholar] [CrossRef]
- Gutiérrez-Gutiérrez, S.C.; Coulon, F.; Jiang, Y.; Wagland, S. Rare Earth Elements and Critical Metal Content of Extracted Landfilled Material and Potential Recovery Opportunities. Waste Manag. 2015, 42, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Wiche, O.; Heilmeier, H. Germanium (Ge) and Rare Earth Element (REE) Accumulation in Selected Energy Crops Cultivated on Two Different Soils. Miner. Eng. 2016, 92, 208–215. [Google Scholar] [CrossRef]
- Hussain, R.; Luo, K. Geochemical Evaluation of Enrichment of Rare-Earth and Critical Elements in Coal Wastes from Jurassic and Permo-Carboniferous Coals in Ordos Basin, China. Nat. Resour. Res. 2020, 29, 1731–1754. [Google Scholar] [CrossRef]
- Adeel, M.; Shakoor, N.; Hussain, T.; Azeem, I.; Zhou, P.; Zhang, P.; Hao, Y.; Rinklebe, J.; Rui, Y. Bio-Interaction of Nano and Bulk Lanthanum and Ytterbium Oxides in Soil System: Biochemical, Genetic, and Histopathological Effects on Eisenia Fetida. J. Hazard. Mater. 2021, 415, 125574. [Google Scholar] [CrossRef]
- Adeel, M.; Shakoor, N.; Ahmad, M.A.; White, J.C.; Jilani, G.; Rui, Y. Bioavailability and Toxicity of Nanoscale/Bulk Rare Earth Oxides in Soil: Physiological and Ultrastructural Alterations in Eisenia fetida. Environ. Sci. Nano 2021, 8, 1654–1666. [Google Scholar] [CrossRef]
- El-Ramady, H. Ecotoxicology of Rare Earth Elements: Ecotoxicology of Rare Earth Elements within Soil and Plant Environments; VDM Verlag Dr. Müller: Saarbrücken, Germany, 2010; ISBN 978-3-639-23680-4. [Google Scholar]
- Franus, W.; Wiatros-Motyka, M.M.; Wdowin, M. Coal Fly Ash as a Resource for Rare Earth Elements. Environ. Sci. Pollut. Res. 2015, 22, 9464–9474. [Google Scholar] [CrossRef] [Green Version]
- Kasowska, D.; Gediga, K.; Spiak, Z. Heavy Metal and Nutrient Uptake in Plants Colonizing Post-Flotation Copper Tailings. Environ. Sci. Pollut. Res. 2018, 25, 824–835. [Google Scholar] [CrossRef] [Green Version]
- Jing, Y.; He, Z.; Yang, X. Role of Soil Rhizobacteria in Phytoremediation of Heavy Metal Contaminated Soils. J. Zhejiang Univ. Sci. B 2007, 8, 192–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwabe, R.; Dittrich, C.; Kadner, J.; Rudi Senges, C.H.; Bandow, J.E.; Tischler, D.; Schlömann, M.; Levicán, G.; Wiche, O. Secondary Metabolites Released by the Rhizosphere Bacteria Arthrobacter Oxydans and Kocuria Rosea Enhance Plant Availability and Soil–Plant Transfer of Germanium (Ge) and Rare Earth Elements (REEs). Chemosphere 2021, 285, 131466. [Google Scholar] [CrossRef]
- Li, F.; Shan, X.; Zhang, T.; Zhang, S. Evaluation of Plant Availability of Rare Earth Elements in Soils by Chemical Fractionation and Multiple Regression Analysis. Environ. Pollut. 1998, 102, 269–277. [Google Scholar] [CrossRef]
- Violante, A.; Cozzolino, V.; Perelomov, L.; Caporale, A.G.; Pigna, M. Mobility and bioavailability of heavy metals and metalloids in soil environments. J. Soil Sci. Plant Nutr. 2010, 10, 268–292. [Google Scholar] [CrossRef] [Green Version]
- Benizri, E.; Kidd, P.S. The Role of the Rhizosphere and Microbes Associated with Hyperaccumulator Plants in Metal Accumulation. In Agromining: Farming for Metals; Van der Ent, A., Echevarria, G., Baker, A.J.M., Morel, J.L., Eds.; Mineral Resource Reviews; Springer International Publishing: Cham, Switzerland, 2018; pp. 157–188. ISBN 978-3-319-61898-2. [Google Scholar]
- Ku, Y.; Rehman, H.M.; Lam, H.M. Possible Roles of Rhizospheric and Endophytic Microbes to Provide a Safe and Affordable Means of Crop Biofortification. Agronomy 2019, 9, 764. [Google Scholar] [CrossRef] [Green Version]
- Kidd, P.; Barceló, J.; Bernal, M.P.; Navari-Izzo, F.; Poschenrieder, C.; Shilev, S.; Clemente, R.; Monterroso, C. Trace Element Behaviour at the Root–Soil Interface: Implications in Phytoremediation. Environ. Exp. Bot. 2009, 67, 243–259. [Google Scholar] [CrossRef]
- Rodríguez, H.; Fraga, R. Phosphate Solubilizing Bacteria and Their Role in Plant Growth Promotion. Biotechnol. Adv. 1999, 17, 319–339. [Google Scholar] [CrossRef]
- Parray, J.A.; Jan, S.; Kamili, A.N.; Qadri, R.A.; Egamberdieva, D.; Ahmad, P. Current Perspectives on Plant Growth-Promoting Rhizobacteria. J. Plant Growth Regul. 2016, 35, 877–902. [Google Scholar] [CrossRef]
- Blake, R.C.; Choate, D.M.; Bardhan, S.; Revis, N.; Barton, L.L.; Zocco, T.G. Chemical Transformation of Toxic Metals by a Pseudomonas Strain from a Toxic Waste Site. Environ. Toxicol. Chem. 1993, 12, 1365–1376. [Google Scholar] [CrossRef]
- Park, C.H.; Keyhan, M.; Wielinga, B.; Fendorf, S.; Matin, A. Purification to Homogeneity and Characterization of a Novel Pseudomonas Putida Chromate Reductase. Appl. Environ. Microbiol. 2000, 66, 1788–1795. [Google Scholar] [CrossRef] [Green Version]
- Rajkumar, M.; Freitas, H. Influence of Metal Resistant-Plant Growth-Promoting Bacteria on the Growth of Ricinus Communis in Soil Contaminated with Heavy Metals. Chemosphere 2008, 71, 834–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.L.; Bilal, S.; Halo, B.A.; Al-Harrasi, A.; Khan, A.R.; Waqas, M.; Al-Thani, G.S.; Al-Amri, I.; Al-Rawahi, A.; Lee, I.-J. Bacillus amyloliquefaciens BSL16 Improves Phytoremediation Potential of Solanum Lycopersicum during Copper Stress. J. Plant Interact. 2017, 12, 550–559. [Google Scholar] [CrossRef] [Green Version]
- Aboushanab, R.; Angle, J.; Chaney, R. Bacterial Inoculants Affecting Nickel Uptake by Alyssum Murale from Low, Moderate and High Ni Soils. Soil Biol. Biochem. 2006, 38, 2882–2889. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Hashem, A.; Abd_Allah, E.F. Bacillus: A Biological Tool for Crop Improvement through Bio-Molecular Changes in Adverse Environments. Front. Physiol. 2017, 8, 667. [Google Scholar] [CrossRef]
- Kabeer, R.; Sylas, V.P.; Praveen Kumar, C.S.; Thomas, A.P.; Shanthiprabha, V.; Radhakrishnan, E.K.; Baiju, K.R. Role of Heavy Metal Tolerant Rhizosphere Bacteria in the Phytoremediation of Cu and Pb Using Eichhornia Crassipes (Mart.) Solms. Int. J. Phytoremediation 2021, 1–13. [Google Scholar] [CrossRef]
- Björklöf, K.; Sen, R.; Jørgensen, K. Maintenance and Impacts of an Inoculated Mer/Luc-Tagged Pseudomonas Fluorescens on Microbial Communities in Birch Rhizospheres Developed on Humus and Peat. Microb. Ecol. 2003, 45, 39–52. [Google Scholar] [CrossRef]
- Castro-Sowinski, S.; Herschkovitz, Y.; Okon, Y.; Jurkevitch, E. Effects of Inoculation with Plant Growth-Promoting Rhizobacteria on Resident Rhizosphere Microorganisms. FEMS Microbiol. Lett. 2007, 276, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ambrosini, A.; de Souza, R.; Passaglia, L.M.P. Ecological Role of Bacterial Inoculants and Their Potential Impact on Soil Microbial Diversity. Plant Soil 2016, 400, 193–207. [Google Scholar] [CrossRef]
- Litchman, E. Invisible Invaders: Non-Pathogenic Invasive Microbes in Aquatic and Terrestrial Ecosystems: Invasive Microbes. Ecol. Lett. 2010, 13, 1560–1572. [Google Scholar] [CrossRef]
- Chowdhury, S.P.; Dietel, K.; Rändler, M.; Schmid, M.; Junge, H.; Borriss, R.; Hartmann, A.; Grosch, R. Effects of Bacillus amyloliquefaciens FZB42 on Lettuce Growth and Health under Pathogen Pressure and Its Impact on the Rhizosphere Bacterial Community. PLoS ONE 2013, 8, e68818. [Google Scholar] [CrossRef] [Green Version]
- Lladó, S.; López-Mondéjar, R.; Baldrian, P. Drivers of Microbial Community Structure in Forest Soils. Appl. Microbiol. Biotechnol. 2018, 102, 4331–4338. [Google Scholar] [CrossRef]
- Li, L.; Ma, J.; Mark Ibekwe, A.; Wang, Q.; Yang, C.-H. Influence of Bacillus Subtilis B068150 on Cucumber Rhizosphere Microbial Composition as a Plant Protective Agent. Plant Soil 2018, 429, 519–531. [Google Scholar] [CrossRef]
- Saad, M.M.; Eida, A.A.; Hirt, H. Tailoring Plant-Associated Microbial Inoculants in Agriculture: A Roadmap for Successful Application. J. Exp. Bot. 2020, 71, 3878–3901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redondo-Gómez, S.; Mesa-Marín, J.; Pérez-Romero, J.A.; López-Jurado, J.; García-López, J.V.; Mariscal, V.; Molina-Heredia, F.P.; Pajuelo, E.; Rodríguez-Llorente, I.D.; Flowers, T.J.; et al. Consortia of Plant-Growth-Promoting Rhizobacteria Isolated from Halophytes Improve Response of Eight Crops to Soil Salinization and Climate Change Conditions. Agronomy 2021, 11, 1609. [Google Scholar] [CrossRef]
- Samaniego-Gámez, B.Y.; Garruña, R.; Tun-Suárez, J.M.; Kantun-Can, J.; Reyes-Ramírez, A.; Cervantes-Díaz, L. Bacillus Spp. Inoculation Improves Photosystem II Efficiency and Enhances Photosynthesis in Pepper Plants. Chil. J. Agric. Res. 2016, 76, 409–416. [Google Scholar] [CrossRef] [Green Version]
- Osorio, S.; Ruan, Y.-L.; Fernie, A.R. An Update on Source-to-Sink Carbon Partitioning in Tomato. Front. Plant Sci. 2014, 5, 516. [Google Scholar] [CrossRef] [Green Version]
- Enebe, M.C.; Babalola, O.O. The Influence of Plant Growth-Promoting Rhizobacteria in Plant Tolerance to Abiotic Stress: A Survival Strategy. Appl. Microbiol. Biotechnol. 2018, 102, 7821–7835. [Google Scholar] [CrossRef] [Green Version]
- Stefan, M.; Munteanu, N.; Stoleru, V.; Mihasan, M.; Hritcu, L. Seed Inoculation with Plant Growth Promoting Rhizobacteria Enhances Photosynthesis and Yield of Runner Bean (Phaseolus coccineus L.). Sci. Hortic. 2013, 151, 22–29. [Google Scholar] [CrossRef]
- Naveed, M.; Mitter, B.; Reichenauer, T.G.; Wieczorek, K.; Sessitsch, A. Increased Drought Stress Resilience of Maize through Endophytic Colonization by Burkholderia Phytofirmans PsJN and Enterobacter Sp. FD17. Environ. Exp. Bot. 2014, 97, 30–39. [Google Scholar] [CrossRef]
- Jamil, M.; Zeb, S.; Anees, M.; Roohi, A.; Ahmed, I.; ur Rehman, S.; Rha, E. shik Role of Bacillus Licheniformis in Phytoremediation of Nickel Contaminated Soil Cultivated with Rice. Int. J. Phytoremediation 2014, 16, 554–571. [Google Scholar] [CrossRef]
- Brantley, S.L.; Liermann, L.; Bau, M. Uptake of Trace Metals and Rare Earth Elements from Hornblende by a Soil Bacterium. Geomicrobiol. J. 2001, 18, 37–61. [Google Scholar] [CrossRef]
- Zhuang, X.; Chen, J.; Shim, H.; Bai, Z. New Advances in Plant Growth-Promoting Rhizobacteria for Bioremediation. Environ. Int. 2007, 33, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Ahemad, M. Enhancing Phytoremediation of Chromium-Stressed Soils through Plant-Growth-Promoting Bacteria. J. Genet. Eng. Biotechnol. 2015, 13, 51–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, X.-F.; Xia, J.-J.; Jiang, C.-Y.; He, L.-Y.; Qian, M. Characterization of Heavy Metal-Resistant Endophytic Bacteria from Rape (Brassica Napus) Roots and Their Potential in Promoting the Growth and Lead Accumulation of Rape. Environ. Pollut. Barking Essex 1987 2008, 156, 1164–1170. [Google Scholar] [CrossRef] [PubMed]
- Lampis, S.; Santi, C.; Ciurli, A.; Andreolli, M.; Vallini, G. Promotion of Arsenic Phytoextraction Efficiency in the Fern Pteris Vittata by the Inoculation of As-Resistant Bacteria: A Soil Bioremediation Perspective. Front. Plant Sci. 2015, 6, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qurashi, A.W.; Sabri, A.N. Bacterial Exopolysaccharide and Biofilm Formation Stimulate Chickpea Growth and Soil Aggregation under Salt Stress. Braz. J. Microbiol. Publ. Braz. Soc. Microbiol. 2012, 43, 1183–1191. [Google Scholar] [CrossRef] [Green Version]
- Fashola, M.; Ngole-Jeme, V.; Babalola, O. Diversity of Acidophilic Bacteria and Archaea and Their Roles in Bioremediation of Acid Mine Drainage. Br. Microbiol. Res. J. 2015, 8, 443–456. [Google Scholar] [CrossRef]
- Seshadri, B.; Bolan, N.S.; Naidu, R. Rhizosphere-Induced Heavy Metal(Loid) Transformation in Relation to Bioavailability and Remediation. J. Soil Sci. Plant Nutr. 2015, 15, 524–548. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.A.; Hussain, I.; Rasheed, R.; Iqbal, M.; Riaz, M.; Arif, M.S. Advances in Microbe-Assisted Reclamation of Heavy Metal Contaminated Soils over the Last Decade: A Review. J. Environ. Manage. 2017, 198, 132–143. [Google Scholar] [CrossRef]
- Fan, B.; Carvalhais, L.C.; Becker, A.; Fedoseyenko, D.; von Wirén, N.; Borriss, R. Transcriptomic Profiling of Bacillus amyloliquefaciens FZB42 in Response to Maize Root Exudates. BMC Microbiol. 2012, 12, 116. [Google Scholar] [CrossRef] [Green Version]
- Silva, R.S.; Antunes, J.E.L.; de Aquino, J.P.A.; de Sousa, R.S.; de Melo, W.J.; Araujo, A.S.F. Plant Growth-Promoting Rhizobacteria Effect on Maize Growth and Microbial Biomass in a Chromium-Contaminated Soil. Bragantia 2021, 80, e2521. [Google Scholar] [CrossRef]
- Mallick, I.; Bhattacharyya, C.; Mukherji, S.; Dey, D.; Sarkar, S.C.; Mukhopadhyay, U.K.; Ghosh, A. Effective Rhizoinoculation and Biofilm Formation by Arsenic Immobilizing Halophilic Plant Growth Promoting Bacteria (PGPB) Isolated from Mangrove Rhizosphere: A Step towards Arsenic Rhizoremediation. Sci. Total Environ. 2018, 610–611, 1239–1250. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, P.; Rathinasabapathi, B.; Ma, L.Q. Arsenic-Resistant Bacteria Solubilized Arsenic in the Growth Media and Increased Growth of Arsenic Hyperaccumulator Pteris vittata L. Bioresour. Technol. 2011, 102, 8756–8761. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, G.; Qi, C.; Cheng, S.; Sun, R. Chemical Speciation and Combustion Behavior of Chromium (Cr) and Vanadium (V) in Coals. Fuel 2016, 184, 42–49. [Google Scholar] [CrossRef]
- Baleizão, C.; Gigante, B.; Sabater, M.J.; Garcia, H.; Corma, A. On the Activity of Chiral Chromium Salen Complexes Covalently Bound to Solid Silicates for the Enantioselective Epoxide Ring Opening. Appl. Catal. Gen. 2002, 228, 279–288. [Google Scholar] [CrossRef]
- Park, J.H.; Bolan, N. Lead Immobilization and Bioavailability in Microbial and Root Interface. J. Hazard. Mater. 2013, 261, 777–783. [Google Scholar] [CrossRef]
- Bridges, C.C.; Zalups, R.K. Molecular and Ionic Mimicry and the Transport of Toxic Metals. Toxicol. Appl. Pharmacol. 2005, 204, 274–308. [Google Scholar] [CrossRef] [Green Version]
- Pagano, G.; Guida, M.; Tommasi, F.; Oral, R. Health Effects and Toxicity Mechanisms of Rare Earth Elements—Knowledge Gaps and Research Prospects. Ecotoxicol. Environ. Saf. 2015, 115, 40–48. [Google Scholar] [CrossRef]
- Wiche, O.; Zertani, V.; Hentschel, W.; Achtziger, R.; Midula, P. Germanium and Rare Earth Elements in Topsoil and Soil-Grown Plants on Different Land Use Types in the Mining Area of Freiberg (Germany). J. Geochem. Explor. 2017, 175, 120–129. [Google Scholar] [CrossRef]
- Du Laing, G.; Rinklebe, J.; Vandecasteele, B.; Meers, E.; Tack, F.M.G. Trace Metal Behaviour in Estuarine and Riverine Floodplain Soils and Sediments: A Review. Sci. Total Environ. 2009, 407, 3972–3985. [Google Scholar] [CrossRef]
- Alfassi, Z.B. , Wai, C.M., Eds. Preconcentration Techniques for Trace Elements; CRC Press: Boca Raton, FL, USA, 1992; ISBN 978-0-8493-5213-3. [Google Scholar]
- China National Analysis Center for Iron and Steel Certificate of Certified Reference Materials. LGC Standards 2021. Available online: https://bit.ly/3A7OQiC (accessed on 12 January 2022).
- Cao, X.; Chen, Y.; Wang, X.; Deng, X. Effects of Redox Potential and PH Value on the Release of Rare Earth Elements from Soil. Chemosphere 2001, 44, 655–661. [Google Scholar] [CrossRef]
- Olaniran, A.; Balgobind, A.; Pillay, B. Bioavailability of Heavy Metals in Soil: Impact on Microbial Biodegradation of Organic Compounds and Possible Improvement Strategies. Int. J. Mol. Sci. 2013, 14, 10197–10228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tóth, G.; Hermann, T.; Da Silva, M.R.; Montanarella, L. Heavy Metals in Agricultural Soils of the European Union with Implications for Food Safety. Environ. Int. 2016, 88, 299–309. [Google Scholar] [CrossRef]
- Krachler, M.; Mohl, C.; Emons, H.; Shotyk, W. Influence of Digestion Procedures on the Determination of Rare Earth Elements in Peat and Plant Samples by USN-ICP-MS. J. Anal. At. Spectrom. 2002, 17, 844–851. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global Patterns of 16S RRNA Diversity at a Depth of Millions of Sequences per Sample. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4516–4522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weißbecker, C.; Schnabel, B.; Heintz-Buschart, A. Dadasnake, a Snakemake Implementation of DADA2 to Process Amplicon Sequencing Data for Microbial Ecology. GigaScience 2020, 9, giaa135. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Koster, J.; Rahmann, S. Snakemake--a Scalable Bioinformatics Workflow Engine. Bioinformatics 2012, 28, 2520–2522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing Mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum | BW NIL | BW R | ZM NIL | ZM R |
|---|---|---|---|---|
| Acidobacteriota | 10.31 | 9.81 | 9.83 | 10.53 |
| Actinobacteriota | 28.98 | 27.88 | 27.62 | 27.39 |
| Bacteroidota | 2.83 | 3.08 | 2.57 | 2.21 |
| Chloroflexi | 7.97 | 7.56 | 7.65 | 8.00 |
| Crenarchaeota | 0.59 | 0.58 | 0.61 | 0.63 |
| Firmicutes | 6.67 | 7.55 | 7.69 | 7.09 |
| Gemmatimonadota | 4.03 | 4.31 | 4.35 | 4.57 |
| Methylomirabilota | 0.74 | 0.50 | 0.66 | 0.74 |
| Myxococcota | 3.11 | 3.33 | 3.72 | 3.94 |
| Patescibacteria | 1.39 | 1.66 | 1.61 | 1.67 |
| Planctomycetota | 7.26 | 7.58 | 7.14 | 6.95 |
| Proteobacteria | 18.40 | 18.37 | 18.73 | 18.05 |
| Verrucomicrobiota | 2.74 | 2.65 | 2.64 | 2.89 |
| Unidentified | 0.72 | 0.81 | 0.64 | 0.79 |
| Species | Treatment | Cr | As | Pb | Co | Cu | Ge | REET |
|---|---|---|---|---|---|---|---|---|
| Z. mays | NIL | 3.86 ± 0.90 | 2.50 ± 0.31 | 1.93 ± 0.89 | 4.14 ± 0.51 | 30.1 ± 5.74 | 0.26 ± 0.16 | 0.68 ± 0.54 |
| R | 2.58 ± 0.11 | 2.34 ± 0.38 | 1.28 ± 0.05 | 1.48 ± 0.18 | 31 ± 1.52 | 0.09 ± 0.004 | 0.17 ± 0.08 | |
| Statistic a | 1.97 | 0.10 | 0.52 | 24.0 | 0.019 | 1.05 | 0.87 | |
| p value | 0.29 | 0.77 | 0.55 | 0.03 | 0.9 | 0.41 | 0.45 | |
| F. esculentum | NIL | 5.15 ± 2.22 | 3.72 ± 0.18 | 1.72 ± 0.16 | 0.15 ± 0.01 | 7.94 ± 2.49 | 0.01 ± 0.004 | 0.77 ± 0.06 |
| R | 1.89 ± 0.34 | 4.14 ± 0.58 | 3.49 ± 0.58 | 2.97 ± 0.30 | 36.1± 8.90 | 0.005 ± 0.001 | 0.96 ± 0.17 | |
| Statistic a | 2.11 | 0.47 | 8.68 | 90.98 | 9.25 | 0.81 | 1.21 | |
| p value | 0.28 | 0.55 | 0.08 | 0.011 | 0.078 | 0.46 | 0.37 |
| Species | Treatment | P | Ca | Si | Fe |
|---|---|---|---|---|---|
| Z. mays | NIL | 1681 ± 181 | 6981 ± 611 | 3137 ± 636 | 88 ± 8 |
| R | 1578 ± 208 | 5975 ± 1162 | 2744 ± 142 | 76 ± 6 | |
| Statistic a | 0.14 | 0.59 | 0.36 | 1.28 | |
| p value | 0.728 | 0.499 | 0.603 | 0.327 | |
| F. esculentum | NIL | 1699 ± 122 | 13,434 ± 692 | 549 ± 34 | 67 ± 4 |
| R | 1953 ± 94 | 17,421 ± 1294 | 611 ± 53 | 53 ± 4 | |
| Statistic a | 2.73 | 7.39 | 0.95 | 6.06 | |
| p value | 0.18 | 0.07 | 0.39 | 0.07 |
| 4a: Soil Physico-Chemical Parameters | |||
| Water content (w/w) | 17.9% | ||
| pH value in aqueous solution | 6.2 | ||
| Conductivity | 32.3 µS/cm | ||
| Organic matter content | 7.7% | ||
| Nitrate concentration | 147 mg/kg | ||
| Ammonium concentration | 0.88 mg/kg | ||
| Phosphate concentration | 136 mg/kg | ||
| Cation exchange capacity | 9.1 cmol/kg | ||
| 4b: Total Concentration and Concentration in Operationally Defined Fractions (µg/g) (mean ± SE) | |||
| Total concentration | Fraction 1 | Fraction 2 | |
| Cu | 175 ± 36 | 0.69 ± 0.04 | 0.84 ± 0.1 |
| Pb | 180 ± 41 | 5.6 ± 0.8 | 31 ± 3.2 |
| Cr | 111 ± 11 | 0.10 ± 0.02 | 0.23 ± 0.01 |
| As | 93 ± 25 | 0.39 ± 0.2 | 0.73 ± 0.2 |
| Ge | 1.84 ± 0.04 | 0.004 ± 0.001 | 0.014 ± 0.001 |
| REET | 157 ± 3.1 | 0.99 ± 0.1 | 2.80 ± 0.2 |
| Ca | 5875 ± 675 | 2282 ± 495 | 232 ± 45 |
| P | 1986 ± 89 | 33.3 ± 6.3 | 25.6 ± 8.3 |
| Fe | 29,337 ± 551 | 4.1 ± 0.4 | 19.4 ± 2.2 |
| Co | 24.3 ± 2.1 | 0.09 ± 0.01 | 0.24 ± 0.02 |
| Si | 141,455 ± 18,019 | 62.7 ± 9.6 | 54.7 ± 5.0 |
| Step | Temperature (°C) | Time (min:sec) | |
|---|---|---|---|
| PCR 1 | |||
| Initial denaturation | 95 | 3:00 | |
| 25 cycles | Denaturation | 98 | 0:20 |
| Annealing | 55 | 0:15 | |
| Elongation | 72 | 0:15 | |
| Final extension | 72 | 5:00 | |
| PCR 2 | |||
| Initial denaturation | 95 | 3:00 | |
| 8 cycles | Denaturation | 98 | 0:30 |
| Annealing | 55 | 0:30 | |
| Elongation | 72 | 0:30 | |
| Final extension | 72 | 5:00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okoroafor, P.U.; Mann, L.; Amin Ngu, K.; Zaffar, N.; Monei, N.L.; Boldt, C.; Reitz, T.; Heilmeier, H.; Wiche, O. Impact of Soil Inoculation with Bacillus amyloliquefaciens FZB42 on the Phytoaccumulation of Germanium, Rare Earth Elements, and Potentially Toxic Elements. Plants 2022, 11, 341. https://doi.org/10.3390/plants11030341
Okoroafor PU, Mann L, Amin Ngu K, Zaffar N, Monei NL, Boldt C, Reitz T, Heilmeier H, Wiche O. Impact of Soil Inoculation with Bacillus amyloliquefaciens FZB42 on the Phytoaccumulation of Germanium, Rare Earth Elements, and Potentially Toxic Elements. Plants. 2022; 11(3):341. https://doi.org/10.3390/plants11030341
Chicago/Turabian StyleOkoroafor, Precious Uchenna, Lotte Mann, Kerian Amin Ngu, Nazia Zaffar, Nthati Lillian Monei, Christin Boldt, Thomas Reitz, Hermann Heilmeier, and Oliver Wiche. 2022. "Impact of Soil Inoculation with Bacillus amyloliquefaciens FZB42 on the Phytoaccumulation of Germanium, Rare Earth Elements, and Potentially Toxic Elements" Plants 11, no. 3: 341. https://doi.org/10.3390/plants11030341