
Evaluation of potential reference genes for real-time qPCR analysis in a biparental beetle, Lethrus apterus (Coleoptera: Geotrupidae)
- Published
- Accepted
- Received
- Academic Editor
- Ilaria Negri
- Subject Areas
- Entomology, Genetics, Molecular Biology
- Keywords
- Insect, Parental care, Housekeeping gene
- Copyright
- © 2017 Nagy et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2017. Evaluation of potential reference genes for real-time qPCR analysis in a biparental beetle, Lethrus apterus (Coleoptera: Geotrupidae) PeerJ 5:e4047 https://doi.org/10.7717/peerj.4047
Abstract
Hormones play an important role in the regulation of physiological, developmental and behavioural processes. Many of these mechanisms in insects, however, are still not well understood. One way to investigate hormonal regulation is to analyse gene expression patterns of hormones and their receptors by real-time quantitative polymerase chain reaction (RT-qPCR). This method, however, requires stably expressed reference genes for normalisation. In the present study, we evaluated 11 candidate housekeeping genes as reference genes in samples of Lethrus apterus, an earth-boring beetle with biparental care, collected from a natural population. For identifying the most stable genes we used the following computational methods: geNorm, NormFinder, BestKeeper, comparative delta Ct method and RefFinder. Based on our results, the two body regions sampled (head and thorax) differ in which genes are most stably expressed. We identified two candidate reference genes for each region investigated: ribosomal protein L7A and RP18 in samples extracted from the head, and ribosomal protein L7A and RP4 extracted from the muscles of the thorax. Additionally, L7A and RP18 appear to be the best reference genes for normalisation in all samples irrespective of body region. These reference genes can be used to study the hormonal regulation of reproduction and parental care in Lethrus apterus in the future.
Introduction
Hormonal regulation in insects generates great interest among entomologists but hormones have only been studied in detail in a few species (Gullan & Cranston, 2014). Insect hormones of particular interest include juvenile hormones, ecdysteroids and neuropeptides. These molecules regulate a vast number of physiological and developmental processes as well as behaviours (Gäde, Hoffmann & Spring, 1997). Studying these hormones used to be difficult considering their small amount and the occasional instability (Gullan & Cranston, 2014). The technical revolution of molecular biology and genetics, however, made it attainable to discover the details of genetic and hormonal regulation in insects (Raikhel, Brown & Belles, 2005). Some of the processes controlled by hormones mentioned above, such as ecdysis (Mykles et al., 2013), are already well described. Nevertheless, there are many other interesting physiological and behavioural mechanisms, like parental care, the hormonal regulation of which are not well understood (Panaitof et al., 2016). One way to increase our understanding of hormonal regulation is to identify patterns of gene expression associated with the hormones in question (Champagne & Curley, 2012).
Real-time quantitative polymerase chain reaction (RT-qPCR) is a commonly used method for analysing gene expression as it is a sensitive, fast and reproducible method; moreover, it requires only a minimal amount of RNA (Radonić et al., 2004). With this method, gene expression levels can be measured simultaneously in several different samples for a limited number of genes. Gene expression analyses with RT-qPCR, however, require some kind of normalisation in order to control the variation caused by stochastic processes occurring during the analytic procedure (Vandesompele et al., 2002). This normalisation is usually achieved by taking into account the expression level of so-called reference genes (VanGuilder, Vrana & Freeman, 2008). These genes are usually selected from housekeeping genes, which produce proteins vital for maintaining fundamental cell functions, like ribosomal or cytoskeletal proteins. Therefore, the expression levels of these reference genes are thought to be relatively stable. Thus, comparing the expression level of genes of interest with the expression levels of the reference genes, we can eliminate the differences caused by the different amount and quality of starting material. With this method we are able to control for differences occurring due to technical errors during sample preparation as well (e.g., RNA isolation and cDNA synthesis, Radonić et al., 2004). Nonetheless, expression levels of the housekeeping genes may also vary considerably under certain circumstances because they can be involved in processes other than maintenance functions of the cell, e.g., apoptosis (Nicholls, Li & Liu, 2012), cytokinesis (D’Souza-Schorey & Chavrier, 2006) and development (Zhou et al., 2015). Therefore, a given housekeeping gene cannot automatically serve as reference gene, and normalisation with unstable reference genes can lead to erroneous quantification results and conclusions (Thellin et al., 1999). Consequently, reference genes must be carefully selected so that their expression levels are similar between the different samples and should not be influenced significantly by different experimental conditions (VanGuilder, Vrana & Freeman, 2008). According to Vandesompele et al. (2002), the combination of two or more reference genes is highly recommended for normalisation to obtain more accurate results. In case of multiple reference genes, it is advised to use the geometric mean for normalisation since it better controls for extreme values and the possible differences between expression levels of the different genes (Vandesompele et al., 2002).
Lethrus apterus (Laxmann, 1770) (Coleoptera: Geotrupidae) is an earth-boring beetle that has biparental care during which the parents provision food for their offspring in advance their hatching (Kosztolányi et al., 2015). This kind of parental care is a complex and relatively rare trait among insects (Smiseth, Kölliker & Royle, 2012) and makes this beetle an outstanding model species for studying the hormonal background of parental care. In order to do so, however, stably expressed reference genes have to be identified.
In recent years, numerous studies aimed to identify stable reference genes in insects (Lord et al., 2010; Ponton et al., 2011; Bansal et al., 2012; Li et al., 2013; Pan et al., 2015; Yu et al., 2016). Nevertheless, there is a lack of reference gene studies that use individuals from natural populations. Our objective in this study was to examine the expression stability of several housekeeping genes in Lethrus apterus across different times of the breeding period in a natural population in order to identify the most stable reference gene(s). With the right combination of reference genes, the expression levels of hormone regulating genes involved in parental care in Lethrus apterus can be examined accurately in the future. Based on the literature (Shi et al., 2013; Liang et al., 2014; Zhu et al., 2014; Yang et al., 2015), we probed eleven housekeeping genes.
Materials and Methods
Sample collection
Samples were collected near Dorogháza, northern Hungary (47°59′29″N, 19°53′36″E) on 16th April, 4th May and 28th May in 2015, which dates corresponded to the beginning, middle and end of the breeding season of Lethrus apterus, respectively. Sample collection was approved by the Nothern Hungarian Inspectorate for Environment Protection and Nature Conservation (No. 9007-8/2014). The first sampling date represents the period of mate choice, while the second and third samplings were done during the period when parents were collecting leaves for the offspring. On each sampling dates, head and thorax samples were collected from eight males and eight females. All tissues were removed from the head capsule and muscle samples were taken from the thorax. Samples were collected in the field in less than five minutes after euthanizing the individuals. Each head and thorax sample was put immediately into separate eppendorf tubes which already contained 600 µl RNAlater® Stabilization Solution (Thermo Fisher Scientific, Waltham, MA, USA), then stored at −20 °C in the laboratory in order to inhibit RNase enzyme activity until RNA extraction.
RNA extraction and cDNA synthesis
Total RNA was isolated from each samples using TRIzol® Reagent (Thermo Fisher Scientific, Waltham, MA, USA) following the manufacturer’s instructions. The extracted RNA was eluted in 15–30 µl RNase-free water, depending on the pellet size. Yield of RNA was quantified by NanoDrop 1000 Spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). To eliminate genomic DNA, samples were treated with RQ1 RNase-Free DNase (Promega, Madison, WI, USA) just before the reverse transcription. First strand cDNA was synthesized from 1 µg DNA-free RNA using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA).
Reference gene selection and primer design
Using a draft genome of Lethrus apterus (unpublished data) eleven reference genes, which were already described as stable reference genes in other arthropods, were selected (Table 1). We manually designed primers (Table 2) using the web-based Sequence Manipulation Suite (Stothard, 2000) and Multiple Primer Analyzer (Thermo Fisher Scientific, Waltham, MA, USA) in order to avoid the forming of possible secondary structures of the primers. To check the specificity of primer pairs and to determine optimal annealing temperature, PCR reactions were performed in 10 µl volumes containing the following components: 10x buffer, 2 mM MgCl2, 0.2 mM dNTP, 0.02 U/µL Taq DNA polymerase enzymes (DreamTaq Green, Thermo Fisher Scientific, Waltham, MA, USA), 0.2 µM forward and 0.2 µM reverse primer and 0.1 µg cDNA. PCR conditions were optimized by determining the optimal annealing temperature using temperature gradient ranging from 54 °C to 62 °C for primer binding. In this study, we used ABI Veriti® 96-Well Thermal Cycler (Applied Biosystems, Foster City, CA, USA). Cycling conditions consisted of a denaturing step at 95 °C for 2 min followed by 40 cycles at 95 °C for 30 sec, at a temperature gradient (54 °C, 56 °C, 58 °C, 60 °C or 62 °C) for 30 s and at 72 °C for 90 s, and finally at 72 °C for 10 min. PCR amplicons were run on 1% agarose gel stained with GelRed™ (Biotium, Fremont, CA, USA).
| Gene | Symbol used | Function | Reference |
|---|---|---|---|
| glyceraldehyde 3-phosphate dehydrogenase | GAPDH | glycolytic enzyme | Liang et al. (2014) |
| tubulin alpha-1 chain | TUB1a | cytoskeletal structural protein | Liang et al. (2014) |
| elongation factor 1-alpha | EF1a | protein synthesis | Liang et al. (2014) |
| elongation factor 2 | EF2 | protein synthesis | Zhu et al. (2014) |
| ADP-ribosylation factor-like protein 1 | ARF1 | GTP-binding protein | Shi et al. (2013) |
| ADP-ribosylation factor 4 | ARF4 | GTP-binding protein | Shi et al. (2013) |
| ribosomal protein S8 | RPS8 | structural constituent of ribosome | Yang et al. (2015) |
| ribosomal protein L4 | RP4 | structural constituent of ribosome | Shi et al. (2013) |
| ribosomal protein L7A | L7A | structural constituent of ribosome | Zhu et al. (2014) |
| ribosomal protein L10 | L10 | structural constituent of ribosome | Zhu et al. (2014) |
| ribosomal protein L18 | RP18 | structural constituent of ribosome | Shi et al. (2013) |
| Gene | GenBank accession number | Primer sequence (5′to 3′)a | Amplicon length (bp) | Tm (°C)b | E (%)c | R2d |
|---|---|---|---|---|---|---|
| GAPDH | KY786279 | F: GCCATTCCAGTAAGTTTTCCATTGAG | 157 | 85.0 | 100.75 | 0.91 |
| R: GCTGTTACTGCTACACAAAAGAC | ||||||
| TUB1a | KY786273 | F: CAGACTGCACGTTGGACTTTAGC | 172 | 83.6 | 100.04 | 0.96 |
| R: TACAGAGGAGATGTTGTCCCCAAG | ||||||
| EF1a | KY786281 | F: AAACCTTTGCGTCTTCCACTACAGG | 184 | 81.7 | 99.83 | 0.94 |
| R: CTTCAGTTGTAAGACCAACAGGTG | ||||||
| EF2 | KY786280 | F: GATGAGAAATCCACATGTCCAG | 244 | 82.0 | 102.00 | 0.86 |
| R: CGACTCCCTAGTATCAAAGG | ||||||
| ARF1 | KY786283 | F: GTATGACAGTAGCTGAAGTTC | 141 | 81.4 | 112.70 | 0.84 |
| R: CTGTTTTGTAAAGCATTGGC | ||||||
| ARF4 | KY786282 | F: TAGTACGGACGGTCAAGTC | 197 | 89.1 | 105.91 | 0.81 |
| R: GTAGACCGTCACCTGTTATGGC | ||||||
| RPS8 | KY786274 | F: CATTATGTACGTACGAGAGGAGGCAACG | 200 | 84.0 | 99.96 | 0.91 |
| R: TCTAAAGGGAGTAGCGTCGATAACG | ||||||
| RP4 | KY786275 | F: TAATGGACCACGACGCTGTATGC | 248 | 84.5 | 100.33 | 0.92 |
| R: CGTACCAGCTTTAGTAATGAGCAAGG | ||||||
| L7A | KY786277 | F: TAGCGACTCAACTGTTCAAGG | 224 | 84.8 | 99.54 | 0.95 |
| R: CCTCAATTGGATCGACGTCATGTG | ||||||
| L10 | KY786278 | F: CGTAGAGCCTCGATAACTTGG | 210 | 84.7 | 99.33 | 0.94 |
| R: TCATGTGCTGGAGCTGATAGG | ||||||
| RP18 | KY786276 | F: TTGTAACCACATGAACGCCTACG | 186 | 85.2 | 99.75 | 0.96 |
| R: AGTTAGCTTTACGTTCACCTACTGG |
Real-time quantitative PCR
RT-qPCR was performed on a QuantStudio 12K Flex Real-Time PCR System (Applied Biosystems, Foster City, CA, USA) using SYBR® Green PCR Master Mix (Applied Biosystems, Foster City, CA, USA) and ROX Passive Reference Dye (Affymetrix, Santa Clara, CA, USA). Amplifications were carried out under the following conditions: initial denaturation at 95 °C for 10 min followed by 40 cycles of 10 sec at 95 °C and for 1 min at the optimal annealing temperature. This was followed by a melting curve analysis in which the temperature raised from 65 °C to 95 °C in sequential steps of 0.05 °C for 1 s. Three technical replicates were performed for each biological sample, and the average cycle threshold (Ct) values of triplicates were calculated. Furthermore, no-template control was done in order to check whether primer-dimers or contamination with amplified PCR product were detectable. Five 5-fold serial dilution was made from cDNA samples to create a standard curve, and the amplification efficiency was determined for each candidate gene. The efficiency (E) values were calculated according to the equation: E = (10(−1∕slope) − 1) × 100, where slope is the slope of the standard curve (Radonić et al., 2004).
Statistical analysis of raw Ct values
In order to examine the differences between sample groups, random intercept mixed-effects models were used with sample id as a random factor for each gene. Significance of fixed terms was investigated by likelihood ratio tests. For likelihood ratio tests models were fitted using Maximum Likelihood estimation. The analyses were carried out using “lme4” (Bates et al., 2014) and “car” (Fox & Weisberg, 2011) packages in the R statistical environment version 3.3.2 (R Core Team, 2016).
Determination of reference gene expression stability
In order to determine the expression stability of the selected reference genes, we used the following methods: geNorm (Vandesompele et al., 2002), NormFinder (Andersen, Jensen & Ørntoft, 2004), BestKeeper (Pfaffl et al., 2004), delta Ct method (Silver et al., 2006) and RefFinder (Xie et al., 2012). For the analyses with the geNorm and NormFinder procedures, the average Ct values were transformed to relative quantities by dividing sample values by the lowest average Ct value. For calculations by BestKeeper, delta Ct method and RefFinder, the untransformed average Ct values were used. All calculations, except the ones done by the web-based RefFinder, were carried out in R with “NormqPCR” package (Perkins et al., 2012).
geNorm calculates the expression stability value M by assessing the mean pairwise expression ratio for each candidate gene against all the other candidates (Vandesompele et al., 2002). The basic assumption of this method is that the expression ratio between two reference genes is identical across the samples. The lower the M value the more stable the expression of the candidate reference gene. Stepwise exclusion of the genes with the highest M value results in the selection of the two most stably expressed reference genes in the tested samples both sharing the same M value. Vandesompele et al. (2002) also suggest not to accept candidate genes as stably expressed reference genes with M value higher than 1.5. Moreover, the procedure determines the normalisation factor by taking the geometric mean of the expression levels from the most stable genes and then additively recalculating with each of the next most stable gene. The pairwise variation, Vn∕n+1 between two sequential normalisation factors is then calculated in order to determine the effect of each newly added gene to the normalisation factor. The optimum number of genes is the lowest number of genes with Vn∕n+1 less than 0.15 (Vandesompele et al., 2002).
NormFinder determines the stability of the candidate reference genes by measuring the intra- and intergroup variation between user specified groups (e.g., male and female groups or treated and control groups) first. Stability values for each candidate gene are then calculated by adding the two sources of variation. The lowest stability value means the most stable expression (Andersen, Jensen & Ørntoft, 2004).
BestKeeper calculates, for each candidate reference gene across the samples, the geometric mean, the arithmetic mean, the minimal and the maximal Ct values, in addition to the average absolute deviation from the arithmetic mean. Genes with the lowest average absolute deviation can be considered as stably expressed reference genes. BestKeeper Index is calculated as the geometric mean of the Ct values of the candidate reference genes. Inter-gene relations are estimated by performing pairwise correlation analyses of all possible reference gene pairs. Furthermore, correlation between the expression level of each candidate gene and the BestKeeper Index is calculated, describing the relation between the index and the contributing genes by the Pearson correlation coefficient, coefficient of determination and the corresponding p-value (Pfaffl et al., 2004).
The delta Ct method compares relative expression of pairs of candidate genes within each sample in order to identify the stably expressed housekeeping genes. If the ΔCt value of the two genes fluctuates when analysed in different samples, it means that one or both genes are variably expressed. If the ΔCt value remains constant, both genes are stably expressed among the samples (Silver et al., 2006).
Each procedure mentioned above uses different algorithms to calculate an expression stability value which represents the suitability of the candidate genes as reference genes, therefore the ranking of the examined genes according to the methods may vary. The web-based tool RefFinder (Xie, 2012) was used in order to combine our results and rank the candidate genes. This user-friendly program integrates the four methods mentioned above. Using the ranking from each program, it assigns an appropriate weight to an individual gene and calculates the geometric mean of their weights for the overall ranking. The lowest rank indicates the most stably expressed gene (Xie et al., 2012).
For each analysis, except for NormFinder, seven sample groups were used: all samples irrespective of body part or sex; head samples irrespective of sex; male head samples; female head samples; thorax samples irrespective of sex; male thorax samples; female thorax samples. The calculation by NormFinder requires subgroup specification, therefore, body regions were set as subgroups for the analysis of all samples. In order to investigate the effect of sexes, male and female subgroups were specified for the analysis of head and thorax sample groups separately. In this way, three sample groups, each divided into two subgroups, were analysed by NormFinder.
Results
Transcriptional profiling of candidate reference genes
Before the evaluation of expression stability of the eleven candidate genes, specificity of each primer pair was checked on 1% agarose gel which showed single products with the expected sizes. Moreover, gene-specific amplification was confirmed by single melting curve peaks. These results indicate that no primer-dimers or nonspecific amplification products were formed. Additionally, no fluorescent signals were detected in the negative control during the RT-qPCR. Each amplicons were sequenced and annotated to the sequences from which the primer design was based in order to check that the correct genes were amplified. The sequences are available in File S1. The efficiency of the eleven candidate genes ranged from 99.33 to 112.70%. The efficiency values and other basic information of the RT-qPCR required based on the guideline of Bustin et al. (2009) are included in Table 2.
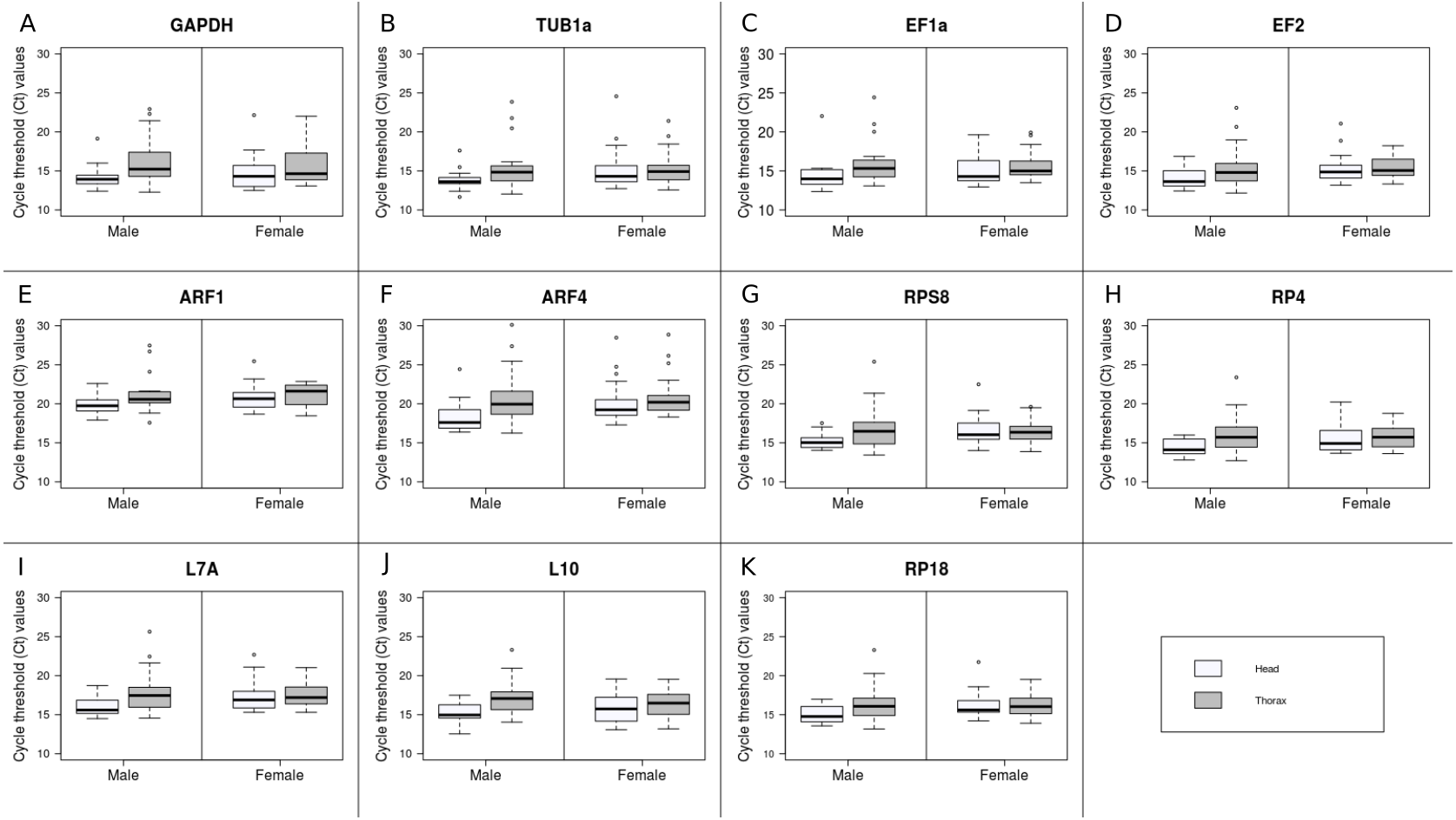
Raw Ct values ranged from 11.66 (TUB1a) to 30.12 (ARF4) (Fig. 1). The mean and standard deviation (SD) of the Ct values across all samples were calculated for each gene (Table 3). Since the mean Ct values ranged between 15 and 30 for all the candidate reference genes, all of them were analysed further (Kozera & Rapacz, 2013). ARF1 had the least variable expression level with the lowest SD value (SD = 1.85), while ARF4 had the most variable expression level (SD = 3.04). Low average Ct values indicate high expression level in TUB1a and EF2 (Ctmean = 15.11), on the other hand, high Ct values of ARF1 (Ctmean = 20.77) indicated low expression.

Figure 1: Expression profiles of the 11 candidate reference genes.
{kind=link}
| Genes | Mean | SD | CV |
|---|---|---|---|
| GAPDH | 15.27 | 2.62 | 0.17157826 |
| TUB1a | 15.11 | 2.64 | 0.17471873 |
| EF1a | 15.44 | 2.34 | 0.1515544 |
| EF2 | 15.11 | 2.05 | 0.13567174 |
| ARF1 | 20.77 | 1.85 | 0.08907078 |
| ARF4 | 20.22 | 3.04 | 0.15034619 |
| RPS8 | 16.38 | 2.12 | 0.12942613 |
| RP4 | 15.52 | 1.94 | 0.125 |
| L7A | 17.27 | 2.1 | 0.12159815 |
| L10 | 16.15 | 1.97 | 0.12198142 |
| RP18 | 16.02 | 1.93 | 0.12047441 |
Based on the likelihood ratio tests, sex had no significant effect on the expression level of the candidate genes. However, significant effect of body region was found in case of six genes: GAPDH, EF1a, ARF1, ARF4, RP4 and L10. The interaction of sex and bodypart had no significant effect on the expression level of the candidates, except for RPS8 (Table 4).
| Gene | Sex | Bodypart | Sex*Bodypart | |||
|---|---|---|---|---|---|---|
| χ2 | P | χ2 | P | χ2 | P | |
| GAPDH | 0.019 | 0.889 | 5.668 | 0.017 | 0.965 | 0.326 |
| TUB1a | 0.642 | 0.423 | 1.960 | 0.162 | 1.995 | 0.158 |
| EF1a | 0.002 | 0.968 | 4.096 | 0.043 | 0.857 | 0.355 |
| EF2 | 1.216 | 0.270 | 2.271 | 0.132 | 2.030 | 0.154 |
| ARF1 | 0.476 | 0.490 | 4.147 | 0.042 | 2.459 | 0.117 |
| ARF4 | 1.417 | 0.234 | 4.885 | 0.027 | 1.289 | 0.256 |
| RPS8 | 0.586 | 0.444 | 2.307 | 0.129 | 4.007 | 0.045 |
| RP4 | 0.356 | 0.551 | 4.272 | 0.039 | 2.384 | 0.127 |
| L7A | 0.556 | 0.456 | 3.087 | 0.079 | 2.901 | 0.089 |
| L10 | 0.246 | 0.620 | 8.509 | 0.004 | 3.319 | 0.068 |
| RP18 | 1.028 | 0.311 | 2.336 | 0.126 | 2.896 | 0.089 |
Notes:
Significant effects are highlighted in bold.
Expression stability of candidate reference genes
Based on geNorm analysis for all samples, eight candidate genes had an M value below the threshold of 1.5 (Table S1). The results show that the lowest M value was 0.390 for RPS8 and L7A. Among the head samples irrespective of sex, all of the tested genes except L10 had an M value below 1.5, and RPS8 and RP18 were co-ranked as the most stable genes from the candidates (M = 0.304). Furthermore, the same two genes had the lowest M value considering male and female head samples separately (M = 0.264 for females and M = 0.346 for males). In case of the thorax samples irrespective of sex, eight genes had an M value below the threshold. RPS8 and L7A were the most stable candidate gene pair with an M value of 0.358. In thorax samples collected from females, RPS8 and RP18 were the most stable genes as well with an M value of 0.222. However, in thorax samples of males, RPS8 and L7A were ranked as the best reference gene pair (M = 0.288).
According to NormFinder, L7A was the most stable gene when calculating with all samples divided into groups of head and thorax samples (Table S2). The second and third genes were RP4 and RPS8, indicating that these are also worth considering as reference genes. In the case of specifying males and females as subgroups within head and thorax samples, L7A was found again to be the most stably expressed gene among the candidate ones. In both head and thorax samples, L7A was followed by similar ranking order: EF2, RP4, RP18 and RPS8 as second, third, fourth and fifth genes, respectively.
Based on BestKeeper, across all samples ARF1 had the lowest mean absolute deviation (MAD) value; however, L7A had the highest correlation r value (Table S3). In the group of head samples irrespective of sex, ARF1 had the lowest MAD value, while among the thorax samples irrespective of sex, L10 was the most stable according to the MAD value. This was surprising as the other programs ranked this gene consistently as one of the least stable genes. On the other hand, in both head and thorax samples, L7A had the highest r value. In head samples of males RPS8 (MAD = 0.827), and of females ARF1 (MAD = 1.122) were the most stable candidate genes. In both male and female head samples, L7A had the highest correlation r value. Considering male thorax samples L10 had the lowest MAD value (MAD = 1.576), while in female thorax samples EF2 was ranked as the most stable with MAD value 1.236. L7A had the highest r value in female thorax samples, however, in male thorax samples EF2 had the highest correlation r value.
According to the delta Ct method, L7A was the most stable gene among the candidates overall with the stability value always below 1.0 (Table S4).
Finally, the candidate genes were evaluated by RefFinder to combine the results of individual methods (Table 5). Using all samples irrespective of body region and sex, and separately the head and thorax samples irrespective of sex, L7A was ranked first, as the most stably expressed gene among the candidate reference genes. In head samples, RP18 was co-ranked with L7A as the most stable reference genes. In thorax samples, RP4 was ranked on the second place. In female head and thorax samples, RP18 was the most stably expressed gene of the candidates. Considering head samples of males, RPS8 was ranked on the first place, while in thorax samples of males, L7A was ranked as the best reference gene. L7A was ranked on the second place in all subgroups, with the exception of male thorax samples, where EF2 was the second best reference gene according to RefFinder.
| Rank | All samples | Head samples | Thorax samples | ||||
|---|---|---|---|---|---|---|---|
| All head samples | Male head samples | Female head samples | All thorax samples | Male thorax samples | Female thorax samples | ||
| 1 | L7A | L7A | RPS8 | RP18 | L7A | L7A | RP18 |
| 2 | RP18 | RP18 | L7A | L7A | RP4 | EF2 | L7A |
| 3 | RPS8 | RPS8 | RP18 | RPS8 | RP18 | RPS8 | EF2 |
| 4 | EF2 | ARF1 | EF2 | EF2 | RPS8 | RP4 | RPS8 |
| 5 | ARF1 | RP4 | TUB1a | ARF1 | EF2 | L10 | RP4 |
| 6 | RP4 | EF2 | RP4 | RP4 | L10 | EF1a | L10 |
| 7 | EF1a | GAPDH | ARF1 | EF1a | ARF1 | RP18 | ARF1 |
| 8 | TUB1a | TUB1a | GAPDH | GAPDH | EF1a | ARF1 | EF1a |
| 9 | L10 | EF1a | L10 | ARF4 | TUB1a | ARF4 | TUB1a |
| 10 | GAPDH | L10 | ARF4 | L10 | ARF4 | TUB1a | GAPDH |
| 11 | ARF4 | ARF4 | EF1a | TUB1a | GAPDH | GAPDH | ARF4 |
Optimal number of reference genes
To determine the minimal number of genes necessary for normalisation, the V-value was computed by geNorm. The results demonstrated that across all samples V2∕3 was the first V-value lower than the cut-off value of 0.15 (Fig. 2). Considering separately the head and thorax samples, V2∕3 was again lower than 0.15. Separate analyses of female and male samples within head and thorax groups showed that V2∕3 was also the first value below the threshold in all cases (results not shown). Therefore, two stably expressed reference genes are sufficient for normalisation in any case of sample classification.
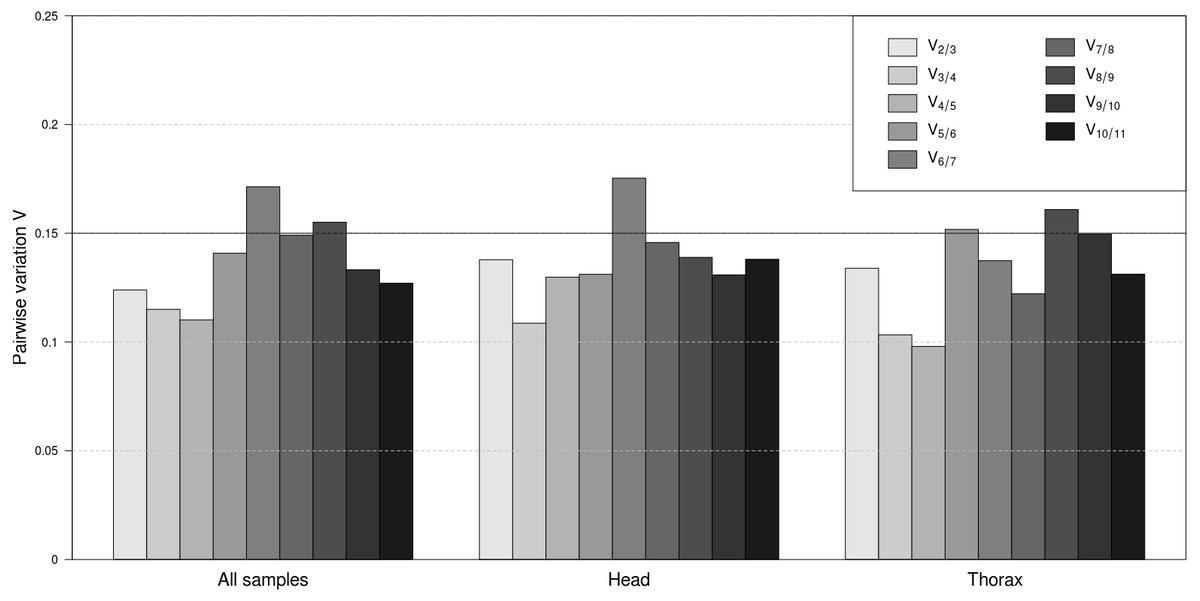
Figure 2: Pairwise variation analyses by geNorm to determine the optimal number of reference genes for accurate normalization.
Pairwise variation for all samples together, as well as separately for head and thorax samples. The lowest number of genes with Vn∕n+1 less than 0.15 means the optimum number of genes.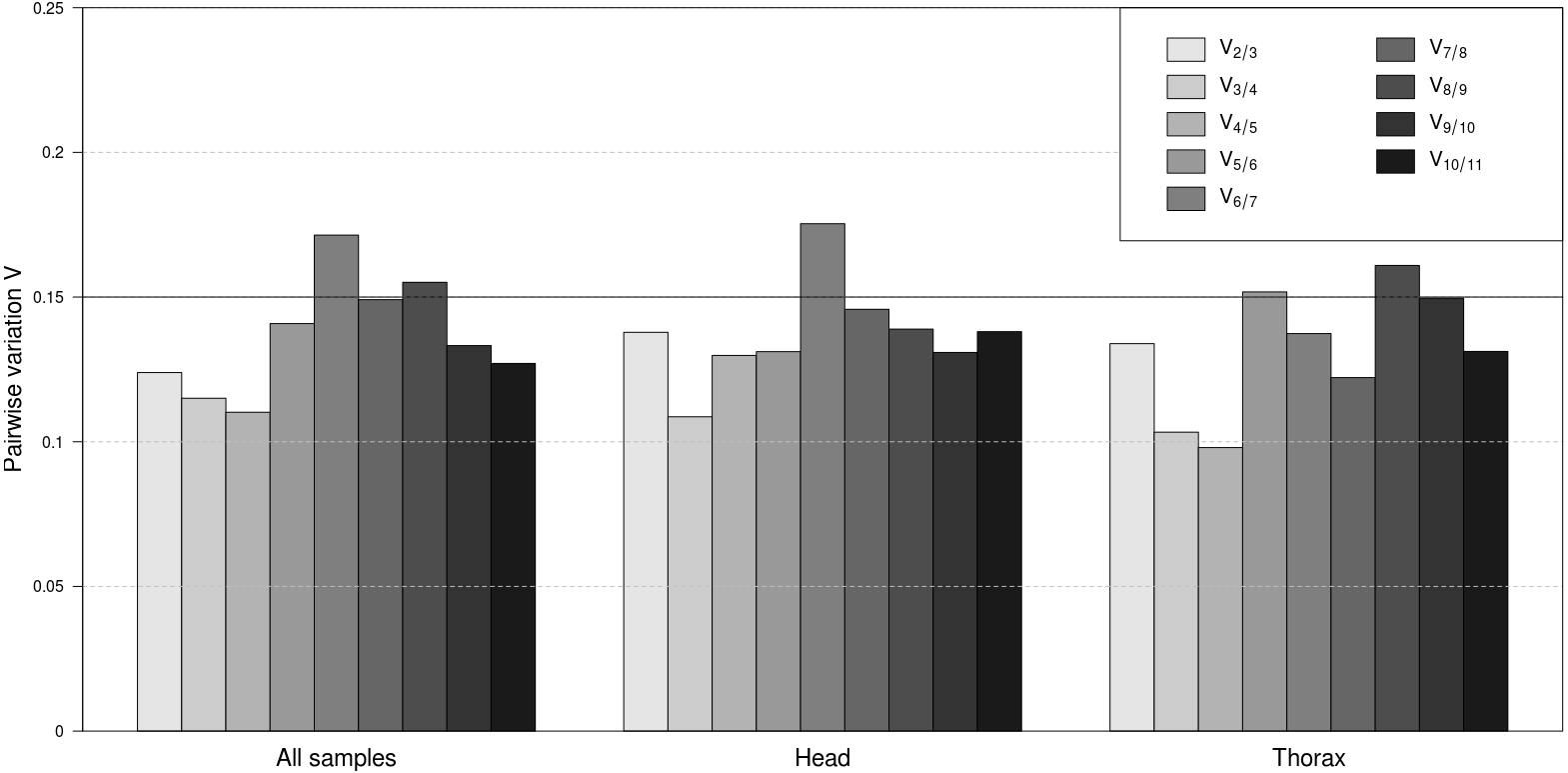{kind=link}
Discussion
RT-qPCR is a widely used method for measuring gene expression levels due to its relatively low cost, high accuracy and sensitivity. A critical step of this method is data normalisation which requires careful selection of reference genes for the given experimental or environmental conditions. With these stably expressed genes, technical errors and variance resulting from the method can be moderated (Udvardi, Czechowski & Scheible, 2008). Several studies have examined the stability of reference genes in various insect species in the past decade and these studies suggest that no universally stable reference gene can be found that is applicable for all species, tissue types and experimental conditions. Hence, it is necessary to identify the most suitable reference genes for the specific circumstances in a given study for a given species (Zhu et al., 2014).
In the present study, variation in expression levels of eleven housekeeping genes were evaluated across a span of 1.5 months covering most of the breeding period of the biparental beetle Lethrus apterus. To date, no study investigated the possible reference genes either in this species, or in the family of Geotrupidae. We analyzed the expression stability of the candidate reference genes by four frequently used programs: geNorm, NormFinder, BestKeeper and comparative delta Ct method. The outcomes of these programs can vary because of the differences in the algorithms. Therefore, the combined use of them ensures more reliable results. For this purpose, RefFinder, a freely available web-based tool was used to calculate a comprehensive ranking value for each candidate gene.
According to the comprehensive ranking by RefFinder, the most stably expressed reference gene was L7A across all samples, irrespective of body region and sex. Based on the results of geNorm analysis, two reference genes are sufficient for normalisation in gene expression analysis in Lethrus apterus during the breeding period. For accurate normalisation, we recommend the use of L7A and RP18 in head samples irrespective of sex. When considering the sexes separately, RPS8 and L7A should be used for head samples of males, and RP18 and L7A for females. In thorax samples irrespective of sex, L7A and RP4 are the best reference genes. In case of thorax samples, L7A and EF2 are recommended for normalisation in males, RP18 and L7A in females.
Consistent with our results, ribosomal proteins are reported to be the best reference genes in many insect species. In a study by Zhu et al. (2014), ribosomal protein L7A was ranked as one of the best reference genes in Spodoptera exigua in different tissues, specific larval physiological stages and male individuals. Studies of other coleopterans gave similar results: RP4 and RP18 were the best reference genes in Leptinotarsa decemlineata (Shi et al., 2013), RPS3 (ribosomal protein S3), RPL13a (ribosomal protein 13a) and RPS18 (ribosomal protein S18) were suitable reference genes for Tribolium castaneum (Lord et al., 2010; Sang et al., 2015), and RPL22e (ribosomal protein 22e) was one of the best reference genes in Mylabris cichorii both in males and females (Wang et al., 2014). In other species, e.g., in Drosophila melanogaste r Rpl32 (ribosomal protein L32) was a suitable reference gene in individuals on different diets (Ponton et al., 2011), and in Aphis craccivora, RPS8, RPL14 (ribosomal protein L14), and RPL11 (ribosomal protein L11) were the three most stable housekeeping genes across different developmental stages and temperature conditions (Yang et al., 2015).
Interestingly, two frequently used reference genes, GAPDH and TUB1a were ranked as less stable genes in this study, beside ARF4, with stability values above the threshold values of all the programs used. L10 was also found to be an unstable candidate gene in all but the geNorm analysis. These results correspond with the findings of Thellin et al. (1999), i.e., housekeeping genes should be evaluated as reference genes across the given experimental conditions in the given species. Based on our results, we recommend to avoid the use of these last four genes for normalisation in studies investigating gene expression patterns during the reproductive period in this species.
Conclusion
By evaluating the stability of eleven candidate housekeeping genes in samples collected during the breeding period of free-living Lethrus apterus, we conclude that two of them provide sufficient reference for normalising target gene expression. In head samples, these two genes appear to be L7A and RP18, whereas in thorax samples L7A and RP4 should be used. In both thorax and head samples of females, RP18 and L7A are the best choices for normalisation. Based on our results, in head samples of males, RPS8 and L7A, while in thorax samples of males, L7A and EF2 are recommended to use. These results provide reliable reference genes that are suitable normalizers for further RT-qPCR investigations on the hormonal regulation in Lethrus apterus.
Supplemental Information
Expression stability ( M) values of the eleven candidate reference genes in the different sample groups as calculated by geNorm
Lower M value indicates more stable expression. The most stable gene pair in each sample group is highlighted in bold.
Stability ranking of the candidate reference genes calculated by NormFinder
As NormFinder requires setting subgroups, the specified subgroups were as follows: head and thorax samples within all samples; male and female samples within head samples; male and female samples within thorax samples, respectively. Because of this requirement we do not have the same sample groups as analysed with the other procedures.
Mean absolute deviation (MAD) and Pearson correlation coefficient (r) of the candidate reference genes in the different samples groups as calculated by BestKeeper
Lower MAD value and higher r value means more stable expression. The most stably expressed genes are highlighted in bold.
Mean standard deviation (SDavg) calculated by delta Ct method
Lower SDavg value indicates more stable expression. The best candidate genes are highlighted in bold.