
Abstract
In recent years, studies have revealed an interaction between two systems of attention—alerting and executive control. Specifically, warning cues increase the influence of cognitive conflict under certain conditions. One of the problems of interpreting this effect is that warning cues can trigger two processes simultaneously—a high arousal state and strategic temporal expectancy. The goal of the present study was to clarify which process underlies the increased congruency effects following a warning event. In two experiments, the influence of warning cues on flanker congruency was examined while controlling for the effects of temporal expectancy and arousal. Experiment 1 revealed a strong effect of warning cues on congruency, even when the warning cues were not temporally predictive. This effect was evident at two short cue-to-target intervals of 100 and 500 ms, but not following a 900-ms interval. Experiment 2 revealed that this effect was not altered even when the warning cues predicted with high certainty that the target would appear at long cue-to-target intervals (e.g., 900 ms). We suggest that the interaction between alerting and executive control reflects the involvement of a subcortical mechanism responsible for increasing arousal.
Similar content being viewed by others

In recent years, interest has grown in how attentional networks interact in order to achieve adaptive behavior. In the present study, we explored the underlying mechanisms of an interaction between a system that is in charge of producing a high level of alertness during performance (i.e., alerting) and a higher system that is involved in complex mental operations such as conflict resolution (i.e., executive control).
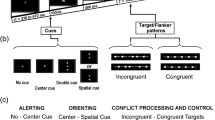
Alerting is often studied by introducing a brief warning signal just before the appearance of an imperative target. The common effect of these warning cues is reduced reaction times (RTs) in trials that include warning cues, as compared with no-cue trials (i.e., the alerting effect). This effect has been tightly linked to arousal and the distribution of norepinephrine (Coull, Nobre, & Frith, 2001; Petersen & Posner, 2012; Witte & Marrocco, 1997). Executive control is commonly tested in tasks that involve responses to a target stimulus while trying to suppress irrelevant conflicting information or response tendencies. Operation of this system has been widely associated with the frontal cortex (e.g., Botvinick, Braver, Barch, Carter, & Cohen, 2001). Elements of alerting and executive control can be tested simultaneously by including warning cues in a task used to measure executive control. This was done as part of a comprehensive test of attention named the “Attention Network Test” (ANT; Fan, McCandliss, Sommer, Raz, & Posner, 2002).
The ANT includes the arrow-flanker task to measure executive control. In this task, participants are presented with a line of five arrows. They are asked to indicate the direction in which the middle arrow points while attempting to ignore irrelevant arrows in close proximity. In the congruent condition, all of the arrows point in the same direction (← ← ← ← ←). In the incongruent condition, the middle arrow points in a different direction than that of the other arrows (← ← → ← ←). Since it is difficult to ignore the irrelevant arrows, RTs are slower in the incongruent than in the congruent condition. The difference in mean RTs between the incongruent and congruent conditions is named the congruency effect, which is used as a measure of the executive network.
One of the conditions in the ANT includes a brief warning cue that precedes a flanker target. Numerous studies that have used the ANT have reported that the flanker congruency effect is larger (i.e., indicating greater conflict) in trials that include warning cues (auditory or visual) than in no-cue trials (e.g., Callejas, Lupiáñez, Funes, & Tudela, 2005; Chica et al., 2012; Costa, Hernández, & Sebastián-Gallés, 2008; Dye, Baril, & Bavelier, 2007; Dye, Green, & Bavelier, 2009; Fan et al., 2009; Ishigami & Klein, 2010; MacLeod et al., 2010; McConnell & Shore, 2011; Redick & Engle, 2006; Weinbach & Henik, 2012a). Specifically, it has been shown that the warning cues induced a larger flanker interference effect, whereas the facilitation effect was not modulated (see Exp. 1 in Weinbach & Henik, 2012a). Why do irrelevant flankers have a greater impact following a warning event? In order to better understand this effect, it is crucial to identify and distinguish between the mental processes involved in this interaction.
One of the problems of interpreting this effect emerges when considering that warning cues often elicit two processes simultaneously, one of which is attributed to an immediate increase in arousal, and the other to increased temporal expectancy toward the target. Temporal expectancy relates to the ability to direct attention to particular moments in time following a warning cue (Coull & Nobre, 1998). Studies that have examined the influence of warning cues on performance have often failed to dissociate between these two processes. This resulted in reports of similar effects that were sometimes attributed to increased arousal and sometimes to temporal expectancy (for examples, see Weinbach & Henik, 2012b). Of current interest to us is the effect of warning cues on cognitive conflict. Although some studies have explored the hypothesis that abrupt arousal following a warning cue is responsible for increasing congruency (e.g., Callejas et al., 2005; Weinbach & Henik, 2012a), others have demonstrated that flanker congruency is larger when the temporal prediction of the cue matches the temporal onset of the target, as compared with a condition in which it does not (Correa, Cappucci, Nobre, & Lupiáñez, 2010).
The aim of the present study was to dissociate the effects of arousal and temporal expectancy on congruency in order to expose the mental process that underlies the greater effect of congruency following a warning cue. To accomplish this, it is necessary to control for temporal expectancy and arousal when examining the effects of warning cues on congruency.
In two experiments, we tested the influences of temporal expectancy and arousal on congruency using the flanker task. In Experiment 1, we attempted to neutralize the effects of temporal expectancy by reducing the temporal information provided by the warning cues. This allowed for examining the pure effect of arousal on congruency. In Experiment 2, the goal was to examine the influence of high temporal expectancy on congruency when arousal was low.
Experiment 1
One of the ways to study the effect of arousal, independent of temporal information, is to increase temporal uncertainty following a warning cue. When a target can appear at various time intervals following a warning cue (i.e., foreperiods), it is more difficult to temporally predict exactly when the target will appear. However, it is not impossible. When using several foreperiods within a block of trials, temporal expectancy develops as time is prolonged, a phenomenon called the “foreperiod effect” (for a review, see Niemi & Näätänen, 1981). Take, for example, an experiment with 90 trials and three possible foreperiods. If 30 trials are presented with each foreperiod, the probability for the target to appear following the first foreperiod is 1/3 (30 out of 90 possible trials). If the target does not appear within the first foreperiod, the probability for it to appear in the second foreperiod is already 1/2 (i.e., 30 out of the 60 possible remaining trials). If it does not appear within the second foreperiod, it is bound to appear within the last foreperiod (i.e., 30 out of the remaining 30 trials), and temporal expectancy reaches its maximum. Because the probability for the target appearance increases as time progresses, temporal expectancy following the cue gradually increases as well, causing RTs to decrease with time (i.e., the foreperiod effect). Some results have indicated that the foreperiod effect reflects flexible top-down anticipation in time and is associated with the frontal cortex (Vallesi, 2010; Vallesi, Shallice, & Walsh, 2007).
One of the techniques used to eliminate temporal expectancy following a warning cue is by using a nonaging foreperiod distribution (Niemi & Näätänen, 1981). This means keeping a constant 50 % probability that the target will appear in any given foreperiod. This makes it impossible to predict when the target will appear following a warning cue. Creating a nonaging foreperiod distribution is done by changing the amount of trials in each foreperiod and adding catch trials (i.e., trials with no target; Table 1 shows how the temporal predictability was calculated as a function of the number of trials in each foreperiod). By using nonaging foreperiods, it is possible to examine the pure effect of arousal following a warning cue, because the warning cue does not reduce uncertainty regarding the temporal onset of the target (Fernandez-Duque & Posner, 1997).
In the first experiment, we examined the influence of a brief warning event that elevates arousal on congruency in the flanker task while using a nonaging foreperiod distribution. We predicted that if the increased congruency effect following warning cues is a result of temporal expectancy, no interaction should occur between warning and flanker congruency. However, if this interaction is due to arousal rather than to top-down temporal preparation, the interaction should still be apparent. In this case, the interaction should be more prominent in earlier foreperiods of 500 ms and shorter, because these time intervals are mainly associated with phasic arousal rather than strategic mental operations that have little time to develop (Hackley, 2009; Hackley et al., 2009).
Method
Participants
A group of 27 undergraduate students (all females, ages ranging from 21 to 26 years) from the Department of Psychology at Ben-Gurion University of the Negev took part in this experiment for course credit or for payment of 30 NIS. All participants reported normal or corrected-to-normal vision, and all signed informed consent prior to their inclusion in the study.
Apparatus
Data collection and stimuli presentation were controlled by a DELL OptiPlex 760 v Pro computer with an Intel Core 2 duo processor E8400, 3 GHz. The stimuli were presented on a Dell E198PF 19-in. LCD monitor. E-Prime software (Psychology Software Tools, Pittsburgh, PA) was used for programming, presentation of the stimuli, and timing operations. Responses were collected through the computer keyboard, and a headphone set was used to deliver an auditory warning cue.
Stimuli
The target stimuli were five black arrows on a gray background, presented at the center of the screen. Each arrow subtended a visual angle of 0.8º from a viewing distance of 57 cm. The arrows were separated by a distance of 0.1º. In the congruent trials, the central target arrow was flanked by arrows pointing in the same direction, and in the incongruent trials, the central arrow was flanked by arrows that pointed in the opposite direction. Participants responded using a keyboard; they pressed the “z” key for a left response, and the “m” key for a right response. On half of the trials, a 2,000-Hz, 50-ms “beep” sound was delivered via headphones.
Procedure and design
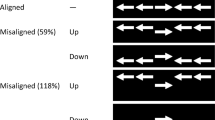
Prior to the beginning of the experiment, participants were instructed to respond according to the direction of a central arrow and to ignore flanking arrows. Each trial began with the appearance of a fixation plus sign that subtended a visual angle of 0.6º (Fig. 1 illustrates a typical trial). On half of the trials, following 2,000 ms, a warning cue (“beep” sound) was presented for 50 ms. Participants were told that they should not respond in any particular way to the auditory tone. Foreperiods of three lengths were possible: 100, 500, and 900 ms. The numbers of trials with the different foreperiods were distributed in such a way as to create nonaging foreperiods and a constant 50 % chance that the target would appear within any given foreperiod. There were 256 trials in the 100-ms foreperiod, 128 trials in the 500-ms foreperiod, and 64 trials in the 900-ms foreperiod. In addition, 64 catch trials were included, in which the target did not appear (see the calculations of probabilities in the “Nonaging” column of Table 1). In the no-cue trials, the time intervals were matched as if there was a cue. In target trials, the target remained in view until the participant’s response or until 2,000 ms had elapsed. After response, a blank screen appeared for 1,000 ms. In total, four experimental blocks were presented, each consisting of 128 trials (for a total of 512 trials). The trials in each block were presented in a random order, and each session began with 12 practice trials.

Example of a typical trial. In this trial, an auditory warning cue was presented prior to an incongruent target
Results
Trials in which an erroneous response was made were excluded from the RT analysis (1.7 %). Extreme RTs—less than 200 ms or greater than 1,000 ms—were also excluded from the analysis (2.4 %). None of the participants responded to the catch trials. One participant was excluded from the analysis for having extreme erroneous responses (more than four standard deviations from the mean). Repeated measures analyses of variance (ANOVAs) included Foreperiod (100, 500, or 900 ms), Warning (warning or no warning), and Flanker Congruency (congruent or incongruent) as factors.
The mean RTs for each condition are presented in Table 2. The analysis revealed a significant main effect of warning, F(1, 25 = 44.89, MSE = 1,424, p < .00001, η2 p = .64, as RTs in trials that included a warning cue were generally faster than those in trials in which the cue was absent. We found a main effect of congruency as well, F(1, 25 = 304.45, MSE = 1,027, p < 00001, η2 p = .92: RTs were slower in the incongruent than in the congruent condition. The main effect of foreperiods was marginally significant, F(2, 50) = 3.12, MSE = 582, p = .052, η2 p = .11. This is because the warning cue induced faster RTs in the 500-ms foreperiod than in the 100-ms and 900-ms foreperiods, F(1, 25) = 6.06, MSE = 433, p < .05, η2 p = .19. It is important to note that no significant difference in RTs emerged between the 100-ms and 900-ms foreperiods in the warning condition, F(1, 25) = 1.23, MSE = 678, p = .27,η2 p = .04. Moreover, the effect of foreperiod in the no-warning condition was not significant, F(2, 50) = 2.37, MSE = 933, p = .10, η2 p = .08.
The three-way interaction between foreperiod, warning, and congruency was not significant, F(2, 50) = 2.12, MSE = 273, p = .13, η2 p = .07, perhaps indicating that the Warning × Congruency interactions were fairly similar between the foreperiods. However, due to the theoretical importance of this effect, planned comparisons were carried out to examine the Warning × Congruency interaction in each of the foreperiods. In the first foreperiod (100 ms), the Warning × Congruency interaction was highly significant, F(1, 25) = 39.39, MSE = 153, p < .00001, η2 p = .61, as was the case in the second, 500-ms foreperiod, F(1, 25) = 26.14, MSE = 272, p < ;.0001, η2 p = .51. In both of the earlier foreperiods, the congruency effect was larger in the warning condition than in the no-warning condition (see Fig. 2). In the last, 900-ms foreperiod, the Warning × Congruency interaction showed the same trend but was not significant, F(1, 25) = 2.87, MSE = 551, p = .10, η2 p = .10.

Mean congruency effects as a function of aging distribution and warning condition. The y-axes represent the congruency effect (in milliseconds) (incongruent RT minus congruent RT), and the x-axes show the foreperiod distribution (in milliseconds). The error bars represent standard errors
Discussion
The results of Experiment 1 demonstrated that the interaction between alerting and executive control is evident even when the warning cue does not provide temporal information. The nonaging foreperiod distribution that was used to reduce temporal expectancy eliminated the common foreperiod effect that results from the gradual increase in temporal expectancy as time is prolonged.
The results support the hypothesis that the Warning × Congruency interaction is not a result of top-down temporal expectancy. The warning cues increased the congruency effect in short foreperiods of 100 ms and 500 ms, but not in the longer, 900-ms foreperiod. This pattern seems to be in line with the proposal that increased arousal underlies the Warning × Congruency interaction. When the time interval between the warning event and the target is 500 ms or shorter, the effects are usually associated with phasic arousal rather than with other, strategic mental operations that may develop later on (Hackley, 2009). Note that the warning cue had the largest effect on the general reduction of RTs in the 500-ms foreperiod. This is in line with previous findings demonstrating that alertness reaches the optimal level at this time interval (Posner & Boies, 1971). However, the difference in the sizes of the interaction between the 100-ms and 500-ms foreperiods was minimal. Furthermore, in the 900-ms foreperiod an influence of the warning cue on the RT reduction as still apparent, but it did not modulate congruency. This pattern may indicate that the Warning × Congruency interaction and the general reduction of RTs following a warning cue do not follow the same time course. The general reduction of RTs following a warning cue has often been associated both with early attentional/perceptual processing stages and with later, response-related processing stages (e.g., Böckler, Alpay, & Stürmer, 2011; Boulinguez, Ballanger, Granjon, & Benraiss, 2009; Hackley, 2009). Some evidence has suggested that the Warning × Congruency interaction might be more sensitive to early processing stages, such as perceptual processing and attention allocation (Weinbach & Henik, 2011, 2012a; see also the elaboration in the General Discussion section). These processes may no longer exert an influence 900 ms after the presentation of the cue. However, the influence of the warning cue on response-related processes (e.g., reduction of the response selection threshold; Hackley, 2009) may last longer and may still have an effect over long foreperiods (e.g., reduction of RTs). This suggestion is supported by a previous study that showed a strong influence of alerting cues on the orienting of attention at short intervals of 100–500 ms, but not following 800 ms, despite a general reduction of RTs at the longer time intervals (Fuentes & Campoy, 2008).
Overall, it seems that the results of Experiment 1 offer a plausible indication that increased arousal following a warning cue induces greater congruency, independent of temporal expectancy. However, some limitations in the nonaging foreperiod manipulation should be addressed. In order to create a nonaging foreperiod and equal 50 % probabilities for the target to appear in each of the foreperiods, significantly more trials must have targets within the first foreperiod. This results in many repetitions of the target in the first foreperiod within each block. It is possible that within a block, the warning cue could prime the first foreperiod, allowing participants to learn an association between the warning cue and target appearance. An alternative interpretation of the results of Experiment 1 would be that although the overall expectancies were equal among the foreperiods, within a block a stronger temporal contingency occurred between the warning cue and the 100- to 500-ms foreperiods, as compared with the 900-ms foreperiod, which had the weakest contingency (see also Lawrence & Klein, 2013).
Accordingly, the goal of Experiment 2 was to examine whether the results of Experiment 1 (i.e., larger congruency effect following a warning cue in earlier foreperiods) could be replicated even when there was a strong contingency between the warning cue and the 900-ms foreperiod, and a weak contingency between the warning cue and the 100-ms foreperiod.
Experiment 2
In Experiment 2, we deliberately increased participants’ temporal expectancy as time prolonged. This was done by simply switching the proportions of trials with the first and last foreperiods, creating a foreperiod distribution called accelerated aging (Baumeister & Joubert, 1969). The total number of trials remained the same as in Experiment 1, but now instead of having equal, 50 % chances for the target to appear in each of the foreperiods, the probability for the target to appear in the first, 100-ms foreperiod was 12.5 % (64 out of the total 512 trials; see also the calculation of probabilities in the “Accelerated Aging” column of Table 1); the probability for target appearance in the second, 500-ms foreperiod was 28.6 % (128 out of the remaining 448 possible trials); and the probability for appearance in the final, 900-ms foreperiod was 80 % (256 trials out of the remaining 320 trials). In this way, expectancy dramatically elevated as foreperiods increased, creating minimum expectancy for the target to appear within the early foreperiods and maximum expectancy for it to appear within the last foreperiod.
If the Warning × Congruency interaction is induced by arousal rather than by temporal expectancy, it should be indifferent to this expectancy manipulation, meaning that the Warning × Congruency interaction should still be most prominent in the early foreperiods. On the other hand, if the interaction is a result of the temporal expectancy induced by the warning cue, the Warning × Congruency interaction should be weakest in the early foreperiods, in which temporal expectancy is minimal.
Method
Participants
A group of 27 undergraduate students (18 females, nine males, ages ranging from 21 to 30 years) from the Department of Psychology at Ben-Gurion University of the Negev took part in this experiment for course credit or for a payment of 30 NIS. All of the participants reported normal or corrected-to-normal vision, and all signed informed consent prior to their inclusion in the study.
Stimuli and apparatus
The stimuli and apparatus were the same as in Experiment 1.
Procedure and design
The procedure was exactly the same as in Experiment 1, except that the proportions of target trials were reversed between the first, 100-ms foreperiod and the final, 900-ms foreperiod. This resulted in 64 trials in the 100-ms foreperiod, 128 trials in the 500-ms foreperiod, 256 trials in the 900-ms foreperiod, and 64 catch trials. Hence, the probabilities of the target appearing in each of the foreperiods were 12.5 %, 28.6 %, and 80 %, respectively.
Results
The erroneous response rate was low (1.8 %), and therefore trials with errors were not analyzed. RTs less than 200 ms or greater than 1,000 ms were also excluded (1.3 %). An ANOVA with Foreperiod (100, 500, or 900 ms), Warning (warning or no warning), and Flanker Congruency (congruent or incongruent) as factors was carried out.
The mean RTs for each condition are presented in Table 3. The analysis revealed significant main effects of warning and congruency—F(1, 26) = 25.13, MSE = 1,738, p < .0001, η2 p = .49, and F(1, 26) = 108.86, MSE = 2,769, p < .00001, η2 p = .80, respectively—with the same pattern as in Experiment 1. In contrast to Experiment 1, we found a main effect of foreperiod, F(2, 52) = 7.97, MSE = 753, p < .001, η2 p = .23. As in Experiment 1, following the warning cue, RTs were faster in the 500-ms foreperiod than in the 100-ms and 900-ms foreperiods, F(1, 26) = 7.7, MSE = 292, p < .05, η2 p = .22. Importantly, in the warning condition, RTs were also faster in the 900-ms foreperiod than in the 100-ms foreperiod, F(2, 52) = 4.12, MSE = 595, p < .052, η2 p = .13.
In addition, we discovered a three-way interaction between foreperiod, warning, and congruency, F(1, 26) = 5.18, MSE = 321, p < .01, η2 p = .16. As was the case in Experiment 1, the Warning × Congruency interaction was highly significant in the first, 100-ms and second, 500-ms foreperiods—F(1, 26) = 21.54, MSE = 445, p < .0001, η2 p = .45, and F(1, 26) = 7.71, MSE = 541, p < .05, η2 p = .22, respectively—but not in the last, 900-ms foreperiod, F(1, 26) = 2.42, MSE = 117, p = .13, η2 p = .08. This was due to a larger congruency effect following warning cues than in no-warning condition for the 100-ms and 500-ms foreperiods, but not for the 900-ms foreperiod (see Fig. 2). An unexpected finding was that in the no-warning condition, the congruency effect was larger in the 900-ms foreperiod than in the 100-ms and 500-ms foreperiods, F(1, 26) = 13.88, MSE = 239, p < .001, η2 p = .34.
Discussion
In Experiment 2, we tested whether high arousal following a warning cue would increase the congruency effect in early foreperiods, even when temporal expectancy was minimal. The results revealed that the Warning × Congruency interaction was significant in the 100-ms and 500-ms foreperiods, but was no longer in the 900-ms foreperiod, although temporal expectancy was highest in this condition. This pattern of results is yet another indication that brief arousing stimuli can act to increase the congruency effect, especially at short foreperiods of 500 ms and less.
An unexpected finding that should be addressed was that in the no-warning condition, which was not influenced by brief arousal, as no arousing cue was presented, we found an increase in the congruency effect in the 900-ms foreperiod as compared with the 100-ms and 500-ms foreperiods. Note that in Experiment 1, in which the expectancies were equal in all foreperiods, no such difference emerged in this condition. This result could indicate that temporal expectancy might have an independent influence on congruency, irrespective of arousal. This is in line with findings by Correa et al. (2010) that showed an increase in the congruency effect following highly predictive symbolic temporal cues. According to these authors, when the target appearance corresponds with the temporal prediction, response preparation is enhanced. This might result in an increased level of activation of the responses associated with both target and flankers and create greater conflict when these responses compete.
General discussion
The major goal of the present study was to understand and distinguish between the mental processes that are elicited by warning cues and that cause greater congruency effects. This follows reports concerning an interaction between alerting and executive control that could be explained as a modulation of higher cognitive functions (i.e., conflict resolution) by an increase in arousal or by temporal expectancy. We demonstrated that when warning cues did not temporally predict the onset time of a target, they still induced an increased flanker congruency effect (Exp. 1). Moreover, this effect seemed to be most prominent at short cue-to-target intervals (i.e., foreperiods of 100 and 500 ms), during which arousal was high and temporal expectancy probably had little time to develop. In Experiment 2, we demonstrated that even when the warning cue temporally predicted target appearance following the longest foreperiod (i.e., 900 ms), the Warning × Congruency interaction was still most prominent at early foreperiods and was not significant at a late foreperiod of 900 ms. In addition, the results revealed that temporal expectancy might have an independent impact on congruency. In the no-warning condition, congruency was greater in the 900-ms foreperiod, during which temporal expectancy was high, than in the 100-ms foreperiod.
The results of the present study confirm the hypothesis that increased immediate arousal following warning cues can generally reduce RTs but also elicits an increased congruency effect. This is especially true when using a short foreperiod in which arousal is high and temporal expectancy is low. In longer foreperiods the pattern is more complex, as congruency can also be influenced by temporal expectancy. This is in line with the idea that arousal and temporal expectancy are dissociable, but their effects are often confounded in many experimental designs (Weinbach & Henik, 2012b). The present study emphasizes the importance of controlling one of these processes when investigating the other.
Previous studies had attempted to discover why warning cues induce greater congruency effects in some conflict tasks (Böckler et al., 2011; Callejas et al., 2005; Fischer, Plessow, & Kiesel, 2010, Fischer et al., 2012; Weinbach & Henik, 2011, 2012a). However, these studies did not provide a conclusive answer regarding what critical process was triggered by the warning cue to cause an increased congruency effect. This is especially important when considering that warning cues can trigger several different processes simultaneously (Weinbach & Henik, 2012b). The present work fills in this gap and shows that the Warning × Congruency interaction at early SOAs results from elevated arousal. The interaction between arousal and executive control can teach us about the path by which lower subcortical mechanisms can modulate the higher cognitive functions associated with the frontal cortex.
Some authors have interpreted the larger flanker congruency effect following warning cues as a direct negative influence or inhibition exerted by arousal on executive control (Callejas et al., 2005; Fan et al., 2009). Recent studies have shown that such a direct impact is not probable, because warning cues do not modulate conflict adaptation effects or electrophysiological measures that are associated with executive control (Böckler et al., 2011). In addition, the Warning × Congruency interaction does not appear in classic executive control tasks such as the Stroop task (Weinbach & Henik, 2012a). These studies suggested that the influence of warning cues on executive control is mediated by other attentional processes. Fischer and colleagues (2010, 2012) suggested that warning cues can modulate executive control by facilitating the link between a stimulus and a response (S–R). When two conflicting responses are presented, the facilitation of the S–R link will create a greater cost in performance following warning cues. However, we have shown that this explanation cannot be exclusive, because warning cues did not modulate the S–R conflict when the relevant and irrelevant dimensions of the target were integrated into one object (Exp. 3 in Weinbach & Henik, 2012a). The Warning × Congruency interaction was apparent only when the relevant and irrelevant information were spatially separated (Exp. 4 in Weinbach & Henik, 2012a). Therefore, it was suggested that warning cues help people detect and process spatial events more efficiently. Namely, when spatial distractors are present, they have greater impact. It seems reasonable that a short increase in arousal can be beneficial for performance under many circumstances. Generally, an increase in arousal reduces the time needed to execute a response. However, when selection is required, this benefit is compromised. In the future, it will be interesting to map which executive functions can be modulated by arousal and whether an optimal level of arousal can also improve different aspects of executive functions under some circumstances.
References
Baumeister, A. A., & Joubert, C. E. (1969). Interactive effects on reaction time of preparatory interval length and preparatory interval frequency. Journal of Experimental Psychology, 82, 393–395.
Böckler, A., Alpay, G., & Stürmer, B. (2011). Accessory stimuli affect the emergence of conflict, not conflict control: A Simon-task ERP study. Experimental Psychology, 58, 102–109.
Botvinick, M. M., Braver, T. S., Barch, D. M., Carter, C. S., & Cohen, J. D. (2001). Conflict monitoring and cognitive control. Psychological Review, 108, 624–652. doi:10.1037/0033-295X.108.3.624
Boulinguez, P., Ballanger, B., Granjon, L., & Benraiss, A. (2009). The paradoxical effect of warning on reaction time: Demonstrating proactive response inhibition with event-related potentials. Clinical Neurophysiology, 120, 730–737.
Callejas, A., Lupiáñez, J., Funes, M. J., & Tudela, P. (2005). Modulations among the alerting, orienting and executive control networks. Experimental Brain Research, 167, 27–37. doi:10.1007/s00221-005-2365-z
Chica, A. B., Thiebaut de Schotten, M., Toba, M., Malhotra, P., Lupiáñez, J., & Bartolomeo, P. (2012). Attention networks and their interactions after right-hemisphere damage. Cortex, 48, 654–653.
Correa, Á., Cappucci, P., Nobre, A. C., & Lupiáñez, J. (2010). The two sides of temporal orienting: Facilitating perceptual selection, disrupting response selection. Experimental Psychology, 57, 142–148. doi:10.1027/1618-3169/a000018
Costa, A., Hernández, M., & Sebastián-Gallés, N. (2008). Bilingualism aids conflict resolution: Evidence from the ANT task. Cognition, 106, 59–86. doi:10.1016/j.cognition.2006.12.013
Coull, J., & Nobre, A. (1998). Where and when to pay attention: The neural systems for directing attention to spatial locations and to time intervals as revealed by both PET and fMRI. Journal of Neuroscience, 18, 7426–7435.
Coull, J., Nobre, A., & Frith, C. (2001). The noradrenergic alpha 2 agonist clonidine modulates behavioural and neuroanatomical correlates of human attentional orienting and alerting. Cerebral Cortex, 11, 73–84.
Dye, M. W. G., Baril, D. E., & Bavelier, D. (2007). Which aspects of visual attention are changed by deafness? The case of the attentional network test. Neuropsychologia, 45, 1801–1811.
Dye, M. W. G., Green, C. S., & Bavelier, D. (2009). The development of attention skills in action video game players. Neuropsychologia, 47, 1780–1789. doi:10.1016/j.neuropsychologia.2009.02.002
Fan, J., Gu, X., Guise, K. G., Liu, X., Fossella, J., Wang, H., & Posner, M. I. (2009). Testing the behavioral interaction and integration of attentional networks. Brain and Cognition, 70, 209–220.
Fan, J., McCandliss, B. D., Sommer, T., Raz, A., & Posner, M. I. (2002). Testing the efficiency and independence of attentional networks. Journal of Cognitive Neuroscience, 14, 340–347. doi:10.1162/089892902317361886
Fernandez-Duque, D., & Posner, M. I. (1997). Relating the mechanisms of orienting and alerting. Neuropsychologia, 35, 477–486.
Fischer, R., Plessow, F., & Kiesel, A. (2010). Auditory warning signals affect mechanisms of response selection: Evidence from a Simon task. Experimental Psychology, 57, 89–97. doi:10.1027/1618-3169/a000012
Fischer, R., Plessow, F., & Kiesel, A. (2012). The effects of alerting signals in action control: Activation of S–R associations or inhibition of executive control processes? Psychological Research, 76, 317–328.
Fuentes, L. J., & Campoy, G. (2008). The time course of alerting effect over orienting in the attention network test. Experimental Brain Research, 185, 667–672.
Hackley, S. A. (2009). The speeding of voluntary reaction by a warning signal. Psychophysiology, 46, 225–233. doi:10.1111/j.1469-8986.2008.00716.x
Hackley, S. A., Langner, R., Rolke, B., Erb, M., Grodd, W., & Ulrich, R. (2009). Separation of phasic arousal and expectancy effects in a speeded reaction time task via fMRI. Psychophysiology, 46, 163–171. doi:10.1111/j.1469-8986.2008.00722.x
Ishigami, Y., & Klein, R. M. (2010). Repeated measurement of the components of attention using two versions of the attention network test (ANT): Stability, isolability, robustness, and reliability. Journal of Neuroscience Methods, 190, 117–128.
Lawrence, M. A., & Klein, R. M. (2013). Isolating exogenous and endogenous modes of temporal attention. Journal of Experimental Psychology: General, 142, 560–572. doi:10.1037/a0029023
MacLeod, J. W., Lawrence, M. A., McConnell, M. M., Eskes, G. A., Klein, R. M., & Shore, D. I. (2010). Appraising the ANT: Psychometric and theoretical considerations of the attention network test. Neuropsychology, 24, 637–659.
McConnell, M. M., & Shore, D. I. (2011). Mixing measures: Testing an assumption of the Attention Network Test. Attention, Perception, & Psychophysics, 73, 1096–1107. doi:10.3758/s13414-010-0085-3
Niemi, P., & Näätänen, R. (1981). Foreperiod and simple reaction time. Psychological Bulletin, 89, 133–162. doi:10.1037/0033-2909.89.1.133
Petersen, S. E., & Posner, M. I. (2012). The attention system of the human brain: 20 years after. Annual Review of Neuroscience, 35, 73–89. doi:10.1146/annurev-neuro-062111-150525
Posner, M. I., & Boies, S. J. (1971). Components of attention. Psychological Review, 78, 391–408. doi:10.1037/h0031333
Redick, T. S., & Engle, R. W. (2006). Working memory capacity and attention network test performance. Applied Cognitive Psychology, 20, 713–721.
Vallesi, A. (2010). Neuroanatomical substrates of foreperiod effects. In A. C. Nobre & J. T. Coull (Eds.), Attention and time (pp. 303–316). Oxford, UK: Oxford University Press.
Vallesi, A., Shallice, T., & Walsh, V. (2007). Role of the prefrontal cortex in the foreperiod effect: TMS evidence for dual mechanisms in temporal preparation. Cerebral Cortex, 17, 466–474.
Weinbach, N., & Henik, A. (2011). Phasic alertness can modulate executive control by enhancing global processing of visual stimuli. Cognition, 121, 454–458. doi:10.1016/j.cognition.2011.08.010
Weinbach, N., & Henik, A. (2012a). The relationship between alertness and executive control. Journal of Experimental Psychology: Human Perception and Performance, 38, 1530–1540. doi:10.1037/a0027875
Weinbach, N., & Henik, A. (2012b). Temporal orienting and alerting. The same or different? Frontiers in Psychology, 3, 236. doi:10.3389/fpsyg.2012.00236
Witte, E., & Marrocco, R. (1997). Alteration of brain noradrenergic activity in rhesus monkeys affects the alerting component of covert orienting. Psychopharmacology, 132, 315–323.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Weinbach, N., Henik, A. The interaction between alerting and executive control: Dissociating phasic arousal and temporal expectancy. Atten Percept Psychophys 75, 1374–1381 (2013). https://doi.org/10.3758/s13414-013-0501-6
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-013-0501-6