Aquatic Plant Dynamics in Lowland River Networks: Connectivity, Management and Climate Change

Abstract
:1. Introduction
- (1)
- (2)
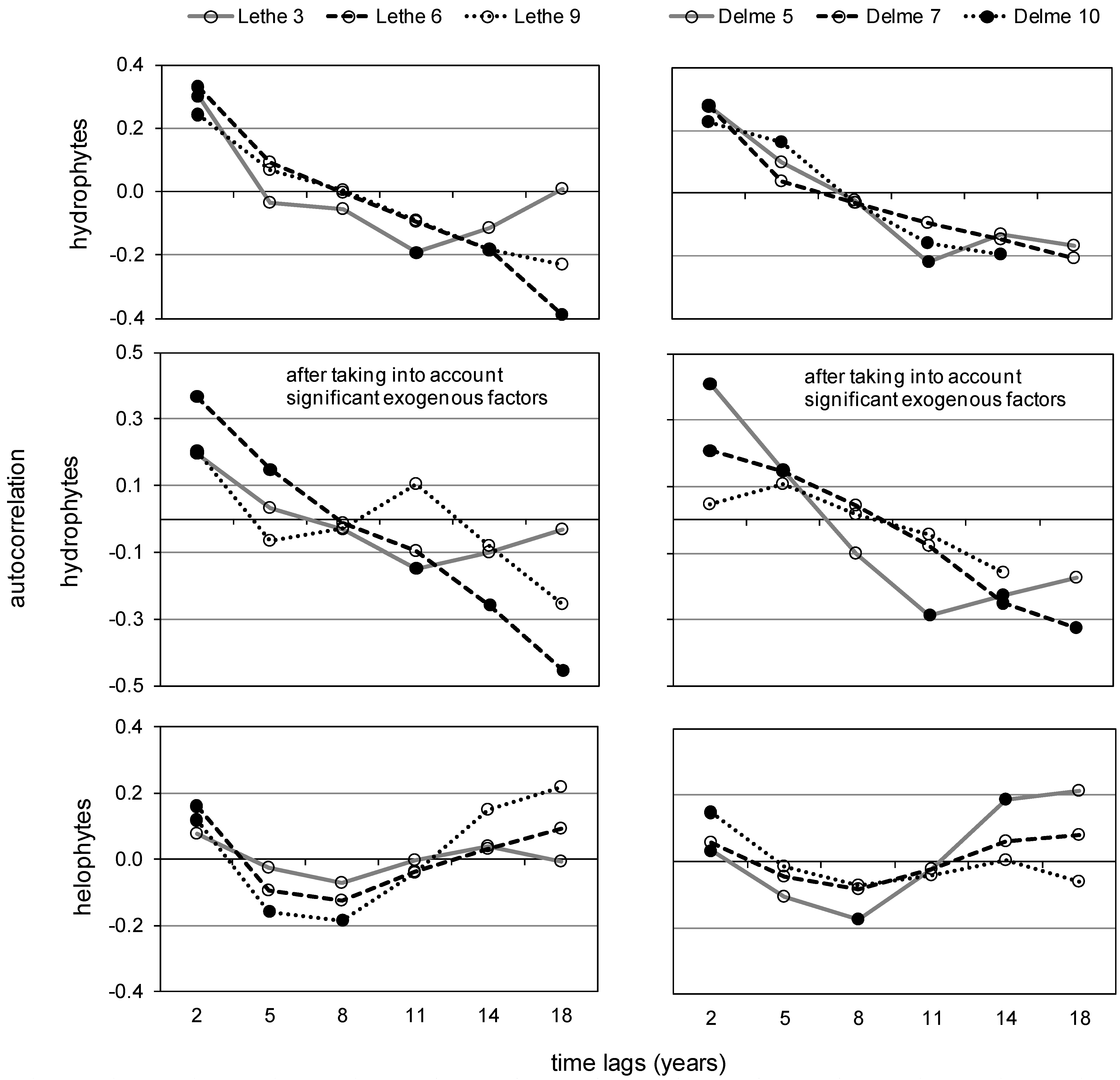
- Yearly autocorrelation in species composition will decrease over time, but will stay positive in the absence of changes in the dynamics of exogenous factors (underlying deterministic gradient). Temporal negative autocorrelation would have to result from “catastrophic” changes (shift in dominant species)—unexpected here [60];
- (3)
- (4)
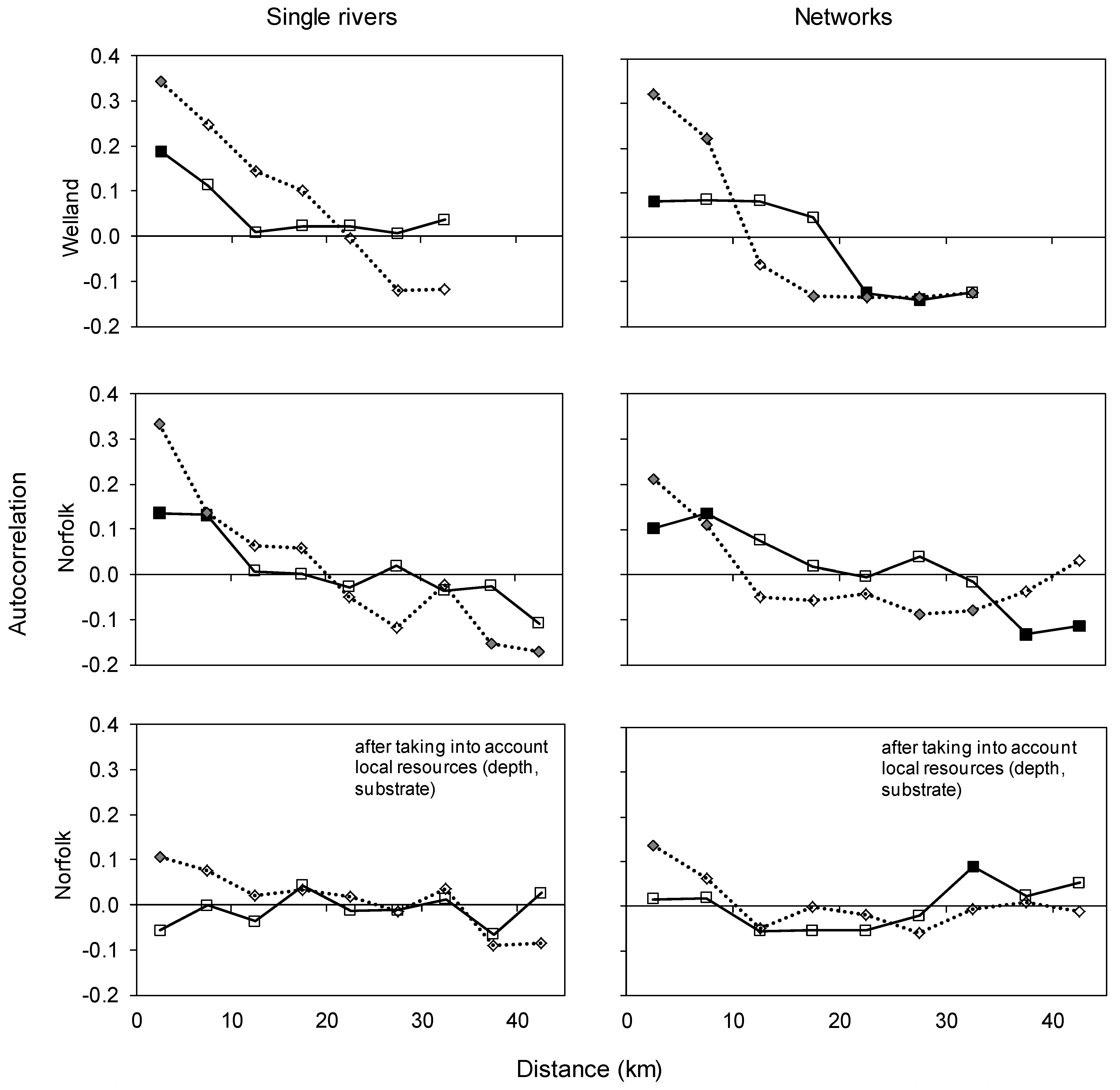
- At longer spatial intervals (tens of km) along the main stem, autocorrelation will decrease and possibly become negative due to differences in local resources creating river zonation. Trends in spatial autocorrelation of species composition after taking into account local resources will be weaker, but more reliably linked to plant dispersal abilities [43];
- (5)
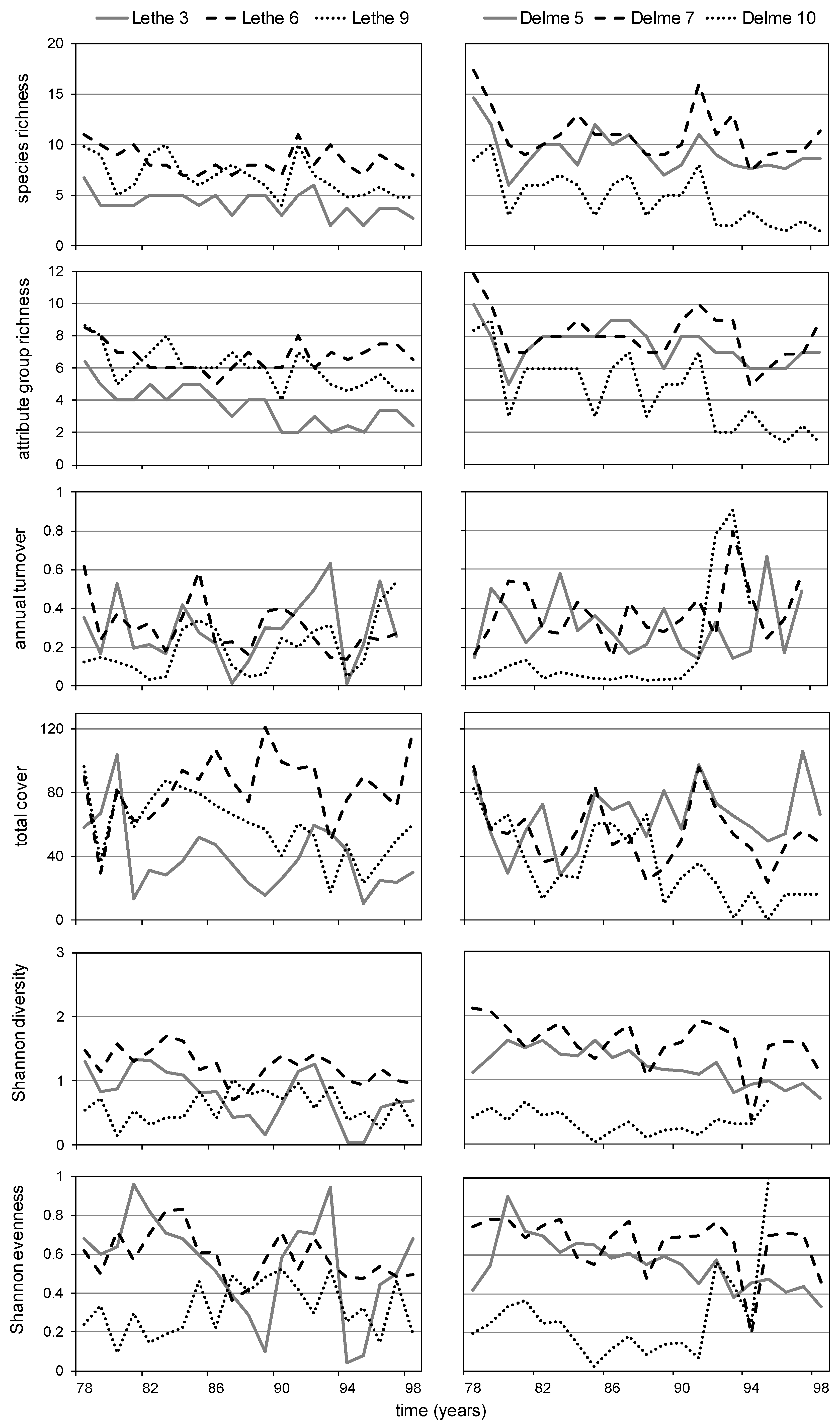
- Richness, cover, diversity, evenness and abundance patterns will fluctuate slightly around a mean value over time (in years) in the absence of changes in the dynamics of exogenous factors [60];
- (6)
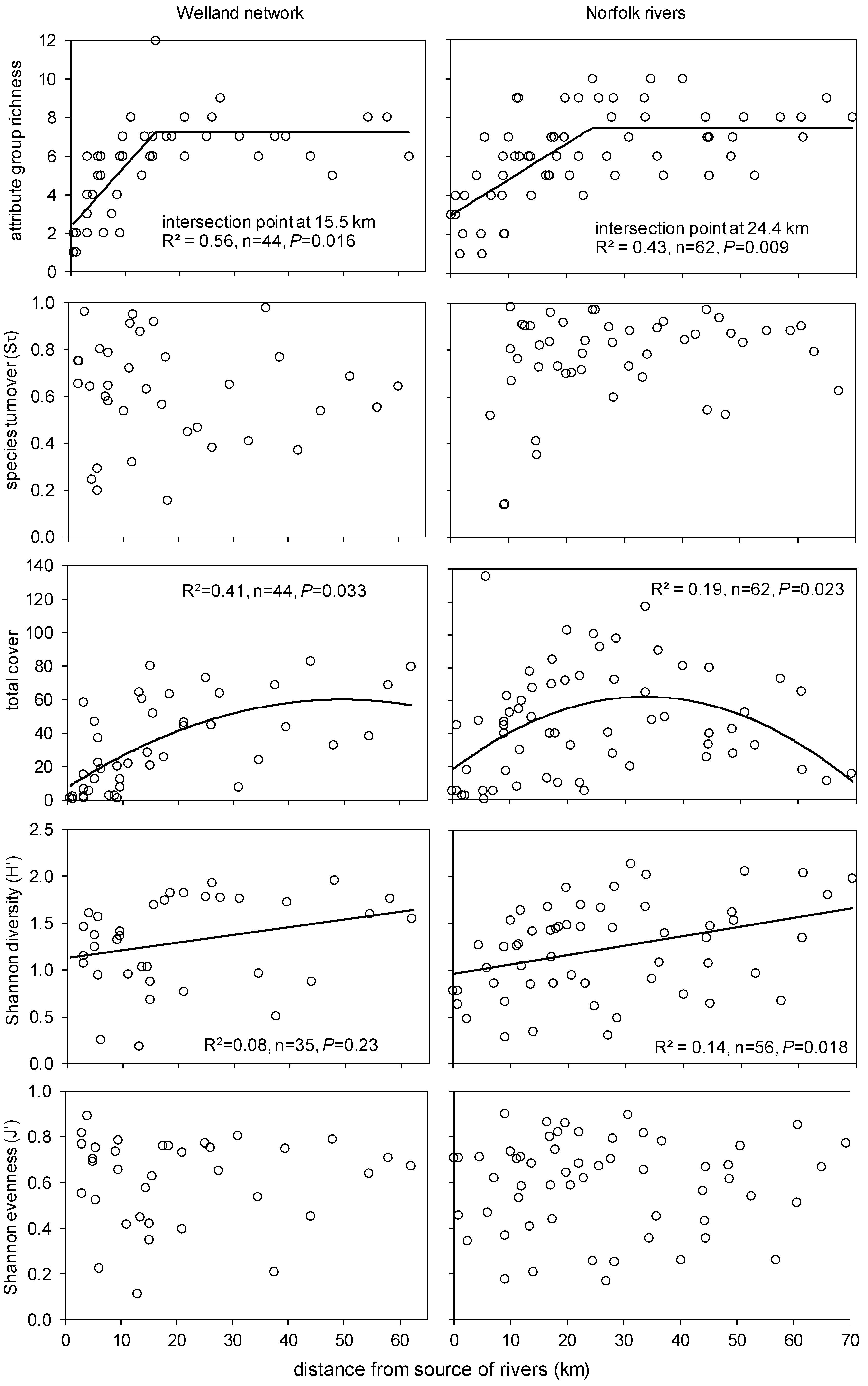
- Species richness increases along individual rivers for strict hydrophytes but not for marginal plants due to a sampling artefact (edge effect);
- (7)
- With richness increasing along rivers (species packing), we expect an increase in evenness and species trait diversity (i.e., attribute groups, sensu Willby et al. [68]);
- (8)
- If biotic gradients (change with time or distance in species composition and community structure) are observed, deterministic exogenous factors can explain them, such as change in climate (changes in magnitude, timing and frequency of high and low flow events, ice scouring, high temperature), management practices (weed cutting, riparian maintenance), biotic competitors (cover of green algae), depth and substrate;
- (9)
- (10)
- (11)
- The increase in hydrophyte richness with distance from source will be less pronounced at network scale than at individual site scale due to dispersal constraints (isolation), especially in the headwaters [62];
- (12)
- Related to that, the regression slope of local hydrophyte abundance as a function of occurrence will be steeper along rivers than across the network [41].
2. Material and Methods
2.1. Study Areas
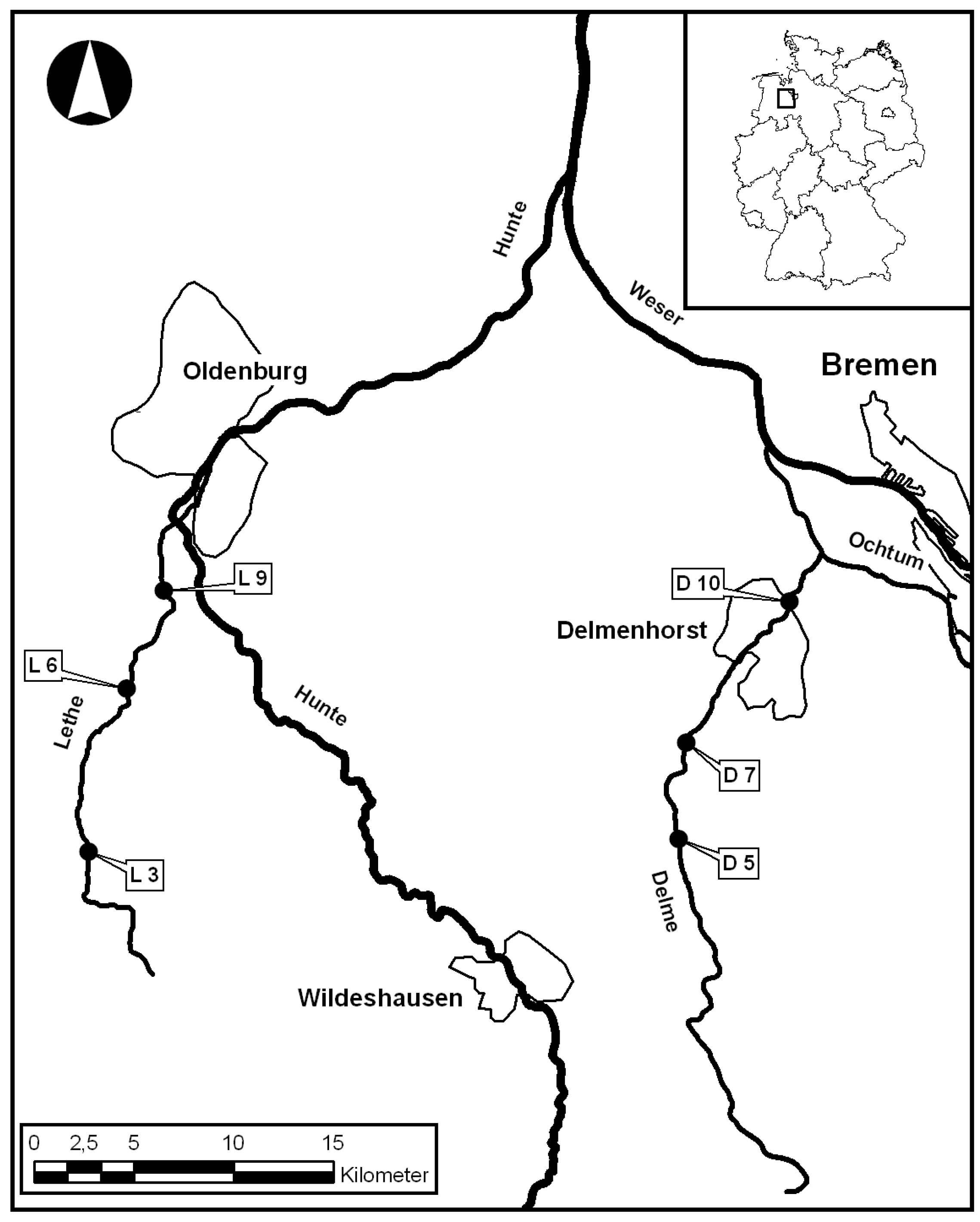
2.1.1. Rivers Lethe and Delme, Lower Saxony, Germany

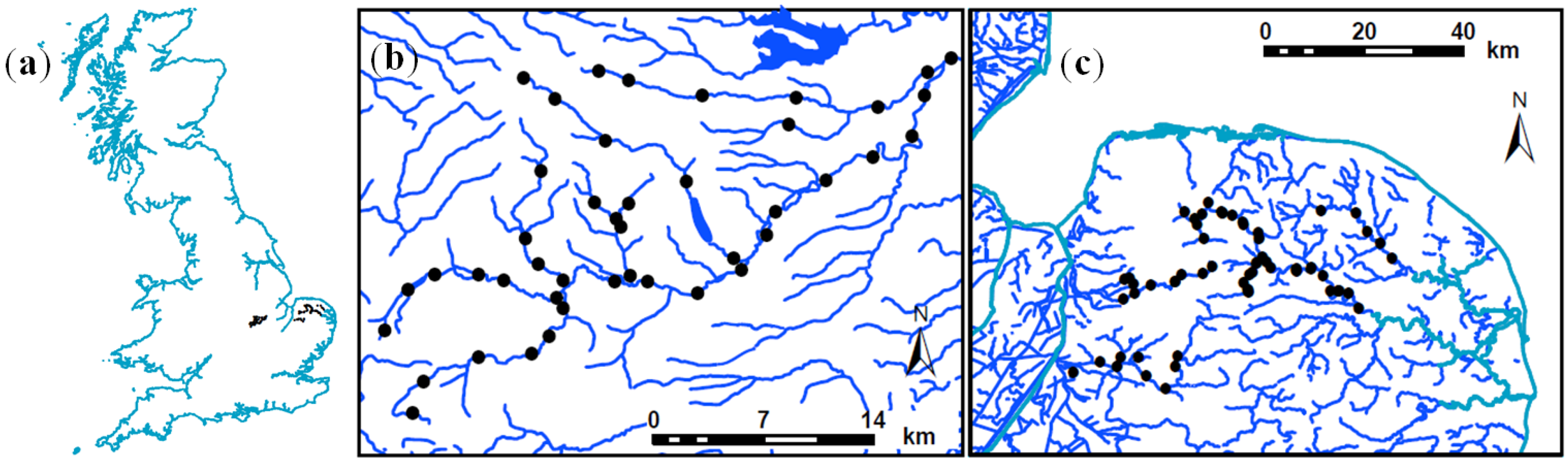
2.1.2. Norfolk Rivers, Norfolk, England
2.1.3. River Welland Network, East Midlands, England

2.2. Field Surveys

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Differences in | L3 | L6 | L9 | D5 | D7 | D10 |
|---|---|---|---|---|---|---|
| Species richness | 0.8 ± 0.3 | 1.0 ± 0.2 | 0.8 ± 0.2 | 2.6 ± 0.4 | 3.4 ± 0.5 | 1.5 ± 0.5 |
| Total cover | 20 ± 4 | 28 ± 6 | 27 ± 4 | 40 ± 5 | 33 ± 4 | 16 ± 5 |
| Attribute group richness | 0.4 ± 0.2 | 0.5 ± 0.2 | 0.6 ± 0.2 | 2.0 ± 0.4 | 1.9 ± 0.4 | 1.4 ± 0.4 |
2.3. Community Structure Indices
2.3.1. Individual Species Cover
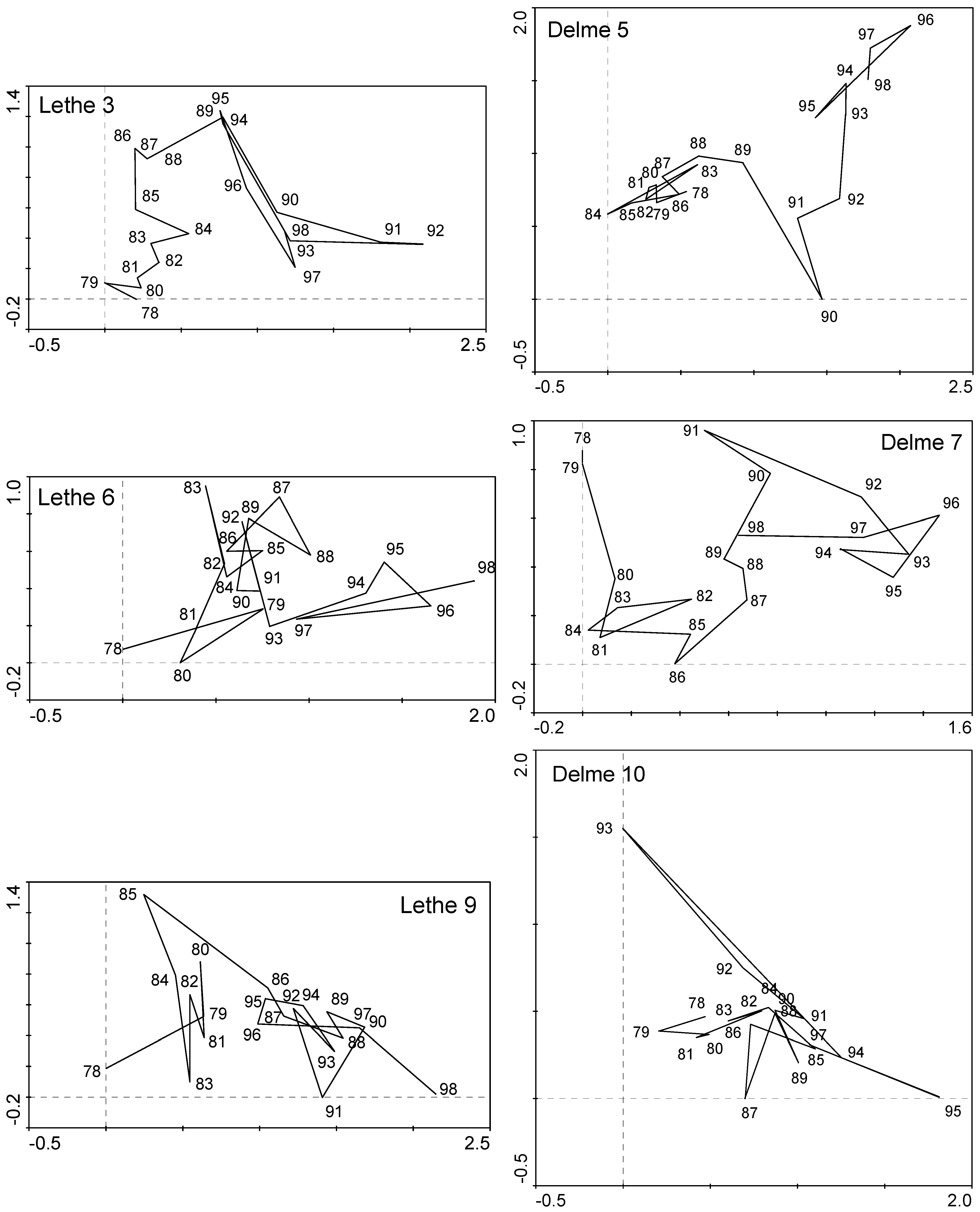
2.3.2. Unconstrained Ordinations
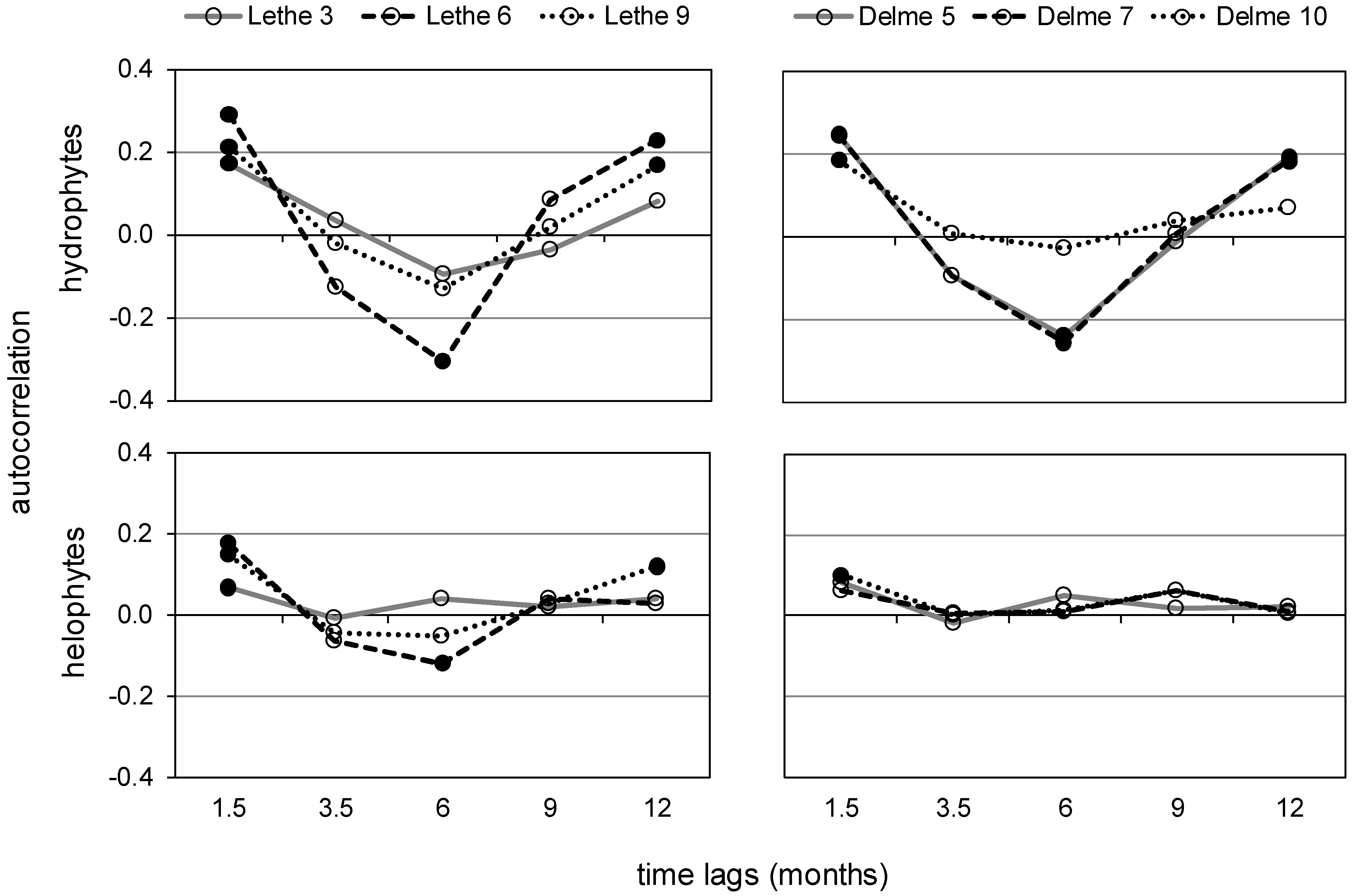
2.3.3. Autocorrelation
2.3.4. Richness, Total Cover and Turnover

2.3.5. Shannon Diversity (H') and Evenness (J')


2.3.6. Species Range-Abundance Patterns
2.4. Environmental Variables
2.4.1. Rivers Lethe and Delme
2.4.2. River Welland and Norfolk Rivers
2.5. Linking Vegetation with Time and Environmental Data
2.6. Statistical Analyses for Spatial Patterns
3. Results
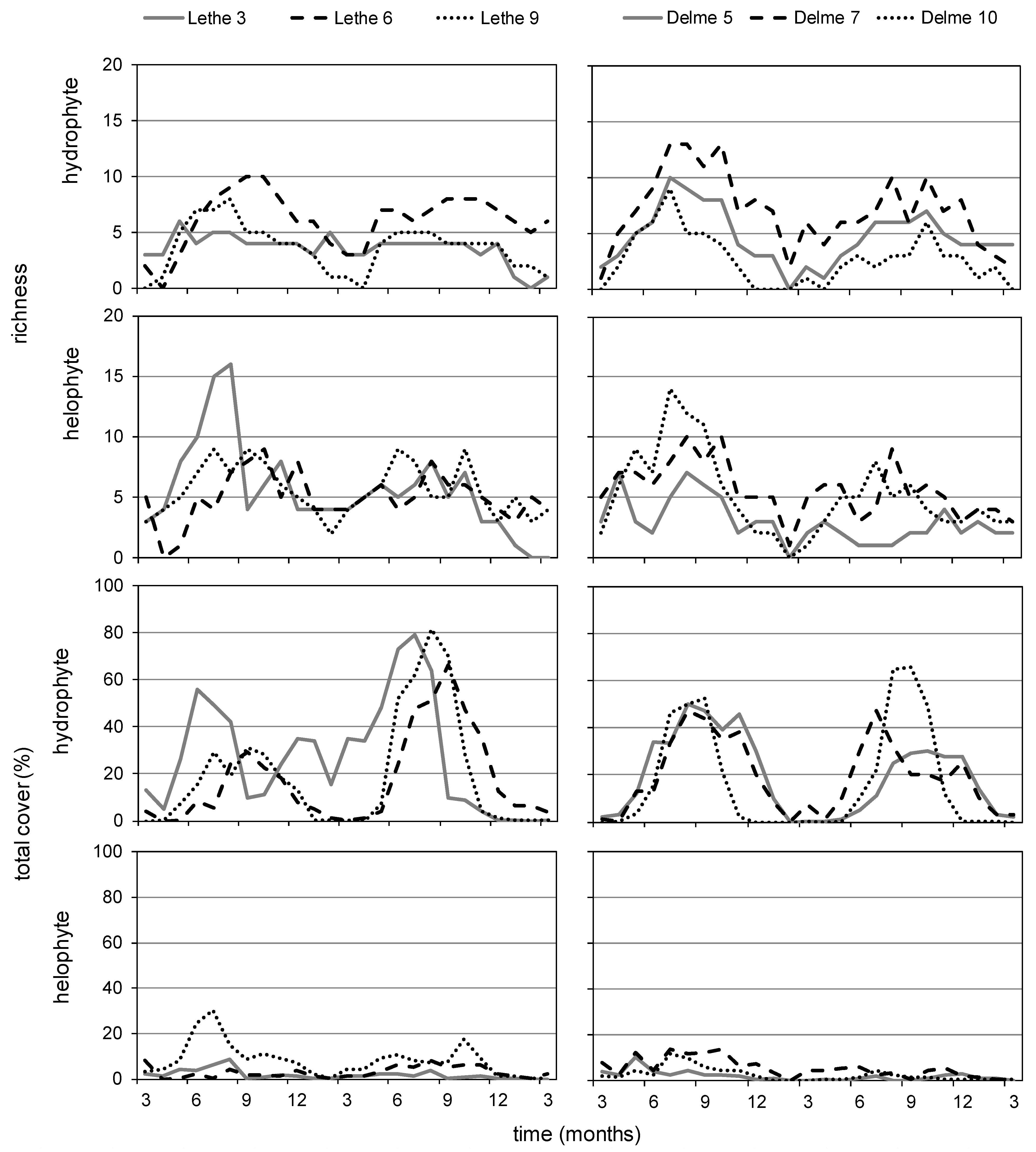
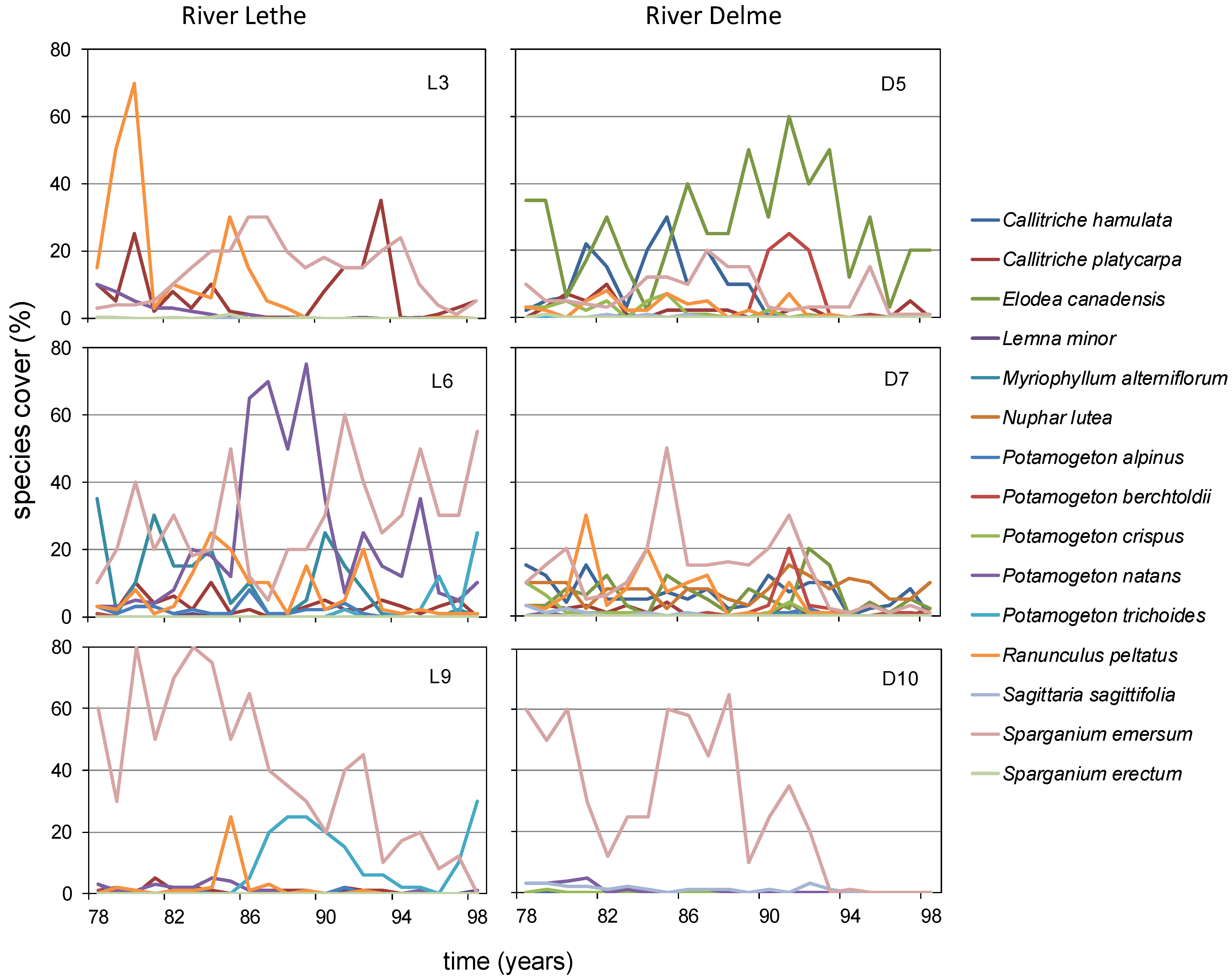
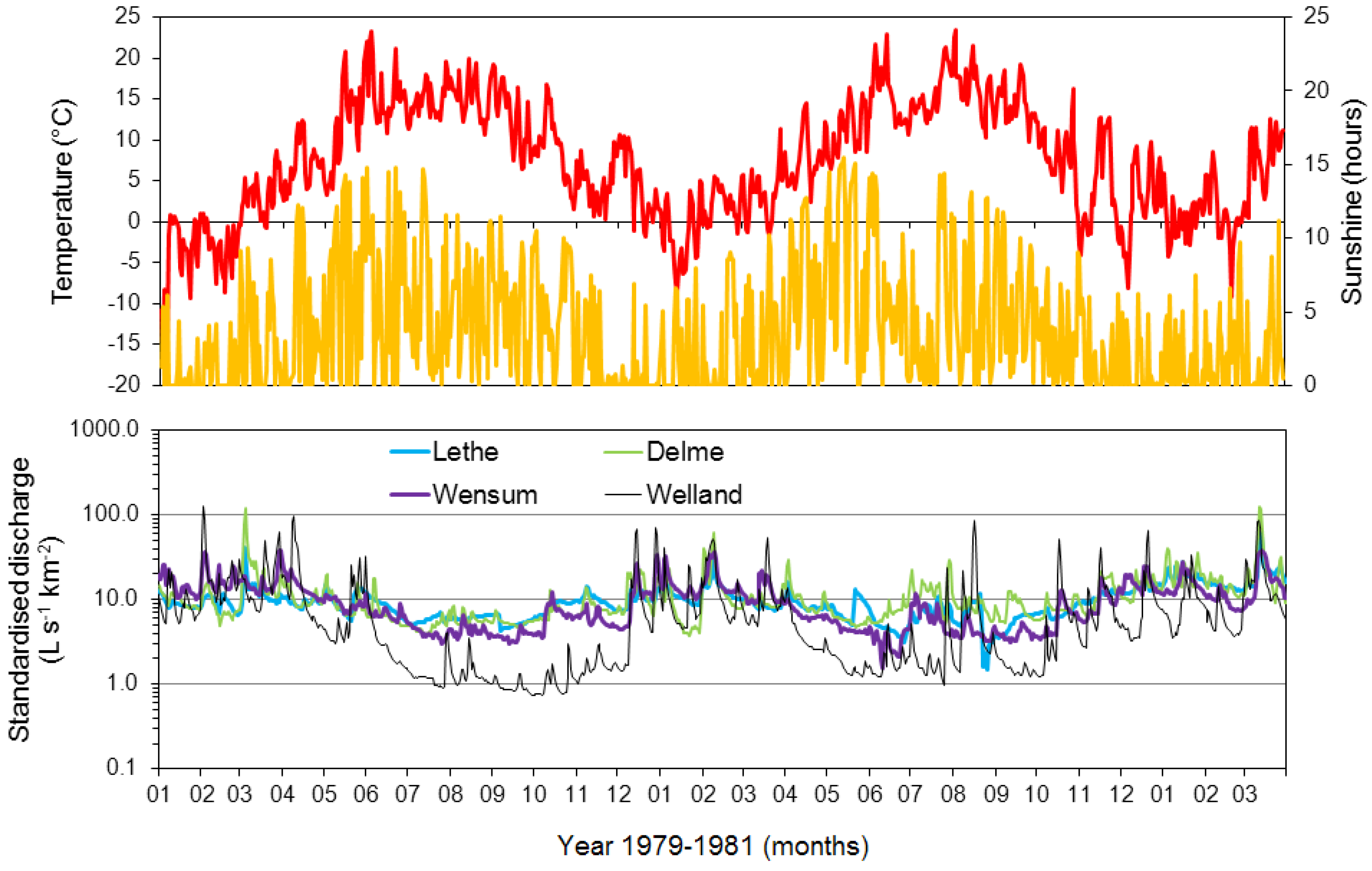
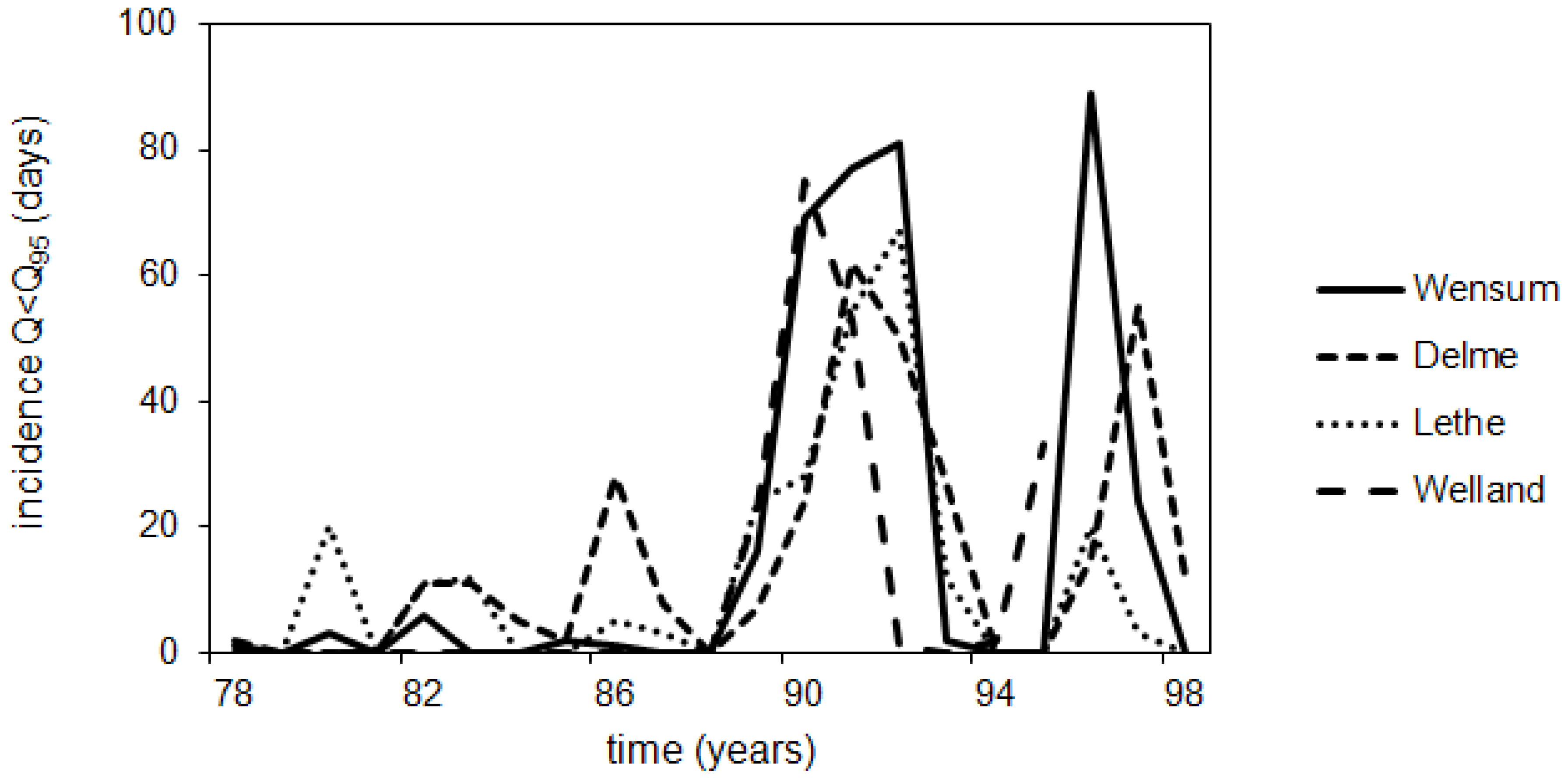
3.1. Lethe-Delme Monthly Changes over Two Years

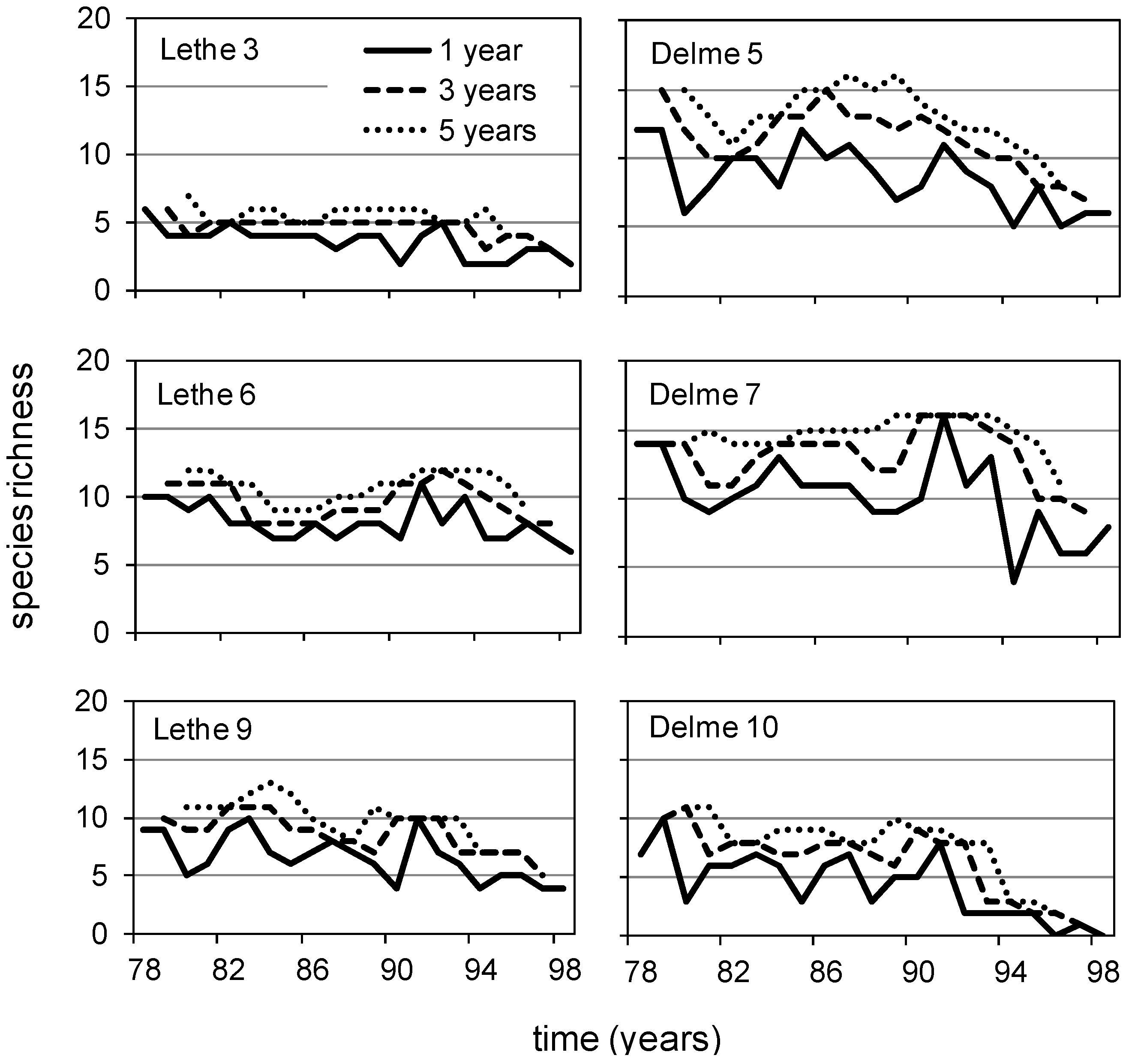
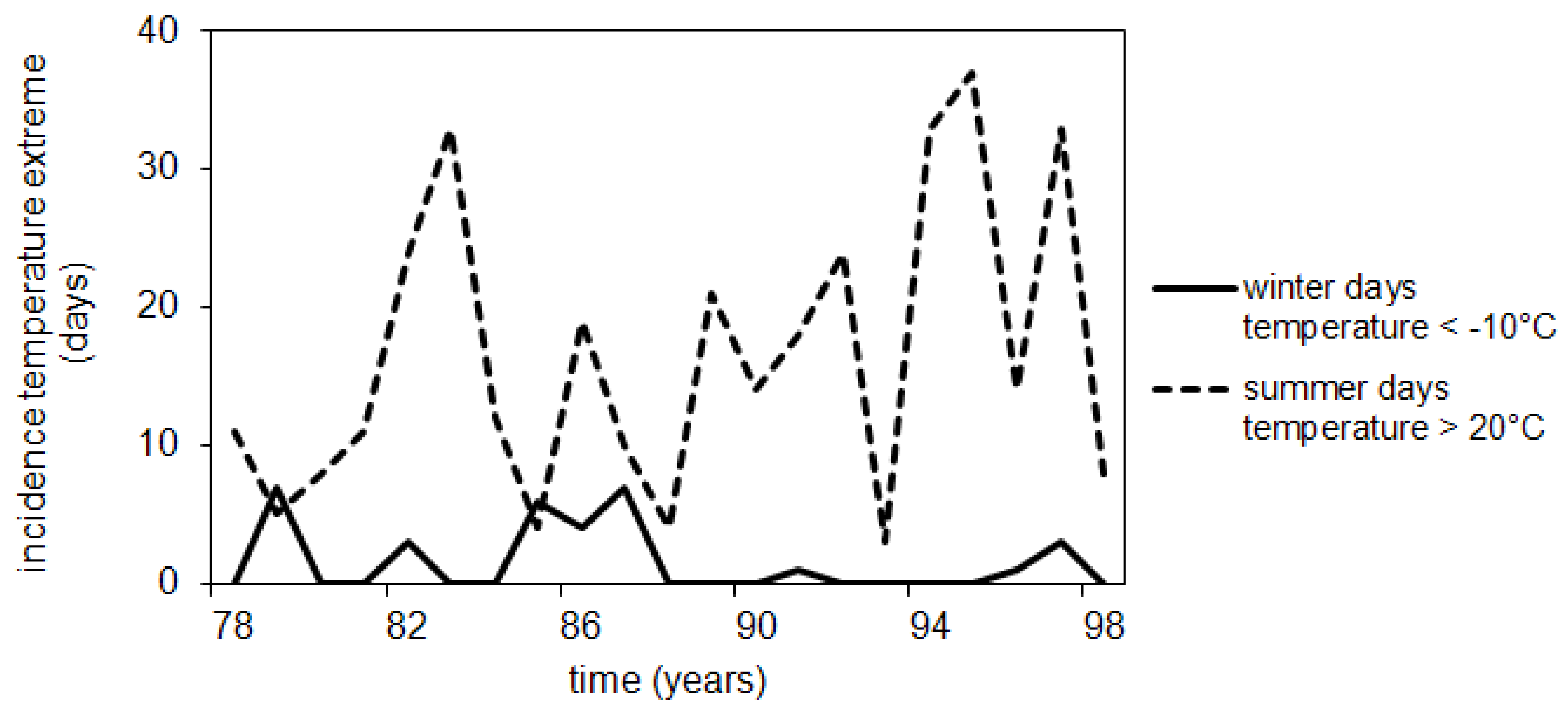
3.2. Lethe-Delme Temporal Changes over 21 Consecutive Years






| Traits | Attributes | Time | Shade | T-10 °C | Q95 |
|---|---|---|---|---|---|
| growth form | free floating surface | −0.20 | 0.01 | 0.08 | −0.02 |
| free floating submerged | 0.11 | 0.00 | 0.01 | 0.13 | |
| anchored, floating leaves | −0.55 | −0.12 | 0.21 | −0.01 | |
| anchored, submerged leaves | −0.39 | −0.07 | 0.15 | 0.11 | |
| anchored, emergent leaves | −0.58 | −0.11 | 0.25 | −0.08 | |
| anchored, heterophylly | −0.46 | −0.10 | 0.18 | 0.00 | |
| vertical shoot architecture | single apical growth point | −0.21 | −0.08 | 0.05 | 0.00 |
| single basal growth point | −0.43 | −0.06 | 0.14 | −0.05 | |
| multiple apical growth point | −0.26 | −0.05 | 0.12 | 0.14 | |
| leaf type | tubular | 0.00 | 0.00 | 0.00 | 0.00 |
| capillary | −0.42 | −0.10 | 0.20 | −0.01 | |
| entire | −0.39 | −0.07 | 0.17 | 0.10 | |
| leaf area | small (<1cm2) | −0.11 | −0.01 | 0.06 | 0.20 |
| medium (1–20 cm2) | −0.46 | −0.11 | 0.18 | −0.03 | |
| large (20–100 cm2) | −0.52 | −0.13 | 0.23 | −0.06 | |
| extra large (>100 cm2) | −0.16 | 0.00 | 0.03 | −0.03 | |
| morphology index (score) | 1 (2) | −0.20 | 0.01 | 0.08 | −0.02 |
| 2 (3–5) | −0.42 | −0.08 | 0.19 | 0.07 | |
| 3 (6–7) | −0.34 | −0.07 | 0.15 | 0.10 | |
| 4 (8–9) | −0.43 | −0.10 | 0.19 | 0.06 | |
| 5 (10) | −0.22 | −0.05 | 0.01 | 0.09 | |
| rooting at nodes | −0.29 | −0.07 | 0.15 | 0.09 | |
| high below-:above-ground biomass | −0.31 | −0.05 | 0.11 | −0.02 | |
| mode of reproduction | rhizome | −0.41 | −0.09 | 0.10 | 0.00 |
| fragmentation | −0.33 | −0.07 | 0.14 | 0.10 | |
| budding | −0.26 | −0.10 | 0.08 | −0.09 | |
| turions | −0.01 | 0.04 | 0.01 | 0.19 | |
| stolons | −0.48 | −0.07 | 0.15 | 0.00 | |
| tubers | −0.38 | 0.00 | 0.06 | −0.13 | |
| seeds | −0.44 | −0.08 | 0.16 | 0.09 | |
| number of reproductive organs/year/individual | low (<10) | 0.11 | 0.00 | 0.01 | 0.13 |
| medium (10–100) | −0.14 | 0.02 | 0.01 | 0.20 | |
| high (100–1000) | −0.48 | −0.09 | 0.19 | 0.04 | |
| very high (>1000) | −0.37 | 0.00 | 0.14 | −0.08 | |
| perennation | annual | −0.25 | −0.05 | 0.12 | 0.18 |
| biennial/short lived perennial | −0.47 | −0.13 | 0.31 | −0.07 | |
| perennial | −0.47 | −0.09 | 0.18 | 0.05 | |
| evergreen leaf | −0.37 | −0.09 | 0.20 | 0.04 | |
| amphibious | −0.52 | −0.10 | 0.18 | 0.02 | |
| gamete vector | wind | −0.35 | −0.05 | 0.09 | 0.14 |
| water | −0.02 | −0.01 | 0.04 | 0.14 | |
| air bubble | 0.23 | 0.06 | −0.12 | 0.33 | |
| insect | −0.33 | −0.08 | 0.21 | 0.00 | |
| self | −0.40 | −0.07 | 0.19 | 0.03 | |
| body flexibility | low (<45°) | −0.44 | 0.00 | 0.13 | −0.11 |
| intermediate (>45°–300°) | −0.22 | −0.02 | 0.13 | 0.08 | |
| high (>300°) | −0.50 | −0.10 | 0.16 | 0.06 | |
| leaf texture | soft | −0.45 | −0.07 | 0.16 | 0.10 |
| rigid | −0.35 | −0.10 | 0.18 | 0.02 | |
| waxy | −0.56 | −0.13 | 0.23 | −0.05 | |
| non-waxy | −0.38 | −0.06 | 0.15 | 0.13 | |
| period of production of reproductive organ | early (March–May) | −0.48 | −0.10 | 0.17 | −0.01 |
| mid (June–July) | −0.40 | −0.07 | 0.15 | 0.11 | |
| late (August–September) | −0.30 | −0.05 | 0.12 | 0.12 | |
| very late (post September) | 0.01 | 0.05 | −0.01 | 0.22 | |
| fruit size | <1 mm | 0.00 | 0.00 | 0.00 | 0.00 |
| 1–3 mm | −0.33 | −0.04 | 0.12 | 0.17 | |
| >3 mm | −0.31 | −0.07 | 0.14 | 0.04 | |
3.3. Wensum Temporal Changes over Three Consecutive Years
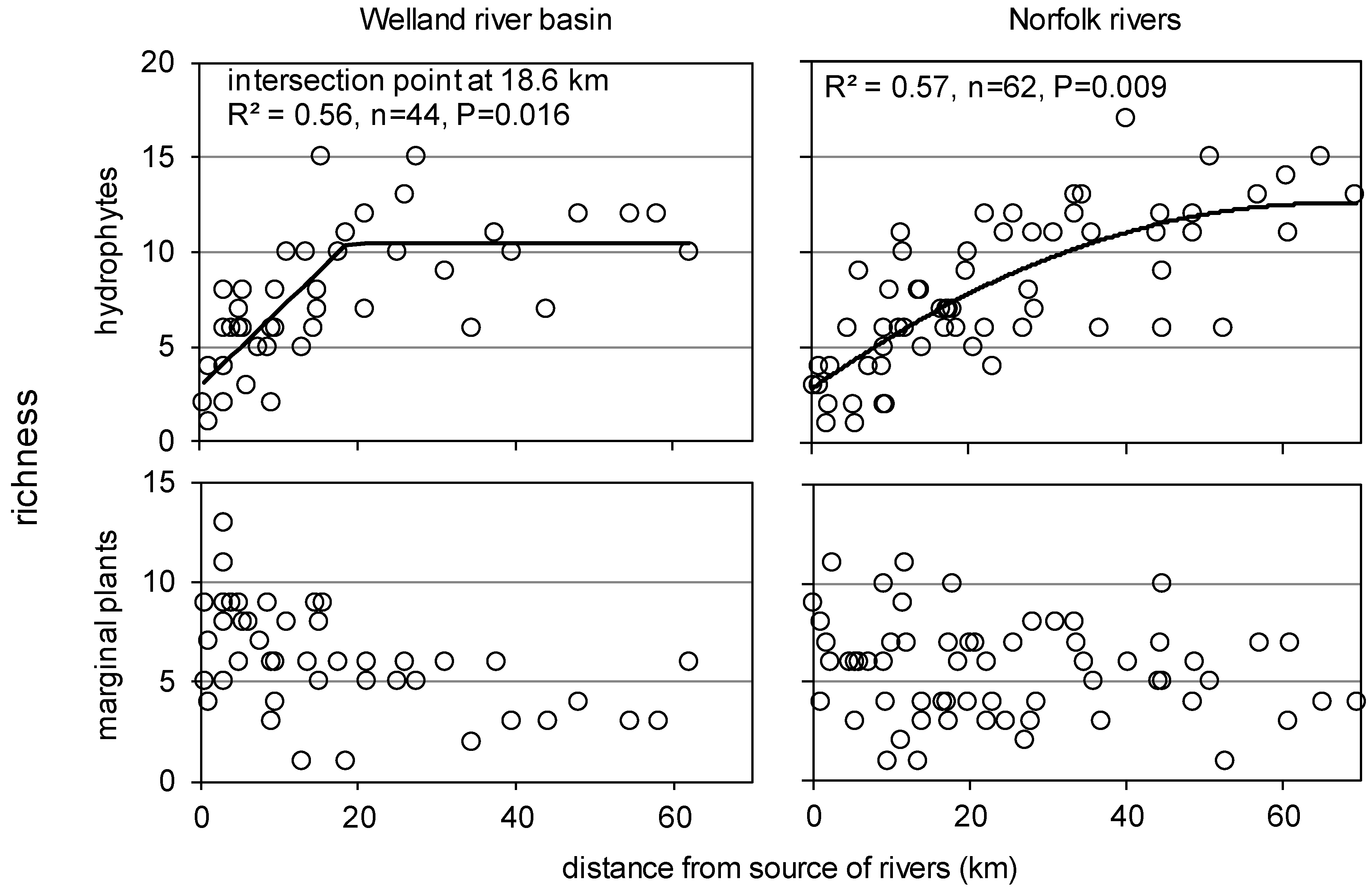
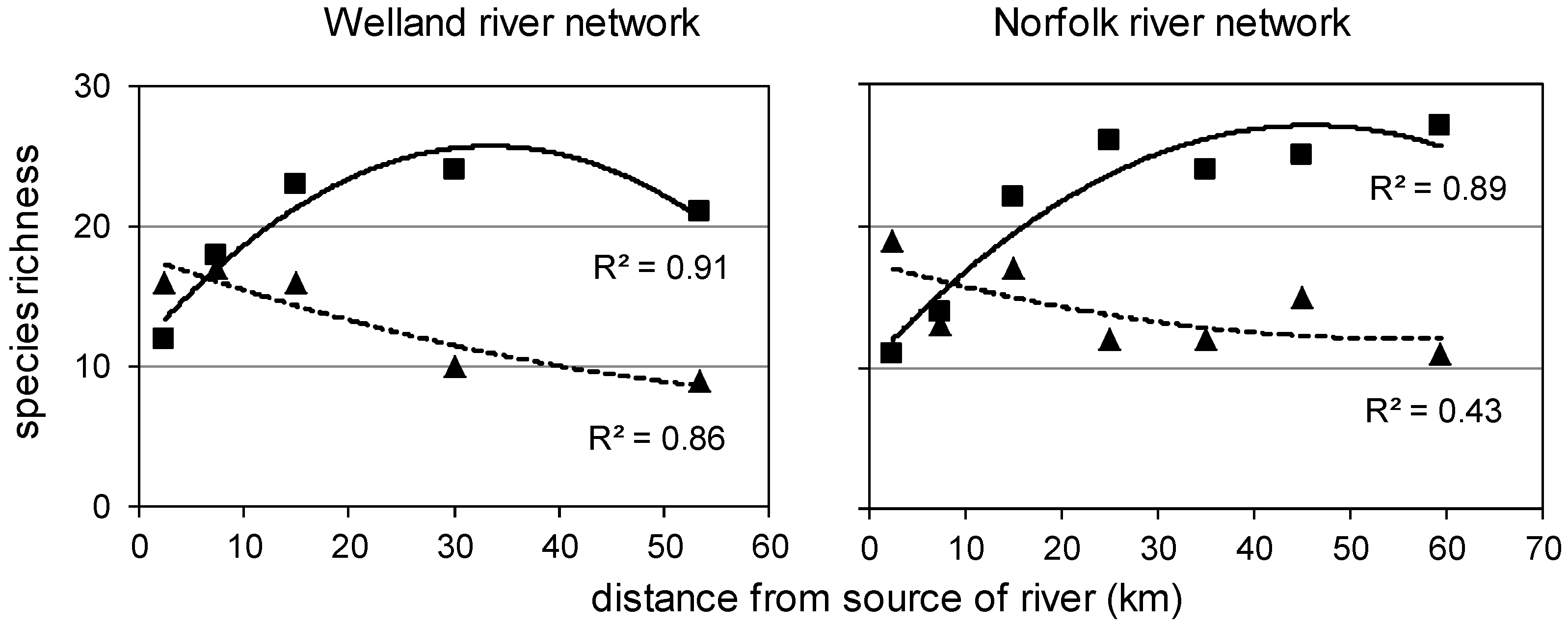
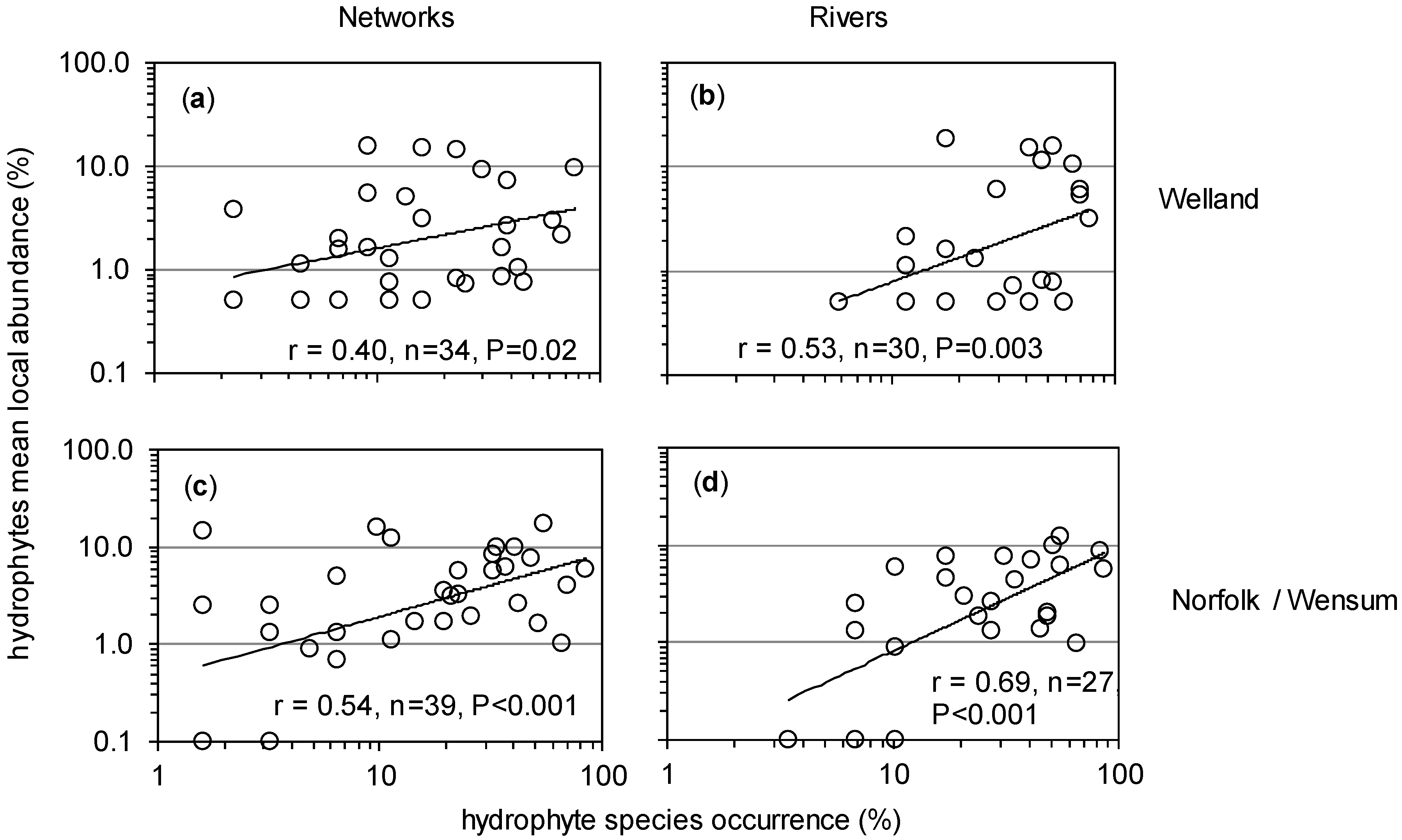
3.4. Spatial Changes along and across River Networks





4. Discussion
4.1. Sampling Design
4.2. Edge Effect: Necessity or Artefact?
4.3. Long Term Changes: Climate, Management Practices and Biotic Interactions
4.4. Short Term Temporal Changes
4.5. Long Term Temporal Changes
4.6. Spatial Connectivity
4.7. Implications
5. Conclusions
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Humboldt, A.; Bonpland, A. Essai sur la Géographie des Plantes; Levrault, Schoell et compagnie: Paris, France, 1805. [Google Scholar]
- Lyell, C. Principles of Geology; John Murray: London, UK, 1830. [Google Scholar]
- Darwin, C. The Origin of Species by Means of Natural Selection, 6th ed.; John Murray: London, UK, 1872. [Google Scholar]
- Warming, E. Lehrbuch der ökologischen Pflanzengeographie. Eine Einführung in die Kenntnis der Pflanzenvereine; Gebrüder Bornträger: Berlin, Germany, 1896. [Google Scholar]
- Hanski, I. Metapopulation dynamics. Nature 1998, 396, 41–49. [Google Scholar] [CrossRef]
- Moilanen, A.; Hanski, I. On the use of connectivity measures in spatial ecology. Oikos 2001, 95, 147–151. [Google Scholar]
- Muneepeerakul, R.; Bertuzzo, E.; Lynch, H.J.; Fagan, W.F.; Rinaldo, A.; Rodriguez-Iturbe, I. Neutral metacommunity models predict fish diversity patterns in Mississippi-Missouri basin. Nature 2008, 453, 220–222. [Google Scholar] [CrossRef]
- Muneepeerakul, R.; Weitz, J.S.; Levin, S.A.; Rinaldo, A.; Rodriguez-Iturbe, I. A neutral metapopulation model of biodiversity in river networks. J. Theor. Biol. 2007, 245, 351–363. [Google Scholar] [CrossRef]
- Labonne, J.; Ravigne, V.; Parisi, B.; Gaucherel, C. Linking dendritic network structures to population demogenetics: The downside of connectivity. Oikos 2008, 117, 1479–1490. [Google Scholar] [CrossRef]
- Venail, P.A.; MacLean, R.C.; Bouvier, T.; Brockhurst, M.A.; Hochberg, M.E.; Mouquet, N. Diversity and productivity peak at intermediate dispersal rate in evolving metacommunities. Nature 2008, 452, 210–214. [Google Scholar] [CrossRef]
- Albert, J.S.; Lovejoy, N.R.; Crampton, W.G.R. Miocene tectonism and the separation of cis- and trans-Andean river basins: Evidence from Neotropical fishes. J. South Am. Earth Sci. 2006, 21, 14–27. [Google Scholar] [CrossRef]
- Burridge, C.P.; Craw, D.; Waters, J.M. River capture, range expansion, and cladogenesis: The genetic signature of freshwater vicariance. Evolution 2006, 60, 1038–1049. [Google Scholar]
- Burridge, C.P.; Craw, D.; Waters, J.M. An empirical test of freshwater vicariance via river capture. Mol. Ecol. 2007, 16, 1883–1895. [Google Scholar] [CrossRef]
- Köhler, F.; Panha, S.; Glaubrecht, M. Speciation and radiation in a river: Assessing the morphological and genetic differentiation in a species flock of viviparous gastropods (Cerithioidea: Pachychilidae). In Evolution in Action; Glaubrecht, M., Ed.; Springer-Verlag Berlin: Heidelberg, Germany, 2010; pp. 513–550. [Google Scholar]
- Vitule, J.R.S.; Skora, F.; Abilhoa, V. Homogenization of freshwater fish faunas after the elimination of a natural barrier by a dam in Neotropics. Divers. Distrib. 2012, 18, 111–120. [Google Scholar] [CrossRef]
- April, J.; Hanner, R.H.; Dion-Cote, A.M.; Bernatchez, L. Glacial cycles as an allopatric speciation pump in north-eastern American freshwater fishes. Mol. Ecol. 2013, 22, 409–422. [Google Scholar] [CrossRef]
- Dias, M.S.; Cornu, J.F.; Oberdorff, T.; Lasso, C.A.; Tedesco, P.A. Natural fragmentation in river networks as a driver of speciation for freshwater fishes. Ecography 2013, 36, 683–689. [Google Scholar] [CrossRef]
- Hrbek, T.; Ferreira da Silva, V.M.; Dutra, N.; Gravena, W.; Martin, A.R.; Farias, I.P. A new species of river dolphin from Brazil or: How little do we know our biodiversity. PLoS ONE 2014, 9, e83623. [Google Scholar]
- Jansson, R.; Nilsson, C.; Renofalt, B. Fragmentation of riparian floras in rivers with multiple dams. Ecology 2000, 81, 899–903. [Google Scholar] [CrossRef]
- Meldgaard, T.; Nielsen, E.E.; Loeschcke, V. Fragmentation by weirs in a riverine system: A study of genetic variation in time and space among populations of European grayling (Thymallus thymallus) in a Danish river system. Conserv. Genet. 2003, 4, 735–747. [Google Scholar] [CrossRef]
- Jansson, R.; Zinko, U.; Merritt, D.M.; Nilsson, C. Hydrochory increases riparian plant species richness: A comparison between a free-flowing and a regulated river. J. Ecol. 2005, 93, 1094–1103. [Google Scholar] [CrossRef]
- Raeymaekers, J.A.M.; Raeymaekers, D.; Koizumi, I.; Geldof, S.; Volckaert, F.A.M. Guidelines for restoring connectivity around water mills: A population genetic approach to the management of riverine fish. J. Appl. Ecol. 2009, 46, 562–571. [Google Scholar] [CrossRef]
- Merritt, D.M.; Nilsson, C.; Jansson, R. Consequences of propagule dispersal and river fragmentation for riparian plant community diversity and turnover. Ecol. Monogr. 2010, 80, 609–626. [Google Scholar] [CrossRef]
- Butcher, R.W. Studies on the ecology of rivers. I. On the distribution of macrophytic vegetation in the rivers in Britain. J. Ecol. 1933, 21, 58–91. [Google Scholar] [CrossRef]
- Boedeltje, G.; Bakker, J.P.; Bekker, R.M.; van Groenendael, J.M.; Soesbergen, M. Plant dispersal in a lowland stream in relation to occurrence and three specific life-history traits of the species in the species pool. J. Ecol. 2003, 91, 855–866. [Google Scholar] [CrossRef]
- Combroux, I.C.S.; Bornette, G. Propagule banks and regenerative strategies of aquatic plants. J. Veg. Sci. 2004, 15, 13–20. [Google Scholar] [CrossRef]
- Pollux, B.J.A.; Santamaria, L.; Ouborg, N.J. Differences in endozoochorous dispersal between aquatic plant species, with reference to plant population persistence in rivers. Freshwater Biol. 2005, 50, 232–242. [Google Scholar]
- Gurnell, A.; Goodson, J.; Thompson, K.; Clifford, N.; Armitage, P. The river-bed: A dynamic store for plant propagules? Earth Surf. Proc. Landf. 2007, 32, 1257–1272. [Google Scholar] [CrossRef]
- Muneepeerakul, R.; Bertuzzo, E.; Rinaldo, A.; Rodriguez-Iturbe, I. Patterns of vegetation biodiversity: The roles of dispersal directionality and river network structure. J. Theor. Biol. 2008, 252, 221–229. [Google Scholar] [CrossRef]
- Kautsky, L. Life strategies of aquatic soft bottom macrophytes. Oikos 1988, 53, 126–135. [Google Scholar] [CrossRef]
- Bornette, G.; Henry, C.; Barrat, M.-H.; Amoros, C. Theoretical habitat templets, species traits, and species richness: aquatic macrophytes in the upper Rhone River and its floodplain. Freshwater Biol. 1994, 31, 487–505. [Google Scholar] [CrossRef]
- Mouw, J.E.B.; Alaback, P.B. Putting floodplain hyperdiversity in a regional context: An assessment of terrestrial-floodplain connectivity in a montane environment. J. Biogeogr. 2003, 30, 87–103. [Google Scholar] [CrossRef]
- Merritt, D.M.; Scott, M.L.; Poff, N.L.; Auble, G.T.; Lytle, D.A. Theory, methods and tools for determining environmental flows for riparian vegetation: Riparian vegetation-flow response guilds. Freshwater Biol. 2010, 55, 206–225. [Google Scholar] [CrossRef]
- Greet, J.; Webb, J.A.; Cousens, R.D. The importance of seasonal flow timing for riparian vegetation dynamics: A systematic review using causal criteria analysis. Freshwater Biol. 2011, 56, 1231–1247. [Google Scholar] [CrossRef]
- Haslam, S.M. River Plants; Cambridge University Press: Cambridge, UK, 1978. [Google Scholar]
- Holmes, N.T.H. Typing British Rivers according to Their Flora; Nature Conservancy Council: Shrewsbury, UK, 1983. [Google Scholar]
- Katz, G.L.; Denslow, M.W.; Stromberg, J.C. The Goldilocks effect: Intermittent streams sustain more plant species than those with perennial or ephemeral flow. Freshwater Biol. 2012, 57, 467–480. [Google Scholar] [CrossRef]
- Holmes, N.T.H.; Whitton, B.A. Submerged bryophytes and angiosperms of the River Tweed and its tributaries. Trans. Bot. Soc. Edinb. 1975, 42, 383–395. [Google Scholar] [CrossRef]
- Haury, J. Patterns of macrophyte distribution within a breton brook compared with other study scales. Landsc. Urban Plan. 1995, 31, 349–361. [Google Scholar] [CrossRef]
- Honnay, O.; Verhaeghe, W.; Hermy, M. Plant community assembly along dendritic networks of small forest streams. Ecology 2001, 82, 1691–1702. [Google Scholar] [CrossRef]
- Riis, T.; Sand-Jensen, K. Abundance-range size relationships in stream vegetation in Denmark. Plant Ecol. 2002, 161, 175–183. [Google Scholar] [CrossRef]
- Riis, T.; Sand-Jensen, K. Historical changes in species composition and richness accompanying perturbation and eutrophication of Danish lowland streams over 100 years. Freshwater Biol. 2001, 46, 269–280. [Google Scholar] [CrossRef]
- Demars, B.O.L.; Harper, D.M. Distribution of aquatic vascular plants in lowland rivers: Separating the effects of local environmental conditions, longitudinal connectivity and river basin isolation. Freshwater Biol. 2005, 50, 418–437. [Google Scholar] [CrossRef]
- Gornall, R.J.; Hollingsworth, P.M.; Preston, C.D. Evidence for spatial structure and directional gene flow in a population of an aquatic plant, Potamogeton coloratus. Heredity 1998, 80, 414–421. [Google Scholar] [CrossRef]
- Andersson, E.; Nilsson, C.; Johansson, M.E. Effects of river fragmentation on plant dispersal and riparian flora. Regul. Rivers Res. Manag. 2000, 16, 83–89. [Google Scholar] [CrossRef]
- Boedeltje, G.; Bakker, J.P.; Ten Brinke, A.; Van Groenendael, J.M.; Soesbergen, M. Dispersal phenology of hydrochorous plants in relation to discharge, seed release time and buoyancy of seeds: The flood pulse concept supported. J. Ecol. 2004, 92, 786–796. [Google Scholar] [CrossRef]
- Pollux, B.J.A.; Jong, M.D.E.; Steegh, A.; Verbruggen, E.; Van Groenendael, J.M.; Ouborg, N.J. Reproductive strategy, clonal structure and genetic diversity in populations of the aquatic macrophyte Sparganium emersum in river systems. Mol. Ecol. 2007, 16, 313–325. [Google Scholar]
- Pollux, B.J.A.; De Jong, M.; Steegh, A.; Ouborg, N.J.; Van Groenendael, J.M.; Klaassen, M. The effect of seed morphology on the potential dispersal of aquatic macrophytes by the common carp (Cyprinus carpio). Freshwater Biol. 2006, 51, 2063–2071. [Google Scholar] [CrossRef]
- Fer, T.; Hroudova, Z. Detecting dispersal of Nuphar lutea in river corridors using microsatellite markers. Freshwater Biol. 2008, 53, 1409–1422. [Google Scholar] [CrossRef]
- Henry, C.P.; Amoros, C.; Bornette, G. Species traits and recolonization processes after flood disturbances in riverine macrophytes. Vegetatio 1996, 122, 13–27. [Google Scholar] [CrossRef]
- Barrat-Segretain, M.H.; Bornette, G. Regeneration and colonization abilities of aquatic plant fragments: Effect of disturbance seasonality. Hydrobiologia 2000, 421, 31–39. [Google Scholar] [CrossRef]
- Puijalon, S.; Piola, F.; Bornette, G. Abiotic stresses increase plant regeneration ability. Evol. Ecol. 2008, 22, 493–506. [Google Scholar] [CrossRef]
- Gurnell, A.; Thompson, K.; Goodson, J.; Moggridge, H. Propagule deposition along river margins: Linking hydrology and ecology. J. Ecol. 2008, 96, 553–565. [Google Scholar] [CrossRef]
- Wiegleb, G. Recherches Méthodologiques sur les Groupemnents Végétaux des Eaux Courantes. In Colloques Phytosociologiques X. Les Végétations Aquatiques et Amphibies 1981; Koeltz Scientific Books: Lille, France, 1983; pp. 69–83. [Google Scholar]
- Tremp, H. Spatial and environmental effects on hydrophytic macrophyte occurrence in the Upper Rhine floodplain (Germany). Hydrobiologia 2007, 586, 167–177. [Google Scholar] [CrossRef]
- Pedersen, T.C.M.; Baattrup-Pedersen, A.; Madsen, T.V. Effects of stream restoration and management on plant communities in lowland streams. Freshwater Biol. 2006, 51, 161–179. [Google Scholar] [CrossRef]
- Demars, B.O.L. Aquatic Vascular Plants in Nitrate-Rich Calcareous Lowland Streams: Do They Respond to Phosphorus Enrichment and Control? Ph.D. Thesis, University of Leicester, Leicester, UK, 2002. [Google Scholar]
- Wiegleb, G. A study of habitat conditions of the macrophytic vegetation in selected river systems in western lower saxony (Federal Republic of Germany). Aquat. Bot. 1984, 18, 313–352. [Google Scholar] [CrossRef]
- Dunn, R.R.; Colwell, R.K.; Nilsson, C. The river domain: Why are there more species halfway up the river? Ecography 2006, 29, 251–259. [Google Scholar] [CrossRef]
- Wiegleb, G.; Herr, W.; Todeskino, D. Ten years of vegetation dynamics in two rivulets in Lower Saxony (FRG). Vegetatio 1989, 82, 163–178. [Google Scholar] [CrossRef]
- Demars, B.O.L.; Harper, D.M. The aquatic macrophytes of an English lowland river system: Assessing response to nutrient enrichment. Hydrobiologia 1998, 384, 75–88. [Google Scholar] [CrossRef]
- Clarke, A.; Nally, R.M.; Bond, N.; Lake, P.S. Macroinvertebrate diversity in headwater streams: A review. Freshwater Biol. 2008, 53, 1707–1721. [Google Scholar] [CrossRef]
- Legendre, P. Spatial autocorrelation: Trouble or new paradigm? Ecology 1993, 74, 1659–1673. [Google Scholar] [CrossRef]
- Tremp, H. Geostatistische Analyse der Strahlwirkung in Fließgewässern am Beispiel der Wasserpflanzen. In der Deutsche Gesellschaft für Limnologie (DGL), Erweiterte Zusammenfassungen der Jahrestagung 2008; Hardegsen: Konstanz, Germany, 2009; pp. 518–523. [Google Scholar]
- Demars, B.O.L.; Trémolières, M. Aquatic macrophytes as bioindicators of carbon dioxide in groundwater fed rivers. Sci. Total. Environ. 2009, 407, 4752–4763. [Google Scholar] [CrossRef]
- Borcard, D.; Legendre, P.; Drapeau, P. Partialling out the spatial component of ecological variation. Ecology 1992, 73, 1045–1055. [Google Scholar] [CrossRef]
- Lennon, J.J.; Koleff, P.; Greenwood, J.J.D.; Gaston, K.J. Contribution of rarity and commonness to patterns of species richness. Ecol. Lett. 2004, 7, 81–87. [Google Scholar]
- Willby, N.J.; Abernethy, V.J.; Demars, B.O.L. An attribute-based classification of European hydrophytes and its relationship to habitat utilisation. Freshwater Biol. 2000, 43, 43–74. [Google Scholar] [CrossRef]
- Wiegleb, G.; Bröring, U.; Filetti, M.; Brux, H.; Herr, W. Long-term dynamics of macrophyte dominance and growth form types in two Northwest German lowland streams. Freshwater Biol. 2014, 59. [Google Scholar] [CrossRef]
- Dawson, H.F.; Castellano, E.; Ladle, M. Concept of species succession in relation to river vegetation and management. Verh. Internat. Verein. Limnol. 1978, 20, 1429–1434. [Google Scholar]
- Walter, R.C.; Merritts, D.J. Natural streams and the legacy of water-powered mills. Science 2008, 319, 299–304. [Google Scholar] [CrossRef]
- Demars, B.O.L. Using Aquatic Macrophytes for Assessing Water Trophic Level in a Lowland River System. Master’s Thesis, Leicester University, Leicester, UK, 1996. [Google Scholar]
- Pope, A. GB rivers. Available online: http://hdl.handle.net/10672/85 (accessed on 28 November 2013).
- Ordnance Survey OpenData™. Available online: https://www.ordnancesurvey.co.uk/ opendatadownload/products.html (accessed on 16 November 2012).
- Holmes, N.T.H. The Use of Riverine Macrophytes for the Assessment of Trophic Status: Review of 1994/95 Data & Refinements for Future Use; Report to the National Rivers Authority Anglian Region; National Rivers Authority: Peterborough, Canada, 1996. [Google Scholar]
- Holmes, N.T.H.; Newman, J.R.; Chadd, S.; Rouen, K.J.; Saint, L.; Dawson, F.H. Mean Trophic Rank: A User’s Manual; Report number E38; Environment Agency: Bristol, UK, 1999; pp. 1–133. [Google Scholar]
- Blanquet, J.B. Plant Sociology: The Study of Plant Communities, 1st ed.; Mc Graw Hill Book Co: New York, NY, USA, 1932. [Google Scholar]
- Ter Braak, C.J.F.; Šmilauer, P. CANOCO Reference Manual and Canodraw for Windows User’s Guide: Software for Canonical Community Ordination (version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- GenStat. VSN International. Available online: http://www.vsni.co.uk/software/genstat (accessed on 1 April 2014).
- Legendre, P.; Legendre, L. Numerical Ecology; Elsevier Science: Amsterdam, the Netherland, 1998. [Google Scholar]
- Tokeshi, M. Niche apportionment or random assortment: Species abundance patterns revisited. J. Anim. Ecol. 1990, 59, 1129–1146. [Google Scholar] [CrossRef]
- Baattrup-Pedersen, A.; Larsen, S.E.; Riis, T. Long-term effects of stream management on plant communities in two Danish lowland streams. Hydrobiologia 2002, 481, 33–45. [Google Scholar] [CrossRef]
- Baattrup-Pedersen, A.; Larsen, S.E.; Riis, T. Composition and richness of macrophyte communities in small Danish streams—Influence of environmental factors and weed cutting. Hydrobiologia 2003, 495, 171–179. [Google Scholar] [CrossRef]
- Arthington, A.H.; Naiman, R.J.; McClain, M.E.; Nilsson, C. Preserving the biodiversity and ecological services of rivers: New challenges and research opportunities. Freshwater Biol. 2010, 55, 1–16. [Google Scholar]
- Monthly Teleconnection Index: North Atlantic Oscillation (NAO). Available online: ftp://ftp.cpc.ncep.noaa.gov/wd52dg/data/indices/nao_index.tim (accessed on 1 April 2014).
- Legendre, P.; Galzin, R.; Harmelin-Vivien, M.L. Relating behavior to habitat: Solutions to the fourth-corner problem. Ecology 1997, 78, 547–562. [Google Scholar]
- Gould, S.J.; Lewontin, R.C. The spandrels of San Marco and the Panglossian paradigm: A critique of the adaptationist programme. Proc. R. Soc Lond. Ser. B Biol. Sci. 1979, 205, 581–598. [Google Scholar] [CrossRef]
- Holmes, N.T.H.; Whitton, B.A. Macrophytes of the River Tweed. Trans. Bot. Soc. Edinb. 1975, 42, 369–381. [Google Scholar] [CrossRef]
- Nilsson, C.; Grelsson, G.; Johansson, M.; Sperens, U. Patterns of plant species richness along riverbanks. Ecology 1989, 70, 77–84. [Google Scholar] [CrossRef]
- Décamps, H.; Tabacchi, E. Species richness in vegetation along river margins. In Aquatic Ecology: Scale, Pattern and Process; Giller, P.S., Hildrew, A., Raffaelli, D.G., Eds.; Blackwell Science: Oxford, UK, 1994. [Google Scholar]
- Peterson, E.E.; Hoef, J.M.V. A mixed-model moving-average approach to geostatistical modeling in stream networks. Ecology 2010, 91, 644–651. [Google Scholar] [CrossRef]
- Wuensch, K.L. Comparing correlation coefficients, slopes, and intercepts. Statistics Lessons. Available online: http://core.ecu.edu/psyc/wuenschk/statslessons.htm (accessed on 13 November 2013).
- Beyer, H.L. Geospatial Modelling Environment, Available online:. Available online: http://www.spatialecology.com/gme/ (accessed on 10 October 2013).
- Demars, B.O.L.; Edwards, A.C. Distribution of aquatic macrophytes in contrasting river systems: A critique of compositional-based assessment of water quality. Sci. Total. Environ. 2009, 407, 975–990. [Google Scholar] [CrossRef]
- Scott, W.A.; Adamson, J.K.; Rollinson, J.; Parr, T.W. Monitoring of aquatic macrophytes for detection of long-term change in river systems. Environ. Monit. Assess 2002, 73, 131–153. [Google Scholar] [CrossRef]
- Daniel, H. Evaluation de la Qualité des cours d’eau par la Végétation Macrophytique. Travail in situ et Expérimental Dans le Massif Armoricain sur les Pollutions par les Macronutriments. Ph.D. Thesis, Ecole Nationale Supérieure Agronomique de Rennes, Rennes, France, 1998. [Google Scholar]
- Bornette, G.; Amoros, C. Aquatic vegetation and hydrology of a braided river floodplain. J. Veg. Sci. 1991, 2, 497–512. [Google Scholar] [CrossRef]
- Martins, S.V.; Milne, J.; Thomaz, S.M.; McWaters, S.; Mormul, R.P.; Kennedy, M.; Murphy, K. Human and natural drivers of changing macrophyte community dynamics over 12 years in a Neotropical riverine floodplain system. Aquat. Conserv. Mar. Freshw. Ecosyst. 2013, 23, 678–697. [Google Scholar]
- Steffen, K.; Becker, T.; Herr, W.; Leuschner, C. Diversity loss in the macrophyte vegetation of northwest German streams and rivers between the 1950s and 2010s. Hydrobiologia 2013, 713, 1–17. [Google Scholar] [CrossRef]
- Breugnot, E.; Dutartre, A.; Laplace-Treyture, C.; Haury, J. Local distribution of macrophytes and consequences for sampling methods in large rivers. Hydrobiologia 2008, 610, 13–23. [Google Scholar] [CrossRef]
- Wilby, R.L.; Cranston, L.E.; Darby, E.J. Factors governing macrophyte status in Hampshire chalk streams: Implications for catchment management. J. Chart. Inst. Water and Environ. Manag. 1998, 12, 179–187. [Google Scholar] [CrossRef]
- Sosiak, A. Long-term response of periphyton and macrophytes to reduced municipal nutrient loading to the Bow River (Alberta, Canada). Can. J. Fish Aquat. Sci. 2002, 59, 987–1001. [Google Scholar] [CrossRef]
- Soulsby, P.G. The effect of a heavy cut on the subsequent growth of aquatic plants in a Hampshire chalk stream. J. Inst. Fish. Manag. 1974, 5, 49–53. [Google Scholar]
- Ham, S.F.; Wright, J.F.; Berrie, A.D. The effect of cutting on the growth and recession of the freshwater macrophyte Ranunculus penicillatus (Dumort.) Bab. var. calcareus (R.W. Butcher) C.D.K. Cook. J. Environ. Manag. 1982, 15, 263–271. [Google Scholar]
- Fox, A.M.; Murphy, K.J. The efficacy and ecological impacts of herbicide and cutting regimes on the submerged plant communities of four British rivers. I. Comparison of management efficacies. J. Appl. Ecol. 1990, 27, 520–540. [Google Scholar] [CrossRef]
- Parker, J.D.; Burkepile, D.E.; Collins, D.O.; Kubanek, J.; Hay, M.E. Stream mosses as chemically-defended refugia for freshwater macroinvertebrates. Oikos 2007, 116, 302–312. [Google Scholar] [CrossRef]
- Rybicki, N.B.; Landwehr, J.M. Long-term changes in abundance and diversity of macrophyte and waterfowl populations in an estuary with exotic macrophytes and improving water quality. Limnol. Oceanogr. 2007, 52, 1195–1207. [Google Scholar] [CrossRef]
- Elger, A.; Willby, N.J.; Cabello-Martinez, M. Invertebrate grazing during the regenerative phase affects the ultimate structure of macrophyte communities. Freshwater Biol. 2009, 54, 1246–1255. [Google Scholar] [CrossRef]
- Haury, J.; Aidara, L.G. Macrophyte cover and standing crop in the River scorff and its tributaries (Brittany, northwestern France): Scale, patterns and process. Hydrobiologia 1999, 415, 109–115. [Google Scholar] [CrossRef]
- Sand-Jensen, K.; Andersen, K.; Andersen, T. Dynamic properties of recruitment, expansion and mortality of macrophyte patches in streams. Int. Rev. Hydrobiol. 1999, 84, 497–508. [Google Scholar]
- Champion, P.D.; Tanner, C.C. Seasonality of macrophytes and interaction with flow in a New Zealand lowland stream. Hydrobiologia 2000, 441, 1–12. [Google Scholar] [CrossRef]
- Barrat-Segretain, M.H.; Amoros, C. Recolonization of cleared riverine macrophyte patches: Importance of the border effect. J. Veg. Sci. 1996, 7, 769–776. [Google Scholar] [CrossRef]
- Barrat-Segretain, M.H.; Amoros, C. Recovery of riverine vegetation after experimental disturbance: A field test of the patch dynamics concept. Hydrobiologia 1996, 321, 53–68. [Google Scholar] [CrossRef]
- Kõrs, A.; Vilbaste, S.; Käeiro, K.; Pall, P.; Piirsoo, K.; Truu, J.; Viik, M. Temporal changes in the composition of macrophyte communities and environmental factors governing the distribution of aquatic plants in an unregulated lowland river (Emajogi, Estonia). Boreal Environ. Res. 2012, 17, 460–472. [Google Scholar]
- Scheffer, M.; Bascompte, J.; Brock, W.A.; Brovkin, V.; Carpenter, S.R.; Dakos, V.; Held, H.; van Nes, E.H.; Rietkerk, M.; Sugihara, G. Early-warning signals for critical transitions. Nature 2009, 461, 53–59. [Google Scholar] [CrossRef]
- Urban, M.C.; Skelly, D.K.; Burchsted, D.; Price, W.; Lowry, S. Stream communities across a rural-urban landscape gradient. Divers. Distrib. 2006, 12, 337–350. [Google Scholar] [CrossRef]
- Amoros, C.; Bornette, G. Connectivity and biocomplexity in waterbodies of riverine floodplains. Freshwater Biol. 2002, 47, 761–776. [Google Scholar] [CrossRef]
- Whitton, B.A.; Boulton, P.N.G.; Clegg, E.M.; Gemmell, J.J.; Graham, G.G.; Gustar, R.; Moorhouse, T.P. Long-term changes in macrophytes of British rivers: 1. River Wear. Sci. Total Environ. 1998, 210/211, 411–426. [Google Scholar] [CrossRef]
- Schweinitz, P.; Poschlod, P.; Zeltner, G.-H.; Kohler, A. Langzeitmonitoring (1970–2010) zur Verbreitung der Makrophyten im Fließgewässersystem Moosach (Münchner Ebene). In der Deutschen Gesellschaft für Limnologie (DGL), Erweiterte Zusammenfassungen der Jahrestagung 2011; Hardegsen: Weihenstephan, Germany, 2012; pp. 98–103. [Google Scholar]
- Kernan, M.; Battarbee, R.W.; Curtis, C.J.; Monteith, D.T.; Shilland, E.M. Recovery of Lakes and Streams in the UK from the Effects of Acid Rain UK Acid Waters Monitoring Network 20 Year Interpretative Report; Environmental Change Research Centre: London, UK, 2010. [Google Scholar]
- UK Environmental Change Network. Available online: http://www.ecn.ac.uk/ (accessed on 1 April 2014).
- Moilanen, A.; Leathwick, J.; Elith, J. A method for spatial freshwater conservation prioritization. Freshwater Biol. 2008, 53, 577–592. [Google Scholar] [CrossRef]
- Hermoso, V.; Linke, S.; Prenda, J.; Possingham, H.P. Addressing longitudinal connectivity in the systematic conservation planning of fresh waters. Freshwater Biol. 2011, 56, 57–70. [Google Scholar] [CrossRef]
- Renöfält, B.M.; Jansson, R.; Nilsson, C. Effects of hydropower generation and opportunities for environmental flow management in Swedish riverine ecosystems. Freshwater Biol. 2010, 55, 49–67. [Google Scholar] [CrossRef]
- Brederveld, R.J.; Jahnig, S.C.; Lorenz, A.W.; Brunzel, S.; Soons, M.B. Dispersal as a limiting factor in the colonization of restored mountain streams by plants and macroinvertebrates. J. Appl. Ecol. 2011, 48, 1241–1250. [Google Scholar] [CrossRef]
- Andersson, E.; Nilsson, C.; Johansson, M.E. Plant dispersal in boreal rivers and its relation to the diversity of riparian flora. J. Biogeogr. 2000, 27, 1095–1106. [Google Scholar] [CrossRef]
- Holmes, N.T.H. The Vegetation of the River Tweed. Ph.D. Thesis, Durham University, Durham, UK, 1975. [Google Scholar]
- Fernandes, C.C.; Podos, J.; Lundberg, J.G. Amazonian ecology: Tributaries enhance the diversity of electric fishes. Science 2004, 305, 1960–1962. [Google Scholar] [CrossRef]
- Besemer, K.; Peter, H.; Logue, J.B.; Langenheder, S.; Lindstrom, E.S.; Tranvik, L.J.; Battin, T.J. Unraveling assembly of stream biofilm communities. Isme J. 2012, 6, 1459–1468. [Google Scholar] [CrossRef]
- Gascon, C.; Smith, M.L. Where rivers meet. Science 2004, 305, 1922–1923. [Google Scholar] [CrossRef]
- Townsend, C.R. The patch dynamics concept of stream community ecology. J. N. Am. Benthol. Soc. 1989, 8, 36–50. [Google Scholar] [CrossRef]
- Grime, J.P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar]
- Townsend, C.R.; Hildrew, A.G. Species traits in relation to a habitat templet for river systems. Freshwater Biol. 1994, 31, 265–275. [Google Scholar] [CrossRef]
- Johansson, M.E.; Nilsson, C.; Nilsson, E. Do rivers function as corridors for plant dispersal? J. Veg. Sci. 1996, 7, 593–598. [Google Scholar] [CrossRef]
- Moggridge, H.L.; Gurnell, A.M.; Mountford, J.O. Propagule input, transport and deposition in riparian environments: the importance of connectivity for diversity. J. Veg. Sci. 2009, 20, 465–474. [Google Scholar] [CrossRef]
- Nilsson, C.; Ekblad, A.; Dynesius, M.; Backe, S.; Gardfjell, M.; Carlberg, B.; Hellqvist, S.; Jansson, R. A comparison of species richness and traits of riparian plants between a main river channel and tributaries. J. Ecol. 1994, 82, 281–295. [Google Scholar] [CrossRef]
- Buckley, L.B.; Urban, M.C.; Angilletta, M.J.; Crozier, L.G.; Rissler, L.J.; Sears, M.W. Can mechanism inform species’ distribution models? Ecol. Lett. 2010, 13, 1041–1054. [Google Scholar]
- Stace, C.A. New Flora of the British Isles, 2nd Ed. ed; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
Appendix 1
| Hydrophytes | Helophytes | Helophytes |
|---|---|---|
| Batrachospermum gelatinosum | Achillea ptarmica | Hydrocotyle vulgaris |
| Callitriche hamulata | Agrostis canina | Juncus articulatus |
| Callitriche obtusangula | Agrostis capillaris | Juncus bufonius |
| Callitriche platycarpa | Agrostis stolonifera | Juncus effusus |
| Elodea canadensis | Alisma plantago-aquatica | Lolium perenne |
| Elodea nuttallii | Alopecurus geniculatus | Lotus pedunculatus |
| Fontinalis antipyretica | Alopecurus pratensis | Lycopus europaeus |
| Hydrocharis morsus-ranae | Bidens cernua | Lysimachia nummularia |
| Lemna gibba | Bidens tripartita | Lysimachia vulgaris |
| Lemna minor | Botrydium granulatum | Lythrum salicaria |
| Leptodictyum riparium | Calamagrostis canescens | Mentha arvensis |
| Myriophyllum alterniflorum | Caltha palustris | Myosotis scorpioides |
| Nitella flexilis | Cardamine amara | Rorippa nasturtium-aquaticum |
| Nuphar lutea | Carex acuta | Persicaria hydropiper |
| Oenanthe aquatica | Carex paniculata | Phalaris arundinacea |
| Persicaria amphibia | Deschampsia cespitosa | Poa palustris |
| Potamogeton alpinus | Eleocharis acicularis | Poa trivialis |
| Potamogeton berchtoldii | Eleocharis palustris | Ranunculus repens |
| Potamogeton crispus | Epilobium hirsutum | Rorippa amphibia |
| Potamogeton natans | Epilobium obscurum | Rorippa palustris |
| Potamogeton perfoliatus | Epilobium palustre | Rorippa sylvestris |
| Potamogeton trichoides | Epilobium roseum | Rumex acetosa |
| Ranunculus peltatus | Equisetum palustre | Rumex hydrolapathum |
| Sagittaria sagittifolia | Galium palustre | Rumex obtusifolius |
| Sparganium emersum | Galium uliginosum | Scirpus sylvaticus |
| Sparganium erectum | Glechoma hederacea | Solanum dulcamara |
| Spirodela polyrhiza | Glyceria fluitans | Stachys palustris |
| Glyceria maxima | Stellaria uliginosa | |
| Holcus mollis | Veronica beccabunga |
| Hydrophytes | Helophytes |
|---|---|
| Alisma plantago-aquatica | Angelica sylvestris |
| Amblystegium riparium | Carex acuta |
| Apium nodiflorum | Carex acutiformis |
| Butomus umbellatus | Cirsium palustre |
| Callitriche spp. | Epilobium hirsutum |
| Elodea canadensis | Equisetum fluitans |
| Elodea nuttallii | Equisetum palustre |
| Fontinalis antipyretica | Glyceria maxima |
| Glyceria fluitans | Iris pseudacorus |
| Lemna gibba | Juncus effusus |
| Lemna minuta/minor | Juncus inflexus |
| Mentha aquatica | Persicaria maculosa |
| Myosotis scorpioides | Petasites hybridus |
| Myriophyllum spicatum | Phalaris arundinacea |
| Nuphar lutea | Ranunculus repens |
| Nymphoides peltata | Rorippa amphibia |
| Oenanthe fluviatilis | Rumex sp. |
| Persicaria amphibia | Scirpus sylvaticus |
| Potamogeton crispus | Scrophularia auriculata |
| Potamogeton natans | Solanum dulcamara |
| Potamogeton obtusifolius | Typha latifolia |
| Potamogeton pectinatus | Urtica dioica |
| Potamogeton perfoliatus | Veronica catenata |
| Potamogeton x salicifolius | |
| Ranunculus subgenus Batrachium | |
| Rhynchostegium riparioides | |
| Rorippa-nasturtium aquaticum | |
| Sagittaria sagittifolia | |
| Schoenoplectus lacustris | |
| Sparganium emersum | |
| Sparganium erectum | |
| Veronica anagallis-aquatica | |
| Veronica beccabunga | |
| Zannichellia palustris |
| Hydrophytes | Helophytes |
|---|---|
| Amblystegium riparium | Agrostis stolonifera |
| Berula erecta | Apium nodiflorum |
| Butomus umbellatus | Caltha palustris |
| Callitriche obtusangula/platycarpa | Carex sp. |
| Ceratophyllum demersum | Epilobium hirsutum |
| Chara globularis | Equisetum fluviatile |
| Elodea canadensis | Equisetum palustris |
| Elodea nuttallii | Eupatorium cannabinum |
| Fontinalis antipyretica | Glyceria maxima |
| Glyceria declinata/notata/fluitans | Iris pseudacorus |
| Groenlandia densa | Juncus inflexus |
| Hippuris vulgaris | Juncus subnodulosus |
| Hottonia palustris | Lycopus europaeus |
| Lemna minor/minuta | Mentha aquatica |
| Lemna trisulca | Mimulus guttatus |
| Myriophyllum spicatum | Myosites scorpioides |
| Myriophyllum verticillatum | Phalaris arundinacea |
| Nitella flexilis | Phragmites australis |
| Nuphar lutea | Ranunculus repens |
| Nymphaea alba | Rorippa nasturtium-aquaticum |
| Oenanthe fluviatilis | Rumex sp. |
| Pellia sp. | Scrophularia auriculata |
| Potamogeton alpinus | Solanum dulcamara |
| Potamogeton crispus | Typha latifolia |
| Potamogeton friesii | Veronica beccabunga/catenata |
| Potamogeton lucens | |
| Potamogeton pectinatus | |
| Potamogeton perfoliatus | |
| Potamogeton pusillus | |
| Ranunculus penicillatus/trichophyllous | |
| Ranunculus circinatus | |
| Ranunculus fluitans | |
| Riccia natans | |
| Sagittaria sagittifolia | |
| Schoenoplectus lacustris | |
| Sparganium emersum | |
| Sparganium erectum | |
| Veronica anagallis-aquatica | |
| Zannichellia palustris |
Appendix 2




© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Demars, B.O.L.; Wiegleb, G.; Harper, D.M.; Bröring, U.; Brux, H.; Herr, W. Aquatic Plant Dynamics in Lowland River Networks: Connectivity, Management and Climate Change. Water 2014, 6, 868-911. https://doi.org/10.3390/w6040868
Demars BOL, Wiegleb G, Harper DM, Bröring U, Brux H, Herr W. Aquatic Plant Dynamics in Lowland River Networks: Connectivity, Management and Climate Change. Water. 2014; 6(4):868-911. https://doi.org/10.3390/w6040868
Chicago/Turabian StyleDemars, Benoît O.L., Gerhard Wiegleb, David M. Harper, Udo Bröring, Holger Brux, and Wolfgang Herr. 2014. "Aquatic Plant Dynamics in Lowland River Networks: Connectivity, Management and Climate Change" Water 6, no. 4: 868-911. https://doi.org/10.3390/w6040868