
Ooctonus vulgatus (Hymenoptera, Mymaridae), a potential biocontrol agent to reduce populations of Philaenus spumarius (Hemiptera, Aphrophoridae) the main vector of Xylella fastidiosa in Europe
- Published
- Accepted
- Received
- Academic Editor
- Ilaria Negri
- Subject Areas
- Agricultural Science, Biodiversity, Entomology, Spatial and Geographic Information Science
- Keywords
- Insect vector, Oophagous, Meadow spittlebug, Parasitoid, Biological control, Natural regulation
- Copyright
- © 2020 Mesmin et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2020. Ooctonus vulgatus (Hymenoptera, Mymaridae), a potential biocontrol agent to reduce populations of Philaenus spumarius (Hemiptera, Aphrophoridae) the main vector of Xylella fastidiosa in Europe. PeerJ 8:e8591 https://doi.org/10.7717/peerj.8591
Abstract
As a vector of Xylella fastidiosa (Wells, 1987) in Europe, the meadow spittlebug Philaenus spumarius (Linnaeus, 1758) (Hemiptera, Aphrophoridae) is a species of major concern. Therefore, tools and agents to control this ubiquitous insect that develops and feeds on hundreds of plant species are wanted. We conducted a field survey of P. spumarius eggs in Corsica and provide a first report of Ooctonus vulgatus Haliday, 1833 (Hymenoptera, Mymaridae) as a potential biocontrol agent of P. spumarius in Europe. To allow species identification, we summarized the main characters distinguishing O. vulgatus from other European species of Ooctonus and generated COI DNA barcodes. Parasitism rates were variable in the four localities included in the survey but could reach 69% (for an average number of eggs that hatched per locality of 109). Based on the geographic occurrences of O. vulgatus obtained from the literature, we calibrated an ecological niche model to assess its potential distribution in the Holarctic. Obviously, several questions need to be addressed to determine whether O. vulgatus could become an effective biocontrol agent of P. spumarius in Europe. So far, O. vulgatus has been reared only from P. spumarius eggs, but its exact host-range should be evaluated to ensure efficiency and avoid non-target effect. The top-down impact of the parasitoid on vector populations should also be assessed on large data sets. Finally, the feasibility of mass rearing should be tested. We hope this report serves as a starting point to initiate research on this parasitoid wasp to assess whether it could contribute to reduce the spread and impact of X. fastidiosa in Europe.
Introduction
Xylella fastidiosa (Wells, 1987) is a xylem-dwelling insect-borne bacterium that originates from the Americas, infects more than 500 species of plants (EFSA, 2015) and causes a variety of scorch-like diseases in many cultivated species (Almeida & Nunney, 2015; EFSA, 2018; Sicard et al., 2018). Studies on the economic impact of X. fastidiosa have primarily focused on the wine and grape industries. Yield reduction and management costs to the California grape industry are estimated at more than US$100 million per year (Tumber, Alston & Fuller, 2014) and a potential introduction of the bacterium in Australia is estimated to cost up to AUD 7.9 billion over 50 years (Hafi et al., 2017).
X. fastidiosa has been recently detected in Europe and is present in Italy (Saponari et al., 2013), France (Denancé et al., 2017), Spain (Olmo et al., 2017), and Portugal (DGAV, 2019). Furthermore, niche modelling has shown that a large part of Europe is climatically suitable for the bacterium (Godefroid et al., 2018; Godefroid et al., 2019). Hence, X. fastidiosa represents a serious threat to European agriculture and natural ecosystems.
The spread of X. fastidiosa depends on several interacting factors, mainly insect vectors and plant communities as well as landscape, climate features and population dynamics of the bacterium itself (Krugner et al., 2019). As a consequence, disease management is complex. Reducing bacterium spread requires acting on a set of different biotic and abiotic factors (Almeida et al., 2005) and modelling approaches may help setting up effective strategies (Fierro, Liccardo & Porcelli, 2019). Here we focus on a possible management strategy to control populations of the most common vector of X. fastidiosa reported in Europe so far: the meadow spittlebug Philaenus spumarius (Linnaeus, 1758) (Hemiptera, Aphrophoridae) (Saponari et al., 2014; Cornara et al., 2016).
P. spumarius is highly polyphagous (Cornara, Bosco & Fereres, 2018), widely distributed in the Palearctic from sea level to high elevation (about 2,000 m; e.g., Halkka, Raatikainen & Vilbaste, 1975; Lees, Dent & Gait, 1983; Drosopoulos & Asche, 1991; Loukas & Drosopoulos, 1992; Quartau, Borges & André, 1992; Stewart & Lees, 1996; Drosopoulos & Remane, 2000), and was probably introduced to the New World (Whittaker, 1973). Its ability to acquire and transmit X. fastidiosa was previously demonstrated (Severin, 1950; Saponari et al., 2014; Cornara et al., 2016).
So far, a few studies have assessed the impact of different insecticides to reduce juvenile populations of P. spumarius in Europe (Dongiovanni et al., 2018; Dader et al., 2019). However, there is a growing awareness of the need to encourage management practices that safeguard harvests, human health, biodiversity and the environment. Thus, the development of effective biological control programs is desirable. Among biocontrol strategies, augmentative biological control consists in enhancing the effectiveness of naturally occurring natural enemies by the periodic release of specimens (Eilenberg, Hajek & Lomer, 2001; Aubertot & Savary, 2005). Compared to classical biological control it eliminates unintended effects of the introduction of new, non-native, parasitoids or predaceous arthropods (Hoy, 2008). However, as for all biological control programs, augmentative biocontrol requires field investigations to identify potential natural enemies of the target pest.
Currently, information about the natural enemies of the meadow spittlebug are scattered (Cornara, Bosco & Fereres, 2018). Species of birds, frogs, arachnids, and insects (Hymenoptera, Diptera, and Coleoptera) occasionally feed on P. spumarius (Phillipson, 1960; Halkka & Kohila, 1976; Harper & Whittaker, 1976; Henderson, Hoffman & Jeanne, 1990; Pagliano & Alma, 1997) but predation by native natural enemies does not appear to be an important source of mortality. Studies are in progress to test whether the invasive assassin bug Zelus renardii Kolenati, 1857 (Hemiptera, Reduviidae) could be used to control populations of P. spumarius in olive orchards (Salerno et al., 2017). However, mass release of this species may be risky for local biodiversity, especially for beneficial arthropods (Ables, 1978). Indeed, it is considered as a generalist predator (Ables, 1978; Cisneros & Rosenheim, 1998; Weirauch, Alvarez & Zhang, 2012; Salerno et al., 2017, but see Cohen & Tang, 1997 who suggest a strong effect of prey body size).
So far, only few parasitoids of P. spumarius have been recorded. Adults are attacked by Verralia aucta (Fallen, 1817) (Diptera, Pipunculidae) in Europe with relatively high parasitism rates in England: in average 31% in females and 46% in males over four years (Whittaker, 1969; Whittaker, 1973). Parasitism by V. aucta has a direct effect on P. spumarius population dynamics because it renders the host sterile (Whittaker, 1973). However, this parasitoid does not have an immediate effect on bacterium transmission because adults are only killed after 10–11 weeks of parasitism (Whittaker, 1969), a period during which they are probably still able to spread the bacterium. Contrastingly, an interesting feature of egg parasitoids is that they kill the host in the egg stage, that is, before it can inflict damage to its host plants (Mills, 2010). In the case of P. spumarius, the insect is killed before it acquires the bacterium from an infected host plant and becomes able to transmit it. A few egg parasitoids have been recorded in the US: Ooctonus vulgatus Haliday, 1833 (Hymenoptera, Mymaridae) and at least two unnamed species of Centrodora (Hymenoptera, Aphelinidae) (Weaver & King, 1954). Indeed, the genus Tumidiscapus, which is cited as parasitoid of P. spumarius in the US (Weaver & King, 1954), is in fact a synonym of Centrodora (Hayat, 1983). However, little is known about the biology and efficacy of egg parasitoids in natura.
In this study, a field survey was conducted to identify major egg parasitoids of P. spumarius in Corsica. We provide a first report of Ooctonus vulgatus in this area. We summarized the main characters separating O. vulgatus from other Palearctic species to facilitate identification and generate COI DNA barcodes to accurately identify the species. Finally, we reviewed the literature and gathered all available occurrence data (i.e., geographical coordinates) of previously detected populations of O. vulgatus. This allowed us to calibrate an ecological niche model linking different climate descriptors to species occurrence data and estimate the potential distribution of the parasitoid in the Holarctic region for comparison with the distribution of P. spumarius.

Figure 1: Parasitism rate of Philaenus spumarius eggs in the four sites sampled in Corsica.
Size of the pie chart is proportional to the total number of eggs that hatched from each locality (n). Slices indicate the relative proportion of O. vulgatus (dark grey) and nymphs of P. spumarius (light grey) that emerged from the pool of eggs. GPS coordinates of sampling localities: 42.984205°N, 9.395287°E (Ersa); 42.338849°N, 9.180636°E (Tralonca); 42.274756°N, 9.487185°E (Canale-di-Verde); 41.931726°N, 9.343731°E (Ventiseri). The map was built with the R package maps, using data from UNESCO (1987) through UNEP/GRID-Geneva.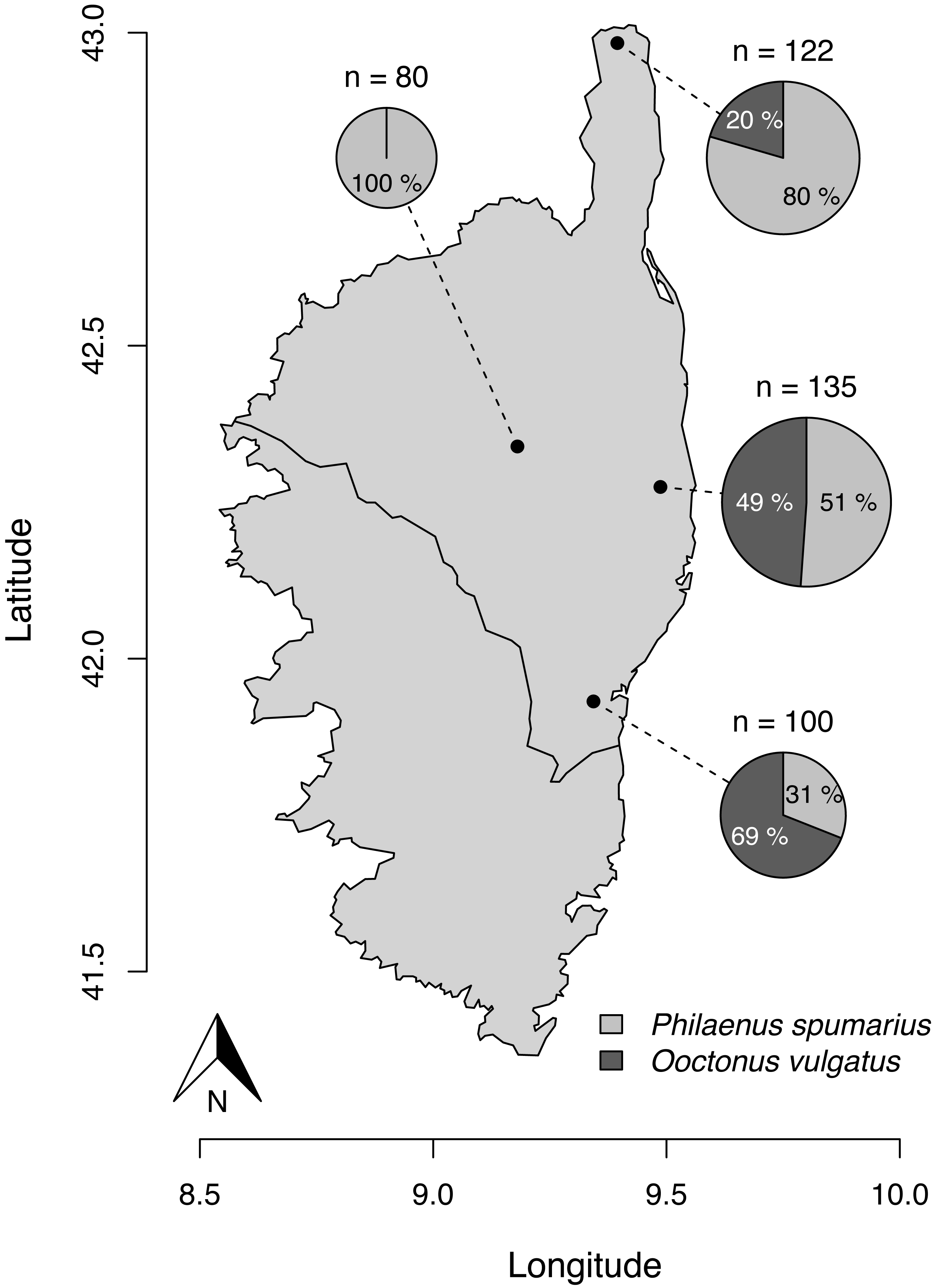{kind=link}
Materials and Methods
Sampling and calculation of parasitism rate
Five to ten handfuls of about eight top branches of Cistus monspeliensis L. 1753 (cut at 50 cm below the end of the branch) were sampled in four localities (Fig. 1). These localities were part of a larger field survey of population dynamics of P. spumarius in Corsica. We targeted C. monspeliensis to maximize our chances to find eggs of P. spumarius. Indeed, we demonstrated in a previous study that, in Corsica, adults of P. spumarius seemed to be mainly associated with this species (Cruaud et al., 2018). Sampling was performed between the 12th and the 15th of February 2019. The back of each leaf (about 900 leaves per handful of branches) was inspected in the laboratory for whitish clusters, which were retained and inspected under a binocular microscope to confirm the presence of eggs of P. spumarius (Appendix S1; 109, 148, 167, 187 eggs obtained per site, hence a total of 611 eggs monitored). The morphological identification of P. spumarius eggs and first stage nymphs was performed using the descriptions of Weaver & King (1954) (Appendix S1). The pieces of leaf containing the eggs were placed on filter papers in unaerated Petri dishes (i.e., without spur) at room temperature (20.2 ± 1.5 °C), with natural light. Filter papers were kept moist by adding drops of water when necessary. Hatching was monitored every morning from the 18th of February to the 15th of March 2019. Emerging nymphs and parasitoids were killed and stored in 70% Ethanol at 4 °C. Parasitism rates were computed in each locality with the following formula: (Costello & Altieri, 1995).
Morphological identification of the parasitoids
Identification to species was performed using the Ooctonus keys by Triapitsyn (2010) and Huber (2012). Specimens were desiccated using HMDS (Heraty & Hawks, 1998) and glued on grey cards. Imaging was performed with a Keyence digital microscope (VHX-5000 Camera color CMOS and the VH-Z100UT lens). Images were then edited in Adobe Photoshop CS6©software.
Molecular identification of the parasitoids
Six individuals were used for molecular identification. Three of them were handled individually (sample codes = XMES00042_0101, XMES00077_0101, XMES00091_0101) and the remaining three were pooled to increase DNA yield (sample code = XMES00041_0189). Total genomic DNA was isolated using the Qiagen DNeasy Blood & Tissue kit without destruction of the specimens. We followed manufacturer’s protocol with the following modifications. Samples (whole insects, without dissection or crushing) were incubated overnight in an Eppendorf thermomixer (temperature = 56 °C, mixing frequency = 300 rpm). To increase DNA yield, two successive elutions (50 µL each) were performed with heated buffer AE (56 °C) and an incubation step of 15 min followed by centrifugation (6,000 g for 1 min at room temperature; see Cruaud et al. (2019) for a detailed description of the protocol). Eppendorf microtubes LoBind 1.5 ml were used for elution and to store DNA at minus 20 °C until PCR amplification. Vouchers were deposited at Centre de Biologie pour la Gestion des Populations (CBGP), Montferrier-sur-Lez, France. The mitochondrial Cytochrome c oxidase I standard barcode fragment (COI) was amplified with a cocktail of M13-tailed primers as detailed in Germain et al. (2013). Unpurified PCR products were sent to Eurofins MWG Operon (Ebersberg, Germany) for sequencing using the M13F and M13R primers (Germain et al., 2013; Ivanova et al., 2007). Both strands for each overlapping fragment were assembled in Geneious v11.1.4 (https://www.geneious.com). Geneious was also used to translate consensus sequences to amino acids to detect premature codon stops. All COI sequences available on BOLD (Ratnasingham & Hebert, 2007) for Ooctonus species were downloaded (last access July 12, 2019) and aligned with the newly generated sequences using MAFFT v7.245 (Katoh & Standley, 2013). A maximum likelihood tree was inferred with raxmlHPC-PTHREADS-AVX version 8.2.4 (Stamatakis, 2014). A rapid bootstrap search (100 replicates) followed by a thorough ML search (-m GTRGAMMA) was conducted. Tree visualization and annotation was performed with TreeGraph 2.13 (Stöver & Müller, 2010).
Species distribution modelling framework
Occurrences of O. vulgatus were retrieved from the literature and the GBIF database (GBIF.org, 2019) (Tables S1 and S2). Two hundred and five occurrences were obtained from the literature (Table S2), eight of which were not included in the analysis as no geographic coordinates were available. Forty occurrences were obtained from GBIF (last access: 22 August 2019; Table S2), but were all discarded due to dubious identification or lack of information on sample origin. Therefore, no occurrence obtained from GBIF could be included in the analysis.
We fitted a correlative model linking different climate descriptors to species occurrences. The Maxent algorithm was chosen to conduct analyses because it does not require absence data (i.e., locations in which we can presume that a species is truly absent) (Phillips, Anderson & Schapire, 2006). We summarized below the main step of our analysis and details are provided in Appendix S2. The mean temperature and precipitation of the wettest, driest, warmest, and coldest quarters as well as precipitation seasonality were extracted from the Worldclim 2.0 database (Fick & Hijmans, 2017) and used as bioclimatic descriptors (Hijmans et al., 2005). In absence of formal knowledge about climatic factors constraining O. vulgatus distribution, we constituted three sets of bioclimatic variables and performed modelling with each of them (Qiao, Soberón & Peterson, 2015; Godefroid et al., 2019). The first set (CLIM1) comprised the mean temperature of the wettest, driest, warmest, and coldest quarters to reflect the impact of temperature constraints on distribution. To highlight the precipitation constraint, we added the precipitation seasonality to CLIM1 and constituted the second set (CLIM2). Finally, we built a third set (CLIM3) by assembling CLIM1 and the precipitation of the wettest, driest, warmest, and coldest quarters to fully account for both extreme temperatures and precipitations in the species distribution models (SDMs). The Maxent algorithm requires a set of locations where the species has been found (here, a random 70% of the available occurrences, the other 30% being used for model validation) and a set of locations where no information about the presence of the species are available (referred to as background points). A total of 10,000 background points were randomly generated in North America and Europe. To render complex response to environmental constraints while reducing model overfitting we first fitted 48 Maxent models using six regularization multiplier (RM) combinations (L, LQ, H, LQH, LQHP, LQHPT with L = linear, Q = quadratic, H = hinge, P = product and T = threshold) and feature class (FC) values (eight values ranging from 0.5 to 4 with increments of 0.5) (Radosavljevic & Anderson, 2014). Optimal FC and RM combinations were determined for each of the three bioclimatic datasets (CLIM1–CLIM3) using the R language (R Core Team, 2019) and the package ENMeval (Muscarella et al., 2014). Optimal parameters were then used to fit a set of 10 replicate Maxent models using 70% of the dataset. The performance of each model was evaluated using the remaining 30% of occurrences using the area under the receiver–operator curve (AUC, Fielding & Bell, 1997) and the true skill statistics (TSS, Allouche, Tsoar & Kadmon, 2006). Models with AUC <0.8 were excluded from further analyses (Vicente et al., 2013). Habitat suitability maps (logistic output ranging from 0 to 1) were transformed into binary projections using the threshold that optimized the TSS statistics on the testing data (Guisan, Thuiller & Zimmermann, 2017). Maxent replicate models were fitted and evaluated using the R package biomod2 (Thuiller et al., 2009).
Two different outputs were generated using the set of model prediction. (i) Binary predictions were averaged to produce the committee (consensus) averaging (Araújo & New, 2007; Marmion et al., 2009) showing the likelihood of the presence of O. vulgatus. This consensus model ranges from 0 (all the models predict absence) to 100% (all the models predict presence) and (ii) the median of the logistic outputs (Guisan, Thuiller & Zimmermann, 2017) of the models that depicts the climate suitability across the different models.
Results
Parasitism rates
Out of the 611 eggs monitored, 437 (i.e., 71.5%) hatched. 277 (63.4%) gave rise to P. spumarius nymphs and parasitoids emerged from 160 eggs (36.6%). All parasitoids were identified as O. vulgatus (Fig. 2). No parasitoid emerged from eggs collected in one of the four localities. We observed parasitism rates of 20.5, 48.9 and 69.0% in the three other localities (Fig. 1).

Figure 2: Morphology of Ooctonus vulgatus Haliday, 1833.
(A) Male antenna (B) Female antenna (C) Habitus. (D) Head front view. (E) Mesosoma lateral view. (F) Male propodeum. (G) Mesosoma dorsal view. All scales = 100 µm except habitus. Photo credit: Jean-Yves Rasplus INRA.{kind=link}
Guidelines for the identification of O. vulgatus
To help identification, we list below the main features that differentiate O. vulgatus from its closest relatives. The genus Ooctonus has been recently revised in the Palearctic and Nearctic regions respectively by Triapitsyn (2010) and Huber (2012). Ooctonus can be distinguished from other genera of Mymaridae by the following set of characters: tarsi 5-segmented, propodeum with diamond-shaped pattern of carinae (Fig. 2F), fore wing venation about one-third the wing length (Fig. 2C), with short marginal and stigmal vein, parastigma with hypochaeta next to proximal macrochaeta (Huber, 2012). In the Holarctic region, O. vulgatus can be distinguished from other species of Ooctonus by the following unique combination of features (Fig. 2): vertex without stemmaticum; mesoscutum without median groove; posterior part of scutellum and frenum smooth with weak sculpture laterally; metanotum and propodeum without reticulate sculpture; propodeum without median carina, but with a pentagonal areole formed by dorsolateral carinae; short petiole, 0.9–1.2x as long as metacoxa; forewing at least slightly truncate apically; females funicle with multiporous placoid sensilla (mps) on F7 and F8 only, F5 and F6 without mps; single row of six bullae inside the female clava; ovipositor at most 1. 4 × as long as metatibia and only slightly exerted beyond apex of gaster.
Molecular identification of the parasitoid
Barcode sequences were successfully generated from all samples. All sequences were identical. Phylogenetic analysis confirmed that the most likely identification was O. vulgatus (Fig. S1).
Species distribution modelling
A total of 200 occurrences (197 obtained from the literature plus the three localities where we sampled O. vulgatus) (Fig. 3A) were used to model the distribution of O. vulgatus in Europe. The optimal Maxent parameters were RM = 4 and FC = hinge; RM = 4 and FC = hinge and RM = 2.5 and FC = hinge for CLIM1, CLIM2, and CLIM3, respectively. With the exception of one model of CLIM2, all models based on these optimal values yielded AUC values >0.8, which indicated that the different bioclimatic data subsets performed well. The consensus model was therefore computed from a set of 29 estimates of climate suitability.
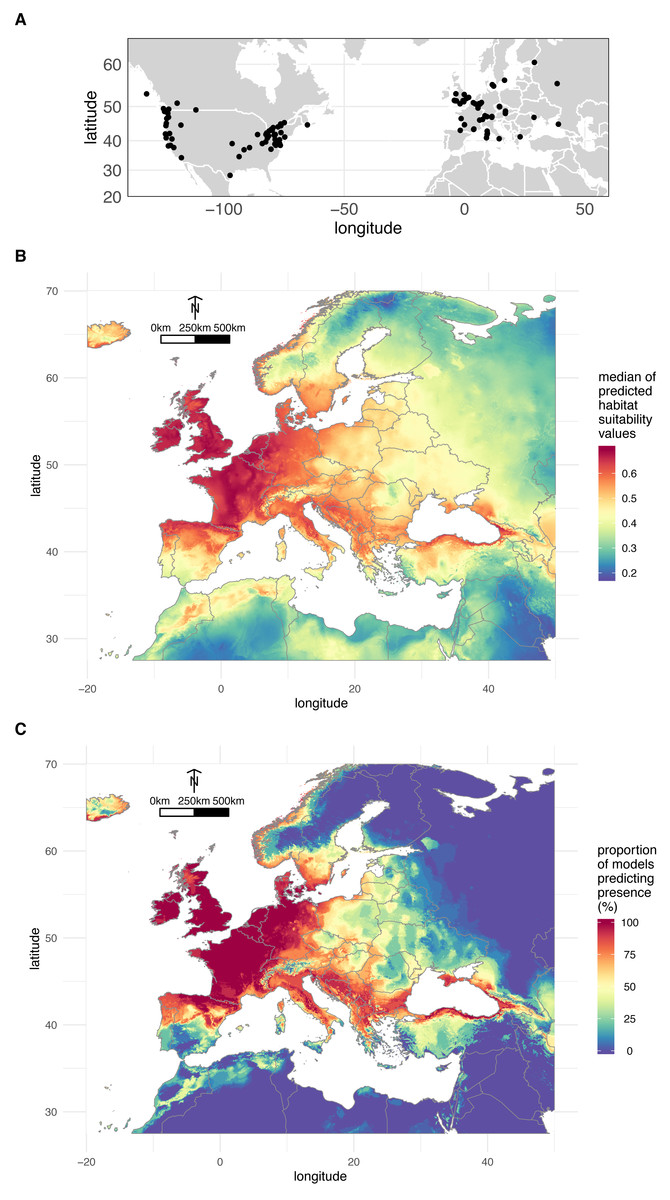
Figure 3: Geographical distribution of O. vulgatus.
(A) Distribution of O. vulgatus occurrences collected from the literature. (B) Consensus model of climate suitability estimated by Maxent: median of model outputs. (C) Consensus model of climate suitability estimated by Maxent: proportion of models predicting O. vulgatus presence in Europe.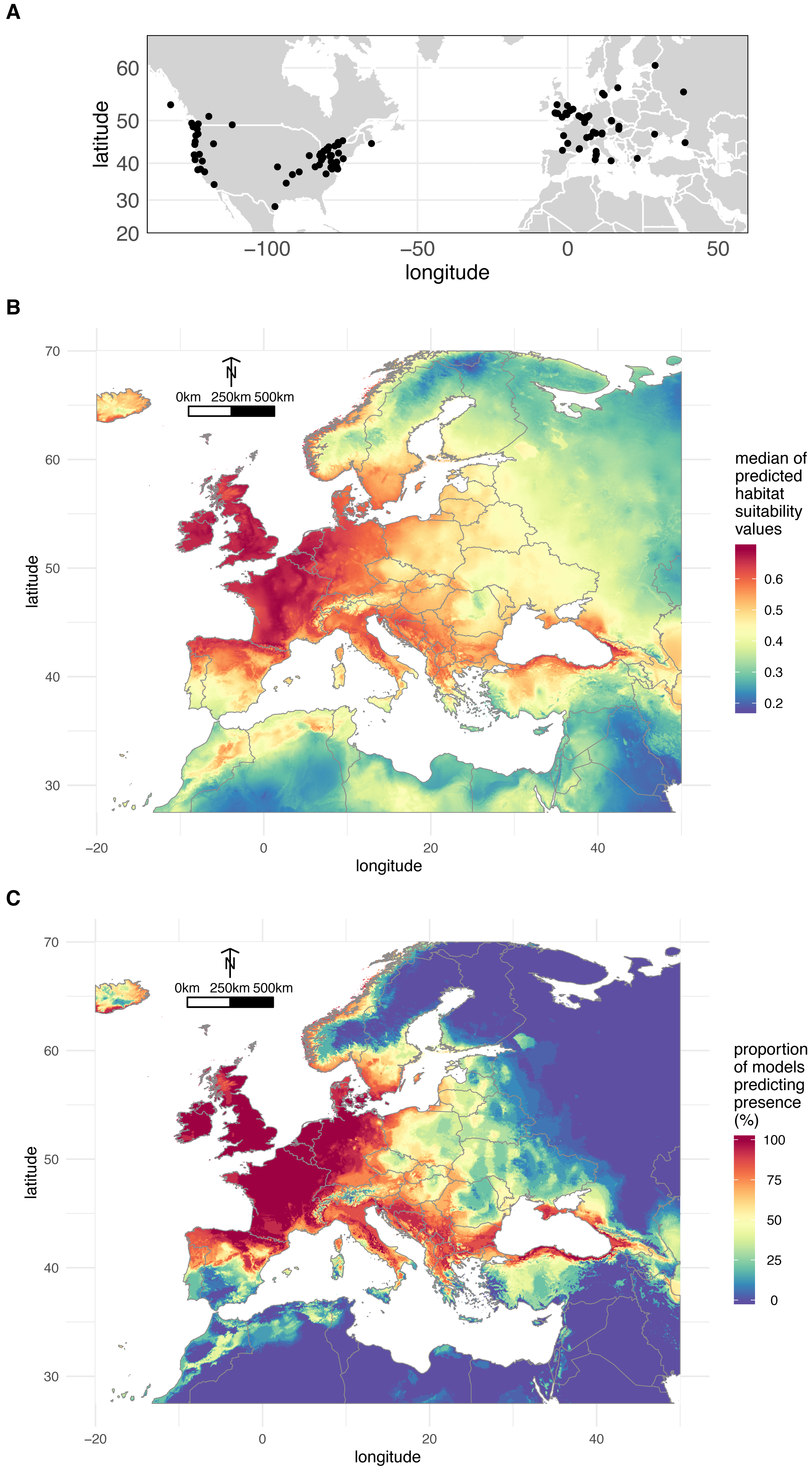{kind=link}
Figure 3B shows the median of the climate suitability values for the 29 models considered. Figure 3C depicts the proportion of the 29 models indicating that the climate is suitable for O. vulgatus. Both Figs. 3B and 3C show that the climate is favorable in very large areas covering most of Western Europe and around the Black Sea. These areas are overlapping with the geographical range of P. spumarius (Cruaud et al., 2018).
Discussion
Ooctonus Haliday, 1833 is a medium-sized genus of Mymaridae containing 37 described species that occur in all zoogeographical regions of the world (Holt et al., 2013) excepted Australasia (Noyes, 2019). O. vulgatus has been reared from the eggs of P. spumarius and studied only once in North America (Weaver & King, 1954). This species is thus poorly known as confirmed by the limited barcoding record. Indeed, only four barcodes are available in BOLD (two from Virginia United States, one from Ontario Canada, and one from British Columbia Canada). As a likely component of aerial plankton, O. vulgatus is expected to be a widespread species distributed in the Holarctic region (ranging from Ireland to the Sakhalin peninsula and from eastern to western coasts of North America, as south as California (Huber, 2012)). The species has been also reported from China (Bai, Jin & Li, 2015) but authors’ illustration casts some doubts about specimen identification. There are only a few unquestionable occurrences in the literature for this species (n = 197). Here, we provide a first report of O. vulgatus in Corsica and assess, for the first time in Europe, its biology as parasitoid of P. spumarius. We also confirm its potential large distribution throughout Europe with modelling approaches. More importantly we show that O. vulgatus potential distribution in Europe (Fig. 3) overlaps that of its host P. spumarius (Cruaud et al., 2018), which is not surprising from a biological point of view but is an interesting result in the framework of biological control. This study is preliminary and predictions, especially because they are based on a limited number of occurrences, are indicative only. This study is a starting point to encourage investigations in other parts of Europe. Sampling efforts should more specifically target areas predicted as suitable for P. spumarius but non-suitable for O. vulgatus such as eastern areas of Europe.
When studied in North America, observed parasitism rates did not exceed 10% of the sampled eggs of P. spumarius (Weaver & King, 1954). Here, we obtained parasitism rates of up to 69%, but absence of parasitism in one sampled site. While we acknowledge sampling four sites is not enough to have a representative view of P. spumarius egg parasitism in Corsica, our results show that parasitism rate can be high though very variable. Further surveys are obviously necessary to better assess the spatial and temporal variability of parasitism rate and understand what is(are) the cause(s) of such variations in Corsica and throughout Europe. Identifying such drivers could open new avenues for conservation biological control against P. spumarius, through the implementation of environments favorable to O. vulgatus in the vicinity of crops susceptible to X. fastidiosa.
The use of mymarids in biological control program has a long history. The most notable instance being the use of Anaphes nitens (Girault, 1928) in several countries to successfully control the eucalyptus weevil, Gonipterus scutellatus Gyllenhal, 1833 (Coleoptera, Curculionidae), which feeds and reproduces on Eucalyptus trees (Doull, 1955). More recently, Cleruchoides noackae Lin and Huber, 2007 has been used in South America to control an invasive sap-feeding pest of Eucalyptus, Thaumastocoris peregrinus Carpintero and Dellapé, 2006 (Hemiptera, Thaumastocoridae) (Martinez, González & Dicke, 2018). Mymarid species were used to control leafhopper vectors of plant pathogens (Hemiptera, Cicadellidae). Anagrus armatus (Ashmead, 1887) regulated Edwardsiana froggatti (Baker, 1925) (Hemiptera, Cicadellidae), a pest of apple in New Zealand, with parasitism rates of the eggs reaching 80% (Dumbleton, 1937). More recently, Cosmocomoidea species were used to target Homalodisca vitripennis (Germar, 1821) (Hemiptera, Cicadellidae) a vector of X. fastidiosa in California (Irvin & Hoddle, 2010). In all these cases, mymarids helped regulate pest population growth.
However, before any attempts to regulate populations of P. spumarius are made, we need to enrich our knowledge on O. vulgatus. In particular, the degree of specificity of the P. spumarius – O. vulgatus interaction needs to be determined to avoid non-target effect of augmentative biocontrol (Van Driesche & Hoddle, 2016). We also need to evaluate our ability to consistently rear O. vulgatus in controlled conditions, one of the key obstacles to the use of mymarids in biological control programs (but see Martinez, González & Dicke, 2018). Finally, parasitoids can have complex effects on vector-borne disease by either increasing (Jeger et al., 2011) or decreasing (Martini, Pelz-Stelinski & Stelinski, 2014) pathogen spread. Further research is still needed to better understand the impact of such tri-trophic interactions on plant disease dynamics. While O. vulgatus does not directly impact transmission capacity of P. spumarius, by killing its host at an early stage of development, it reduces the number of vectors that may acquire the bacterium from an infected host-plant and become able to transmit it.
Again, we consider this study as a starting point to encourage research on this parasitoid wasp to assess whether it could contribute to reduce the spread and impact of X. fastidiosa in Europe. Increasing egg parasitism of P. spumarius in the fall might significantly reduce population size in the next year and possibly the transmission of the bacterium, without resorting to chemical treatments.
Supplemental Information
Illustration of the hatching experiment to quantify parasitism rates in four sampling sites
Bunches of branches of Cistus monspeliensis were collected on four sites, at the following GPS coordinates: 42.984205°N, 9.395287°E (Ersa);
42.338849°N, 9.180636°E (Tralonca); 42.274756°N, 9.487185°E (Canale-di-Verde); 41.931726°N, 9.343731°E (Ventiseri).
After inspection of the back of each leaf of C. monspeliensis collected in the Corsican maquis, parts of leaf containing whitish clusters were cut and put on moistened filter paper in Petri dishes (A). (B) shows a zoom on the leaf, the white arrow pointing to the P. spumarius egg cluster. Leaf cuttings were observed under a binocular microscope to confirm the presence of P. spumarius eggs. (C–E) show several groups of P. spumarius eggs. The orange pigmented spots and the black shields that can both be seen especially in (C) and (D) are respectively the future eyes of the larva and the characteristic “egg burster” used by the larva to break the egg shell (Weaver & King, 1954). Panel E shows an egg cluster embedded in the frothy cement described by Weaver & King (1954). In panel D, the egg on the right has already hatched whereas the egg on the left hasn’t. Finally, panel F shows a first instar larva of P. spumarius in dorsal view while panel G show a male of O. vulgatus in ventral view.
Supplementary methods
This appendix provides details regarding the species distribution modelling framework used in the study
Maximum likelihood phylogenetic tree including the newly generated sequences and all COI barcodes available in BOLD for Ooctonus species (last access July 12,2019)
Bootstrap (100 replicates) at nodes. The newly generated sequences are in red. The only four sequences identified as O. vulgatus in BOLD are in blue.