
Abstract
Our spatial perception is not always veridical. Indeed, systematic distortions in localization have been found to result from orienting of attention. Distorted localization is inferred from tasks wherein the subject reports the location of centrally presented parallel (vernier) line stimuli. Particularly, prior to the presentation of the lines, a shift of attention toward peripheral cues produces a mislocalization of the line stimuli away from the cues (termed the attentional repulsion effect [ARE]). However, if the shift of attention is induced after target presentation, by reversing the order of stimulus presentation, a substantial mislocalization toward the cues (attentional attraction effect [AAE]) is found. The purpose of this study was to identify whether the AAE arises from the modulation in the same processes as the ARE. While an interocular presentation of cues to one eye and vernier lines to the other was previously shown to eliminate the ARE, the AAE persists across both the interocular and monocular conditions (both the cues and vernier lines are presented to the same eye). Considering Ono and Watanabe’s (2011) suggestion that memory traces may be involved in generating the AAE, this prospect was examined by having participants delay their response for a short (100 ms) or long (1,000 ms) period of time. The magnitude of AAE was larger with a longer delay, consistent with the involvement of visual memory. Next, to directly examine the role of spatial working memory, the attentional attraction task was embedded within either a spatial memory task (remembering the locations of one or three squares) or a color memory task (remembering the color of one or three squares). Only high spatial memory load reduced the magnitude of AAE. Our results suggest the AAE relies on changes to different visual processes than does the ARE and involves spatial working memory.
Similar content being viewed by others
Orienting attention to specific locations in the visual field confers many advantages, such as faster and more accurate processing of the attended stimuli, as compared with stimuli at unattended locations (1980; Henderson, 1991; Posner, Snyder, & Davidson, 1980; Yantis & Hillstrom, 1994; for recent reviews, see Carrasco, 2011; Theeuwes, 2010). Stimuli at attended locations enjoy priority processing and are often perceived as occurring earlier than other, simultaneously presented stimuli at unattended locations (Stelmach & Herdman, 1991). Furthermore, stimuli at attended locations enjoy enhanced spatial resolution (Tsal & Shalev, 1996; Yeshurun & Carrasco, 1999; although see Schneider & Komlos, 2008) and are localized more accurately (Müller & Rabbit, 1989; Tsal & Bareket, 1999). Concurrent with these advantages, shifts of attention also cause spatial distortions that briefly impair localization at unattended locations (Ono & Watanabe, 2011; Suzuki & Cavanagh, 1997). The purpose of the present study is to provide further understanding of the processes underlying attention-driven spatial distortions.
Suzuki and Cavanagh (1997) were the first to find that shifts of attention to the periphery caused vernier lines presented at fixation to be displaced away from the peripheral cues. They termed this phenomenon the attentional repulsion effect (ARE). Their standard ARE paradigm consisted of the brief presentation of two peripheral cues positioned along a diagonal (top-right and bottom-left vs. Top left and bottom right), followed 100–200 ms later by two vertically aligned vernier lines presented at fixation. Observers would misperceive the top vernier line to be displaced to the left of the bottom line following cues in the top-right and bottom-left positions and to the right of the bottom line for cues in the top-left and bottom-right positions. The ARE appears to be a robust phenomenon that has consequences for subsequent stages of visual processing, since presenting cues near object contours distorts the shape of objects (Fortenbaugh, Prinzmetal, & Robertson, 2011; Kosovicheva, Fortenbaugh, & Robertson, 2010) and similarly produces a bias in the bisection of lines (Toba, Cavanagh, & Bartolomeo, 2011). Taken together, these studies provide strong evidence of a systematic distortion of objects and locations, congruent with repulsion away from the attended location.
As is implied by the name, there is good evidence to suggest that the ARE is indeed attentional in nature. Suzuki and Cavanagh (1997) found that the ARE was caused by both reflexive and voluntary shifts of attention and could not be attributed to figural aftereffects or apparent motion. Converging evidence for the role of attention comes from Pratt and Arnott (2008), who found that manipulations capable of modulating reaction time (RT) based effects of attention also modulated the ARE. For instance, onset, offset, and onset–offset cues, which produce comparable RT effects when presented in isolation (Pratt & McAuliffe, 2001), produced comparable levels of ARE. The magnitude of ARE could vary with the amount of attentional capture, with single onset–offset cues producing greater repulsion effects than a color singleton. Furthermore, the magnitude of ARE is also sensitive to the feature similarity between the cue and the target (Folk, Remington & Johnston, 1992), with target-similar cues causing greater ARE (Gozli & Pratt, 2012). Thus, attentional effects predicted from previous RT experiments were replicated with the ARE, consistent with the key role of attention in the misperception of space.
Interestingly, by simply switching the order of presentation of the stimuli, such that the vernier lines are presented before the peripheral cues, a substantial attentional attraction effect (AAE) is observed (Ono & Watanabe, 2011). That is, observers perceived the top vernier line to be closer to the cues, as opposed to farther away from the cues in the case of ARE, with maximal AAEs found with target–cue stimulus onset asynchronies (SOAs) of 100–200 ms. Similar to the ARE, the AAE also appears to depend on attentional resources. Using an attentional control set to elicit voluntary shifts of attention, Ono and Watanabe verified the existence of an AAE, albeit with a reduced magnitude, as compared with those generated with reflexive shifts of attention.
While it seems that ARE and AAE are both generated through shifts of attention, it is unknown whether or not both effects rely on attention affecting the same underlying mechanism. To say both effects are attentional in nature is unspecific, since attentional biases occur at multiple stages of information processing (Desimone & Duncan, 1995). There is, indeed, some preliminary indication that the two effects are driven through different mechanisms. Suzuki and Cavanagh (1997) reasoned that the ARE might arise from changes in the receptive fields (RFs) of position-encoding cells in early visual regions (see also Tsal & Shalev, 1996). Testing the idea of an early locus for the ARE, DiGiacomo and Pratt (2012) examined the magnitude of the effect across monocular, binocular, and interocular (cues presented to one eye and vernier targets to the other) conditions. The assumption was that if the ARE does not persist in the interocular condition, the interaction between cue and target representations at early stages of visual processing is necessary in causing the ARE. The authors found that the interocular presentation eliminated the effect, suggesting that the ARE relies on cue–target interaction at early stages of visual processing. The early locus of the ARE is also consistent with the findings of Pratt and Turk-Browne (2003), who reported ARE in both perceptual judgment and manual actions, suggesting that the effect exists early in processing before perceptual and actions streams are separated.
In the present study, we are concerned with examining whether the AAE, unlike the ARE, is driven by alterations in visual working memory representation. An indication that the AAE and the ARE have distinct loci comes from the study by Chein, Ono, and Watanabe (2011), who placed cues both before and after the target vernier lines and found that the two effects produce summative distortions of visual space, with the AAE being larger than the ARE. In addition, by presenting the cues twice (once simultaneous with the target and once before or after the targets, depending on ARE/AAE paradigm), they found that reducing attentional shifts eliminated the AAE, whereas the ARE still persisted. Although changes in the early visual RFs may still play a partial role in the generation of the AAE, similar to the ARE (DiGiacomo & Pratt, 2012; Suzuki & Cavanagh, 1997), they may not be the only driving force behind the AAE. Specifically, the AAE may, at least partially, rely on changes to working memory traces (as suggested by Ono & Watanabe, 2011). In other words, the shift of attention after the presentation of the targets may distort the memory representation of the target location.
Given the potential role of working memory in the AAE, the present study proceeds to test this role in two steps. The first step attempts to rule out the possibility that the AAE, like the ARE, is driven by modulation of RFs of position-encoding cells in early visual regions. To do so, in Experiment 1, we subject the AAE to the interocular manipulation used by DiGiacomo and Pratt (2012) that successfully eliminated the ARE. By confirming that the AAE can persist with interocular presentation of targets and cues, we open up the possibility of attributing this effect to higher-level processes, including working memory representation. Subsequently, the second step is to directly evaluate the hypothesis proposed by Ono and Watanabe (2011) that a working memory trace may play a role in the AAE, by incorporating a delay between the offset of the stimulus and when participants are allowed to respond. A longer delay will consolidate the effect of cues on the representation of target location in working memory (e.g., Sheth & Shimojo, 2001). Using a similar logic, Sheth and Shimojo found that the tendency to mislocalize items closer to the fovea and other salient items in the display increased with a longer delay duration. Should the AAE involve the distortion of memory traces, we would expect to find increased levels of spatial distortion with a longer delay. Following Experiment 1, we test the role of working memory in the AAE by having participants perform the task under low/high spatial (Experiment 2) or nonspatial (Experiment 3) memory load conditions. Finding an attenuation of the magnitude of the AAE under spatial memory load would provide a more direct support for the role of this type of memory in the generation of the AAE.
Experiment 1
The key finding of DiGiacomo and Pratt (2012) was that the ARE existed with monocular viewing (cues and targets presented to a single eye) but was completely eliminated with interocular viewing (cues and targets presented to different eyes). If the AAE is also exclusively driven by early visual RFs alteration, as seems to be the case in the ARE, the attraction effect should disappear with interocular viewing. If, however, an AAE is found under both monocular and interocular viewing, the underlying mechanism must be different from the ARE. In addition, we investigate the role of visual working memory in AAE by adding a short (100 ms) or long (1,000 ms) delay before participants respond. If the AAE is due to attentionally mediated changes in RFs, the magnitude of the attraction effect should remain unchanged over time. On the other hand, if the AAE does rely on memory traces, the attraction effect should become more pronounced over time, since the cues have more time to exert a bias on the memory trace of the vernier locations.
Method
Participants
Ten undergraduate students (4 male) participated in the experiment for additional course credit. Each participant had normal or corrected-to-normal vision and provided written consent prior to undergoing the experiment. Participants were all unaware of the purpose of the experiment. All experimental protocols were approved by the Research Ethics Board at University of Toronto.
Apparatus
Stimuli were presented on a 19-in. ViewSonic Graphics Series G90fB monitor at a resolution of 1,024 × 768 that operated at a refresh rate of 85 Hz. Participants sat in a dimly lit room and viewed the monitor at a distance of 48 cm fixed with the assistance of a chinrest. Portable Liquid-Crystal Apparatus for Tachistoscopic Occlusion (PLATO) goggles were used to facilitate precise monocular and binocular presentations of stimuli, turning opaque to occlude vision in either eye.
Procedure
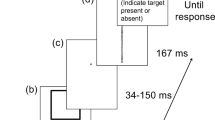
The basic attentional attraction paradigm consists of the presentation of two vernier lines before two peripheral circular cues (Fig. 1). Each trial began with the presentation of a white central fixation cross, subtending 0.32o × 0.32o, for 1,000 ms on a black background. Next, two white vernier lines, each 1.44o long and 0.1o wide (distance from horizontal midline = 2°), were presented for 100 ms. The top vernier line could occupy three positions: either directly above the bottom vernier line or horizontally displaced 0.36o to the left or right. After an SOA of 200 ms, two circular cues were presented for 50 ms in diagonal pairs, either top-left with bottom-right or top-right and bottom-left. The cues were 1.44o in diameter, and their centers deviated 6.32o from the display center.

Basic trial sequence of the attentional attraction paradigm. In this paradigm, cue presentation is preceded by target presentation (opposite to attentional repulsion). The target (i.e., top vernier line) could appear to the left, right, or directly above the bottom vernier line
Participants viewed the stimuli in both the monocular and interocular conditions in a counterbalanced and blocked order. In the monocular condition, the participant viewed both the vernier lines and the circular cues with the same eye, with either the right or left lens of the goggles occluded (at an equal likelihood) for the duration of the trial. In the interocular condition, participants would view the vernier lines with one eye and the circular cues with the other, as the LCD goggles would alternate the occlusion between the presentation of the lines and cues. Each eye was equally likely to see either the vernier lines or the circular cues. In both conditions, both lenses were opened at the end of the trial for 1,000 ms before the next trial commenced.
Participants were asked to indicate whether the top vernier line was displaced to the left or right (a forced choice) with the z and / keys, respectively and were prompted to make a response upon hearing a beep 100 or 1,000 ms after the cues disappeared. Participants performed 20 practice trials prior to completing 360 test trials, with an opportunity to take a break every 60 trials.
Results and discussion
Trials on which participants pressed any key other than “left” or “right” or had RTs that exceeded 2.5 standard deviations from the mean were excluded from analysis. In addition, 2 participants were replaced for performing with less than 75% accuracy (this accuracy measure was calculated after excluding trials on which the top vernier line was vertically aligned with the bottom vernier line).
The percentages of left responses for the left, central, and right vernier lines were computed, and the data are displayed in Fig. 2. We confined the analysis to trials with the central vernier line. Percentage of left responses was analyzed by a 2 (view: monocular or interocular) × 2 (cue: top-left or top-right) × 2 (delay: 100 or 1,000 ms) ANOVA. A main effect of cue, F(1, 9) = 22.89, p < .01, ηp 2 = .766, p rep Footnote 1 = .98] revealed a robust AAE; participants were more likely to indicate that the top vernier line was displaced to the left when a top-left cue was present and displaced to the right in the case of a top-right cue. There were no main effects of view, F(1, 9) = 1.02, p = .35, ηp 2 = .102, or delay, F(1, 9) < 1. Importantly, there was no view × cue interaction, F(1, 9) = 1.25, p = .30, ηp 2 = .152, JZS Bayes FactorFootnote 2 = 2.46, since AAEs were found in both viewing conditions. Nevertheless, it could be argued that the absence of a view × cue interaction may be due to low statistical power, since the AAE seems larger in the monocular condition (M ± SE = .23 ± .04, Cohen's d = 1.73), as compared with the interocular condition (M ± SE = .17 ± .05, Cohen's d = 1.17). The important point, however, is that the AAE in the interocular viewing condition is significantly greater than zero, t(9) = 3.48, p < .01. The interaction between delay and cue was significant, F(1, 9) = 15.63, p < .01, ηp 2 = .691, p rep = .92, with greater AAEs found with the longer delays (M ± SE = 0.26 ± 0.04), as compared with shorter delays (M ± SE = 0.13 ± 0.04). Finally, the two-way interaction between viewing condition and delay, F(1, 9) = 2.64, p = .148, ηp 2 = .264, and the three-way interaction were both not reliable, F(1, 9) < 1, JZS Bayes factor = 4.30.

Percentage of left responses on the attentional attraction task in Experiment 1 across the monocular and interocular conditions with delayed responses of 100 ms (left panel) and 1,000 ms (right panel). Error bars denote 95% confidence intervals
The main finding from the first experiment is that AAE was found in both monocular and interocular conditions, unlike the absence of the ARE under interocular viewing as reported by DiGiacomo and Pratt (2012). Although we cannot rule out the role of early RFs in the AAE, our findings indicate that the AAE is not driven solely by changes to the RFs of these early position-encoding cells (unlike the ARE). Moreover, AAEs were found to be more pronounced with the longer delay, consistent with the findings of Yamada, Miura, and Kawabe (2011) that AAE begins to emerge with target–probe SOAs of longer than 600 ms. Hence, it is possible that visual working memory is involved in the AAE. The following two experiments were constructed to directly evaluate this hypothesis.
Experiment 2
The larger AAEs found with longer delays in the previous experiment are consistent with the notion that distortion of memory traces may be the source of this effect. To explicitly test this memory hypothesis, a spatial memory task was added to the AAE paradigm in the present experiment. Should the AAE require spatial working memory resources, the AAE should be attenuated under the circumstance that these resources are partially occupied.
Method
Participants
Sixteen different undergraduate students (9 male) participated in this experiment to receive additional course credit. Each participant had normal or corrected-to-normal vision and provided written consent prior to undergoing the experiment.
Apparatus and procedure
The same monitor and chinrest setup was used as in Experiment 1. To test whether spatial working memory was involved, a spatial load task was added to the AAE paradigm (Fig. 3). A white fixation cross spanning 0.5o × 0.5o was presented for 1,000 ms on a black background prior to the spatial load presentation. Participants were asked to remember the locations of either one green square (low load) or three green squares (high load) within an invisible 3 × 3 grid positioned in the middle of the screen. The grid spanned 4o × 4o, and individual squares were 0.6o × 0.6o. The squares were presented for 500 ms, followed by a blank screen (200 ms) and then the vernier display. After participants made a response to the vernier task, they were presented with a green probe square for 200 ms and were asked to indicate whether the square was in the same location as that seen at the beginning of the trial or in a different location. The participants were given 20 practice trials before performing a total of 360 test trials and took a break every 120 trials.

In Experiment 2, a spatial working memory load was added before the AAE task. Participants were given low (one item) or high (three items) spatial working memory load and were probed at the end of the trial with one square
Results and discussion
Trials on which an incorrect response was made to the visual working memory task or participants had an RT that exceeded 2.5 standard deviations from the mean on either the visual working memory or vernier task were excluded from analysis. Six participants were replaced for having accuracies below 75% on the spatial working memory task or vernier judgment task. After replacing these participants, average accuracy on the spatial working memory task was 94% under low load and 89% under high load.
Figure 4 shows the percentage of left responses to left, central, and right vernier lines, across the low and high spatial memory load conditions. Similar to Experiment 1, we confined analysis to those trials where the top line was centered, using a 2 (load: low or high) × 2 (cue: top left or top right) ANOVA. A significant effect of cue, F(1, 15) = 55.88, p < .001, ηp 2 = .788, p rep ≈ 1, revealed an ARE. No main effect of load was found, F(1, 15) < 1. Importantly, the interaction of load and cue was significant, F(1, 15) = 5.63, p = .03, ηp 2 = .273, p rep = .60, with a larger AAE in the low-load condition (M ± SE = .42 ± .06, Cohen's d = 1.94) than in the high-load condition(.33 ± .06, Cohen's d = 1.56). Thus, this experiment supports a role for spatial working memory in the AAE, since it appears that engaging this memory reduces the AAE.

Percentage of left responses on the AAE task under low and high spatial working memory load conditions (Experiment 2). Error bars represent 95% confidence intervals
Experiment 3
The previous experiment showed that the AAE is sensitive to spatial working memory loads, suggesting that the effect does partially rely on spatial working memory. It seems reasonable that the AAE selectively involves spatial working memory, but to confirm this, Experiment 3 repeats the previous experiment but with a feature (color) visual working memory load. If the AAE does rely specifically on spatial working memory, the color memory task should not modulate the attraction effect. If, on the other hand, the effect of load is not limited to spatial working memory, the AAE should be reduced under high visual feature memory load.
Method
Participants
Sixteen different participants (7 male) at the University of Toronto participated in this experiment for additional course credit. All participants indicated that they had normal or corrected-to-normal vision and provided consent to participate in the experiment.
Apparatus and procedure
The monitor and chinrest setup was identical to that in previous experiments. Participants were given either a low or a high color visual working memory load prior to the vernier judgment task of the AAE task (see Fig. 5). Following a white fixation cross spanning 0.5o × 0.5o, the color memory load task was presented on a black background for 500 ms at the beginning of each trial and consisted of one colored square (low load) or three differently colored squares (high load) in any of three possible positions (either in the center of the screen or 2o above or below the midline). Squares were 0.6o × 0.6o in size. Participants were asked to remember the colors of the squares. A 200-ms blank screen followed the offset of the memory display, after which participants were presented with the vernier display. Upon providing a response to the vernier task, participants were shown one colored square in any of the three positions for 200 ms and were asked to indicate whether that color was presented at the beginning of the trial. Possible colors for the squares included white, blue, green, red, yellow, purple, and turquoise. Participants were given 20 practice trials before completing 360 test trials, with a break every 120 trials.

In Experiment 3, participants were given a low (one item) or high (three items) color visual working memory load while performing the AAE task
Results and discussion
Trials on which an incorrect response was made to the color visual working memory task, an inappropriate key was pressed as a response, or RTs to the color visual working memory or vernier task exceeded 2.5 standard deviations of the mean were not considered for analysis. Three participants were replaced for having below 75% accuracy on either the color visual working memory or vernier judgment task. The remaining participants maintained good accuracy on the color visual working memory task, with 93% accuracy on low loads and 84% on high loads.
The percentages of left responses to left, central, and right vernier lines, across both low and high color visual working memory loads, are shown in Fig. 6. Similar to previous experiments, we confined the analysis to trials with the central vernier line with a 2 (load: low or high) × 2 (cue: top-left, bottom-right pair or top-right, bottom-left pair) ANOVA. There was a significant effect of cue, F(1, 15) = 86.077, p < .001, ηp 2 = .852, p rep ≈ 1, revealing an AAE. There was no effect of load, F(1, 15) = 1.32, p > .05, ηp 2 = .123, and, more important, no interaction between load and cue, F(1, 15) < 1, JZS Bayes factor = 4.16. The magnitudes of the AAE under low and high load were, respectively, .49 (SE = .06, Cohen's d = 2.32) and .47 (SE = .06, Cohen's d = 2.11).

Percentage of left responses under conditions of low (one item) and high (three items) color visual working memory load (Experiment 3). Error bars denote 95% confidence intervals
General discussion
The aim of the present research was to examine whether the AAE and the ARE are driven by changes in the same visual processes and, if not, identify what does underlie the AAE. While both the ARE and the AAE share the commonality in that they are both driven by shifts of attention (Au, Ono, & Watanabe, 2013; Ono & Watanabe, 2011; Suzuki & Cavanagh, 1997), it seems that the AAE cannot be driven solely by changes to the RFs of early visual cells (in contrast to the ARE). By employing the same interocular manipulation used by DiGiacomo and Pratt (2012) that was able to eliminate the ARE by preventing cue and target information from interfering with each other at early stages of visual processing, we found in Experiment 1 that the AAE continued to persist.Footnote 3 Previous findings that the ARE and AAE produced summative effects upon consecutive presentation and the mitigation of the AAE, but not the ARE, by reducing the magnitude of attentional shifts (Chien et al., 2011) also support the notion that the AAE and the ARE rely on different processes. The differential time course of these two phenomena also highlights their differences, with the ARE existing at SOAs between 100 and 200 ms and AAE begining to emerge at SOAs of greater than 600 ms (Yamada et al., 2011). To begin exploring what produces the AAE, we tested the hypothesis first proposed by Ono and Watanabe that biased memory traces may be involved in the process. Specifically, we manipulated the delay before the localization response, to examine whether the effect is susceptible to memory distortions over time in working memory. Our findings of the increased magnitude of AAE over a longer period of delay support the notion that a memory bias might indeed be involved. To confirm that the AAE relies on visuospatial working memory, we manipulated spatial memory load in Experiment 2. An increased spatial memory load attenuated the strength of the AAE, providing evidence that the AAE relies on visuospatial memory resources. To ensure that the reduced magnitude of the AAE was not simply due to increased cognitive load, Experiment 3 employed a nonspatial working memory task (in this case, memory for colors), which did not modulate the AAE.Footnote 4 We conclude that the AAE does not share the same mechanism that produces the ARE and is, instead, dependent on spatial working memory resources.
Previous work suggests that attentional orienting is necessary for the AAE, since the effect is obtained only when the cues are attended (Au et al., 2013; Chien et al., 2011; Ono & Watanabe, 2011). Of course, that is not to say that attentional orienting is sufficient for obtaining the AAE. Instead of eliminating AAE, our viewing manipulation (interocular presentation) and occupying spatial working memory both only attenuated the AAE, suggesting that other, lower-level processes underlie this effect. Nonetheless, we demonstrated the involvement of a spatial working memory representation of the targets in the later stages of visual processing. It is at these later stages that mechanisms that compare and integrate locations may produce the AAE. It is possible that the cues served as landmarks toward which participants consistently biased the location of the target in the encoding process or averaged to a location between the cue and the true position of the vernier (Hubbard & Ruppel, 2000; Sheth & Shimojo, 2001; Yamada, Kawabe, & Miura, 2008). For instance, Zimmerman, Fink and Cavanagh (2013) reported that a peripheral stimulus was consistently attracted toward a more foveal anchor, regardless of the temporal order between the two. They proposed that this was the result of summing the neural response of the locations of the anchor and target, with the more foveal component carrying additional weight, due to cortical magnification. Through either of these processes, a distorted memory representation of the target being closer to the cue is formed.
In addition, Sheth and Shimojo (2001) proposed that the memory of the spatial relationship between two objects may decay over time, causing the remembered distance to be shorter at the time of response than at presentation. They postulated that this error arises during transfer of information from retinal coordinates stored more accurately in iconic memory to the relatively longer lasting but less accurate working memory storage. Supporting this, Sheth and Shimojo found that localization errors were reduced when participants responded after a period of 200 ms, as compared with 500 ms and 2 s, noting that items exist in iconic memory up to a maximum of 500 ms (Sperling, 1960). Thus, it is also possible that the AAE may arise from a deteriorated representation within the visual working memory system.
The AAE could also be explained by the possible involvement of motion signals. Although Suzuki and Cavanagh (1997) showed that the ARE was not due to apparent motion, this has not been demonstrably refuted with respect to the AAE. Chien et al. (2011) acknowledged this possibility, suggesting that the perceived location of the target could have been retrospectively influenced by the perceived motion of the cue. As a reviewer helpfully pointed out, this phenomenon has been previously referred to as the Fröhlich effect, where the initial position of a stimulus that starts moving from an unseen position is mislocalized in the direction of the motion (Fröhlich, 1923). Since both the AAE and Fröhlich effect produce mislocalizations in the same direction with similar stimuli present, it may be prudent to consider the processes underlying the Fröhlich effect. Similar to the AAE, attention plays a role, since the mislocalization in the Fröhlich effect has been attributed to the time it takes neural processing to allocate attention to the moving object and, in the midst of this, this process does not extrapolate completely the trajectory of the object to correctly determine its initial position (Hubbard & Motes, 2005; Müsseler & Aschersleben, 1998). Kirschfeld and Kammer (1999) have proposed that the Fröhlich effect is, in fact, the interaction between focal attention and metacontrast processes, such that the backward masking makes it difficult for the initial position to be perceived. These attention models suggest that better allocation of attention can attenuate the Fröhlich illusion. On the other hand, other models suggest that the disruption of spatiotemporal integration processes underlie the illusion. Eagleman and Sejnowski (2000) have proposed that unexpected external events render the existing internal representation inaccurate and integration of the unexpected event interrupts its continuity. As a result, a new integration process must take place, and the delay in “resetting” the internal representation causes it to be temporally offset from the actual onset of the external event. Further studies should attempt to tease apart the attentional and nonattentional components to distinguish between the AAE and the Fröhlich effect or determine whether they are in fact the same. Regardless, our findings show that spatial working memory does play a role in at least the AAE.
Most research on attention has focused on what happens when attention precedes the stimuli, but the AAE phenomenon illustrates how attention can retroactively affect the representation of preceding stimuli. Moreover, its basis does not appear to be as straightforward as the ARE, since visual working memory appears to be involved. The findings of this study persuade us to support the suggestion of Ono and Watanabe (2011) that attentional phenomena need to be studied in the context of time. In the particular case of the ARE and AAE, it is interesting that the differential time course of attentional shifts can produce such drastically different spatial consequences, causing attentional biases at distinct stages of visual processing of targets.
Notes
p rep (probability of replication) refers to the observed statistical power (1 − ß).
The JZS Bayes factor (Rouder, Speckman, Sun, Morey, & Iverson, 2009) estimates the likelihood ratio of the null hypothesis over alternative hypotheses (e.g., Bayes factor of 2 indicates that the null effect is twice more likely than its rejection). We report this estimate only for the null effects that carry theoretical importance.
Although we may not have detected a reduction in AAE magnitude in the interocular condition due to statistical power, the relevant point is that AAE did persist under interocular condition. Therefore, despite the potential role of early visual RFs in AAE, this effect cannot be exclusively attributed to early visual processes.
It is worth noting that the same memory load manipulations have not yet been applied to the ARE paradigm. Although whether or not the ARE will be affected by memory load remains an empirical question, we speculate that the ARE will be impervious to memory manipulations, due to its lower-level origin (DiGiacomo & Pratt, 2012).
References
Au, R. K., Ono, F., & Watanabe, K. (2013). Spatial distortion induced by imperceptible visual stimuli. Consciousness and Cognition, 22(1), 99–110.
Carrasco, M. (2011). Visual attention: The past 25 years. Vision Research, 51(13), 1484–1525.
Chien, S. E., Ono, F., & Watanabe, K. (2011). Mislocalization of Visual Stimuli: Independent Effects of Static and Dynamic Attention. PloS One, 6(12), e28371.
Desimone, R., & Duncan, J. (1995). Neural mechanisms of selective visual attention. Annual Review of Neuroscience, 18(1), 193–222.
DiGiacomo, A., & Pratt, J. (2012). Misperceiving space following shifts of attention: Determining the locus of the attentional repulsion effect. Vision Research, 64, 35–41.
Eagleman, D. M., & Sejnowski, T. J. (2000). Motion integration and postdiction in visual awareness. Science, 287(5460), 2036–2038.
Folk, C. L., Remington, R. W., & Johnston, J. C. (1992). Involuntary covert orienting is contingent on attentional control settings. Journal of Experimental Psychology: Human Perception and Performance, 18, 1030–1044.
Fortenbaugh, F. C., Prinzmetal, W., & Robertson, L. C. (2011). Rapid changes in visual-spatial attention distort object shape. Psychonomic Bulletin & Review, 18(2), 287–294.
Fröhlich, F. W. (1923). Überr die Messung der Empfindungszeit. Zietschrift für Sinnesphysiologie, 54, 58–78.
Gozli, D. G., & Pratt, J. (2012). Attentional repulsion effect despite a colour-based control set. Visual Cognition, 20(6), 696–716.
Henderson, J. M. (1991). Stimulus discrimination following covert attentional orienting to an exogenous cue. Journal of Experimental Psychology: Human Perception and Performance, 17(1), 91–106.
Hubbard, T. L., & Motes, M. A. (2005). An effect of context on whether memory for initial position exhibits a Fröhlich effect or an onset repulsion effect. The Quarterly Journal of Experimental Psychology Section A, 58(6), 961–979.
Hubbard, T. L., & Ruppel, S. E. (2000). Spatial memory averaging, the landmark attraction effect, and representational gravity. Psychological Research, 64(1), 41–55.
Kirschfeld, K., & Kammer, T. (1999). The Fröhlich effect: a consequence of the interaction of visual focal attention and metacontrast. Vision Research, 39(22), 3702–3709.
Kosovicheva, A. A., Fortenbaugh, F. C., & Robertson, L. C. (2010). Where does attention go when it moves? Spatial properties and locus of the attentional repulsion effect. Journal of Vision, 10(12), 1–13.
Müller, H. J., & Rabbit, P. M. (1989). Spatial cuing and the relation between the accuracy of "where" and "what" decisions in visual search. Quarterly Journal of Experimental Psychology, 41(4), 747–773.
Müsseler, J., & Aschersleben, G. (1998). Localizing the first position of a moving stimulus: the Frohlich effect and an attention-shifting explanation. Perception Psychophysics, 60(4), 683–695.
Ono, F., & Watanabe, K. (2011). Attention can retrospectively distort visual space. Psychological Science, 22(4), 472–477.
Posner, M. I., Snyder, C. R., & Davidson, B. J. (1980). Attention and the detection of signals. Journal of Experimental Psychology: General, 109(2), 160–174.
Pratt, J., & Arnott, S. R. (2008). Modulating the attentional repulsion effect. Acta Psychologica, 127(1), 137–145.
Pratt, J., & McAuliffe, J. (2001). The effects of onsets and offsets on visual attention. Psychological Research, 65(3), 185–191.
Pratt, J., & Turk-Browne, N. B. (2003). The attentional repulsion effect in perception and action. Experimental Brain Research, 152(3), 376–382.
Rouder, J. N., Speckman, P. L., Sun, D., Morey, R. D., & Iverson, G. (2009). Bayesian t tests for accepting and rejecting the null hypothesis. Psychonomic Bulletin & Review, 16(2), 225–237.
Schneider, K. A., & Komlos, M. (2008). Attention biases decisions but does not alter appearance. Journal of Vision, 8(15), 1–10.
Sheth, B. R., & Shimojo, S. (2001). Compression of space in visual memory. Vision Research, 41(3), 329–341.
Sperling, G. (1960). The information available in brief visual presentations. Psychological Monographs, 74(11), 1–29.
Stelmach, L. B., & Herdman, C. M. (1991). Directed attention and perception of temporal order. Journal of Experimental Psychology: Human Perception and Performance, 17(2), 539–550.
Suzuki, S., & Cavanagh, P. (1997). Focused attention distorts visual space: An attentional repulsion effect. Journal of Experimental Psychology: Human Perception and Performance, 23(2), 443–463.
Theeuwes, J. (2010). Top–down and bottom–up control of visual selection. Acta Psychologica, 135(2), 77–99.
Toba, M. N., Cavanagh, P., & Bartolomeo, P. (2011). Attention biases the perceived midpoint of horizontal lines. Neuropsychologia, 49(2), 238–246.
Tsal, Y., & Bareket, T. (1999). Effects of attention on localization of stimuli in the visual field. Psychonomic Bulletin & Review, 6(2), 292–296.
Tsal, Y., & Shalev, L. (1996). Inattention magnifies perceived length: The attentional receptive field hypothesis. Journal of Experimental Psychology: Human Perception & Performance, 22(1), 233–243.
Yamada, Y., Kawabe, T., & Miura, K. (2008). Mislocalization of a target toward subjective contours: Attentional modulation of location signals. Psychological Research, 72(3), 273–280.
Yamada, Y., Miura, K., & Kawabe, T. (2011). Temporal course of position shift for a peripheral target. Journal of Vision, 11(6), 1–12.
Yantis, S., & Hillstrom, A. P. (1994). Stimulus-driven attentional capture: Evidence from equiluminant visual objects. Journal of Experimental Psychology: Human Perception and Performance, 20(1), 95–107.
Yeshurun, Y., & Carrasco, M. (1999). Spatial attention improves performance in spatial resolution tasks. Vision Research, 39(2), 293–306.
Zimmerman, E., Fink, G., & Cavanagh, P. (2013). Perifoveal spatial compression. Journal of Vision, 13(5), 1–9.
Acknowledgments
This study was supported by an Undergraduate Student Research Award (USRA) from the Natural Sciences and Engineering Research Council of Canada (NSERC) awarded to A.C., an NSERC discovery grant awarded to J.P., and an NSERC doctoral scholarship awarded to D.G.G.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Chow, A., Gozli, D.G. & Pratt, J. Examining the locus of the attentional attraction effect. Atten Percept Psychophys 76, 2389–2397 (2014). https://doi.org/10.3758/s13414-014-0724-1
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-014-0724-1