
Abstract
The Morris water maze is a popular task for examining spatial navigation and memory in rats. Historically, emphasis has been put on extramaze cues as the primary environmental feature guiding navigation and spatial memory formation. However, other features of the environment may also be involved. In this experiment, we trained rats on the spatial version of the Morris water maze over four days. A probe test was given 24 h after training, in which the shape of the pool either remained the same as during training or was changed to a different shape. Mass training of a new platform position in one training session was performed in a pool of one of these two shapes, with a second probe test being done 24 h afterward. The results showed that spatial training produces a spatial preference for the trained location in the probe test when the pool shape remains the same. However, changing the shape of the pool eliminates this preference. All groups learned the new platform position during mass training and also expressed a spatial preference for the mass-trained quadrant when tested 24 h later. The results from these experiments implicate the use of pool shape in guiding spatial navigation in the water maze and as a critical environmental feature represented in spatial memory.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Spatial navigation is a behavior composed of an organism’s abilities to actively move throughout its environment and to determine its own position as well as the positions of other features within that environment. Spatial navigation and spatial memory are tightly linked processes because the ability to successfully move along routes to locate objects or find locations that are associated with rewards depends on previous experience with those specific environmental features (Burgess, Maguire, & O’Keefe, 2002): The knowledge of how to get to a certain place necessitates a lasting memory of where that place is. The neurological basis of spatial navigation and memory in the mammalian brain has been studied for decades, and evidence shows that it is composed of several distinct neural systems and structures with their own unique functional contributions (Devan, Goad, & Petri, 1996; Dumont & Taube, 2015; Ekstrom et al., 2003; Moser, Kropff, & Moser, 2008; Sutherland, Whishaw, & Kolb, 1988). One central brain structure critical in spatial navigation and memory is the hippocampal formation.
The hippocampal formation is a cortical structure located in the medial temporal lobe. Genetic, biochemical, electrophysiological, and lesion evidence all point to the hippocampus as having a critical role in spatial navigation and memory processes (Abel et al., 1997; Moser, Krobert, Moser, & Morris, 1998; Sutherland, Whishaw, & Kolb, 1983). The electrophysiological activity patterns of distinct cell types in the hippocampus and parahippocampal tissues also suggest a role in spatial functions (Bostock, Muller, & Kubie, 1991; Clark, Harris, & Taube, 2012; Derdikman et al., 2009).
The Morris water maze has long been used as a task for examining spatial navigation and memory in rats. It is a simple task that uses a pool of opaque water with a submerged platform located inside. In order to escape the cool water, the rat must find the hidden platform, and training is thought to induce a spatial memory for this hidden platform location. Indeed, hundreds of studies over the years have implicated the hippocampus in spatial memory and the successful completion of this task (D’Hooge & De Deyn, 2001; McDonald & Hong, 2000; Vorhees & Williams, 2006). The most widely accepted explanation of how rats solve the spatial version of the water maze is by using the extramaze cues within the global environment (O’Keefe & Nadel, 1978). These environmental cues include visible items in the room, such as posters, doors, computers, windows, and so forth. It has been argued that the location of the platform is learned by determining the relationship between the distal cues and the platform location. Covering the pool with a black curtain does not allow for successful water maze learning and eliminates the expression of spatial memories after learning (Morris, 1984). However, multiple variations of the water maze exist, and manipulations to training protocols have revealed that several strategies are available to the animal for solving the task (McDonald & White, 1994). Determining which environmental features the rat uses, as well as how spatial memories form in the hippocampus in reference to these features, has been the subject of recent debate (Bannerman et al., 2012; Hamilton, Akers, Weisend, & Sutherland, 2007).
The water maze environment can be divided into at least two separate spaces. The first is the intramaze (local) environment, which contains everything inside the pool walls, including the shape of the pool, the distance between the platform and the pool wall, and any intramaze cues that might be available. The second is the extramaze (global) environment, which contains everything outside the pool walls but inside the experiment room. This would include the distances between extramaze cues as well as the distance between the platform and the room wall or the maze itself and the room wall. The physical features that are present, where they are, and the room shape are all components of the extramaze environment (Cheng & Newcombe, 2005). Some evidence has shown that certain aspects of spatial navigation can transfer between multiple contexts, and these aspects would thus constitute more general, environment-independent spatial processes (Dudchenko & Zinyuk, 2005; Taube & Burton, 1995).
Many studies have provided evidence for alternative explanations of how rats solve the standard spatial version of the water maze, as well as highlighting the contributions that different neural systems provide to spatial navigation in this version of the task. It has also been argued that the types of search strategies employed by rats change over the course of training (Graziano, Petrosini, & Bartoletti, 2003). Rats can also successfully solve the spatial version of the water maze with varying degrees of available extramaze information (Baldi, Lorenzini, & Bucherelli, 2003). In some instances, their search may be either idiothetic (i.e., internally guided), using path integration (Whishaw & Jarrard, 1996), or procedural, using cue-guided motor sequences (McDonald & White, 1994). Rats may also perform the water task equally well regardless of cue salience (Young, Choleris, & Kirkland, 2006), indicating that flexibility in their use of both search strategies and the available environmental features is high.
Hamilton, Akers, Weisand, and Sutherland (2007) and later Hamilton et al. (2009) trained rats on the spatial-version water maze, and for the probe test they moved the pool to another position in the room. The procedures in this experiment were designed in such a way that if rats had formed a spatial location memory using extramaze cues, they would swim to a particular spatial location in the room, but if they had formed a directional memory, they would swim to a different location. Directional learning was found in all variations of the task used, except when the pool walls were visually eliminated by raising the water level. These findings indicate that the pool itself may have an important role in guiding navigation—specifically, heading direction.
With this in mind, we conducted a related behavioral study using the water maze and multiple training rooms, in which rats were given standard spatial training in one room and then a probe test in a different room (Clark, Bettenson, Woolford, Horwood, & McDonald, 2015). None of the extramaze information was the same between the rooms, and the only spatial feature that remained constant was the shape of the pool. It was found that spatial training in one room led to a consistent heading direction in the rats’ swimming when they were given a probe in a new room: When placed in the new pool for the first time, the rats would consistently swim in the same direction. Spatial disorientation, a procedure in which a rat is put in an opaque box and slowly spun around for 1 min, thereby eliminating the rat’s previous directional orientation, was shown to eliminate this directional swimming during the probe. This study revealed that heading direction can be guided by information independent of any extramaze cues. One possibility was that the shape of the pool combined with head direction information was mediating the retention of the directional response in the new room.
In the present study we sought to manipulate the shape of the pool itself, to determine whether it plays a critical role in the formation of spatial memories in the water maze. Rats were trained on the spatial version of the Morris water maze in pool with one of two different shapes. After successful learning, the pool shape was changed, and a probe test was performed to determine how pool shape influences the retrieval of spatial memory. Mass training to a new spatial location followed this, and a probe test was given 24 h later in the mass-trained pool shape. In this way, we could observe the effects of pool shape on new learning, as well. Given the findings of past research, we believed that rats would use the pool shape as a critical piece of environmental information to guide spatial navigation and memory (Clark et al., 2015; Hamilton et al., 2009). We hypothesized that a change in pool shape would eliminate spatial preference during the probe test and eliminate interference from previous training during mass training, resulting in enhanced new learning.
Materials and method
Subjects
Thirty male Long Evans rats 90 days of age were used for the study. Upon arrival, the rats were pair-housed and kept on a 12-h light/12-h dark cycle, with lights turning on at 7:30 and off at 19:30. The rats had ad libitum access to both water and food. The rats were allowed seven days of acclimatization in their home cages to reduce stress induced from travel. After this period, all rats were handled for 5 min a day for four days to familiarize them with the male experimenter and with being manipulated. The weight range at the start of the experiment was between 300 and 350 g. All procedures were in accordance with the regulations set out by the Canadian Council of Animal Care and were approved by the University of Lethbridge Animal Care Committee.
Pools
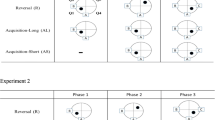
Two different pool shapes were used in these experiments (Fig. 1): a white circular pool with a diameter of 127 cm and a total surface area of 12,667 sq. cm, as well as a white square inlay made of plastic that fit into the circular pool, with a wall length of 90 cm and a total surface area of 8,100 sq. cm (63% of the circle). The pool walls were 48 cm in height and the water was filled up to 33 cm, leaving 15 cm of exposed wall inside the pools. The top of the platform was 2 cm below the surface of the water. When the square pool was placed inside the circular pool, it was held in place with transparent tape that was outside of the rats’ field of vision and therefore was not a local, intramaze cue. The square pool was kept in the same position throughout all training procedures. For analysis, the pools were divided into four equal-sized quadrants indicated by the cardinal directions (N W S E). All behavioral training and testing occurred in the same room, which was 3 m wide by 4.5 m long. The extramaze cues included three large geometric-shaped posters placed on the white walls, a computer, an upright storage shelf, and the experimenter (Fig. 1B).

Experimental design of the pool shape experiments: (A) Order of the pool shapes and platform locations. (B) Layout of the room used in the entire experiment; the X is where the experimenter was located throughout.
Behavioral procedures
A three-phase procedure was used to assess the effect of pool shape on rats’ spatial navigation behaviors. For Phase 1, the rats were trained on the standard spatial version of the Morris water maze. A hidden escape platform was located in the SW quadrant of the pool. Animals were given eight trials per day over four days (32 in total), with a random start point used for each trial (i.e., NE, SW, NW, SE, SW, NE, NW, or SW). All eight trials were completed in a single training session. Rats were placed in the pool facing the wall and were given 60 s to find the platform. If they did not find the platform, they were removed from the water and placed on the platform. After finding the platform or being aided by placement upon it, they remained there for 10 s and were then brought back to their holding cage, where they waited for the next trial.
After Phase 1, the rats were assigned to either the New group, in which the pool shape changed for Phase 2, or the Same group, in which the pool shape remained the same as in the previous training. Because the square pool was placed inside the circular pool, it was necessarily smaller and provided a smaller surface area available for navigation. To ensure that the observable effects seen were due to the fact that the pool shape was changed, and not because of the order of the shape change, surface area limitations, or movement restrictions, pool shape was counterbalanced within the groups (Fig. 1). Half of the rats in the New group (n = 7) were trained in the square first and then tested in the circle, and the other half (n = 8) were trained in the circle first and then in the square. Half of the rats in the Same group were trained in the circle throughout all training procedures (n = 8), and the other half were trained in the square throughout all training procedures (n = 7).
Phase 2 consisted of two parts. First the platform was removed, and the rats were given a 30-s probe test in either the same or the new shape. Respectively in each experiment, the Same group probe was done in the pool shape that had been used in Phase 1 training, and the New group probe was done in the new shape. This was done to determine whether the place memory formed during Phase 1 was dependent on pool shape. After this probe, the platform was placed in the NE quadrant, opposite that in Phase 1, and the rats were mass-trained to this new position. This consisted of 15 random start-point trials occurring within a 2-h time period. The pool shape used for this part of Phase 2 was identical to the shape used during the probe test for each condition (Fig. 1). This was done to examine the influence of pool shape on new spatial learning.
Phase 3 consisted of a 30-s probe test done 24 h after the completion of Phase 2. The pool shape used for Phase 3 was identical to that used in Phase 2 for each condition. This was done in order to determine how well the new location was remembered.
Measures and statistics
For all training procedures, the latency and path length to find the platform during a trial were used as the measures of learning. For all probe tests, the percentages of swim time spent in the individual quadrants were used as a measure of spatial memory. Initial heading error (1–2 s) was calculated from the line between where the rat was placed in the pool and the platform location, and was used as a measure of heading angle. Heading error was measured as the average deviation from this line between seconds 1 and 2 of the trial, instead of from the exact start of the trial. The 1-s delay was meant to ensure that the rat had spun around, because the rats were placed into the pool facing the wall, and the spin could add unnecessary variability. The raw data for all measures were taken using the Noldus Ethovison XT software and a ceiling camera directly above the pool. Mixed-model analysis of variance (ANOVA), with group as the between-subjects factor and day or quadrant as the repeated measures factor, was used to analyze both the learning curves and probe tests. In the case of significant interactions, post-hoc pairwise comparisons with a Bonferroni correction were conducted. Since there was not a cohort effect in terms of whether the animals were trained in a circle or a square, and since equal numbers of animals from both groups were trained in the circle and the square, the effect of training pool shape was not included in our statistical analyses. Instead, the Same group was simply compared to the New group. All of the statistics were calculated with SPSS 21 (IBM, Armonk, New York, NY), and the GraphPad Prism software (GraphPad, La Jolla, CA) was used to make all of the graphs.
Results
Acquisition during Phase 1 is represented as the averages of the latencies and path lengths to find the platform on that day of training (eight per day). Over the four days of training, both groups significantly reduced their latencies and path lengths. The first-day average latencies for the Same and New groups were 27.5 and 27.8 s, and the last-day average latencies were 6.8 and 8.2 s, respectively (Fig. 2A). Two-way mixed-model ANOVA revealed a significant effect of day [F(3, 84) = 62.473, p < .001, ηp2 = .691]. No significant differences were observed between the groups during this phase of training [F(1, 28) = 0.038, p > .05], nor was there an interaction effect [F(3, 84) = 0.450, p > .05]. The first-day average path lengths for the Same and New groups were 8.3 and 7.6 m, and the last-day average path lengths were 1.7 and 2.0 m, respectively (Fig. 2B). Two-way mixed-model ANOVA again revealed that there was a significant effect of day [F(3, 84) = 78.456, p < .001, ηp2 = .737]. No significant differences were observed between the groups during this phase of training [F(1, 28) = 0.902, p > .05], nor did we observe an interaction effect [F(3, 84 = 0.845, p > .05]. This indicates that both groups learned over the four days of training and that no differences were observed between them during Phase 1.

(A) Latencies to find the platform during Phase 1 standard training. Each point is the average of eight trials; training was eight trials a day for four days. (B) Path lengths to find the platform during standard training. Each point is the average of eight trials, and training was eight trials a day for four days. Error bars indicate ± 1 SE. *p < .05
The first part of Phase 2 consisted of a 30-s probe test with the platform removed from the pool. The percentage of the total swim time spent in the target quadrant the (Phase 1 training quadrant) was compared to the average swim time in the other three quadrants (Fig. 3). The Same group appeared to spend more time in the target quadrant than in the others, whereas no preference was observed in the New group. A two-way mixed-model ANOVA revealed significant effects of quadrant [F(1, 28) = 34.201, p < .001, ηp2 = .550] and group [F(1, 28) = 15.797, p < .001, ηp2 = .361], as well as their interaction [F(1, 28) = 15.797, p < .001, ηp2 = .361]. Given these results, a post-hoc pairwise comparison revealed a significant difference between the target and averaged quadrants for the Same group (p < .001), but not for the New group (p = .196). These results indicate that when the pool shape used during a probe is the same as that used during training, a spatial preference is seen. However, when the pool shape is different from that used during training, this preference disappears.

The 30-s probe test done 24 h after completing standard training, comparing the percentages of swim time spent in the quadrant where the pretrained platform was located versus an average of the other three quadrants. The Same group probe was done in the same pool shape used in standard training. The New group probe was done in the new pool shape, different from the one used in standard training. Error bars indicate ± 1 SE. *p < .001
The second part of Phase 2 involved mass training, wherein the hidden platform was moved to the opposite quadrant of the pool. Mass training is represented as the individual trial latencies and path lengths to find the platform. Both groups significantly reduced their latency times over mass training. The first-trial latency for the Same group was 33.5 s, whereas the latency for the New group was 36.3 s. By the end of training, the Same and New groups had latencies of 6.9 and 5.3 s, respectively (Fig. 4A). A two-way mixed-model ANOVA revealed an effect of trial [F(14, 392) = 19.338, p < .001, ηp2 = .409], but no effect of group [F(1, 28) = 0.004, p > .05] and no interaction [F(14, 392) = 0.698, p > .05]. The first-trial path length for the Same group was 8.7 m, whereas the path length for the New group was 9.0 m. By the end of training, the Same and New groups had path lengths of 1.7 and 1.4 m, respectively (Fig. 4B). A two-way mixed-model ANOVA revealed an effect of trial [F(14, 392) = 17.024, p < .001, ηp2 = .378], but no effect of group [F(1, 28) = 0.001, p > .05] and no interaction [F(14, 392) = 0.359, p > .05]. Both groups significantly reduced their path lengths over mass training.

(A) Latencies to find the platform during Phase 2 mass training to a new platform position. (B) Path lengths to find the platform during mass training to a new platform position. Mass training was 15 trials over the course of 2 h. Error bars indicate ± 1 SE.
Next, 24 h after mass training, a second probe test was conducted. The percentage of swim time spent in the target quadrant (the Phase 2 mass-training quadrant) was compared to the average percentage swim time of the other three quadrants (Fig. 5). A two-way repeated measures ANOVA revealed a significant effect of quadrant [F(1, 28) = 28.886, p < .001, ηp2 = .508], but no group effect [F(1, 12) = 0.283, p > .05] and no interaction [F(1, 28) = 0.283, p > .05]. The results from the 24-h probe test showed that the rats rapidly formed a memory for a new platform position, regardless of shape.

The 30-s probe test done 24 h after completing mass training, comparing the percentages of swim time spent in the quadrant where the mass-trained platform was located versus an average of swim times for the other three quadrants. The Same group probe occurred in the same pool shape used in both standard and mass training. The New group probe occurred in the same pool shape used in massed training, but different from the shape during standard training. Error bars indicate ± 1 SE. *p < .001
Heading error
Finally, initial heading error was analyzed. The initial heading error was defined as the average deviation from the line between the starting point and the platform location over the time period from 1 to 2 s during the trial. A one-way ANOVA revealed a significant difference between the New group and the Same group [F(1, 28) = 5.512, p < .05]. To assess unsystematic errors that might be related to the differing pool shapes, we also performed Levene’s test for the homogeneity of variances. Levene’s test showed that there was no significant difference in variances between the New and Same group heading directions [F(1, 28) = 0.183, p = .682], which suggests that systematic error accounted for much of the observed variance. These results indicated that changing the pool shape can impair a rat’s initial heading angle. The swim paths can be found in Fig. 6A.

(A) Initial swim paths for Probe 1 in the New and Same groups. (B) Initial heading error was measured as the average deviation from the line between the starting point and the platform location over the interval from second 1 to 2 of the trial. Error bars indicate ± 1 SE. *p < .05
Discussion
In this experiment, we showed that pool shape has an important role in the formation of spatial memories in the Morris water maze. After four days of standard training, changing the shape of the pool eliminated the spatial preference seen when the pool remained the same. Pool shape change resulted in an elimination of spatial preference during the probe test. Both groups successfully learned the new platform position during massed reversal, even with the pool shape change in the New group. Similarly, both groups had a preference for the mass-trained quadrant during a subsequent probe trial.
External cues or pool shape?
Extramaze cues are distinct cues, whereas pool shape is a geometrical and local feature. Changing the pool shape after distributed training eliminates the spatial preference it normally produces. This same effect is produced by removing all of the extramaze cues through the use of a curtain (Morris, 1984). In our experiment, even though all of the extramaze cues were present throughout every phase, the impairment was still seen. If removing extramaze cues but keeping pool shape, as well as removing pool shape but keeping extramaze cues, both result in the same impairment, this suggests that rats do not use a single feature and/or representation in their environment to guide spatial navigation. Rats may accomplish successful navigation using many different features of the environment, or they may employ different strategies (Hamilton et al., 2007; Huxter, Thorpe, Martin, & Harley, 2001; Skinner, Horne, Murphy, & Martin, 2010).
Consistent with this idea, the orientation of head direction cells, recorded from various regions of the hippocampal formation, can be determined by either extramaze cues or maze geometry, depending on the salience of the two environmental features in relation to each other (Clark, Harris & Taube, 2012). The more extramaze information is available, the more likely it is that head direction cells will tune their orientation with reference to the cues. Less extramaze information increases the likelihood that the cells will tune with reference to the maze shape. These findings offer neurophysiological evidence of changing cue salience in guiding spatial navigation.
Clark et al.’ (2015) study supports these neurophysiological findings by suggesting that heading direction might be mediated by local cues such as pool shape in the absence of learned extramaze cues. The finding from the present study, that heading direction changes with the shape of the pool despite the learned extramaze cues being static, also suggests that heading direction is impacted by local and not just extramaze cues.
These findings fit with the work done by Hamilton, Akers, Weisend, and Sutherland (2007), and later by Hamilton et al. (2009), showing that the pool wall is a primary feature of water maze training that guides heading direction and that the expression of learned spatial location was only possible when the pool walls were removed. However, rats are also fully capable of learning hidden platform locations using only pool shape (Pearce, Ward-Robinson, Good, Fussell, & Aydin, 2001), and distinct individual features of these shapes guide this learning (Pearce, Good, Jones, & McGregor, 2004). Tommasi and Thinus-Blanc (2004) showed not only that rats could use the shape of the training environment in a hidden-food task to learn the location of the food, but that this ability could be transferred across shapes. This is somewhat inconsistent with our results; nonetheless, collectively these works lead us to conclude that a large part of spatial learning can be done without extramaze cues and instead relies on the apparatus geometry.
Probe tests
The important tests in our experiment were the probes, which yielded the percentages of total swim times in each quadrant, a size-invariant measure of spatial preference. Even if one pool were twice the size of another, a rat with a strong spatial preference would still swim to the appropriate quadrant and swim within it. The size of the water maze pools used in rat experiments is not the same across laboratories, ranging in diameter from 1.2 m (Graziano, Petrosini, & Bartoletti, 2003) to 1.5 m (Hamilton et al., 2007) to 2 m (Morris, Steele, Bell, & Martin, 2013), and spatial preferences are still observed in all of these cases. In the present study, the magnitude of the observed effects suggests that both arrangements of shape change resulted in the elimination of spatial preference, regardless of pool size.
Interestingly, the Phase 3 probe test done 24 h after mass training disproved our initial hypothesis that if a novel pool shape could disrupt spatial preference, it would also enhance new spatial learning. This result was predicted because of a competition effect between two different representations: one for the previous platform location, and the other for the new one. If the expression of the first spatial representation was dependent on pool shape, which we found it to be, then learning a new platform location in that same shape would be a competition between the new and the old, and so would result in a slower learning curve and possibly in no stable preference 24 h after mass training. A new shape would then be a distinct training environment and offer novel spatial associations, which could be made without interference from the old ones made during standard training, so that learning would be faster and a preference more easily seen. This, however, was not the case, because both groups learned equally well, and both the Same and the New group probes showed equal preference.
The results of the Clark et al. (2015) study are also important regarding the probe results, because one possible explanation for the disruption of spatial memory following the pool shape manipulations in the present experiment is that it was simply a novelty effect. That is, during the first probe trial the different pool shape was novel, and it is possible that spatial navigation was disrupted simply due to a neophobia effect. According to this interpretation, the disrupted spatial behavior during Probe 1 did not occur because crucial information (pool shape) had been removed, but simply because the normal navigational behavior was disrupted by a competing behavioral response to novelty. The results from the Clark study suggest that this was not the case. In that study, rats were trained on the standard spatial version of the water task and then transferred to a completely new context for the mass-training procedures. The rats transferred to this novel context continued to show a place response based on head direction, and probably on pool shape, acquired in the familiar context.
The present results provide some insight concerning the type of environmental information that rats use during spatial navigation. Local cues, such as the geometric shape of the training environment, appear to be an important feature of spatial navigation and spatial memory (Hamilton et al., 2007). Different components of navigation are known to rely on different brain structures and neural processes, such as on head direction representations in the subiculum (Taube, Muller, & Rank, 1990), place cell formation in the hippocampus proper (O’Keefe, 1976), and cue-guided navigational processes in the striatum (McDonald & White, 1994). Geometric modules have been proposed, potentially involving parahippocampal cortical structures (Cheng & Newcombe, 2005), and the influence of individual features of geometric components such as corner angles have also been studied behaviorally (Pearce et al., 2004; Kosaki, Austen, & McGregor, 2013). Exploring the biological basis of these different spatial processes will be key to understanding how organisms move through their environment. Future work in this domain may help us understand the complexity of how animals navigate and what they use in their environments, as well as the neural circuitry behind these behaviors.
References
Abel, T., Nguyen, P. V., Barad, M., Deuel, T. A., Kandel, E. R., & Bourtchouladze, R. (1997). Genetic demonstration of a role for PKA in the late phase of LTP and in hippocampus based long-term memory. Cell, 88, 615–626.
Baldi, E., Lorenzini, C. A., & Bucherelli, C. (2003). Task solving by procedural strategies in the Morris water maze. Physiology & Behavior, 78, 785–793. doi:https://doi.org/10.1016/S0031-9384(03)00064-7
Bannerman, D. M., Bus, T., Taylor, A., Sanderson, D. J., Schwarz, I., Jensen, V., . . . Sprengel, R. (2012). Dissecting spatial knowledge from spatial choice by hippocampal NMDA receptor deletion. Nature Neuroscience, 15, 1153–1159. doi:https://doi.org/10.1038/nn.3166
Bostock, E., Muller, R. U., & Kubie, J. L. (1991). Experience-dependent modifications of hippocampal place cell firing. Hippocampus, 1, 193–205.
Burgess, N., Maguire, E. A., & O’Keefe, J. (2002). The human hippocampus and spatial and episodic memory. Neuron, 35, 625–641. doi:https://doi.org/10.1016/S0896-6273(02)00830-9
Cheng, K., & Newcombe, N. S. (2005). Is there a geometric module for spatial orientation? Squaring theory and evidence. Psychonomic Bulletin & Review, 12, 1–23. doi:https://doi.org/10.3758/BF03196346
Clark, B. J., Hong, N. S., Bettenson, B. J., Woolford, B. J., Horwood, L., & McDonald, B. J. (2015). Maintained directional navigation across environments in the Morris water task is dependent on vestibular cues. Journal of Experimental Psychology: Animal Behavior Processes, 41, 301–308. doi:https://doi.org/10.1037/xan0000066
Clark, B. J., Harris, M. J., & Taube, J. S. (2012). Control of anterodorsal thalamic head direction cells by environmental boundaries: Comparison with conflicting distal landmarks. Hippocampus, 22, 172–187. doi:https://doi.org/10.1002/hipo.20880
D’Hooge, R., & De Deyn, P. P. (2001). Applications of the Morris water maze in the study of learning and memory. Brain Research Reviews, 36, 60–90.
Derdikman, D., Whitlock, J. R., Tsao, A., Fyhn, M., Hafting, T., Moser, M. B., & Moser, E. I. (2009). Fragmentation of grid cell maps in a multicompartment environment. Nature Neuroscience, 12, 1325–1332. doi:https://doi.org/10.1038/nn.2396
Devan, B. D., Goad, E. H., & Petri, H. L. (1996). Dissociation of hippocampal and striatal contributions to spatial navigation in the water maze. Neurobiology of Learning and Memory, 66, 305–323.
Dudchenko, P. A., & Zinyuk, L. E. (2005). The formation of cognitive maps of adjacent environments: Evidence from the head direction cell system. Behavioral Neuroscience, 119, 1511–1523. doi:https://doi.org/10.1037/0735-7044.119.6.1511
Dumont, J. R., & Taube, J. S. (2015). The neural correlates of navigation beyond the hippocampus. Progress in Brain Research, 219, 83–102.
Ekstrom, A. D., Kahana, M. J., Caplan, J. B., Fields, T. A., Isham, E. A., Newman, E. L., & Fried, I. (2003). Cellular networks underlying human spatial navigation. Nature, 425, 184–188.
Graziano, A., Petrosini, L., & Bartoletti, A. (2003). Automatic recognition of explorative strategies in the Morris water maze. Journal of Neuroscience Methods, 130, 33–44.
Hamilton, D. A., Akers, K. G., Weisend, M. P., & Sutherland, R. J. (2007). How do room and apparatus cues control navigation in the Morris water task? Evidence for distinct contributions to a movement vector. Journal of Experimental Psychology: Animal Behavior Processes, 33, 100–114. doi:https://doi.org/10.1037/0097-7403.33.2.100
Hamilton, D. A., Akers, K. G., Johnson, T. E., Rice, J. P., Candelaria, F. T., & Redhead, E. S. (2009). Evidence for a shift from place navigation to directional responding in one variant of the Morris water task. Journal of Experimental Psychology: Animal Behavior Processes, 35, 271–278. doi:https://doi.org/10.1037/a0013260
Huxter, J. R., Thorpe, C. M., Martin, G. M., & Harley, C. W. (2001). Spatial problem solving and hippocampal place cell firing in rats: Control by an internal sense of direction carried across environments. Behavioural Brain Research, 123, 37–48.
Kosaki, Y., Austen, J. M., & McGregor, A. (2013). Overshadowing of geometry learning by discrete landmarks in the water maze: Effects of relative salience and relative validity of competing cues. Journal of Experimental Psychology: Animal Behavior Processes, 39, 126–139. doi:https://doi.org/10.1037/a0031199
McDonald, R. J., & Hong, N. S. (2000). Rats with hippocampal damage are impaired on place learning in the water task when overtrained under constrained conditions. Hippocampus, 10, 153–161.
McDonald, R. J., & White, N. M. (1994). Parallel information processing in the water maze: Evidence for independent memory systems involving dorsal striatum and hippocampus. Behavioral and Neural Biology, 61, 260–270.
Morris, R. (1984). Developments of a water-maze procedure for studying spatial learning in the rat. Journal of Neuroscience Methods, 11, 47–60.
Morris, R. G. M., Steele, R. J., Bell, J. E., & Martin, S. J. (2013). N-methyl-d-aspartate receptors, learning and memory: Chronic intraventricular infusion of the NMDA receptor antagonist d-AP5 interacts directly with the neural mechanisms of spatial learning. European Journal of Neuroscience, 37, 700–717. doi:https://doi.org/10.1111/ejn.12086
Moser, E. I., Krobert, K. A., Moser, M. B., & Morris, R. G. (1998). Impaired spatial learning after saturation of long-term potentiation. Science, 281, 2038–2042.
Moser, E. I., Kropff, E., & Moser, M. B. (2008). Place cells, grid cells, and the brain’s spatial representation system. Annual Review of Neuroscience, 31, 69–89.
O’Keefe, J. (1976). Place units in the hippocampus of the freely moving rat. Experimental Neurology, 51, 78–109.
O’Keefe, J., & Nadel, L. (1978). The hippocampus as a cognitive map. Oxford, UK: Oxford University Press, Clarendon Press.
Pearce, J. M., Good, M. A., Jones, P. M., & McGregor, A. (2004). Transfer of spatial behavior between different environments: Implications for theories of spatial learning and for the role of the hippocampus in spatial learning. Journal of Experimental Psychology: Animal Behavior Processes, 30, 135–147. doi:https://doi.org/10.1037/0097-7403.30.2.135
Pearce, J. M., Ward-Robinson, J., Good, M., Fussell, C., & Aydin, A. (2001). Influence of a beacon on spatial learning based on the shape of the test environment. Journal of Experimental Psychology: Animal Behavior Processes, 27, 329–344. doi:https://doi.org/10.1037/0097-7403.27.4.329
Skinner, D. M., Horne, M. R., Murphy, K. E., & Martin, G. M. (2010). Rats’ orientation is more important than start point location for successful place learning. Journal of Experimental Psychology: Animal Behavior Processes, 36, 110–116. doi:https://doi.org/10.1037/a0015773
Sutherland, R. J., Whishaw, I. Q., & Kolb, B. (1983). A behavioural analysis of spatial localization following electrolytic, kainate- or colchicine-induced damage to the hippocampal formation in the rat. Behavioural Brain Research, 7, 133–153.
Sutherland, R. J., Whishaw, I. Q., & Kolb, B. (1988). Contributions of cingulate cortex to two forms of spatial learning and memory. Journal of Neuroscience, 8, 1863–1872.
Taube, J. S., & Burton, H. L. (1995). Head direction cell activity monitored in a novel environment and during a cue conflict situation. Journal of Neurophysiology, 74, 1953–1971.
Taube, J. S., Muller, R. U., & Ranck, J. B. (1990). Head-direction cells recorded from the postsubiculum in freely moving rats: I. Description and quantitative analysis. Journal of Neuroscience, 10, 420–435.
Tommasi, L., & Thinus-Blanc, C. (2004). Generalization in place learning and geometry knowledge in rats. Learning and Memory, 11, 153–161. doi:https://doi.org/10.1101/lm.60904
Vorhees, C. V., & Williams, M. T. (2006). Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nature Protocols, 1, 848–858.
Whishaw, I. Q., & Jarrard, L. E. (1996). Evidence for extrahippocampal involvement in place learning and hippocampal involvement in path integration. Hippocampus, 6, 513–524.
Young, G. S., Choleris, E., & Kirkland, J. B. (2006). Use of salient and non-salient visuospatial cues by rats in the Morris Water Maze. Physiology & Behavior, 87, 794–799.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Bye, C.M., Hong, N.S., Moore, K. et al. The effects of pool shape manipulations on rat spatial memory acquired in the Morris water maze. Learn Behav 47, 29–37 (2019). https://doi.org/10.3758/s13420-018-0319-0
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13420-018-0319-0