
Abstract
In the contingent-capture protocol, singleton cues that have a target’s searched-for feature capture attention, but cues that do not have the target’s searched-for feature do not, a result labeled the contingent-capture effect. The contingent-capture effect is usually regarded as evidence for the observers’ ability to establish search settings for certain nonspatial features in a top-down manner. However, in recent years it has become increasingly clear that selection history is also a powerful mediator of attentional capture. In this vein, it has been suggested that contingent-capture effects could emerge as a result of (intertrial) priming: The idea is that features that have been encountered previously in the target are primed, so that cues that have these features automatically capture attention in a subsequent encounter. Here we tested a strong version of the priming account of the contingent-capture effect. We wanted to know whether cues that had target features would capture attention when the corresponding features were not part of the instructions (i.e., when the corresponding features were task-irrelevant). The results suggested that a strong version of the priming account of contingent capture is not supported. In five experiments, we found little evidence that the contingent-capture effect could be explained by (intertrial) priming of task-irrelevant features alone. These results show that processes beyond priming through task-irrelevant features are critical for contingent-capture effects.
Similar content being viewed by others

The results of spatial-cueing studies are often regarded as evidence for humans’ ability to attend willingly to specific features, such as shape, color, or spatial frequency, in a top-down manner (Folk, Remington, & Johnston, 1992). Folk et al. showed that spatial pretarget cues lead to validity effects (i.e., faster responses to targets that appear at the same location as a preceding cue [valid trials] than to targets that appear at a different location [invalid trials]) only when these cues have the specific features that participants have been instructed to search for. These are the top-down matching cues. When the cues do not have the searched-for target feature (i.e., in the case of a nonmatching cue), the cues do not lead to a validity effect. This pattern of results is labeled the contingent-capture effect and has been replicated many times under various conditions (e.g., Carmel & Lamy, 2014; Eimer, Kiss, Press, & Sauter, 2009; Goller & Ansorge, 2015; for a review, see Büsel, Voracek, & Ansorge, 2018). It is regarded as evidence that people are able to attend to certain nonspatial features in a top-down way, because cues that have the target’s critical searched-for feature capture attention as if the observers were willingly attending to the corresponding features.
However, in recent years it has become increasingly clear that selection history is a powerful mediator of attentional selection (e.g., Awh, Belopolsky, & Theeuwes, 2012; see also Theeuwes, 2018). Selection history denotes a set of implicit biases in the attentional selection process, including statistical learning (e.g., Umemoto, Scolari, Vogel, & Awh, 2010), (intertrial) priming effects (e.g., Belopolsky, Schreij, & Theeuwes, 2010), and reward-based biases (e.g., Anderson, 2016). Of most interest in the context of the present study, prior research has suggested that contingent-capture effects could be explained by priming (Belopolsky et al., 2010; see also Folk & Remington, 2008; Theeuwes, 2013, 2018). The general idea is straightforward: In almost every study that has reported contingent-capture effects, participants searched for a target with a specific feature (e.g., red color), and the target that was selected therefore had this feature in each and every trial. It is hence possible that merely seeing the corresponding feature in the target on every trial primed the attentional system, so that matching cues (cues with the corresponding feature) captured attention. Studies that have reported priming in contingent-capture experiments have usually investigated intertrial priming effects from the target features of a preceding trial to the cue features of a given trial (Folk & Remington, 2008). Some studies have failed to find the effect (Ansorge & Horstmann, 2007; Irons, Folk, & Remington, 2012). It is known, however, that priming does not only work from one trial to the next. Instead, priming effects can accumulate over several trials when the same feature is repeatedly encountered in the attended objects (e.g., Kristjánsson & Campana, 2010; Kruijne, Brascamp, Kristjánsson, & Meeter, 2015; Maljkovic & Nakayama, 1994; but see Ansorge & Becker, 2012).
In contingent-capture studies, cumulative priming could emerge in different ways. First, it is conceivable that priming effects could accumulate because observers repeatedly encounter the searched-for features in the targets when a nonmatching cue is presented between the targets. If all stimuli in nonmatching cueing displays remained unattended, the cumulative priming between two subsequent encounters of targets would not be interrupted by the stimuli in nonmatching cueing displays.Footnote 1 Second, in matching-cue conditions, in which the cue and target colors are the same, cumulative priming could theoretically take effect both between the target and cue and between the cue and target. Although it should be noted that invalid preceding trials can mitigate cueing effects—that is, they might even diminish cumulative priming to some extent (Goller & Ansorge, 2015)—it is possible that with a single, fixed searched-for target feature, at least two immediately succeeding matching valid trials would further enhance the cumulative priming effect, because matching valid cues are usually attended, and the corresponding feature would be primed and not down-weighed.Footnote 2
These considerations show that the effect of priming from target to cue (from one trial to the next), which has been reported by some studies but not by others, might only be the tip of the iceberg of an otherwise stronger effect that accumulates over several subsequent repetitions of the corresponding feature in the attended object—for example, as in contingent-capture effects measured with blocked target features (Theeuwes, 2013). Hence, we use the term “priming” instead of “intertrial priming” in what follows, to emphasize the fact that priming from a preceding trial’s target to a given trial’s cue is maybe only part of the story. Only when we speak about specific priming effects from a preceding trial n–1’s target feature to a cue’s feature in a current trial n will we use the term “intertrial priming.”
It is still unclear, however, to what extent the priming account is a real alternative to the contingent-capture theory and the idea that observers are able to set top-down attentional control settings for nonspatial features at will. In fact, it is debatable how priming relates to classical top-down control of attention and to what extent voluntary feature-based attention is important in priming, as well. Becker (2018), for instance, pointed out that priming is in many cases dependent on the task relevance of the corresponding features, and she used this as an argument against the idea that priming is automatic and impervious to top-down control. We concur with the view that the task relevance of features in priming is an important issue in this context. If priming was independent of the features’ task relevance, it would be possible for contingent-capture effects to emerge based on feature repetition alone, without any sort of instruction directing attention at the corresponding features. This sort of priming account could indeed be regarded as a true alternative to the top-down contingent-capture effect. If this was not the case, and priming as such depends on the instructions and corresponding feature-based attention, the notion that priming is a true alternative to contingent capture would be debatable (cf. Becker, 2018).
We, therefore, think that it is important to clarify to what extent the emergence of contingent-capture effects is possible through priming by uninstructed task-irrelevant features, and to what extent priming itself depends on the instructions, and thus on the instilled task relevance of the corresponding features. Regarding existing research on the matter: On the one hand, a body of evidence suggests that priming effects are in many cases contingent on, or at least enhanced by, the task relevance of the corresponding features (e.g., Fecteau, 2007; Kristjánsson, 2006). On the other hand, priming is not in all cases restricted to task-relevant features. Kristjánsson reported for the priming of pop-out effect that at least target color consistently gave rise to significant priming effects, even when it was not task-relevant. Indeed, most theories of visual attention assume that the selection of an object leads to the selection and, hence, the processing of all its features, regardless of their task relevance (e.g., Duncan, 1998), so that it may very well be possible for even task-irrelevant target features to be primed. Prior work has addressed the question of the extent to which task-irrelevant object features are processed when the corresponding object is attended to. Remington and Folk (2001) investigated whether attending to a specific object leads to the processing of all its features, irrespective of the present goals. In an attentional-cueing study, the authors measured congruence effects between the response features in a foil (an object that was presented in the target display in addition to the target and carried the response features of the target) and in a target. When a cue directed attention to the foil, a task-relevant feature dimension of the foil was processed, but a task-irrelevant feature dimension was not. At the same time, however, the target’s task-irrelevant feature dimension was processed and did interfere with the required response. This shows, on the one hand, that attention to an object is not sufficient to ensure the processing of all its feature dimensions. On the other hand, if an attended object is a target, task-irrelevant feature dimensions are apparently processed. Therefore, even task-irrelevant target features could give rise to priming effects, as has been reported at least for color (Kristjánsson, 2006).
These considerations support a strong version of a priming account of contingent capture: Contingent capture could emerge as a result of the repetition of target features and, hence, of the priming by these features even when they are not, per se, task-relevant, and therefore not attended to. In the present study, we were interested in whether or not empirical evidence can be found for such a radical alternative explanation of contingent-capture effects. We were interested in the extent to which contingent capture (validity) effects would emerge based on the mere repetition of uninstructed, and in this sense task-irrelevant, target features. To shed light on this question, we conducted a set of experiments in which the target was defined by a particular feature that participants were instructed to search for. Crucially, the target also had a second feature that could, in principle, be used to locate it, but this feature was not task-relevant (i.e., it was not pointed out to the participants in the instructions). Prior to the targets, singleton cues appeared that either had the relevant searched-for target feature or a task-irrelevant target feature. The research question was whether cues with primed task-irrelevant target features would capture attention, and if they did, whether they would account for the full contingent-capture effect. Because priming effects, and the effects of selection history more broadly, can accumulate over several trials (Kristjánsson & Campana, 2010, for a review), we were interested not only in the effects that emerged from one trial to the next (Exps. 3 to 5), but also in whether irrelevant-feature cues that were primed in each trial, because they always had the same (but uninstructed) target-discriminating feature as all of the targets, would lead to overall validity effects (Exps. 1 and 2).
Experiment 1
In Experiment 1, participants searched for two possible target colors. These task-relevant features formed the top-down feature set (i.e., both these colors were task-relevant, as participants were instructed to search for these colors). Participants were informed that one target would be presented in each trial in one of two possible colors (e.g., red or blue). Which target color was realized in a trial was randomly chosen. In addition, each target consisted of a specific type of disk–ring composite,Footnote 3 by which the target could in principle also be discriminated from all other items in the target display: Each of several gray disks in the target display was surrounded by a colored ring, only one of which was of the target color (see Fig. 1). Importantly, each color ring either fit neatly around its gray disk or was loosely fitted around the disk. In the latter case, there was a small gap between the inner disk and the outer color ring (henceforth, “gap items”). For half of all participants, the target was neatly fitting throughout the experiment and the distractors were gap items, and for the other half of the participants, the target was a gap item in each trial throughout the experiment, and the distractors in the target display were all neatly fitting. This disk–ring composite feature was clearly visible (and, as Exp. 3 showed, it could also be used successfully to search for the target when participants were instructed to do so). However, the target’s unique composite feature was not instructed as a to-be-searched for feature in Experiments 1 and 2, so it was deemed task-irrelevant. There was no necessity for the participants to incorporate the unique disk–ring composite feature into their top-down set of searched-for target features, because the colors also did the job.

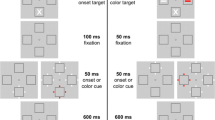
Example sequences of events that illustrate the general procedure in each trial in Experiments 1 and 2. See the online publications for the color version of the figure. In Experiment 1, participants were instructed to search for two target colors (e.g., red and green; here, a red target is depicted). In Experiment 2, they searched for just one color. The illustrated trials depict sequences with a valid cue of a top-down matching color (two sequences at the top) and of a top-down nonmatching color (two sequences at the bottom). The disk–ring composite type of the target was unique. For half of the participants, the targets were gap items among neatly fit distractors in the target display. This is depicted in the present case. (For the other half of participants, the targets were neatly fit items among gap distractors. This is not depicted.) The composite of the cue either could match the composite of the target (target-similar composite cues) or it could differ form the composite of the target (target-dissimilar composite cues). Note that when the cue had a target-dissimilar composite, the cue was not a singleton in the composite dimension, only in the color dimension. This was different in Experiment 3 (cf. Fig. 5)
Critically, with this setup, it was possible to assess the validity effects by cues that had the task-relevant feature (color) and by cues that had the task-irrelevant feature (disk composite). If merely seeing the feature in the target primed the attentional system so that the corresponding feature captured attention, validity effects should emerge regardless of whether the corresponding features were relevant or irrelevant to the task at hand. If, however, the instructions and the top-down search setting were critical, only instructed, task-relevant target feature cues should capture attention. There should be no (additional) validity effect from cues that had the task-irrelevant target feature as well.
In addition to this measure of major importance, the design allowed us to assess intertrial priming of color from trial n−1 to trial n. We used two colors, and hence we were able to assess whether validity effects were stronger when the cue in the given trial n had the same color as the target in trial n−1 (Folk & Remington, 2008). This measure allowed us to assess whether we found an indication of the influence of intertrial priming on the contingent-capture effects among at least the instructed features, thereby replicating the results of prior studies (Folk & Remington, 2008).
Method
Participants and general considerations regarding power and sample size
Twenty-four participants took part (16 male, eight female; Mage = 27.5 years, SDage = 8.5 years). With this sample size, the experiment should be well powered, given the typically large sizes of contingent-capture effects. Indeed, studies reporting contingent-capture effects have typically employed sample sizes no larger than the sample size of the present study (e.g., Carmel & Lamy, 2014; Eimer et al., 2009; Goller & Ansorge, 2015). Certainly, smaller effects of the task-peripheral features could be overlooked by a single study. We therefore replicated Experiment 1’s results in Experiment 2. As an additional measure to deal with power issues, we will report Bayes factors (Marsman & Wagenmakers, 2016), which allow for quantifying the evidence in favor of the null hypothesis in case that there is no evidence of capture effects for primed but task-irrelevant features.
The participants had normal or corrected-to-normal vision, as well as normal color vision as assessed by Ishihara color plates. Informed consent was obtained from each participant at the beginning of the experiments. No approval from the ethics committee was needed for this type of study, according to institutional guidelines at the University of Vienna.
Apparatus and stimuli
All stimuli were presented on an LCD monitor, with a refresh rate of 60 Hz and a resolution of 1,280 × 1,024 pixels. All stimuli were presented against a gray background. Stimuli were centered at a distance of 4.9° of visual angle from the screen center, where a small fixation cross (width and height ~ 0.5°) was presented at the beginning of each trial. The stimuli consisted of circles with a diameter of 2.0°. At each position, an inner disk (diameter: 1.0°) in dark gray (CIE-Lab color coordinates: 35.2, 2.4, − 8.7) was surrounded by a colored ring. The colors used were yellow (CIE-Lab: 56.5, − 17.2, 44.8), green (CIE-Lab: 53.2, − 59.0, 34.0), red (CIE-Lab: 56.8, 72.5, 50.4), blue (CIE-Lab: 52.2, 75.5, − 156.9), turquoise (CIE-Lab: 55.4, − 33.2, − 24.2), and pink (CIE-Lab: 56.9, 80.8, − 68.3). Importantly, there could be a small gap (0.25°, filled in with the background color, CIE-Lab: 55.0, − 2.3, − 14.4) between the inner circle and the colored ring, which was then filled in with the background color. These were the “gap items.” For another set of stimuli, this gap was not present—that is, the inner circle in dark gray fit neatly with the colored ring. These were the “neatly fit” stimuli.
In the cueing display as well as in the target display, four such disk–ring composite stimuli appeared at four display locations, at an upper left, an upper right, a lower left, and a lower right location (see Fig. 1). Only in the target display, four tilted (i.e., two leftward- and two rightward-tilted) Ts (width and height: 0.5°) were presented in light gray (CIE-Lab: 78.9, − 4.0, − 19.5). The Ts always appeared inside of the dark-gray center of each of the disks, so that there was a clear contrast between the dark gray and the light gray of the Ts.
Design
Participants worked through 1,152 trials in nine blocks of 128 trials each. After each block there was a self-paced break, and participants were informed about their overall performance in the experiment up to that point. Participants were instructed to search for a color target that could have one of two possible colors (this was the information about the target that was given in the instructions), and they had to respond to the orientation of a tilted T inside the target item. They had to press the j key on a standard keyboard with their right index finger when the T was tilted rightward, and they had to press the f key with their left index finger when the T was tilted leftward. In addition, in the target displays the target could be a gap item and the three distractors would be neatly fit (half of the participants), or the target could be neatly fit and the distractors would be gap items (the other half of the participants). The specific disk–ring composite of the target was not pointed out to the participants in the instructions, but it was present throughout all trials.
The succession of events in each trial was as follows: Each trial started with a fixation cross. After 1 s, the cueing display was presented (see Fig. 1). In the cueing display, four stimuli were presented. One of these items was always a cue—that is, it was presented as a feature singleton (with a unique color relative to the other, nonsingleton items in the cueing display, and in half of the trials also with a unique disk–ring composite—here, the same composite as the target). The cue either had one of the target colors (targets were always in one of two specific colors; see above) or had a nontarget color. The other items in the cueing display were always presented in a color that was different from that of the cue. The target colors and the color of the nonmatching cue were chosen from the three colors red, green, and blue, which were balanced across participants. When the target colors were red and green, the color of the nonsingletons in the cueing display was yellow in each trial. When the target colors were red and blue, the nonsingletons in the cueing display were pink, and when the target colors were blue and green, the nonsingletons in the cueing display were turquoise. In each case, the two remaining nonsingleton colors were presented as distractors in the target display, to ensure that the target was not a color singleton. After 50 ms the cueing display disappeared and a blank interval was presented, in which only the fixation cross remained on the screen for 200 ms. After this time interval, the target display was presented for 200 ms. In the target display, four items were presented, only one of which had one of the searched-for target colors (i.e., this item was the target). Additionally, the target was a specific and unique disk–ring composite in its display. The other items in the target display were presented in two different colors—that is, one color was used for two items, but the target was not a singleton in the color dimension, because three colors were used for the four items. Within each of the items presented in the target display was a T, tilted left- or rightward. In two of the items, the T was always tilted leftward, and in the other two items the T was tilted rightward. Participants had to respond to the orientation of the T that appeared inside the target (as described above). After the target screen disappeared, a blank screen was presented, and participants had to respond within a time window of 2 s. If participants failed to respond, or if their response was incorrect, the corresponding trial counted as an error trial. Trial-wise feedback was only given during the practice phase of 20 trials, at the beginning of the experiment. The experiment started when participants had responded correctly at least 70% of the time in the practice trials. If this was not achieved, the practice was repeated.
The target and the cue appeared in each trial randomly and unpredictably at one of the four possible locations. The steps of the variables cue location, target location, cue color (target-matching vs. target-nonmatching color), and cue composite (target-similar vs. target-dissimilar composite) were combined orthogonally. Trial type was selected pseudo-randomly on each trial, but each condition or trial type appeared equally often.
Results
Reaction times
Reaction times (RTs) were measured as the time interval between the onset of the target display and the participant’s button press to the orientation of the T inside the target. The data of one participant were removed because her or his error rate exceeded 40%. For the remaining participants, errors (15.2%) were removed, and RTs that deviated by more than 2.5 SDs (2.2% of the trials) from the mean of the corresponding condition were also removed. The remaining individual mean correct RTs were subjected to an analysis of variance (ANOVA), with the within-subjects variables cue validity (valid vs. invalid), cue color match (matching vs. nonmatching color), and cue composite similarity (target-similar vs. target-dissimilar composite). We found a main effect of cue validity, F(1, 22) = 17.79, p < .001, ηp2 = .45, and an interaction between cue validity and color match, F(1, 22) = 49.03, p < .001, ηp2 = .69. Other effects were not significant (all nonsignificant ps > .24, all nonsignificant Fs < 1.48). Post-hoc t tests confirmed a cueing effect when the cue color matched the top-down set, 70 ms, t(22) = 6.30, p < .001. When the cue color did not match the top-down set, there was no significant cueing effect, − 9 ms, t(22) = − 1.2, p = .21. See Fig. 2.

Results of Experiment 1. Depicted on the left are the mean reaction times (RTs) and error rates in the valid and invalid conditions, with color-matching and color-nonmatching cues. The filled circles correspond to the data with target-similar composite cues, and the nonfilled circles correspond to the data with target-dissimilar composite cues. Error bars represent within-subjects standard errors (Cousineau, 2005). Depicted on the right are the cueing effects (RT in invalid minus RT in valid cue conditions) by matching color cues. Compared are the cueing effect when the target in trial n–1 had the color of the cue in trial n versus the cueing effect when the target in trial n–1 did not have the color of the cue in trial n
Since the results showed no significant effect of the variable composite similarity, we also obtained an estimate of the extent that our data favored the null over the alternative hypothesis. To this end, we compared the overall validity effect (invalid minus valid RT) for target-similar composite cues to the overall validity effect for target-dissimilar composite cues by use of a Bayesian paired test using the JASP software (Marsman & Wagenmakers, 2016). This analysis with the default setting of 0.707 for the scale parameter of the Cauchy prior yielded a Bayes factor (BF10) of 0.73, suggesting that the null was more likely than the alternative hypothesis, albeit only slightly (i.e., the null hypothesis was 1.37 times more likely than the alternative hypothesis).
Errors
The percentage of errors in each condition was arcsine-transformed and subjected to an ANOVA like the one above. The results mirrored those for RTs. There was a main effect of cue validity, F(1, 22) = 23.93, p < .001, ηp2 = .52, and also an interaction between cue validity and color match, F(1, 22) = 26.10, p < .001, ηp2 = .54. Post-hoc t tests showed a large validity effect with color-matching cues (error rate invalid minus valid) of 9.4%, t(22) = 6.16, p < .001, but not with color-nonmatching cues, 0.3%, t(22) = 0.33, p = .75. Other effects were not significant (all nonsignificant ps > .22, all nonsignificant Fs < 1.59).
We also computed BF10 as described for the RTs, by comparing the overall validity effects for target-similar composite cues to the overall validity effects for target-dissimilar composite cues. This led to a value of 0.21, thus, favoring the null over the alternative hypothesis (the null hypothesis is four to five times more likely than the alternative).
Intertrial priming by color
For the color-matching conditions, we collected correct RTs on trials in which the target color in the preceding trial n–1 matched the cue color in trial n, and in trials in which the target color in trial n–1 did not match the cue color in trial n. In each condition, RTs deviating by more than 2.5 SDs from the mean were removed, and means were then computed for each participant (by averaging across the data from the target-similar composite and the target-dissimilar composite cues), so that a paired t test could be calculated between matching color cue conditions in which the target color in trial n–1 was similar versus dissimilar to the cue color in trial n. This t test was significant, t(22) = 2.74, p < .05. When the target color in trial n–1 was similar to the matching cue’s color in a trial n, the validity effect from these cues was 18 ms larger than when the target color in trial n–1 was different from the matching cue’s color in trial n. However, the validity effect of the matching cues was still significant if the cue color was not primed by the color of the target in trial n–1 (p < .01). See also Fig. 2.
Because priming can occur at an object level (Kristjánsson, Ingvarsdóttir, & Teitsdóttir, 2006) and it is, therefore, possible that only cues that matched the target with respect to all of its features would be primed, we checked whether the priming effects for composite-similar and composite-dissimilar cues differed. There was no statistically significant difference (p = .85). If anything, the priming effects were larger when the cue was dissimilar to the composite of the target (19 ms; p < .05), as compared to when the composite of the cue was similar to that of the target (17 ms; one-tailed p = .055).
We performed the same analysis with the error rates. The cueing effect in error rates was numerically also larger when the target in trial n–1 was of the same color as the matching cue in trial n (10%), as compared to when the target color in trial n–1 was dissimilar to that of the matching cue in trial n (9%). This difference was, however, far from significant, t(22) = 0.16, p = .87. No difference was apparent between the priming effects of target-similar versus target-dissimilar composite cues (p = .25).
Discussion
The results of Experiment 1 showed that cues that had the task-irrelevant target feature did not capture more attention than cues that did not have this feature. This was true even though the corresponding feature was clearly visible and was presented in the target in each trial (and as we will see, Exp. 3 would show that the corresponding feature could in principle be used to locate the target). A regular contingent-capture effect was found for the feature that participants were instructed to search for. Cues that had the target colors led to a strong validity effect, whereas cues that did not have the target color did not.
Together, the results are at odds with the strong version of the priming account of the contingent-capture effect. If attentional capture by the cues’ features were present based on feature repetition of any target features without instructions directed at the corresponding feature, we should also have found substantial capture effects by cues that had the task-irrelevant feature. This was not the case. A complication of Experiment 1 might be that the task-irrelevant target feature was realized in a different dimension (composites) than the instructed target feature (color). Because a change between dimensions is costly in terms of processing efficiency (Found & Müller, 1996; Müller, Heller, & Ziegler, 1995), a priming effect for the task-irrelevant target features might have been prevented by this delay (e.g., the corresponding information might have been available too late to affect capture by the cues similar to a task-irrelevant target feature). We will get back to this important point in the final two experiments.
Interestingly, we also found evidence of intertrial priming by the instructed task-relevant target features in the present Experiment 1. Attentional validity effects were stronger with cues that had the target color of trial n–1 than with cues that had the other target color. This result replicates the findings of some prior studies, and it suggests that intertrial priming by instructed target features is a factor in contingent-capture studies (Folk & Remington, 2008).
Experiment 2
In Experiment 2, we aimed at a conceptual replication of Experiment 1’s task-irrelevant conditions, but this time we used an electrophysiological measure of attentional capture in addition to the behavioral RTs and error rates (e.g., Eimer et al., 2009). This was done in order to check whether we had overlooked weaker capture of attention by the target-similar, and thus primed but task-irrelevant, composite cues, because their capture effects could be more short-lived and transient than the capture effects by color (cf. Theeuwes, 2010). Thus, in Experiment 1, at the time of the overt response, the capture effect by the target-similar composite cues could have already been past. However, a cue-elicited lateralized event-related potential could still demonstrate a more fleeting capture effect. This was tested in Experiment 2.
Method
Participants
Twenty-four participants took part (12 males, 12 females; Mage = 23.54 years, SDage = 1.77 years). They had normal or corrected-to-normal vision, as well as normal color vision as assessed by Ishihara color plates. Informed consent was obtained from each of the participants at the beginning of the experiment.
Apparatus, stimuli, and procedure
The experiment was conducted with a CRT monitor with a refresh rate of 60 Hz and a resolution of 1,280 × 1,024 pixels. The stimulus design was similar to that in Experiment 1, except for the following changes: Participants were instructed to search for only one color (either red, green, or turquoise, balanced across participants). The colors used were red (CIE-Lab: 64.2, 74.4, 59.1), green (CIE-Lab: 65.4, − 70.8, 55.0), turquoise (CIE-Lab: 66.2, − 36.4, − 35.3), pink (CIE-Lab: 63.1, 79.9, − 86.0), and yellow (CIE-Lab: 64.8, − 22.9, 57.6). All stimuli were presented against a light gray background (CIE-Lab: 66.8, − 6.4, − 24.1) and had a dark gray inner circle (CIE-Lab: 49.4, − 7.2, − 19.1).
As in Experiment 1, the cue was always a color singleton. In color-nonmatching cue conditions, the cue appeared in a nontarget color, and the nonsingletons in the cueing display appeared in a different nontarget color. The target was always a nonsingleton color—that is, in each target display, three colors were presented. The target appeared in the target color, and the three additional items in two different colors (see also Fig. 1).
Electroencephalogram—Data recording and preprocessing
An electroencephalogram (EEG) was recorded at 1,000 Hz by a full-band DC-EEG system (neuroConn GmbH, Ilmenau, Germany), with active electrodes (Brain Products, actiCAP system) mounted in a cap (EASYCAP GmbH, Herrsching, Germany) at the 64 positions of the 10–10 system. Electrode impedances were kept below 10 kΩ (Kappenman & Luck, 2010). The ground electrode at AFZ served as an online reference. The signal was re-referenced offline to the average of both mastoids and 40-Hz low-pass filtered with a finite impulse response filter (FIR), with a cutoff frequency of 45 Hz (− 6 dB) and a transition bandwidth of 10 Hz.
The epoched EEG data were subjected to an artifact rejection procedure for a time interval from − 100 ms (prior to cue onset) to 400 ms (following cue onset). Vertical electrooculogram (VEOG) epochs were excluded if values exceeded ± 60 μV. The difference between two electrodes, one at each of the outer canthi, was used for the horizontal EOG (HEOG). Such epochs were excluded if the values exceeded ± 30 μV. Furthermore, we excluded signals from the event-related potentials’ (ERPs’) average separately at each electrode if the signal exhibited very low activity (less than a 0.5-μV difference between subsequent samples for a time period of at least 500 ms), very high signal changes (more than 50 μV/ms), or values exceeding ± 80 μV. Also, baseline correction was performed for each entire epoch with respect to the 100-ms interval before cue onset. N2pc components to the cue displays were quantified on the basis of the mean amplitudes obtained in a 190- to 270-ms (see Ansorge et al., 2011; Grubert & Eimer, 2013) time window after stimulus onset at lateral posterior electrodes PO7 and PO8. ERPs were computed, and the analysis of the ERP data was based only on trials with a correct behavioral response.
Results
Behavioral results
Reaction times
Errors (11.4%) and RTs deviating by more than 2.5 SDs from the mean of the corresponding condition were removed (2.2% of the trials). The resulting mean correct RTs were fed into a repeated measures ANOVA with the variables cue validity, cue color match, and cue composite similarity. This led to a main effect of color match, F(1, 23) = 5.67, p < .05, ηp2 = .20, and a main effect of cue validity, F(1, 23) = 78.39, p < .001, ηp2 = .77. It also led to an interaction between the variables color match and cue validity, F(1, 23) = 113.84, p < .001, ηp2 = .83. Post-hoc t tests showed a strong cueing effect when the color of the cue was top-down matching, 147 ms, t(23) = 11.23, p < .001, but not when the cue was not matching, 5 ms, t(23) = 0.61, p = .55. Other effects in the ANOVA were not significant (all nonsignificant ps > .54, all nonsignificant Fs < 0.39). See also Fig. 3.

Behavioral results of Experiment 2. Depicted are the mean correct reaction times (RTs) and error rates in the valid and invalid conditions with color-matching and color-nonmatching cues. The filled circles correspond to the data with target-similar composite cues, and the nonfilled circles correspond to the data with target-dissimilar composite cues. Error bars represent within-subjects SEs (Cousineau, 2005)
Again, the BF10 was computed as in Experiment 1, by comparing the overall validity effect with target-similar composite cues to the overall validity effect with target-dissimilar composite cues. The result was 0.18, providing evidence in favor of the null hypothesis (i.e., the null hypothesis in Exp. 2 was about five times more likely than the alternative). We also collapsed the data from Experiments 1 and 2 and computed a Bayesian test again with the collapsed data, by comparing the overall validity effect with target-similar composite cues to the overall validity effect with target-dissimilar composite cues. BF10 was 0.31, again favoring the null over the alternative hypothesis.
Errors
Error rates were arcsine-transformed and fed into an ANOVA like the above. This led to a main effect of cue validity, F(1, 23) = 88.96, p < .001, ηp2 = .80, and to an interaction between cue validity and color match, F(1, 23) = 61.23, p < .001, ηp2 = .73. There was a strong validity effect with color-matching cues (12.4%), t(23) = 10.57, p < .001, but no validity effect with color-nonmatching cues (0.3%), t(23) = 1.02, p = .32. Other effects in the ANOVA were not significant (nonsignificant Fs < 2.36, ps > .14).
BF10 was 0.37, again computed by comparing the overall validity effect with target-similar composite cues to the overall validity effect with target-dissimilar composite cues. We again collapsed the data over Experiments 1 and 2, and performed the Bayes factor analysis again. The resulting BF10 was 0.22.
Electroencephalogram results
Event-related potentials
Mean ERP amplitudes of correct trials were subjected to a repeated measures ANOVA, with the within-subjects variables cue color match (matching vs. nonmatching color), cue composite similarity (target-similar vs. target-dissimilar composite), and hemisphere (contralateral vs. ipsilateral); see also Fig. 4. We found a main effect of cue match, F(1, 23) = 6.38, p = .019, ηp2 = .22, as well as an interaction between cue color match and hemisphere, F(1, 23) = 27.73, p < .001, ηp2 = .56. In the matching color condition, the mean ERP amplitude was significantly smaller contralateral (1.51 μV) than ipsilateral (2.17 μV), t(23) = − 3.38, p = .003. This result reflects a typical N2pc. In the nonmatching color condition, the opposite was the case: The mean ERP amplitude was significantly higher contralateral (1.44 μV) than ipsilateral (1.13 μV), t(23) = 3.61, p = .002. Other effects in the ANOVA were not significant (nonsignificant Fs < 2.11, ps > .16).

Event-related potentials (ERPs) for color-matching (left panel) and color-nonmatching (right panel) cues in Experiment 2. The graphs show the EPRs at electrode positions PO7/PO8, in microvolts. Negative values are plotted upward. The light lines illustrate the ipsilateral ERPs, and the dark lines the contralateral ERPs. The two smaller graphs at the bottom of each panel illustrate the ERPs split for target-similar (in the left panels) and target-dissimilar (in the right panels) composite cues
To confirm that there truly was no effect of composite similarity, we subjected the amplitude differences (contra – ipsilateral) with target-similar composite cues and with target-dissimilar composite cues to a paired Bayesian test. The BF10 was 0.26, thus confirming the null hypothesis.
Discussion
Experiment 2 replicated the major results of Experiment 1. We found no indication of attentional capture by cues primed with task-irrelevant uninstructed target features. Singleton-color cues that had a composite similar to the target did not capture more attention than did singleton-color cues that had a composite dissimilar from the target. This was true despite the fact that the target had the corresponding composite in each and every trial, and target-similar composite cues would thus have been primed throughout the blocks. We did not observe a behavioral cueing effect, nor did the analysis of the ERP data indicate attentional capture by composite-similar cues.
In contrast to this negative result, we replicated the contingent-capture effect by color, and also, in the ERPs, found a cue-elicited N2pc with color-matching cues. With nonmatching color cues, we found a reversed effect (i.e., more negativity ipsilateral to the cue), probably indicating attentional suppression (e.g., Hickey, Di Lollo, & McDonald, 2009).
Experiment 3
In Experiments 1 and 2, we showed that a cue’s validity effect based on its similarity to a target’s composite type does not emerge when participants search for colors. In Experiment 3, we changed the instructed roles of the specific composite of the target and the color of the target, so that the instructions informed participants not about the target colors, but only about the specific composite of the target. This was done (1) to demonstrate that a contingent-capture effect based on the composite of the target could be obtained when participants were instructed to search for this feature, and (2) to test whether color as a task-irrelevant feature would foster more evidence of capture effects by intertrial priming.
To note, color is a more dominant perceptual feature than others (e.g., shapes; cf. Theeuwes, 1992). Color has also repeatedly featured in demonstrations of intertrial priming of capture, including the first experiment of the present study. It may thus well be that more evidence for (intertrial) priming of capture would emerge with color as a task-irrelevant target feature (i.e., a feature not contained in the instructed top-down set). Just as in Experiment 1, we again used two different target colors, to check for the effects of selection history on capture by color cues.
Method
Participants
Thirty new participants took part (24 female, six male; Mage = 22.2 years, SDage = 4.8 years). Two participants were removed from the analysis because they had error rates exceeding 40%.
The experimental setup was identical to that in Experiment 1, with the following exceptions: (1) Participants were now instructed to search for the specific target composite (e.g., for the gap item vs. the neatly fitting item). Which composite was used for the target was fixed throughout all trials but was balanced across participants. (2) The composite-matching singleton cues were presented among composite-nonmatching nonsingletons in the cueing display, and the composite-nonmatching singleton cues were presented among composite-matching nonsingletons in the cueing display (see Fig. 5). This was different from Experiments 1 and 2, in which target-dissimilar composite cues were presented among other target-dissimilar composite items. With the procedure in Experiment 3, we were able to check whether participants adopted a singleton detection mode (Bacon & Egeth, 1994) for the “odd composite” rather than for a specific target composite. If participants adopted a singleton search mode, composite-nonmatching cues were expected to also capture attention, because they were presented as singletons in the composite feature dimension.

Example sequences of events that illustrate the general procedure in each trial of Experiment 3. See the online publication for the color version of the figure. Participants were instructed to search for a specific composite type as their target. In the depicted example, participants would be instructed to search for the gap items among neatly fitting composite items. (The other half of the participants, not depicted, were instructed to search for neatly fitting composite items among gap items.) The illustrated trials depict a sequence with a valid cue of a top-down matching composite type (the two sequences at the top) and of a top-down nonmatching composite type (the two sequences at the bottom). Notice that in Experiment 3, when the cue had a nonmatching composite, the cue was also a singleton in the composite dimension—that is, the other three items had the matching composite, in this case. This was different from in Experiments 1 and 2 (cf. Fig. 1). In addition, the targets were always presented in one of two possible colors (here, the target is always depicted in red). This fact was not pointed out to participants in the instructions. Cues could be either similar to one of the target colors or dissimilar to both of the target colors
Apart from these manipulations, the design was identical to that of Experiment 1, and so were the predictions. Intertrial priming of capture by task-irrelevant target features should show up as stronger validity effects for target-similar color-singleton cues with a color similar to the last seen (n–1) target than for cues with a color dissimilar from the last target. Unless participants now incorporated the task-irrelevant and uninstructed target colors into their top-down sets after they realized that colors could be used to search for the targets, we expected to see no capture by the task-irrelevant singleton-cue colors, independent of intertrial priming (i.e., independent of whether or not a target-similar color cue was of the same color as its preceding target).
Results
Reaction times
RTs were collected in each of the conditions. Errors (18.2%) and outliers that deviated by more than 2.5 SDs from the mean of the corresponding condition (2.2%) were excluded, and the correct mean RTs were subjected to an ANOVA with the variables cue validity, cue composite match, and cue–target color similarity. This analysis led to a highly significant main effect of cue validity, F(1, 27) = 18.15, p < .01, ηp2 = .40, as well as two-way interactions between cue validity and composite match, F(1, 27) = 35.93, p < .001, ηp2 = .57, and cue validity and color similarity, F(1, 27) = 10.90, p < .01, ηp2 = .29. There was also an interaction between all variables, F(1, 27) = 4.45, p < .05, ηp2 = .14.
Post-hoc t tests revealed a strong validity effect when the composite was top-down matching and the cue color was similar to a target color, 73 ms, t(27) = 6.19, p < .001. There was also a strong, albeit smaller, validity effect when only the composite was matching but the color of the cue was dissimilar to both target colors, 43 ms, t(27) = 4.79, p < .001. There was no effect when the color of the cue was similar to a target color but the composite was nonmatching, − 5 ms, t(27) = 0.56, p = .58. The cueing effect was significantly reversed when the color of the cue was dissimilar to the target colors and the cue composite was nonmatching, − 20 ms, t(27) = − 3.41, p < .01. See Fig. 6.

Results of Experiment 3. Depicted on the left are the mean correct reaction times (RTs) and error rates in valid and invalid conditions with composite-matching and composite-nonmatching cues. The filled circles correspond to the data for target-similar color cues. The nonfilled circles correspond to the data for target-dissimilar color cues. Error bars represent the within-subjects SEs (Cousineau, 2005). Depicted on the right are the cueing effects (RT in invalid minus RT in valid cue conditions) by target-similar color cues. Compared are the cueing effects when the target in trial n–1 had the color of the cue in trial n versus the cueing effect when the target in trial n–1 did not have the color of the cue in trial n
Errors
The errors in each condition were counted and the percentages of errors were arcsine–square root transformed. The resulting data were subjected to an ANOVA like the one above. With error rates, we found main effects of composite match, F(1, 27) = 18.28, p < .001, ηp2 = .40, and of cue validity, F(1, 27) = 31.39, p < .001, ηp2 = .54. Furthermore, an interaction emerged between cue validity and composite match, F(1, 27) = 23.32, p < .001, ηp2 = .48. This interaction indicated a contingent-capture effect by the cue’s composite match. There was a strong cueing effect with composite-matching cues, 8.2% t(27) = 5.95, p < .001, but this effect was absent with composite-nonmatching cues, − 0.4%, t(27) = 0.16, p = .88. Other effects were not significant (all nonsignificant Fs < 3.32, all nonsignificant ps > .08).
Intertrial priming by color
We next computed the mean correct RTs in each condition in which the color of the cue was similar to that of one of the targets (i.e., for cues that would have been top-down matching cues in Exp. 1). We then split up these data according to whether or not the (potentially matching) cue was of the same color as the target in trial n−1. For this purpose, the data were averaged over the two steps of the variable composite match, and we compared validity effects when the target color in trial n−1 was the cue color in trial n with those trials in which the target color in n−1 was not the cue color in trial n. Unlike in Experiment 1, we found no evidence of intertrial priming of capture by color cues, 2 ms, t(27) = 0.27, p = .79. The BF10 of a corresponding Bayesian test was 0.25, indicating that the null hypothesis was four times more likely than the alternative. Also, when the results were analyzed separately for matching and nonmatching composite cues, no evidence of priming was found (ps < .28, BF10s < 0.35). We conducted the same analysis with error rates. This analysis also showed no trend toward a significant result, t(27) = 0.10, p = .92. The corresponding BF10 was 0.22, again indicating that the null hypothesis was about four to five times more likely than the alternative hypothesis. In addition, when the data were analyzed separately for matching and for nonmatching composite cues, no significant effects were obtained (ps < .67, BF10s < 0.22).
Discussion
Crucially, the results of Experiment 3 showed that participants were able to set up a top-down search set for the particular composite type of the target, because this led to a contingent-capture effect from target-matching composite cues. Thus, singleton capture can be ruled out. This time, we also found a capture effect by the cues carrying a task-irrelevant, not instructed, but potentially helpful target feature: color. Interestingly, we nonetheless observed no intertrial priming of capture by the target-similar color cues from one trial to the next. Whether a target-similar color cue was or was not of the same color as the target in trial n–1 did not matter for the strength of the capture effect by the target-similar cues. These results point to a different origin of the color-based cueing effects when color was irrelevant to the task (as in Exp. 3) versus when it was task-relevant (as in Exp. 1). Maybe the color-based cueing effect in Experiment 3 reflected an implicit form of learning of color–target associations that is different from priming and more akin to statistical learning (cf. Lanthier, Wu, Chapman, & Kingstone, 2015). Such implicit attentional guidance might not lead to the incorporation of the target-associated feature into the top-down set of explicitly instructed relevant target features. For example, whereas instructed relevant target features could be actively maintained in a working memory representation, the same might not be true of an implicitly learned target-feature association that could be represented in some form of perceptual memory (cf. Hamann & Squire, 1997; Jiang, 2018).
What is also remarkable is that the cueing effects reversed with cues that did not have either the target’s composite or color. We think that, in the present context, the most likely explanation for this is that the nonsingletons in the cueing display had the searched-for target feature, in the case of a nonmatching composite cue (see Fig. 5), and these items, therefore, captured attention to some extent. However, other explanations, such as an origin of the effect in terms of object-file updating in working memory, are conceivable (e.g., Carmel & Lamy, 2014; see also Schoeberl, Ditye, & Ansorge, 2018). Another possibility is that the effect was due to the inhibition of recently visited distractors (see Campana & Casco, 2009).
Experiment 4
Experiments 1–3 showed that validity effects were found with cues that had features participants were instructed to search for. There was little (Exp. 3) or no (Exps. 1 and 2) enhancement of the validity effects when the cues had task-irrelevant target features, making it unlikely that the contingent-capture effect could be explained by priming through uninstructed features. However, as we briefly discussed following Experiment 1, the task-irrelevant target features in Experiments 1–3 were defined in a different feature dimension than the instructed features. It is therefore possible, for example, that participants may have simply down-weighted the corresponding feature dimension and disregarded the whole dimension, because they were told that a different feature dimension was task-relevant (cf. Found & Müller, 1996; Müller et al., 1995). In Experiment 4, we therefore tested to what extent the results we obtained in Experiments 1–3 would generalize to conditions in which both the task-relevant instructed target features and the task-irrelevant target features were defined in the same feature dimension. Participants now viewed four crosses. One of these crosses was the target, and participants responded to its orientation (i.e., whether the target was tilted or straight). In each of the crosses, two of the four arms were colored. Participants were instructed that the target cross always would contain one of two possible colors (blue and green in Exp. 4a, and red and brown in Exp. 4b); in each trial, the target also contained one of two fixed, task-irrelevant colors that participants were not instructed to search for (red and brown in Exp. 4a, and blue and green in Exp. 4b). That is, the target always had one arm in an instructed, searched-for color, and one arm in a noninstructed, task-irrelevant color. After the instructions, participants knew only the pair of task-relevant colors, but not the pair of task-irrelevant colors. Prior to each target display, cueing displays with a color cue were presented. The cues were either blue, green, red, or brown and were primed by the preceding target color in half of the trials.
If cueing effects were based on feature priming, regardless of the instructions, we expected all primed color cues to capture attention, regardless of the instructions. If the cues captured attention contingent on their match to the instructed top-down search settings, we expected only cues in the instructed target colors to capture attention.
In addition, the experimental setup allowed us to investigate the impact of intertrial priming from one trial to the next, by instructed target features as well as by noninstructed, task-irrelevant target features.
Method
Participants
In all, 21 participants (13 female, eight male; Mage = 21.62 years, SDage = 2.62 years) took part in Experiment 4a (instructed target colors green and blue), and 23 participants (16 female, seven male; Mage = 20.96 years, SDage = 2.18 years) took part in Experiment 4b (instructed target colors red and brown).
Stimuli and procedure
All stimuli were presented on an LCD monitor with a refresh rate of 60 Hz and a resolution of 1,280 × 1,024 pixels. The stimuli were presented against a light gray background (CIE Lab color coordinates: 105, − 1.1, − 28.7). The stimuli were crosses in gray (CIE Lab coordinates: 65.9, − 3.7, − 15.5) in which two of the four arms appeared in color. The colors used for this experiment were red (CIE Lab: 66.4, 89.6, 82.1), green (CIE Lab: 67.6, − 75.2, 47.8), blue (66.8, 41.2, − 117.9), violet (65.6, 95.8, − 80.3), brown (66.0, 0.3, 54.5), yellow (118.0, − 23.0, 101.0), and black (0, 0, 0). In principle, targets could be localized by searching either for red and brown or for blue and green, because one arm of the target cross was in each case red or brown, and another arm was always blue or green. The distractor crosses in the target display also had two arms that were colored. The colors used for this purpose were yellow, black, and violet. The colors used for the singleton cues in the cueing display were red, green, blue, and brown. Thus, all of the cues were target-similar, but only half of them were task-relevant (instructed), whereas half of them were task-irrelevant (uninstructed). The distractors in the cueing display were violet in each trial.
For the sequence of events on each trial, see Fig. 7. Participants viewed a fixation cross at the beginning of each trial. After 1 s, a cueing display was presented for 50 ms, in which four colored rings (line width: 0.2°, diameter: 3.2°) were presented at 4.8° eccentricity in the corners of a virtual square centered at screen center—that is, at an upper left, upper right, lower left, and lower right location. After 50 ms, the cueing display was turned off and an interval display was presented for 100 ms, in which only the fixation cross remained on the screen. After this, the target display was presented. In that display, four crosses were presented (the crosses were composed of two orthogonal bars, 2.1° × 0.7°). Each of the crosses was centered at one of the same positions as the colored rings. In each trial, two of the crosses were tilted and two were straight. Participants’ task was to indicate whether the target cross (the cross in which the target color appeared: blue or green for Exp. 4a, red and brown for Exp. 4b) was tilted (f key) or straight (j key). The target display was presented for 300 ms. After this, the target display was turned off and a blank display was presented in which only the fixation cross remained on the screen. Now participants had 1.5 s to respond. After each correct response, there was a short interval of 500 ms before the next trial started. If participants did not respond correctly or were too slow, they received the written feedback Wrong, which was displayed for 500 ms.

Sequences of events in Experiments 4a, 4b, and 5. See the online publication for the color version of the figure. In all cases, participants were instructed to search for two colors: either red and brown or blue and green. In the present example, let us assume that participants were instructed to search for red and brown. As can be seen, the target (highlighted by a broken circle in the rightmost boxes) always had one of the searched-for colors (here, red) and one of the colors that was not pointed out to the participants during the instructions (here, green). Cues could be either top-down matching (i.e., having a task-relevant, instructed color) or top-down nonmatching (i.e., having a task-irrelevant color, which was not pointed out in the instructions). In Experiment 4, the participants’ task was to respond to the orientation of the target cross (i.e., whether it was tilted or straight; in the present case, the target cross is tilted). In Experiment 5, participants were instructed to respond with four designated keys to the orientation of the target color inside the cross (i.e., whether it was presented in the top, bottom, left, or right arm of the cross)
In total, participants worked through 1,280 trials in miniblocks of 80 trials each. The trial type—whether the trial was valid or invalid, and which target and which cue were presented—was randomly chosen on each trial. Before the experiment started, participants received a few trials of practice, and the experiment started when participants reached an accuracy level of 80% (eight out of ten trials correct).
Results
Reaction times
The data from Experiments 4a and 4b were merged. Data of two participants were excluded from the analysis because their error rate exceeded 40%. Notice that the analyses were also performed for each of the experiments separately. Since there were no qualitative differences in the results, we analyzed the results of both experiments together in a mixed-design ANOVA, with the within-subjects variables cue validity (valid vs. invalid), target color n–1 (target in trial n–1 = cue color in trial n vs. target in trial n–1 ≠ cue color in trial n), and cue color relevance (instructed, relevant color cue vs. task-irrelevant color cue). Experiment (4a vs. 4b) was a between-participants variable. RTs were collected from trials on which participants responded correctly. This resulted in the exclusion of 6.0% of the trials in which participants responded incorrectly. Next, RTs that deviated by more than 2.5 SDs from the mean of the corresponding condition were removed (2.6% of trials), and the results were fed into the ANOVA. This led to main effects of cue color relevance, F(1, 41) = 9.40, p < .01, ηp2 = .19, and cue validity, F(1, 41) = 38.18, p < .001, ηp2 = .49. Most importantly, the ANOVA also led to a two-way interaction between cue color relevance and cue validity, F(1, 41) = 25.86, p < .001, ηp2 = .39. Post-hoc t tests showed that overall, only instructed, task-relevant color cues reliably captured attention, 24 ms, t(41) = 7.21, p < .001, whereas noninstructed, task-irrelevant color cues did not, 4 ms, t(41) = 1.60, p = .12. The ANOVA also led to a two-way interaction between target color in trial n–1 and cue validity, F(1, 41) = 9.80, p < .01, ηp2 = .20, indicating that the cueing effect was larger overall when the cue’s color was primed by the target in trial n–1, 19 ms, t(41) = 7.28, p < .001, than when the cue’s color was not primed, 9 ms, t(41) = 3.32, p < .01.
The between-participants variable experiment only interacted with the variable cue color relevance, F(1, 41) = 4.83, p < .05, ηp2 = .11. Other effects were not significant (all nonsignificant Fs < 1.58, all nonsignificant ps > .22). Planned t tests were conducted to assess the cueing effects in each of the conditions separately. When the cue had an instructed feature and was primed by the target in trial n–1, there was a significant cueing effect, 30 ms t(41) = 7.22, p < .001. Likewise, when the cue was not primed but had an instructed feature, there was a reliable cueing effect, 18 ms, t(41) = 4.83, p < .001. We also observed a significant cueing effect when the cue was primed but did not have an instructed target feature, 7 ms, t(41) = 2.29, uncorrected p < .05. There was no cueing effect when the cue did not have an instructed target feature and was not primed, 1 ms, t(41) = 0.29, p = .77. For the results, see Fig. 8.

Results of Experiments 4a and b. In each case, the upper two panels depict the means of the reaction times (RTs), with instructed, task-relevant color cues (on the left) and uninstructed, task-irrelevant color cues (on the right). The filled circles correspond to the data when the cue’s color was primed by the target color in trial n–1, and the nonfilled circles correspond to the data when the cue’s color was not primed. The two panels on the bottom on each side show the error rates in the corresponding conditions. Error bars represent the within-subjects SEs (Cousineau, 2005)
Errors
A corresponding ANOVA with the arcsine square-root-transformed error rates led to a main effect of cue color relevance, F(1, 41) = 8.47, p < .01, ηp2 = .18, and to a main effect of cue validity F(1,41) = 8.10, p < .01, ηp2 = .17. There was also a marginally significant interaction between cue validity, cue color relevance, and experiment, F(1, 41) = 4.09, p = .05, ηp2 = .09. Other effects were not significant (nonsignificant Fs > 3.76, nonsignificant ps > .06).
Planned t tests with each of the experimental conditions revealed no reliable effects (all ts < 1.99, all ps > .053; if anything, the effect closest to significance, with p = .053, was for task-irrelevant feature cues in the unprimed conditions). Since this result indicated that some speed–accuracy trade-off could be involved, we repeated the planned t tests with the inverse efficiency scores (IESs; Townsend & Ashby, 1978). This led to a significant cueing effect when the cue was primed and had an instructed target feature, 36 ms, t(41) = 4.01, p < .001. With task-relevant feature cues that were not primed, the cueing effect was also significant, 24 ms, t(41) = 3.66, p < .01. There were no significant effects, however, with the task-irrelevant feature cues, ts < 1.07, ps > .29.
Discussion
Overall, the result of Experiments 4a and 4b corroborated the results of Experiments 1–3. We found a highly significant validity effect for cues that had features participants were instructed to search for. Overall, cues that had task-irrelevant features did not capture attention. However, with task-irrelevant feature cues, we observed a cueing effect when the corresponding cue color was primed by the preceding target. Yet this effect was only observed without correction for an opposite tendency in error rates and without correction for multiple comparisons, and it was still smaller than the effects we observed with task-relevant feature cues, even when the corresponding task-relevant cue color was not primed [it was smaller by 10 ms; t(41) = 2.00, p = .052].
Interestingly, we also found an effect of intertrial priming from one trial to the next. However, this time, the intertrial priming effect was not restricted to the task-relevant features. That is, the lack of a three-way interaction between cue validity, target color n–1, and cue color relevance indicated that intertrial priming generalized to task-irrelevant target features. This result shows that indeed, under certain conditions, instructions concerning the corresponding feature are not important for the emergence of intertrial priming of capture effects (cf. Kristjánsson, 2006).
What is remarkable is that the contingent-capture effect (the stronger validity effect for cues that had the task-relevant feature than for cues that had the uninstructed, and in this respect task-irrelevant, feature) was found even though the intertrial priming effect emerged for both task-relevant and task-irrelevant features. Granted that priming by task-irrelevant features is possible under appropriate conditions, if priming could explain the full range of attentional capture by top-down matching cues (cf. Theeuwes, 2013), we would have expected the task-irrelevant feature cues to capture attention to the same extent as the task-relevant feature cues. Because this was not the case, Experiment 4 suggests that intertrial priming by uninstructed, task-irrelevant features cannot account for the full range of the contingent-capture effect, even though the same features create a significant cueing effect once they are made relevant by the instructions, and even though relevant and irrelevant features are from the same feature dimension (here, color).
Experiment 5
In Experiment 4, we again obtained a capture effect by instructed, task-relevant features, and features that were irrelevant to the task at hand produced no significant validity effect overall, though these features were also primed and were also associated with the targets. Importantly, this was shown for instructed and for task-irrelevant features that were even defined in the same feature dimension of color. However, in Experiment 4 we found that the intertrial priming effect generalized to the task-irrelevant features. In other words, the intertrial priming effect emerged independently of the instructions concerning the corresponding features. But the target-defining features in Experiment 4 were special in one respect: Both the instructed and task-irrelevant color features carried the same response-relevant target orientation information. That is, when the target was tilted, both the square in the task-relevant color and the square in the task-irrelevant color were tilted. Likewise, when the target stood straight, both colored squares within the stimulus stood straight. It is thus possible that this response relevance or response congruence was important for the priming effects. For example, it could be that response-congruent target information was incorporated into the target-object files, with the consequence that priming generalized to the response-congruent task-irrelevant features (cf. Kristjánsson, 2006; see also Kristjánsson et al., 2008).
To test whether this could have been the case, we conducted Experiment 5 as a control experiment. The difference from Experiment 4 was that a four-choice response was now given based on the relative location of the corresponding color in a straight cross (i.e., whether the instructed color bar of a cross was at the top, at the bottom, or on the left or right within the cross). Again, each target cross was indicated by two colors, only one of which was instructed, but now the task-irrelevant target color marked a cross bar that was incongruent to the response required for the instructed target color. This should undermine priming by the task-irrelevant colors if such priming depends on the integration of response-congruent colors into joint target-object representations.
Method
Participants
Twenty-one participants took part (12 male, nine female; Mage= 22.29 years, SDage = 5.24 years). The participants were again students who participated in return for course credit.
Apparatus, stimuli, and design
These were the same as in Experiment 4, with the exception that now all crosses were straight and the participants’ task was to respond to the location of the square with the searched-for color inside of the target cross. Participants placed their right index finger on the 5 key of the number pad, and responses were given with the 2 key when the color in the target cross was at the bottom, with the 8 key when the color was at the top, with the 4 key when the target was in the left arm of the target, and with the 6 key when the color was in the right arm of the target cross.
Results
Reaction times
Incorrect trials were removed (3.2%), and trials that deviated more than 2.5 SDs from the mean of the corresponding condition were removed (2.2% of the trials). We conducted an ANOVA on the mean RTs in each condition, with the variables cue validity, target color n–1, and cue color relevance. This led to a main effect of cue color relevance, F(1, 20) = 24.39, p < .001, ηp2 = .55, and a main effect of cue validity, F(1, 20) = 65.30, p < .001, ηp2 = .77. In addition, there was an interaction between the variables cue validity and cue color relevance, F(1, 20) = 50.82, p < .001, ηp2 = .72. We found a cueing effect with task-relevant color cues, 41 ms, t(20) = 9.16, p < .001, but not with task-irrelevant color cues, 2 ms, t(20) = 0.52, p = .61. Other effects in the ANOVA were not significant (all nonsignificant Fs < .34, all nonsignificant ps > .56). As regards the effect of intertrial priming, we computed the Bayes factor: The data for the two steps of the variable cue color relevance were averaged, and a Bayesian test was conducted comparing the overall validity effects (invalid minus valid RT) with cues similar to one of the target’s colors in trial n–1 versus cues dissimilar to one of the target’s colors in trial n–1. The BF10 was 0.27, favoring the null hypothesis over the alternative hypothesis. For the results, see Fig. 9.

Results of Experiment 5. The upper two panels depict the means of the reaction times (RTs), with instructed, task-relevant color cues (on the left) and uninstructed, task-irrelevant color cues (on the right). The filled circles correspond to the data when the cue’s color was primed by the target color in trial n–1, and the nonfilled circles correspond to the data when the cue’s color was not primed. The two panels on the bottom show the error rates in the corresponding conditions. Error bars represent the within-subjects SEs (Cousineau, 2005)
Errors
Arcsine-transformed error rates were fed into an ANOVA like the one above. This led to a main effect of cue validity, F(1, 20) = 14.27, p < .01, ηp2 = .42, and to an interaction between the variables cue validity and cue color relevance, F(1, 20) = 20.59, p < .001, ηp2 = .51: There was a cueing effect with task-relevant color cues, t(20) = 4.64, p < .001, but not with task-irrelevant color cues, t(20) = 1.59, p = .13.
Discussion
Experiment 5 again showed a contingent-capture effect. However, this time we found no indication of intertrial priming of attentional capture. Since we only intended to eliminate the priming effect for the uninstructed, task-irrelevant features, it is surprising that intertrial priming ceased to exert an influence not only for the task-irrelevant feature cues, but also for the task-relevant feature cues. Maybe this had to do with the higher importance of the exact relative position of the target-color square within crosses for responses that required more scrutiny than the judgment of the global figure orientation in Experiment 4. This, in turn, could have lowered the relative strength of the contribution of color to priming effects per se. What exactly the reason was for the absence of the intertrial effect, however, is not so important in the present context. What is important is that the capture effect again clearly emerged only with the cues carrying a relevant, instructed feature, but not with the cues carrying an uninstructed, and in this respect irrelevant, feature, as well as that we found not the slightest indication of intertrial priming of these irrelevant features. This is further evidence for the independence of the contingent-capture effect of the relevant cues from priming and shows that feature repetition of task-irrelevant features from one target to the next cue is not sufficient to mimic validity effects by truly target-relevant cues.
General discussion
In the present study, we investigated to what extent target features that are task-irrelevant (i.e., that participants are not instructed to search for) would foster the emergence of attentional capture mimicking contingent-capture effects by cues that had searched-for features. Answering this question is important, because it has been suggested that the contingent-capture effect might emerge as a consequence of priming resulting from feature repetitions in selection history (e.g., Belopolsky et al., 2010; Theeuwes, 2013). In light of theories that postulate that the processing of an object leads to cortical enhancement of all of its object features (e.g., Duncan, 1998), the mere processing of the target could be sufficient for the enhancement and, hence, also the priming of all its features, regardless of their task relevance. Indeed, prior studies have sometimes reported (Kristjánsson, 2006) priming by completely task-irrelevant features. According to this view, stronger validity effects of target-similar than of dissimilar cues (i.e., “contingent-capture effects”) could emerge as a result of priming based on the mere processing of the object comprising the target, without any need for the instruction-based task relevance of the corresponding feature value.
The present results, however, are at odds with this idea: We found little evidence that seeing task-irrelevant features in the target alone gave rise to attentional capture effects by, thus, primed target-similar cues. In Experiments 1 and 2, in which participants searched for color and the target had a specific disk composite, we found no attentional capture of cues that had the target disk composite, over and above the attentional capture that was explained by the color cues alone. This was the case even though the specific disk composite was realized in the target in every trial, so that even cumulative priming would have been possible (cf. Kruijne et al., 2015), to some extent. In contrast, in Experiment 3, in which participants searched for the disk composite, cues that had the corresponding disk composite captured attention. It is true that in Experiment 3, task-irrelevant color cues that participants were not instructed to search for also captured attention. However, we found no indication of intertrial priming by the corresponding color from one trial to the next. This suggests that the moderate cueing effect by color in Experiment 3 was due to some other attentional guidance principle. It is possible that participants incorporated the feature color into their search setting at least in some trials, but that this experiment was then one of several instances in which priming of capture failed entirely, even with regard to relevant features (see Ansorge & Horstmann, 2007; Irons et al., 2012). It is also possible that a form of statistical learning gave rise to the attentional-capture effects by color in Experiment 3 (cf. Lanthier et al., 2015). In any case, the magnitude of the validity effect of the task-irrelevant target colors was clearly smaller than the effects with cues of a task-relevant color in Experiments 1 and 2.
One could object that the task-relevant and task-irrelevant features were defined in different dimensions in Experiments 1–3. It is, therefore, possible that participants down-weighted the task-irrelevant feature dimension (Found & Müller, 1996; Müller et al., 1995), and that the features thus could not give rise to attentional capture through priming. To find out whether this could have been the case, we conducted Experiments 4 and 5, in which the instructed, task-relevant feature of the target was a color and the uninstructed, task-irrelevant feature was a color, too. Dimensional down-weighting of the uninstructed, task-irrelevant target feature was therefore not possible. The results of these experiments further corroborated our conclusions from Experiments 1–3. In Experiment 4, we did find evidence for intertrial priming by the task-relevant features from one trial to the next and, a bit surprisingly, by the task-irrelevant feature too. However, there were also clear indications that contingent-capture effects were independent of priming, reflected in significant and much higher validity effects of the task-relevant feature cues only, even when these were not primed, meaning that overall, priming by task-irrelevant features cannot account for contingent-capture effects. Task-irrelevant color cues did not even lead to a reliable validity effect overall (i.e., they were not significant when RT differences were weighted by opposite error tendencies). Given this apparent dissociation between the validity effects of relevant cues, on the one hand, and the intertrial priming effects, on the other, it is difficult to maintain the position that the contingent-capture effects could be (exclusively) based on priming. If the contingent-capture effects were explained by priming alone, cueing effects should have emerged to approximately the same degree for primed task-relevant and task-irrelevant target features.
The pattern of results that emerged in Experiment 5 was slightly different from that of Experiment 4, but it was still in agreement with the conclusion that priming by uninstructed, task-irrelevant features cannot lead to substantial attentional-capture effects. In Experiment 5, we found no indication of intertrial priming from one trial to the next. It is difficult to pin down the exact reason for this lack of an intertrial priming effect even with task-relevant features. We speculate that the task may have been a critical factor. In Experiment 5, we used a four-alternative forced choice (4AFC) task. Since the congruency of the response from one trial to the next is sometimes considered to be important for the emergence of intertrial priming effects (e.g., Lamy, Yashar, & Ruderman, 2010), the use of a 4AFC task could have undermined intertrial priming, because it increased the ratio of trial-by-trial response-incongruent relative to trial-by-trial response-congruent trials. Whatever was the exact reason for the absence of an intertrial priming effect, we still found evidence in line with a contingent-capture effect, in which attentional capture was restricted to the cues with an instructed, task-relevant feature. Cues with a task-irrelevant feature again had no validity effect.
Limitations of the present study and future directions
One problematic aspect of the present study is that we defined the task relevance of the target features only by instruction. That is, participants were informed about the task-relevant features, and the task-irrelevant features were not pointed out to them in the instructions. But, because the task-irrelevant features in principle could also be used to find the target, we cannot know for sure whether participants incorporated the task-irrelevant features in their search settings. As regards Experiments 1, 2, and 5, this is a minor problem, because validity effects for cues that had a task-irrelevant target feature were entirely absent. However, the possibility that participants incorporated task-irrelevant features into their search settings complicates the interpretation at least of Experiments 3 and 4, in which validity effects (Exp. 3) and intertrial priming effects (Exp. 4) were found with task-irrelevant features. As regards Experiment 3, it remains unclear whether the moderate validity effect by color that we observed despite color being task-irrelevant was due to the fact that participants searched actively for the corresponding feature, at least in some trials. Likewise, it is unclear whether the intertrial priming by task-irrelevant colors that we observed in Experiment 4 was at least partly driven by an explicit strategy to look for the corresponding color.
A second issue that complicates the interpretation of the present study is the possibility that task-irrelevant features were inhibited and, therefore, failed to capture attention. In fact, there are some indications in the data that inhibition could have played a role, at least in Experiments 4 and 5: We found faster responses with task-relevant than with task-irrelevant feature cues. It is conceivable that this slowing reflected inhibition of task-irrelevant features. We think, however, that this alternative explanation for finding no validity effect by task-irrelevant feature cues is not very likely. First of all, it is not clear why participants would have adopted the strategy to actively inhibit task-irrelevant colors. Inhibition would be a detrimental strategy, because the corresponding colors were realized in the target. Second, there are alternative explanations for the relative slowing of the response times with task-irrelevant cues: (1) Responses with task-relevant colors could simply have been faster because, in a subset of trials, valid cues speeded responding; (2) in the case of a task-relevant feature cue, the cue could have primed the corresponding search setting (e.g., Büsel, Pomper, & Ansorge, 2018; Irons et al., 2012; see also Moore & Weissman, 2010), so that responding to the target was faster when the target was similar to a previously presented cue.
A third issue is that skeptics might argue that priming by task-irrelevant features just takes longer to evolve than does priming by task-relevant features. This, for example, could explain the fact that we found a smaller validity effect for task-irrelevant color cues in Experiment 3 than in Experiments 1 and 2, in which color was relevant. Namely, the targets were presented in two different colors, and color, therefore, did not remain constant from one trial to the next. What might happen if the color remained constant over a whole block of trials? Clearly, this is an empirical question and could be addressed by future studies. We think, however, that the evidence against a strong version of the priming account is still compelling. Notice that the composite feature that was task-irrelevant in Experiments 1 and 2 did remain the same in the target in each trial, and still no capture effects were found.
Finally, we would point out that in the present study we tested a very strong version of a priming account of the contingent-capture effect: We mainly considered the possibility that contingent-capture effects emerge based on mere feature repetition of attended targets, without task relevance of the corresponding features. Clearly, our study is almost silent with respect to the possibility that priming by task-relevant features plays an important role in contingent-capture effects. There was at least a small hint in our data suggesting that even priming by task-relevant features cannot explain the contingent-capture effect: In Experiment 4, we found statistically indistinguishable intertrial priming effects by task-irrelevant and task-relevant target colors. At the same time, the attentional validity effect was stronger, and actually was exclusively present, for task-relevant as compared to task-irrelevant features cues. The lack of a three-way interaction in this experiment points toward a dissociation between the contingent-capture effect and priming. However, this is a null finding, and the intertrial priming effect by task-irrelevant features in Experiment 4 was not expected. Therefore, there is clearly a need for more research, both theoretical and empirical, to address the question of the extents to which the contingent-capture effect could reflect voluntary attention and to which automatic processes such as priming by task-relevant features could be involved. In two (Exps. 1 and 4) of the three experiments in which we assessed the intertrial priming of task-relevant features, reliable priming effects from one trial to the next were found. Future studies, therefore, will have to further address the question of the extent to which the contingent-capture effects depends on flexible search settings and the extent to which automatic processes, such as priming by task-relevant features, are involved.
Conclusions
Together, the present results suggest that a strong version of the priming account (priming by mere target feature repetition, without task relevance of the corresponding features) is not supported. In none of the five experiments we conducted did we find substantial evidence of validity effects (or of enhancement of validity effects) when the cue had one of the target’s task-irrelevant but primed features; only relevant cues carrying an instructed, and thus searched-for, feature captured attention. It remains unclear, however, to what extent priming by task-relevant features is responsible for contingent capture. This question will need to be addressed by future studies.
Notes
It is also possible that in nonmatching cueing displays, participants’ attention is equally likely to be captured by each stimulus with equal probabilities (e.g., by chance), or that participants’ attention is first captured by the nonmatching singleton and then deallocated back to screen center (cf. Theeuwes, Atchley, & Kramer, 2000). Although the latter option is unlikely (cf. Ansorge, Kiss, Worschech, & Eimer, 2011; Schoeberl, Goller, & Ansorge, 2018), the former remains a possibility. If participants attended to one of the stimuli in the nonmatching cueing displays, cumulative priming by successive targets of the same color would be undermined by intermediate attention to a differently colored stimulus.
It is unlikely that the priming from cue to target within a given trial directly accounts for a spatial validity effect, because this form of priming is likely not location-specific (cf. Büsel, Pomper, & Ansorge, 2018; Irons et al., 2012); priming of the target features by a matching cue might, however, contribute to a cumulative effect of priming, especially over several immediately succeeding valid trials.
The task-irrelevant target-discriminating feature of Experiments 1 and 2 was labeled a composite, although it could also be that another feature that was correlated with the disk–ring composite, such as the size of the circumference of the ring or the thickness of the ring discriminated target and distractors. Thus, we use the term composite as one of several alternative labels that we could have used, without assuming that searching for a particular disk–ring composite would have been the only way that participants could have searched for targets by their task-irrelevant features. This is important to note, because the target’s task-irrelevant discriminating feature was, thus, not necessarily more difficult to process than the relevant feature;,it was not the case that only the task-irrelevant feature required conjunction search, whereas the relevant feature did not. In fact, in Experiment 3 we showed that participants could efficiently search for targets by the “composite feature” if they were instructed to do so.
References
Anderson, B. (2016). The attention habit: How reward learning shapes attentional selection. Annals of the New York Academy of Sciences, 1369, 24–39. https://doi.org/10.1111/nyas.12957
Ansorge, U., & Becker, S. (2012). Automatic priming of attentional control by relevant colors. Attention, Perception, & Psychophysics, 74, 83–104. https://doi.org/10.3758/s13414-011-0231-6
Ansorge, U., & Horstmann, G. (2007). Preemptive control of attentional capture by color: Evidence from trial-by-trial analysis and ordering of onsets of capture effects in RT distributions. Quarterly Journal of Experimental Psychology, 60, 952–975. https://doi.org/10.1080/17470210600822795
Ansorge, U., Kiss, M., Worschech, F., & Eimer, M. (2011). The initial stage of visual selection is controlled by top-down task set: New ERP evidence. Attention, Perception, & Psychophysics, 73, 113–122. https://doi.org/10.3758/s13414-010-0008-3
Awh, E., Belopolsky, A., & Theeuwes, J. (2012). Top-down versus bottom-up attentional control: A failed theoretical dichotomy. Trends in Cognitive Science, 16, 437–443. https://doi.org/10.1016/j.tics.2012.06.010
Bacon, W. F., & Egeth, H. E. (1994). Overriding stimulus-driven attentional capture. Perception & Psychophysics, 55, 485–496. https://doi.org/10.3758/BF03205306
Becker, S. I. (2018). Reply to Theeuwes: Fast feature-based top-down effects, but saliency may be slow. Journal of Cognition, 1, 28. https://doi.org/10.5334/joc.23
Belopolsky, A., Schreij, D., & Theeuwes, J. (2010). What is top-down about contingent capture? Attention, Perception, & Psychophysics, 72, 326–341. https://doi.org/10.3758/APP.72.2.326
Büsel, C., Pomper, U., & Ansorge, U. (2018). Capture of attention by target-similar cues during dual-color search reflects reactive control among top-down selected attentional control settings. Psychonomic Bulletin & Review. Advance online publication. https://doi.org/10.3758/s13423-018-1543-5
Büsel, C., Voracek, M., & Ansorge, U. (2018). A meta-analysis of contingent-capture effects. Psychological Research. Advance online publication. https://doi.org/10.1007/s00426-018-1087-3
Campana, G., & Casco, C. (2009). Repetition effects of features and spatial position: Evidence for dissociable mechanisms. Spatial Vision, 22, 325–338. https://doi.org/10.1163/156856809788746318
Carmel, T., & Lamy, D. (2014). The same-location cost is unrelated to attentional settings: An object-updating account. Journal of Experimental Psychology: Human Perception and Performance, 40, 1465–1478. https://doi.org/10.1037/a0036383
Cousineau, D. (2005). Confidence intervals in within-subject designs: A simpler solution to Loftus and Masson’s method. Tutorials in Quantitative Methods for Psychology, 1, 42–45. https://doi.org/10.20982/tqmp.01.1.p042
Duncan, J. (1998). Converging levels of analysis in the cognitive neuroscience of visual attention. Philosophical Transactions of the Royal Society B, 353, 1307–1317. https://doi.org/10.1098/rstb.1998.0285
Eimer, M., Kiss, M., Press, C., & Sauter, D. (2009). The roles of feature-specific task set and bottom-up salience in attentional capture: An ERP study. Journal of Experimental Psychology: Human Perception and Performance, 35, 1316–1328. https://doi.org/10.1037/a0015872
Fecteau, J. H. (2007). Priming of pop-out depends upon the current goals of observers. Journal of Vision, 7(6), 1:1–11. https://doi.org/10.1167/7.6.1
Folk, C. L., & Remington, R. W. (2008). Bottom-up priming of top-down attentional control settings. Visual Cognition, 16, 215–231. https://doi.org/10.1080/13506280701458804
Folk, C. L., Remington, R. W., & Johnston, J. C. (1992). Involuntary covert orienting is contingent on attentional control settings. Journal of Experimental Psychology: Human Perception and Performance, 18, 1030–1044. https://doi.org/10.1037/0096-1523.18.4.1030
Found, A., & Müller, H. J. (1996). Searching for unknown feature targets on more than one dimension: Investigating a “dimension-weighting” account. Perception & Psychophysics, 58, 88–101. https://doi.org/10.3758/BF03205479
Goller, F., & Ansorge, U. (2015). There is more to trial history than priming of attentional capture. Attention, Perception, & Psychophysics, 77, 1574–1584. https://doi.org/10.3758/s13414-015-0896-3
Grubert, A., & Eimer, M. (2013). Qualitative differences in the guidance of attention during single-color and multiple-color visual search: Behavioral and electrophysiological evidence. Journal of Experimental Psychology: Human Perception and Performance, 39, 1433–1442. https://doi.org/10.1037/a0031046
Hamann, S. B., & Squire, L. R. (1997). Intact perceptual memory in the absence of conscious memory. Behavioral Neuroscience, 111, 850–854. https://doi.org/10.1037/0735-7044.111.4.850
Hickey, C., Di Lollo, V., & McDonald, J. J. (2009). Electrophysiological indices of target and distractor processing in visual search. Journal of Cognitive Neuroscience, 21, 760–775. https://doi.org/10.1162/jocn.2009.21039.
Irons, J. L., Folk, C. L., & Remington, R. W. (2012). All set! Evidence of simultaneous attentional control settings for multiple target colors. Journal of Experimental Psychology: Human Perception and Performance, 38, 758–775. https://doi.org/10.1037/a0026578
Jiang, Y. V. (2018). Habitual versus goal-driven attention. Cortex, 102, 107–120. https://doi.org/10.1016/j.cortex.2017.06.018
Kappenman, E. S., & Luck, S. J. (2010). The effects of electrode impedance on data quality and statistical significance in ERP recordings. Psychophysiology, 47, 888–904. https://doi.org/10.1111/j.1469-8986.2010.01009.x
Kristjánsson, Á. (2006). Simultaneous priming along multiple feature dimensions in a visual search task. Vision Research, 46, 2554–2570. https://doi.org/10.1016/j.visres.2006.01.015
Kristjánsson, Á, & Campana, G. (2010). Where perception meets memory: A review of repetition priming in visual search tasks. Attention, Perception, & Psychophysics, 72, 5–18. https://doi.org/10.3758/APP.72.1.5
Kristjánsson, Á., Ingvarsdóttir, Á., & Teitsdóttir, U.D. (2008). Object- and feature-based priming in visual search, Psychonomic Bulletin & Review, 15, 378–384. https://doi.org/10.1037/a0026578
Kruijne, W., Brascamp, J. W., Kristjánsson, Á., & Meeter, M. (2015). Can a single short-term mechanism account for priming of pop-out? Vision Research, 115, 17–22. https://doi.org/10.1016/j.visres.2015.03.011
Lamy, D., Yashar, A., & Ruderman, L. (2010). A dual-stage account of inter-trial priming effects. Vision Research, 48, 1274–1279. https://doi.org/10.1016/j.visres.2010.01.008
Lanthier, S. N., Wu, D., Chapman, C. S., & Kingstone, A. (2015). Resolving the controversy of the proportion validity effect: Volitional attention is not required, but may have an effect. Attention, Perception, & Psychophysics, 77, 2611–2621. https://doi.org/10.3758/s13414-015-0956-8
Maljkovic, V., & Nakayama, K. (1994). Priming of pop-out: I. Role of features. Memory & Cognition, 22, 657–672. https://doi.org/10.3758/BF03209251
Marsman, M., & Wagenmakers, E.-J. (2016). Bayesian benefits with JASP. European Journal of Developmental Psychology, 14, 545–555. https://doi.org/10.1080/17405629.2016.1259614
Moore, K. S., & Weissman, D. H. (2010). Involuntary transfer of a top-down attentional set into the focus of attention: Evidence from a contingent attentional capture paradigm. Attention, Perception, & Psychophysics, 72, 1495–1509. https://doi.org/10.3758/APP.72.6.1495
Müller H. J., Heller D., & Ziegler J. (1995). Visual search for singleton feature targets within and across feature dimensions. Perception & Psychophysics, 57, 1–17. https://doi.org/10.3758/BF03211845
Remington, R., & Folk, C. (2001). A dissociation between attention and selection. Psychological Science, 12, 511–515. https://doi.org/10.1111/1467-9280.00394
Schoeberl, T., Ditye, T., & Ansorge, U. (2018). Same-location costs in peripheral cueing: The role of cue awareness and feature changes. Journal of Experimental Psychology: Human Perception and Performance, 44, 433–451. https://doi.org/10.1037/xhp0000470.
Schoeberl, T., Goller, F., & Ansorge, U. (2018). Top-down matching singleton cues have no edge over top-down matching nonsingletons in spatial cueing. Psychonomic Bulletin & Review. Advance online publication. https://doi.org/10.3758/s13423-018-1499-5
Theeuwes, J. (1992). Perceptual selectivity for color and form. Perception & Psychophysics, 51, 599–606. https://doi.org/10.3758/BF03211656
Theeuwes, J. (2010). Top-down and stimulus-driven control of visual selection. Acta Psychologica, 135, 77–99. https://doi.org/10.1016/j.actpsy.2010.02.006
Theeuwes, J. (2013). Feature-based attention: It is all bottom-up priming. Philosophical Transactions of the Royal Society B, 368, 20130055. https://doi.org/10.1098/rstb.2013.0055
Theeuwes, J. (2018). Visual selection: Usually fast and automatic; seldom slow and volitional. Journal of Cognition, 1, 29. https://doi.org/10.5334/joc.13
Theeuwes, J., Atchley, P., & Kramer, A. F. (2000). On the time course of top-down and bottom-up control of visual attention. In S. Monsell & J. Driver (Eds.), Control of cognitive processes: Attention and performance XVIII (pp. 105–125). Cambridge, MA: MIT Press.
Townsend, J. T., & Ashby, F. G. (1978). Methods of modeling capacity in simple processing systems. In J. Castellan & F. Restle (Eds.), Cognitive theory (Vol. 3, pp. 200–239). Hillsdale, NJ: Erlbaum.
Umemoto, A., Scolari, M., Vogel, E. K., & Awh, E. (2010). Statistical learning induces discrete shifts in the allocation of working memory resources. Journal of Experimental Psychology: Human Perception and Performance, 36, 1419–1429. https://doi.org/10.1037/a0019324
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Schoeberl, T., Goller, F. & Ansorge, U. Testing a priming account of the contingent-capture effect. Atten Percept Psychophys 81, 1262–1282 (2019). https://doi.org/10.3758/s13414-019-01672-0
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-019-01672-0