
Depicting the mating system and patterns of contemporary pollen flow in trees of the genus Anadenanthera (Fabaceae)
- Published
- Accepted
- Received
- Academic Editor
- Yu Liang
- Subject Areas
- Biodiversity, Genetics, Plant Science, Forestry, Population Biology
- Keywords
- Atlantic forest, Biparental inbreeding, Categorical paternity, Conservation genetics, Microsatellite markers, Gene flow
- Copyright
- © 2021 Feres et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2021. Depicting the mating system and patterns of contemporary pollen flow in trees of the genus Anadenanthera (Fabaceae) PeerJ 9:e10579 https://doi.org/10.7717/peerj.10579
Abstract
Anadenanthera (Fabaceae) is endemic to the Neotropics and consists of two tree species: A. colubrina (Vell.) Brenan and A. peregrina (L.) Speg. This study examined the mating system and contemporary gene flow of A. colubrina (Acol) and A. peregrina (Aper) in a highly fragmented area of the Atlantic Forest to provide valuable information that informs conservation strategies. Reproductive adults from forest remnants [nA. colubrina = 30 (2.7 ha), nA. peregrina = 55 (4.0 ha)] and progeny-arrays (nA. colubrina = 322, nA. peregrina = 300) were genotyped for seven nuclear microsatellite markers. Mating system analyses revealed that A. colubrina is a mixed mating species (tm = 0.619) while A. peregrina is a predominantly outcrossing species (tm = 0.905). For both Anadenanthera species, high indices of biparental inbreeding were observed (Acol = 0.159, Aper = 0.216), resulting in low effective pollination neighborhood sizes. Categorical paternity analysis revealed different scales of pollen dispersal distance: the majority of crossings occurring locally (i.e., between nearby trees within the same population), with moderate pollen dispersal coming from outside the forest fragments boundaries (Acolmp = 30%, Apermp = 35%). Nevertheless, pollen immigration from trees outside the populations for both species suggests that the populations are not reproductively isolated. This study highlights the importance of evaluating both mating system and contemporary gene flow for a better understanding of the biology of Anadenanthera species. This information should be considered to ensure the effective conservation and management practices of these plant species.
Introduction
Within in the semi-deciduous Brazilian Atlantic Forest, Ribeirão Preto is a region that has been significantly affected by land conversion for sugarcane agricultural practices. The municipality is currently among the largest producers of sugarcane in Brazil. Floristic survey data in this region indicate that Atlantic Forest remnants in Ribeirão Preto are distributed across 102 fragments (ranging from 1.5 to 247.0 ha), of which 54% are less than 10 ha (Kotchetkoff-Henriques, 2003). Among these fragments, 37 areas show a high density (more than 20 individuals/ha) of Anadenanthera, the target genus of this study. Plants of this genus often occur in monospecific clusters, known locally in Brazil as angicais. This population structure may be related to allelopathic effects that have been discussed in previous analyses of this genus (Silva et al., 2010). The regional landscape is marked by high concentrations of Anadenanthera scattered along highways and in municipal parks, especially in deciduous forest areas that are not suitable for agriculture, thus highlighting the importance of this genus in the region.
Occurring in Latin America and the Caribbean, Anadenanthera consists of two species and four varieties (Altschul, 1964). Anadenanthera colubrina (Vell.) Brenan includes two varieties: A. colubrina var. colubrina, which grows in northeastern Argentina and southeastern Brazil, and A. colubrina var. cebil (Griseb.) Altschul, which occurs in northern Argentina, Bolivia, Brazil, Paraguay, and Peru (Altschul, 1964). Anadenanthera peregrina (L.) Speg. consists of A. peregrina var. peregrina, which occurs in the northwest of Brazil, Guyana, Colombia, Venezuela, and the Antilles, and A. peregrina var. falcata (Benth.) Altschul which is found in southern Brazil and Paraguay. An in-depth botanic description of these species and their varieties, including detailed information on cultural and pharmacological uses can be found in Torres & Repke (2012).
Both species of the genus can be used in landscaping, and their excellent quality wood can be used in construction and shipbuilding (Lorenzi, 2002; Carvalho, 2003). The bark and the wood have tannin (used in traditional medicine) and a colorant that can be extracted from the bark (used in tanneries) (Lorenzi, 2002; Monteiro et al., 2006). Anadenanthera nuts contain alkaloids that were used by indigenous peoples in pre-colonial times, principally in Peru, Chile, Bolivia, and Argentina, and are thought to have potential medicinal properties (Weber et al., 2011; Torres & Repke, 2012). Anadenanthera species are considered honey plants as they provide bees with pollen and nectar (Souza et al., 2007). These species are widely recommended for the recovery of degraded and eroded areas, for riparian replacement in poor, shallow soils, and as a component of Agroforestry systems (Durigan & Silveira, 1999; Monteiro et al., 2006).
As the reduction of population size becomes more common due to ongoing loss and degradation of habitats in the Brazilian Atlantic Forest, it is essential to assess the mating parameters since they affect the genetic diversity of future generations and thus demonstrate a species’ responsiveness to anthropogenic factors. Over generations, mating parameters (e.g., effective dispersal distances of gene flow, and outcrossing rates) determine how genes are recombined and maintained in plant populations, providing the basis for the continuation of the evolutionary processes (e.g., Feres et al., 2009, 2012a; Barrett, 2010; Gonela et al., 2013; Guidugli et al., 2016; Souza et al., 2018; Cuénin et al., 2019; Garcia et al., 2019; Seoane et al., 2019). In highly fragmented landscapes, even low levels of gene dispersal are sufficient to counteract the long-term detrimental effects of inbreeding and loss of genetic diversity resulting from genetic drift, founder effects, and genetic erosion (e.g., Young, Boyle & Brown, 1996; Ellstrand, 2014). As an illustrative example, in the same fragmented landscape where Adenanthera species occurs, a gene flow study involving the allogamous species Carianiana estrellensis (Lecythidaceae) revealed a lack of inbreeding as a consequence of extensive gene immigration and high outcrossing rates (Guidugli et al., 2016). Therefore, to formulate biological diversity conservation policies and management programs, mainly in fragmented landscapes, an understanding of the mating system parameters (e.g., outcrossing, selfing and biparental inbreeding rates) in addition to estimates of contemporary gene flow in plant populations are indispensable (e.g., Allard, 1999; Finkeldey, 2005; Guidugli et al., 2016; Zhang et al., 2019; Lompo et al., 2020).
In this context, and considering the resilience of Anadenanthera species in this hyperfragmented region and their economic importance, there is considerable interest in increased knowledge of mating parameters in natural populations of A. colubrina and A. peregrina. We focused our efforts on characterizing the mating parameters for these insect-pollinated and autochorous tropical tree species. Our specific goals were to: (1) document the mating system estimates of A. colubrina and A. peregrina, (2) apply paternity analysis to quantify realized pollen dispersal distances, and (3) identify pollen immigration for A. colubrina and A. peregrina. The study of these genetic parameters in trees of the Anadenanthera genus in Atlantic Rainforest remnants will provide a useful starting point for conservation and management practices.
Materials and Methods
Study species
Anadenanthera belongs to Fabaceae, subfamily Caesalpinioideae (LPWG, 2017). Individuals [A. colubrina and A. peregrina] are heliophyte climax trees that can reach up to 35 m in height, and a diameter at breast height (DBH) up to 120 cm. The distribution of Anadenanthera appears to be largely natural, except for the probable anthropogenic introduction of A. peregrina into the West Indies (Altschul, 1964). The species of this small genus occur in savannas (i.e., Brazilian cerrado), and in wet and dry forests throughout tropical and temperate America (Altschul, 1964). These species tolerate sandy and shallow soils and light shading during the juvenile phase. A. colubrina (Figs. 1A–1D) has nitid, smooth to reticulated pods, and glandular anthers, while A. peregrina (Figs. 1E–1H) has dull, scurfy to verrucose pods, and eglandular anthers (Altschul, 1964). Anadenanthera colubrina var. colubrina and A. peregrina var. falcata—the focal taxa of this study—are hermaphrodites with actinomorphic flowers that are pollinated by bees and small insects. In São Paulo, Brazil, both species bloom from August to October (Fig. 1). The fruits, which are pods with 8–16 seeds, ripen from August to September of the following year (Fig. 1). The seeds of both species are brown, slightly flattened, orbicular, without a wing, and range from 10 to 20 mm in diameter (Altschul, 1964) and from 0.10 g to 0.17 g in weight. Due to autochorous seed dispersal, the dry seeds are dispersed primarily over short distances (Lorenzi, 2002). Despite their economic, environmental, and cultural importance, no study has yet examined the conservation of these valuable species.

Figure 1: Pictures of Anadenanthera colubrina and A. peregrina illustrating its vegetative and reproductive characteristics.
Highlight for the stem (A), pods (B), flowers (C), and suber (D) of A. colubrina. Details of the stem (E), pods (F), suber (G), and leaves and fruits (H) of A. peregrina are also shown.{kind=link}
Study site
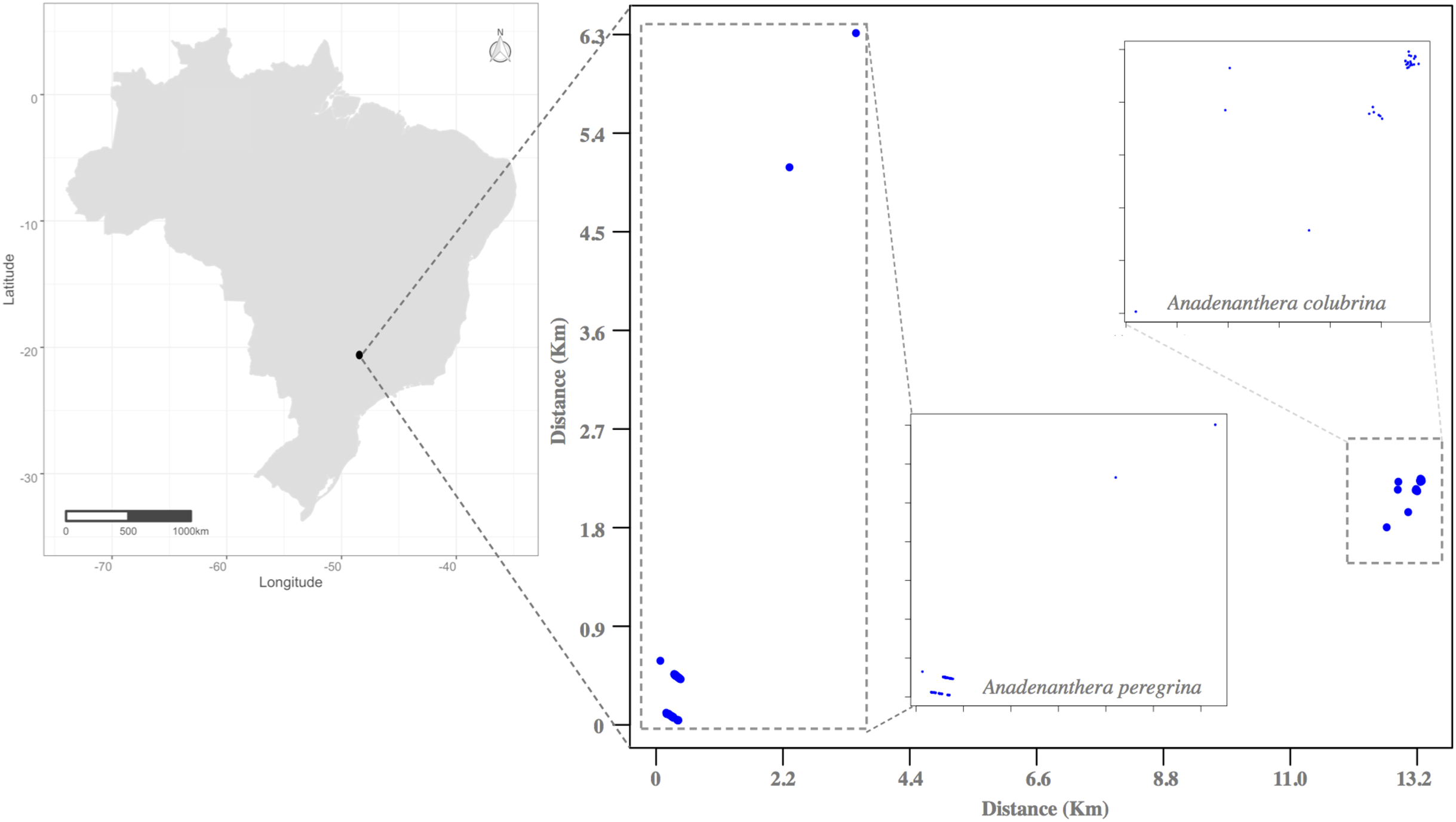
This study was conducted in Ribeirão Preto, State of São Paulo, Southeastern Brazil, during the drought seasons of 2011 and 2012 in small, semi-deciduous forest fragments (Fig. 2). The history of forest devastation of Ribeirão Preto region began with the increase of the area for coffee production in the late nineteenth century and the massive arrival of European immigrants after the abolition of slavery in Brazil in the early twentieth century. The degradation of the few forest remnants that persist in the region continues due to the extensive production of sugarcane and intense urban expansion (Kotchetkoff-Henriques, 2003; Victor et al., 2005; Guidugli et al., 2016). Today this Region is known as one of the largest sugarcane producers in Brazil. In this context, to perform the mating system and pollen flow studies, we selected and sampled all individuals from an isolated area for each Anadenanthera species: Acol (A. colubrina, 21°17′19.86″ S, 47°48′49.29″ O, 2.7 ha, Fig. 2) and Aper (A. peregrina, 21°23′51.98″ S, 47°51′49.23″ O, 4.0 ha, Fig. 2). The distances among Acol and Aper adult trees were ~2.5 m to 1,300 m. In both angicais, we observed little regeneration, as these clusters are located along roadsides and profoundly disturbed by anthropogenic activities of urban sprawl and agriculture. Small remnants occur within 100 and 1,300 m of the sampled angicais, and other fragments containing the species occur between 11.0 and 20.0 km away.
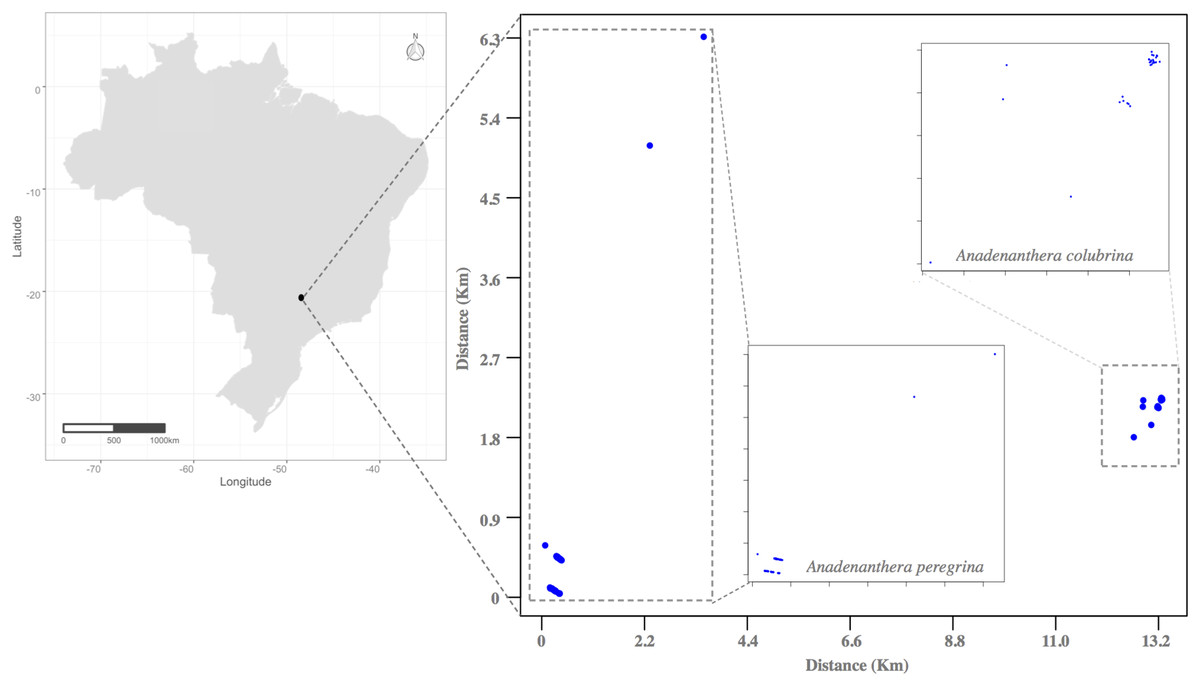
Figure 2: Location of the Atlantic Forest fragments studied in the municipality of Ribeirão Preto (São Paulo State, Southeastern Brazil), where individuals of the species Anadenanthera colubrina and A. peregrina were sampled.
{kind=link}
Sample collection and genetic analysis
Leaf material for Anadenanthera colubrina (Acol) and A. peregrina (Aper) was sampled from 30 and 55 adult trees, respectively, which were identified and mapped with a GPS device (Garmin, USA). The adult trees (DBH 1.4–3.9 m) were those that produced fruits during the sampling period. In addition, on average, 10 and 15 random fruits were collected from 10 adult seed-trees each of A. colubrina and A. peregrina population that flowered in 2011 and 2012, respectively. All fruits were separated, identified, and stored in paper bags (one fruit per bag) to maintain the provenance of fruit, seed, and seed-tree. The identification of seeds by fruit allowed us to determine the correlation of paternity between and within fruits. Seeds were planted in a nursery of forest seedlings of the Campus of the University of São Paulo, Ribeirão Preto. Leaf samples were obtained from 322 Acol progeny arrays (28, 33, and 30 open-pollinated progeny from one, each of eight, and one seed-trees, respectively). Likewise, we obtained leaf samples of 300 Aper progeny arrays (30 open-pollinated progeny from each seed-tree). Samples from all 85 adult trees and 622 progeny were conserved at −20 °C until DNA extraction. Research permit to collect the Anadenanthera species was approved by SisGen (number A380C57).
Genomic DNA extraction and microsatellite amplification conditions were described in Alzate-Marin et al. (2009) and Feres et al. (2012b), respectively. For A. colubrina, we scored the loci Acol02, Acol05, Acol15, Acol16, Acol17, Acol18, Acol20. A different set of primers (Acol09, Acol11, Acol12, Acol13, Acol14, Acol15, Acol20) was used for A. peregrina due to cross-amplification issues (Feres et al., 2012b). Electrophoretic conditions for these loci were described in Feres et al. (2012b).
Linkage disequilibrium, Hardy-Weinberg equilibrium, and null alleles tests
The linkage disequilibrium for all the analyzed loci was tested using FSTAT 2.9.3.2 program (Goudet, 2002) applying the Bonferroni correction (Rice, 1989). Tests for deviations from Hardy-Weinberg equilibrium for adult trees and progeny arrays were examined based on Monte Carlo permutations of alleles using the adegenet package (Jombart, 2008) implemented in R 3.3.1 (R Core Team, 2018). The presence and frequency of null alleles were estimated following the method of Brookfield (1996) using the PopGenReport package (Adamack & Gruber, 2014) in R.
Determination of mating system
Genotypes from the offspring (n = 322 for A. colubrina and n = 300 for A. peregrina) and seed-trees were evaluated using the mating system program MLTR version 3.2 (Ritland, 2008). This program took into account that all loci may contain null alleles even if there are none (Ritland, 2008). The parameters estimated for individual families (n = 10 families) and at the population level, included tm (multilocus outcrossing rate), ts (single-locus outcrossing rate), 1– tm (selfing rate), tm–ts (biparental inbreeding or mating among relatives), and rp(m) (multilocus paternity correlation). The average number of pollen donors Nep = 1/rp(m) (Ritland, 1989) and the inbreeding coefficient for seed-trees and offspring were also calculated. For all parameters, the standard error (SE) was calculated from 1,000 bootstrap replicates with resampling among families.
Categorical parentage analysis
Progeny arrays for both Anadenanthera species (Acol = 322, Aper = 300) were examined to estimate gene flow through pollen dispersal, considering all adult trees (n = 30 for A. colubrina and n = 55 for A. peregrina) as paternal candidates. Paternity analyses were performed according to the maximum likelihood method (Marshall et al., 1998) integrated in the CERVUS program v.3.0.3 (Kalinowski, Taper & Marshall, 2007). We carried out 50,000 simulated genotypes to achieve the critical value of Delta (Δcrit), considering 90% of putative fathers were sampled and a genotyping error ratio of 1%. This genotyping error was set up to minimize the effects of null alleles on paternity estimates (Dewoody, Nason & Hipkins, 2006; Kalinowski, Taper & Marshall, 2007). After Δcrit was calculated, the paternity assignment was performed to achieve the value of Delta (Δ; i.e., the difference between the highest and the second highest LOD scores). Paternity analyses for A. colubrina and A. peregrina were performed using all loci and confidence level of 95%. In the paternity analyses, only the potential father with Δ > Δcrit was considered to be the true parent of the analyzed progenies. For both Anadenathera species, pollen dispersal distances were calculated for progeny arrays considering the geographic coordinates of the seed-tree and the assigned pollen parent within the studied population. Furthermore, pollen immigration rate (mp) was estimated as the percentage of genotypes not assigned to a candidate parent within the population as proposed by Smouse & Sork (2004). We also calculated the cryptic pollen flow (Cgf; i.e., the value that expresses the proportion of genotypes assigned to a candidate father within the sampled area when the true father is located outside there) as 1 – (1 – P2)n, where n is the number of candidate fathers within the population, and P2 represents the combined non-exclusion probability of the second parent, when the mother parent is known (Dow & Ashley, 1996). P2 values were calculated from adult trees of A. colubrina and A. peregrina using the CERVUS program (Kalinowski, Taper & Marshall, 2007). Finally, considering a circular area around each seed-tree of A. colubrina and A. peregrina, we calculated the effective pollination neighborhood area (Aep = 2πσ2) from the variance of pollen dispersal distances (σ2) according to Levin (1988).
Results
Linkage disequilibrium, Hardy-Weinberg equilibrium, and null alleles frequency
For both A. colubrina and A. peregrina, the linkage disequilibrium was not significant between loci for adults and progeny arrays after a sequential Bonferroni correction for k tests (k = 21, p < 0.0024). In addition, no significant departures from HWE were observed after a Bonferroni adjustment (p > 0.007) for adult and progeny arrays of both species. PopGenReport revealed that locus Acol18 may contain null alleles at moderate frequencies (0.138) in adult trees of A. colubrina (Table S1). For A. peregrina, frequencies for locus Acol20 ranged from 0.074 (adult trees) to 0.126 (progeny arrays; Table S1). However, we chose to include Acol18 and Acol20 in this study and accounted for null alleles in our mating system and paternity analyses.
Mating system of Anadenanthera species
The average multilocus outcrossing rates estimated for Anadenanthera colubrina (tm = 0.619) and A. peregrina (tm = 0.905) were significantly different from unity, indicating that A. colubrina is a mixed mating species while A. peregrina is a predominantly outcrossing species with a likely absence of self-incompatibility mechanisms in both species (Table 1). Estimates of individual multilocus outcrossing rates showed variation between sampled seed-trees, with high and low observed values for the ten open-pollinated families analyzed, varying from 0.19 to 1.2 for A. colubrina and from 0.83 to 1.2 for A. peregrina (Table 1).
| Parameters | A. colubrina | A. peregrina |
|---|---|---|
| Number of seed-trees/progeny arrays | 10/322 | 10/300 |
| Multilocus outcrossing rate (tm) | 0.619 [0.57–0.66] | 0.905 (0.89–0.92] |
| Multilocus outcrossing rate by family | ||
| Family [01] | 1.200 [1.02–1.37] | 0.791 [0.77–0.82] |
| Family [02] | 0.866 [0.72–1.01] | 1.200 [1.19–1.21] |
| Family [03] | 0.677 [0.57–0.79] | 0.875 [0.84–0.91] |
| Family [04] | 0.296 [0.25–0.34] | 0.875 [0.84–0.91] |
| Family [05] | 0.655 [0.55–0.76] | 0.833 [0.80–0.86] |
| Family [06] | 0.713 [0.59–0.84] | 0.901 [0.87–0.93] |
| Family [07] | 0.628 [0.52–0.74] | 1.200 [1.19–1.21] |
| Family [08] | 0.624 [0.52–0.73] | 0.830 [0.80–0.86] |
| Family [09] | 0.477 [0.40–0.56] | 0.931 [0.90–0.96] |
| Family [10] | 0.196 [0.01–0.23] | 0.961 [0.92–1.00] |
| Single-locus outcrossing rate (ts) | 0.460 [0.42–0.50] | 0.690 [0.67–0.71] |
| Mating rate among relatives (tm - ts) | 0.159 [0.12–0.19] | 0.216 [0.20–0.24] |
| Selfing rate (1 – tm) | 0.381 [0.34–0.43] | 0.095 [0.08–0.11] |
| Multilocus paternity correlation (rp) | 0.465 [0.41–0.52] | 0.677 [0.62–0.74] |
| Multilocus paternity correlation within fruits | 0.905 [0.84–0.97] | 0.805 [0.73–0.88] |
| Multilocus paternity correlation among fruits | 0.383 [0.33–0.44] | 0.298 [0.24–0.36] |
| Average number of pollen donors: Nep | 2.15 | 1.48 |
| Average number of pollen donors within fruits | 1.10 | 1.24 |
| Average number of pollen donors among fruits | 2.61 | 3.35 |
| Fixation index in adult-trees | 0.202* | 0.091* |
| Fixation index in progeny arrays | 0.393* | 0.119* |
Notes:
( ) 95% confidence interval obtained by 1,000 bootstraps. Confidence intervals that fall within 1 (for tm and ts estimates) or 0 (for tm–ts estimate) are not significant.
We detected biparental inbreeding (Ritland, 2002) for both Anadenanthera species in the studied populations (tm − ts ranged from 0.159 for A. colubrina to 0.216 for A. peregrina), indicating that 15.9% and 21.6% of crosses were realized between related individuals. The fixation index observed in the adults (FA. colubrina = 0.202, FA. peregrina = 0.091) and progeny arrays (FA. colubrina = 0.393, FA. peregrina = 0.119) of both species were statistically different from zero, indicating inbreeding (Table 1).
The multilocus paternity correlation (rp(m)) was high and different from zero (0.465 for A. colubrina and 0.677 for A. peregrina). However, these values were even higher when we divided the analysis into within and among fruit for each seed-tree. Within fruit, the same parents were responsible for 90.5% (A. colubrina) and 80.5% (A. peregrina) of crossings. Among fruit, we observed 38.3% (A. colubrina) and 29.8% (A. peregrina) of crossings generated by the same trees. These results indicate that the progenies are composed of a mixture of half-sibs and full-sibs. Based on paternity correlation, it was estimated that 2.15 and 1.48 effective pollen donors mated with each seed-tree in both A. colubrina and A. peregrina, respectively. However, different fruits of the same seed-tree received pollen from approximately three individuals (2.6 for A. colubrina, 3.35 for A. peregrina), whereas a single fruit may have received pollen from only one tree (Table 1).
Paternity analysis for Anadenanthera species
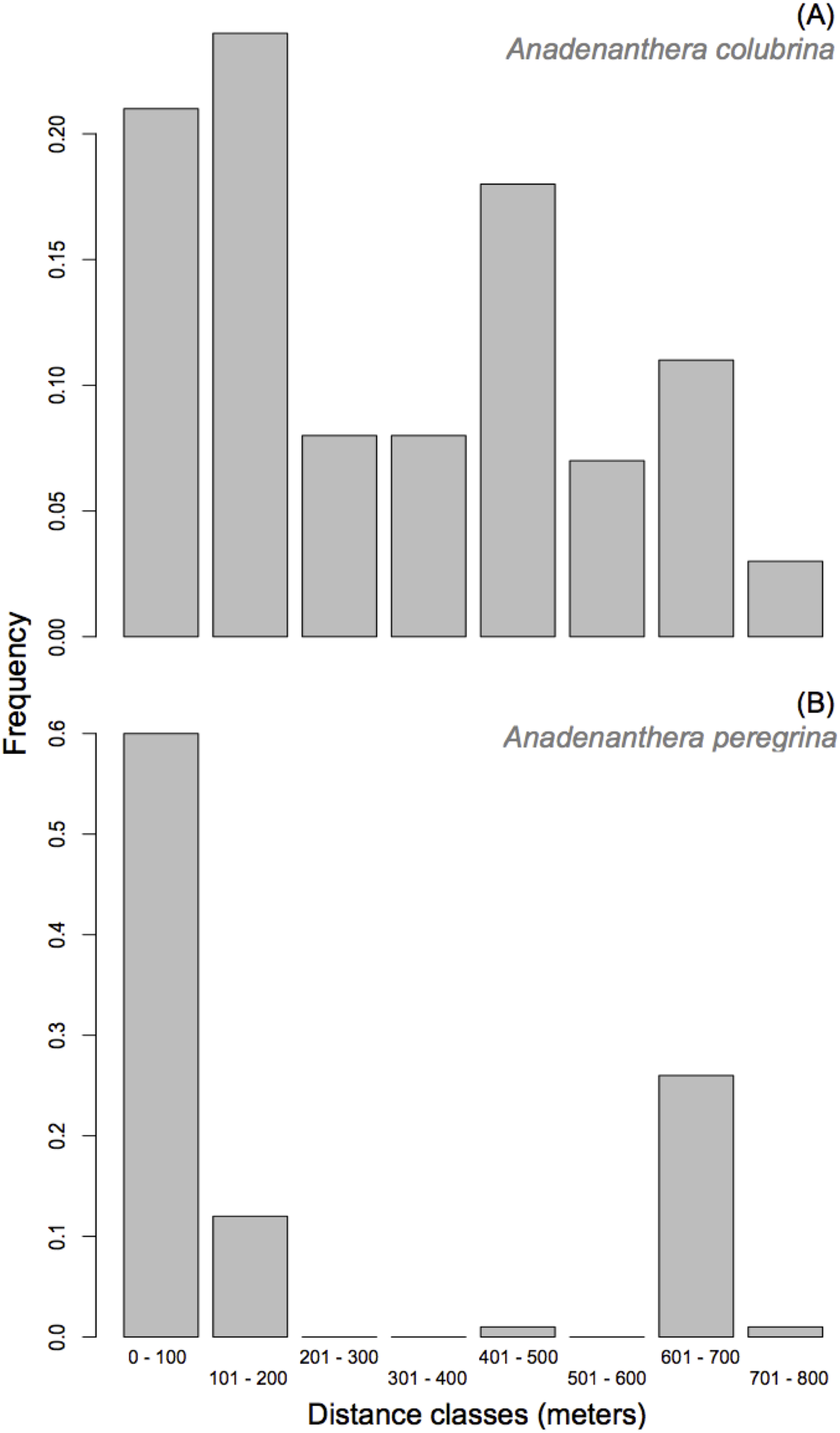
Anadenanthera colubrina—A paternal parent was assigned to 70% of the population (225 out of 322 progeny arrays) with a minimum 80% confidence. Consistent with the results for the mating system, 103 of the 225 assigned paternities were the result of self-fertilization. Paternity was designated for 81 of the 322 progeny arrays (25%) with 95% confidence. Further, 22 of the 30 adult trees contributed paternity to offspring from the ten seed-trees analyzed, though only five donated pollen to 52% of offspring. The percentage of progenies that do not have a parent tree assigned within the studied population (i.e., pollen immigration) was moderate (mp = 30.0%). Since the cryptic gene flow had a probability of 6.3%, the total pollen migration for progeny arrays was 36%, which indicates that this small A. colubrina population is not genetically isolated given the exclusion probability (P2 = 0.919; see Table S2) and sampling of all adult trees within the population. Effective pollen dispersal distances within the population were relatively short, with 53% of pollination occurrences at distances up to 300 m (Fig. 3). However, 39% of the effective pollination was observed over long distances (>500 m; Fig. 3). Overall, the pollen dispersal distance ranged from 7 to 769 m, with an average of 299.8 ± 8.4 m (SE). The effective pollination neighborhood (Aep) for A. colubrina ranged from 0.57 to 5.73 ha, with an average of 2.54 ha between seed-trees, corresponding to a circle around a seed-tree with a radius of 90 m.
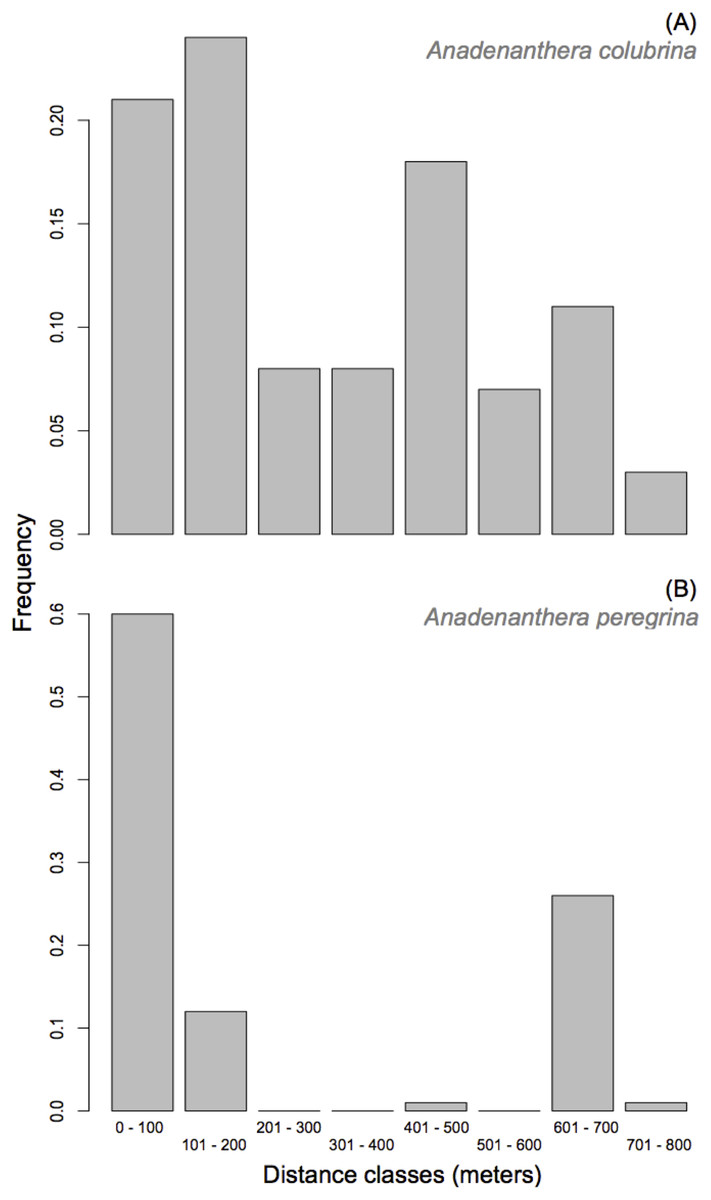
Figure 3: Frequency distribution of pollen dispersal distance in Anadenanthera colubrina (A) and A. peregrina (B).
{kind=link}
Anadenanthera peregrina—For this species, a paternal parent was assigned to 64% of the population (193 of 300 progeny arrays) with a minimum 80% confidence. Paternity was designated with 95% confidence for 60 of the 300 sampled progeny arrays (20%). Further, 45 of the 55 adult trees contributed pollen to offspring from the analyzed 10 seed-trees, and only 11 pollinated 50% of offspring. The percentage of pollen immigration was moderate (mp = 35.0%). Since the cryptic gene flow had a probability of 5.4%, the total pollen immigration for progeny arrays was 40.4%, indicating that the forest fragment is not genetically isolated, given the exclusion probability (P2 = 0.970; see Table S2) and sampling of all adult trees within the population, a result similar to A. colubrina. Although 27% of the effective pollination was observed over long distances (>700 m; Fig. 3), 72% of dispersal distances within the population were relatively short (i.e., up to 200 m; Fig. 3). The effective pollination neighborhood (Aep) ranged from 0.54 to 307.3 ha, with an average of 17.16 ha between seed-trees, corresponding to a circle around a seed-tree with a radius of 234 m.
Discussion
To our knowledge, this is the first study assessing the mating system and gene flow patterns of the economically and ecologically important leguminous trees A. colubrina and A. peregrina. The mating system was studied in two small natural clusters (angicais) of these species located in Ribeirão Preto, Brazil, an area in which the forest has been highly fragmented for at least 100 years. Both species of Anadenanthera presented a mixed-mating system, with selfing, mating between relatives, and a limited number of pollen donors. However, A. peregrina presents a higher outcrossing rate than A. colubrina. These estimates are comparable to those reported in field experiments in another region of Brazil (Mato Grosso State), where A. peregrina is identified as an allogamous species with a self-pollination rate of 9.48% (Costa, Contini & Melo, 2003). Protandry is also reported for both Anadenanthera species as one of the cross-pollination strategies for the species (Costa, Kageyama & Mariano, 1992; Costa, Contini & Melo, 2003). However, to confirm that mixed-mating is indeed a specific characteristic of these species, more populations in other regions and biomes in Brazil must be analyzed.
The estimates for reproductive parameters for both Anadenanthera species show a wide variety among the sampled seed-trees, which may indicate that the selfing rate fluctuates between reproductive events. Furthermore, our results suggest that the reproductive system of A. colubrina and A. peregrina may not be resilient to reductions in population size, since the results for biparental inbreeding indicate mating among relatives. Although we have not investigated the spatial genetic structure in these populations due to the small sample sizes mainly for A. colubrina, it can be inferred based on the occurrence of pollen flow over relatively short distances and the biparental inbreeding observed in the studied populations. These results are consistent with other tree species in fragmented populations that have shown elevated levels of mating between related individuals (Ismail et al., 2012; Tambarussi et al., 2017; Souza et al., 2018) and inbreeding (Fuchs, Lobo & Quesada, 2003; Jump & Peñuelas, 2006; Kettle et al., 2007; Dick et al., 2008; Vranckx et al., 2011; Zhang et al., 2012; Finger et al., 2014; Tambarussi et al., 2017; Seoane et al., 2019; but see Lompo et al., 2020). Such findings could be explained by the genetic structure of natural clusters (i.e., angicais) in the parental generation, with near-neighbor related trees resulting from seed dispersal close to the seed-tree (autochoric dispersion), profuse flowering, and pollinator behavior. In the present study, the pollination biology of Anadenathera species was not investigated; however, other authors have reported that inflorescence of Anadenanthera species are foraged intensively by native bees, including Trigona spinipes (Fabr.), Plebeia sp., Frieseomelitta sp., and the exotic bee Apis mellifera (Teixeira, Oliveira & Viana, 2007; Trovão et al., 2009). Therefore, the population structure of reproductive adult trees in the fragmented habitat and pollinator behavior are key aspects in the crossing rates observed herein and may play an essential role in connecting the few remaining forest fragments.
Gene flow by pollen dispersal outside the boundaries of forest fragments has been reported for several animal-pollinated plant species (Nason & Hamrick, 1997; Sato et al., 2006; Nazareno & Carvalho, 2009; Ottewell et al., 2012; Côrtes et al., 2013; Saro et al., 2014; Guidugli et al., 2016; Garcia et al., 2019; Skogen et al., 2019; Lompo et al., 2020). Even in an extremely fragmented landscape, such as the Ribeirão Preto region, we observed a moderate frequency (30–35%) of pollen immigration for both Anadenanthera species, showing that pollen flow outside the edges of small forest fragments is feasible. In this same region, Guidugli et al. (2016) also reported extensive pollen immigration (i.e., up to 53%) for the allogamous tree species Cariniana estrellensis in a forest fragment size of 8.0 ha. The main point raised by these two studies, and with potential relevance for conservation strategies in Ribeirão Preto region, is that populations of tree species in small forest fragments are not genetically isolated.
In practical terms, our study presents basic information that should be considered for in situ strategies, and when collecting seeds for breeding, conservation, and restoration programs. For seed collection, it should be considered that all seeds in a fruit of A. colubrina and A. peregrina are a mixture of half- and full-sibs, and that seed-trees of both species were pollinated by multiple pollen donors. Hence, we recommend collecting one seed from many separate fruits through the whole crown of each seed-tree. However, further fine-scale spatial genetic structure studies should be documented for these populations to avoid collecting seeds from genetically related seed-trees. With regards to gene flow, although the estimates are biased due to the low power of exclusion of the seven loci used herein, we can conclude that the Anadenanthera populations received a moderate percentage of immigrant pollen. Although pollen dispersal occurred predominantly over relatively short distances in the A. colubrina and A. peregrina populations, pollen flow over long distances was also observed (Fig. 3). These dispersal patterns are relevant for the in situ conservation of remaining local populations since gene flow over long distances can enhance and/or increase connectivity between the remaining fragments of Anadenanthera species. Indeed, this suggests that angicais could play the role of ecological stepping stones while also enabling the viability of local pollinators. Several empirical studies have shown the importance of connectivity for processes of recolonization after local extinction, as well as in the maintenance of metapopulations across fragmented landscapes (e.g., Hanski, 1994; Metzger, 2000; Antongiovanni & Metzger, 2005; Rubio & Saura, 2012; Laurance et al., 2018), as appears to be the case with angicais in the Ribeirão Preto region.
The concept of connectivity is essential, since species survive in fragmented habitats (or are dispersed heterogeneously throughout the landscape) and depend on their natural capacity for gene flow across landscape matrices. Generally, the matrix acts as a barrier to the movement of seed and pollen dispersal, restricting local populations (e.g., Metzger, 2000; Dick, Etchelecu & Austerlitz, 2003; Antongiovanni & Metzger, 2005; Uezu, Metzger & Vielliard, 2005; Ayram et al., 2016). The intensity of gene flow between habitats is regulated by the permeability of the matrix and the inherent ability of each species for pollen and seed flow across these environments (e.g., Metzger, 2000; Uezu, Beyer & Metzger, 2008). Therefore, considering the importance of connectivity between fragments to maintain forest remnant viability and biodiversity, efforts have been made to reduce habitat isolation. Several strategies have proven to increase connectivity between fragments, notably the establishment of corridors along riparian forests (e.g., Rosot et al., 2018), increasing the porosity of the matrix (e.g., Viana & Pinheiro, 1998; Rodrigues et al., 2009; Rubio & Saura, 2012). Such measures must be based on empirical data, especially related to the flow capacity of natural species particular to each environment (e.g., McRae et al., 2012; Torrubia et al., 2014). As highlighted by Ribeiro et al. (2009), small remaining forest patches (less than 50 ha) play an important role in connectivity in the fragmented landscape, reducing the average isolation among forest populations from 3,532 m to 1,441 m.
Conclusions
Based on the study of gene flow and the mating system of Anadenanthera species in Ribeirão Preto region, we suggest that angicais play an important role connecting the few remaining local fragments. Thus, strategies for the conservation and maintenance of diversity for angicais and other species should be based on the information reported herein. Nevertheless, further studies assessing mating parameters on a spatial-temporal scale are necessary to strengthen our understanding of the mating systems and gene dispersal patterns of these tropical plant species.
Supplemental Information
Microsatellite data for Anadenanthera colubrina and Anadenanthera peregrina.
Each two columns represents a specific locus. Individuals (adults and progenies) are listed in each row.