
Colonization Dynamics of Potential Stowaways Inhabiting Marinas: Lessons from Caprellid Crustaceans


1
Departamento de Biología, Facultad de Ciencias del Mar y Ambientales, Campus de Excelencia Internacional del Mar (CEIMAR), Universidad de Cádiz, Avenida República Saharaui s/n, 11510 Puerto Real, Spain
2
Laboratorio de Biología Marina, Departamento de Zoología, Facultad de Biología, Universidad de Sevilla, Avda Reina Mercedes 6, 41012 Seville, Spain
3
Instituto Universitario de Investigación Marina (INMAR), Campus de Excelencia Internacional del Mar (CEIMAR), Universidad de Cádiz, 11510 Puerto Real, Spain
*
Author to whom correspondence should be addressed.
Water 2022, 14(17), 2659; https://doi.org/10.3390/w14172659
Submission received: 31 July 2022
/
Revised: 22 August 2022
/
Accepted: 24 August 2022
/
Published: 28 August 2022
(This article belongs to the Special Issue Biological Invasions in the Marine Environment)
Abstract
:Despite the fact that stowaway transport is an increasingly common invasion pathway, its key ecological aspects, such as colonization dynamics of potential invaders on human transport vectors, are poorly understood. Considering that marinas may function as stepping-stone habitats for the spread of marine invaders, they provide a unique opportunity for understanding the first steps in the stowaway pathway. One of the dominant groups inhabiting marinas are caprellid crustaceans. These small invertebrates have demonstrated a high potential for introduction and establishment far beyond their distribution range. Here, we analyzed the colonization dynamics of caprellid species inhabiting marinas of Cádiz Bay (Spain) on plastic collectors and discussed their implications on invasion success. We found that collectors largely mirrored the resident caprellid community assemblages after just one month of deployment, showing that caprellid species inhabiting marina structures have a high capacity to colonize new artificial substrates. Moreover, the two non-indigenous species found, Caprella scaura and Paracaprella pusilla, successfully established dense and well-structured populations in the collectors (including all sex/age groups considered). We suggest a potential interplay between colonization ability and stowaway organisms’ invasion success. Furthermore, collectors used are supported as a useful tool in the early detection of non-indigenous species and their monitoring in relation to resident species.
1. Introduction
Increasing global connectivity through human transport networks is unintentionally moving a massive number of stowaway organisms into novel biogeographical regions [1,2,3]. Indeed, non-indigenous species (NIS) transported as stowaways on ships are prevalent in most marine ecosystems throughout the world [4], posing one of the greatest threats to marine biodiversity and ecosystem functioning [5]. Understanding the ways humans carry out this transport is needed to reduce this threat [6].
Invasion is a multi-step process that essentially include: (1) the species’ uptake by a human transport vector (e.g., ship’s hull fouling or ballast water) at the ‘donor’ location; (2) movement of the human vector to a new location, beyond the species’ distribution range; (3) establishment of a self-sustaining population within the ‘recipient’ location; and (4) spread of species to nearby habitats [7,8,9]. While later stages receive the greatest research effort, the contingent nature of the process makes the first steps on the introduction pathway pivotal in the likelihood of biological invasion [10]. Therefore, understanding the interaction between potential stowaway source populations at the donor location and human transport vectors is fundamental to shed light on this growing invasion pathway.
In marine ecosystems, transport hubs, such as marinas and commercial ports, may function as stepping-stones habitats in the invasion pathway. They enable many fouling organism species that are unintentional transported as stowaways on ships (either in the ships’ hull or in ballast water) to colonize areas beyond their distribution ranges [11]. While commercial ports are usually considered the first entry point of NIS into a new region, marinas are critical habitats for their successful establishment and subsequent spread. This is partly due to the larger surface available for species colonization in marinas in comparison with commercial ports, but also since recreational boats remain moored more time than commercial ships (making it easier for stowaways to colonize recreational boats) [12]. Furthermore, recreational boating is a high-risk NIS dispersal vector, lacking a proper regulation, which could facilitate the NIS spread not only among marinas, but also into adjacent natural habitats [13]. A crucial step at the beginning of the stowaway pathway is the colonization of human transport vectors (or vector uptake). Reaching and colonizing anthropogenic vectors in a transport hub (such as marinas) may be a determinantal filter to stop stowaway transport events. However, little information exists about potential traits that may favor this early stage in the stowaway pathway. For example, Ros et al. [14] found that peracarid crustaceans with a documented history of anthropogenic transport had a higher propensity for local dispersal (i.e., tendency to move about short distances) in comparison with other peracarids that had a restricted distribution, lacking a history of human-mediated introductions. This supports the notion that a high tendency to move about short distances may increase the chances to reach and colonize human transport vectors, the frequency of transport, and, therefore, promote success in the invasion process [15]. However, the initial dispersal of immigrating individuals from a source population is only the first step in the colonization process and not all dispersing individuals are necessarily good colonizers. From a metapopulation perspective, colonization can in turn be described as the result of different processes, starting with the emigration of individuals from a source population (i.e., local species pool) and the subsequent dispersal across a more or less suitable habitat matrix. Then, colonizing individuals face the challenges of surviving and reproducing in a novel empty niche. The final stage generally takes place when the viability of a population is no longer dependent on the arrival of ‘propagules’ (i.e., colonizing individuals) [16,17]. Despite colonization abilities vary among marine organism species [18], ecological studies generally ignore this potential variation at the beginning of the stowaway pathway. Indeed, the capacity of potential stowaways to colonize human transport vectors has rarely been experimentally assessed in marine ecosystems. There are few quantitative and comparative data documenting the patterns of spatial and temporal variation on which colonization occurs in this context. Understanding the colonization dynamics of potential stowaways inhabiting transport hubs on human transport vectors can provide valuable insight into the early stages of the invasion process. This in turn may improve stowaway risk assessments.
One of the dominant groups among the fouling communities inhabiting marina structures are caprellid amphipods (e.g., [19,20]). These small peracarid crustaceans live as epibionts on a wide variety of sessile species encrusting marina structures, including macroalgae, bryozoans or hydrozoans (especially in those basibionts with an arborescent and complex structure). Furthermore, they can also reach high densities in submerged complex artificial substrates (e.g., ropes or nets) [21]. Despite their limited ability for autonomous dispersion (due to its lack of larval stage and its reduced swimming appendages), some caprellid species have demonstrated a high potential for the introduction and establishment far beyond their distribution range, causing significant impacts on receiving communities [22]. It has been proposed that caprellids species inhabiting marina structures can reach and colonize encrusting basibiont species on the submerged surface of human transport vectors (e.g., boats’ hull), which could serve as a refuge during anthropogenic transport [20]. However, little is known about how these first steps in the stowaway pathway occurs and why some caprellid species fail to spread while others do [14].
We aim to investigate the colonization dynamics of caprellid species inhabiting transport hubs (specifically marinas) on new artificial substrates (i.e., plastic collectors) and its potential implications on invasion success (considered here as the potential to spread via anthropogenic vectors). We hypothesized that the deployment of new artificial substrates for one and three months would mirror the resident caprellid community by providing empty niches for the local pool of species. Therefore, we expect that if caprellid species inhabiting marina structures (i.e., local basibionts) are good colonizers, caprellid assemblages will differ only among the marinas investigated in the two periods. Additionally, we assessed the efficacy of collectors for early detection of non-indigenous caprellid species and their monitoring in relation to resident species.
2. Materials and Methods
2.1. Study Area
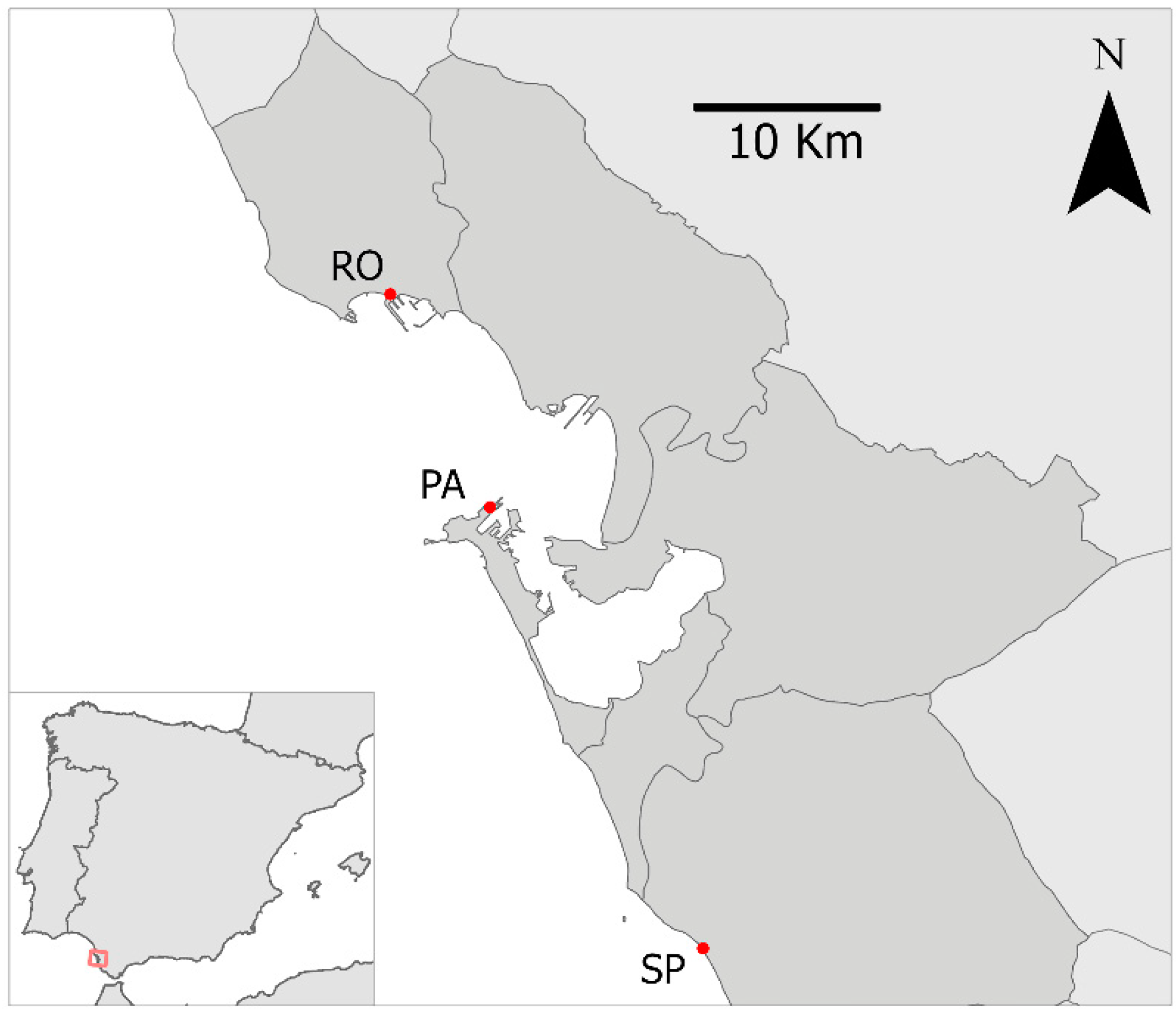
The study was carried out in three marinas of Cádiz Bay, southern Spain (Figure 1), Sancti Petri marina (Chiclana de la Frontera), Rota marina (Rota) and Puerto América marina (Cádiz). This region is adyacent to the Strait of Gibraltar, a major maritime route that is influenced by intense commercial and recreational maritime traffic, which can favour the entry and spread of NIS. Indeed, we selected this area since it is considered a hot spot for biological invasions (e.g., [20,23]).
2.2. Experimental Design and Field Procedure
In this study, we used plastic collectors based on three-dimensionally folded PVC meshes. Ros et al. [14,24] showed the efficacy of this substrate for collecting a high abundance and diversity of peracarid species compared with other methodologies and Scribano et al. [25] its suitability for studying their population dynamics. The structural complexity of these collectors simulates the habitat complexity provided by the local fouling basibionts on which epibiont organisms live. Collectors were deployed in the water column at a 1-m depth (the same depth at which boat hulls tend to be more heavily fouled [26]) through a rope anchored to floating pontoons. Collectors were deployed for two time periods (one month and three months, respectively) to investigate the medium-term colonization ability of caprellid species inhabiting marinas and explore differences in population structure. Eighteen collectors were randomly placed at each marina in April 2017 (54 in total). At each time considered (one month and three months after placement), nine collectors were carefully retrieved from each marina and their associated macrofauna was sorted, preserved in situ in 70% ethanol and enclosed in plastic pots following the protocol established by Ros et al. [24]. Simultaneously with the retrieval of collectors, a complementary sampling focused on the nearest potential source of ‘propagules’ was conducted to characterize the local pool of caprellid species associated with the marina’s fouling community. To avoid the effect of patchiness and to adequately sample local caprellid diversity, each replicate consisted of the collection by hand of a pool of basibionts (mainly macroalgae, bryozoans and hydrozoans) present in the pontoon until a wet volume of 200 mL was collected (see e.g., [27,28]). After draining, the volume of each replicate was ca. 50 mL (coinciding with the volumes of the collectors used). Just after collection, samples were stored in plastic bags and preserved in 70% ethanol.
2.3. Laboratory Processing and Classification of Collected Individuals
The associated macrofauna was carefully separated from their host substrate (i.e., local basibionts and collectors). Caprellid crustaceans were sorted and identified at the species level under a binocular microscope. The abundance of caprellids was expressed as the number of individuals/1000 mL of substrate for suitable comparison due to the different structures of the substrate types. For this purpose, the volume of the basibionts per replicate was measured as the difference between the initial and final volume when placed into a graduated cylinder with a fixed amount of water [29,30]. The mean volume of collectors was 50 mL. Caprellid species were classified as native (N), introduced (I), or cryptogenic (C) for the study area (i.e., the Mediterranean and East Atlantic coast). This last category hosted species traditionally considered native in this region (i.e., species that have been established in the Mediterranean and the East Atlantic coast for hundreds of years) but having unknown origin and a long history of documented anthropogenic transport [31]. To investigate the population characteristics of the introduced caprellid species, specimens were classified as mature females (with developed oostegites), premature females (with undeveloped oostegites), males, or juveniles according to visible signs of maturity or prematurity. The criterion for determining the sex/age group of the specimens was based on Ros et al. [32].
2.4. Statistical Analyses
A three-factor orthogonal and balanced analysis of variance (ANOVA) was used to explore the effect of the type of host substrate (fixed factor with two levels: collectors and local basibionts), time of deployment (fixed factor with two levels: one month and three months), and locality of deployment (random factor with three levels: Sancti Petri, Rota, and Puerto América) in the total abundance and number of caprellid species, as well as in the abundance of each of the caprellid species found in the present study. Previously to ANOVA, the heterogeneity of variances was tested using Cochran’s C-test. If variances were significantly different (p < 0.05), data were transformed with the square root (x + 1) or Ln (x + 1) transformation to achieve homogeneity of variance. Where variances remained heterogeneous even with data transformation, untransformed data was analyzed. ANOVA is a robust statistical test and is relatively unaffected by heterogeneity of variances, particularly in balanced experiments [33]. In such cases, the level of significance was reduced to p < 0.01 to reduce Type I error. Normality was not tested since ANOVA analysis in balanced experiments is robust enough even for non-parametric data [34]. Finally, where ANOVA indicated significant differences for a given factor, the source of the difference was identified using Student-Newman-Keuls (SNK) post-hoc tests. Differences in the caprellid community structure among substrate types in the study periods were tested by the use of a permutational multivariate analysis of variance (PERMANOVA). The experimental design was the same as used for the ANOVA. Analysis was based on the Bray-Curtis similarity. Significance p-values were obtained by computing 9999 permutations of residuals under a reduced model [35]. Pair-wise comparisons were then used between factors to determine where significant differences occurred within the caprellid assemblage. Univariate analyses were performed with GMAV5 [34] and multivariate analyses were conducted using PRIMER v.6 plus PERMANOVA package [36].
3. Results
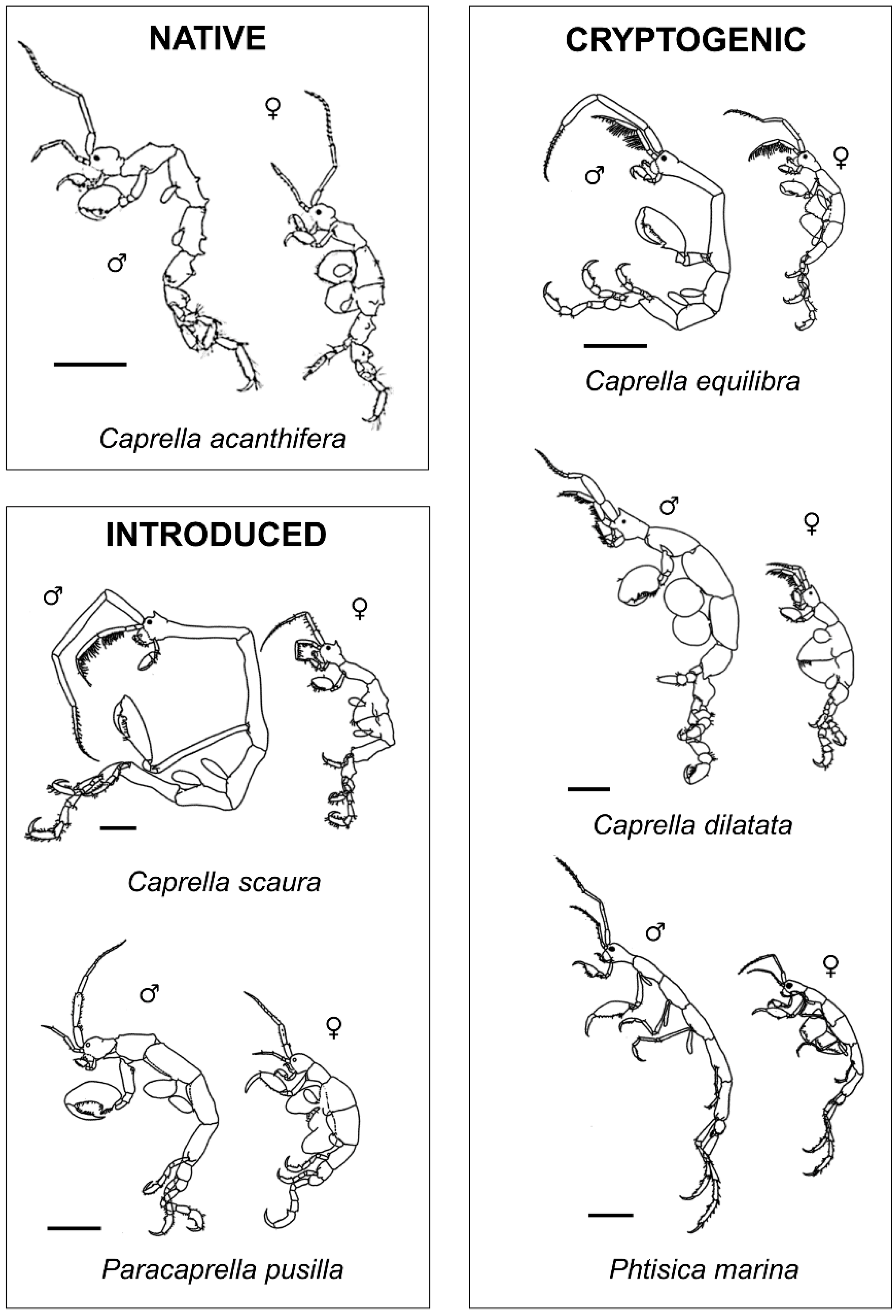
Overall, 3340 caprellids belonging to 3 genera and 6 species were collected during the present study. A total of six caprellid species were found in assemblages colonizing the collectors and five species in the local basibionts. Considering both collectors and local basibionts, we found: 2 introduced species (I), Caprella scaura Templeton, 1836 and Paracaprella pusilla Mayer, 1890; 3 cryptogenic species (C), Phtisica marina Slabber, 1749, Caprella equilibra Say, 1818 and Caprella dilatata Krøyer, 1843; and 1 native species, Caprella acanthifera Leach, 1814 (Figure 2, Table S1).
3.1. Influence of Factors on Caprellid Community Assemblages
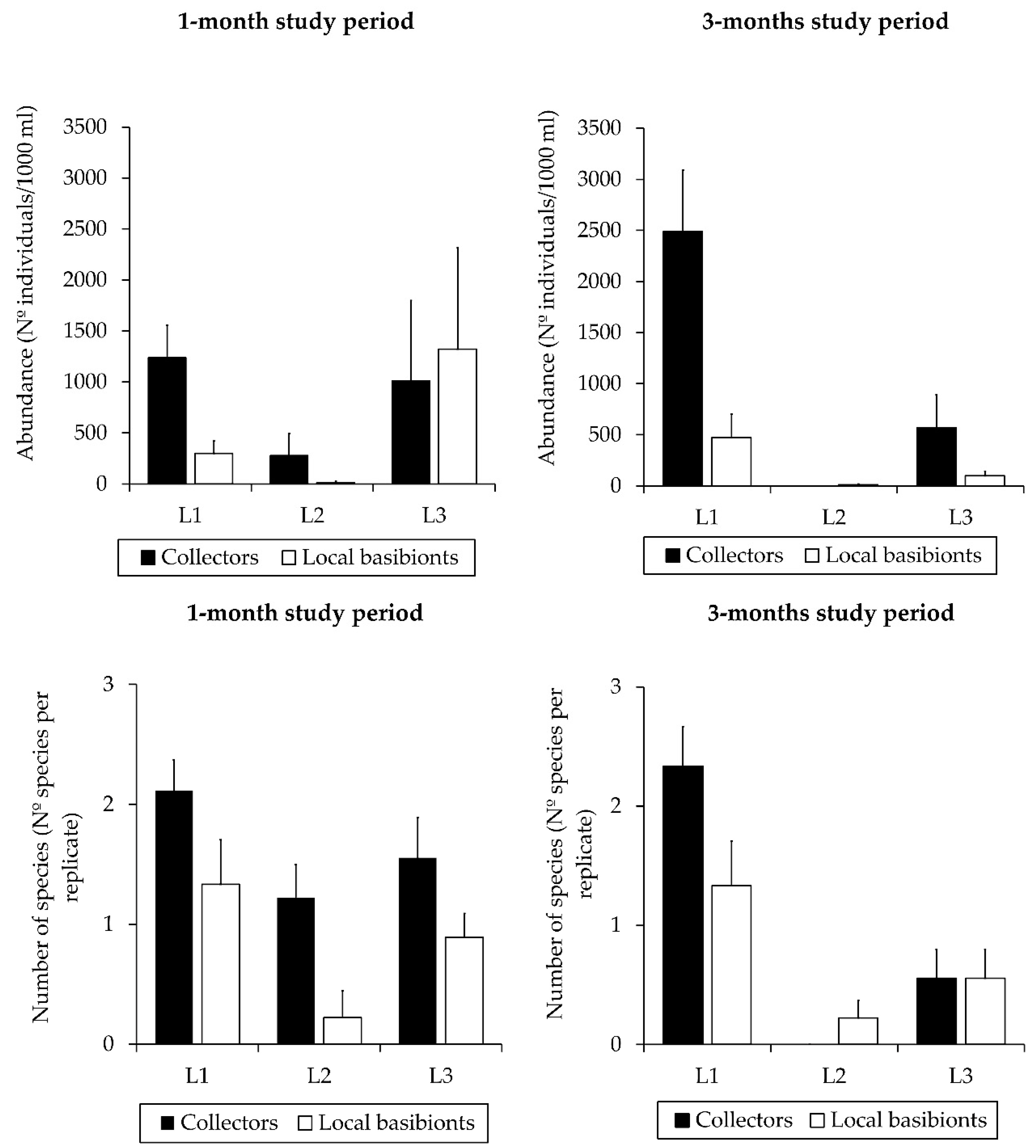
The ANOVA analysis (Table 1) revealed significant differences in total abundance (F = 33.82; p < 0.001) and number of species (F = 26.15; p < 0.001) for the ‘locality’ factor, indicating that both the total number of caprellids per 1000 mL and the number of species per replicate were significantly higher in the marina of Sancti Petri, followed by Puerto América marina and Rota (Figure 3). With regards to the factor ‘substrate’, we did not find significant differences in the total abundance and the number of species between the types of host substrate (i.e., collectors and local basibionts) (Table 1). However, it should be noted that we found a higher number of individuals and a greater number of species in the collectors than in the local basibionts in most of the marinas surveyed in both time periods (1-month study period and 3-months study period) (Figure 3). There were no significant differences in total abundance and number of species when the ‘time’ factor was considered (Table 1).
Focusing on the ANOVA analyses for each caprellid species (Table S2), when we found significant differences, the locality was always the most important factor in determining differences in abundance. The introduced species Paracaprella pusilla, showed a significant interaction between the factors ‘locality’, ‘substrate’, and ‘time’ (F = 13.05; p < 0.001), determining its dominance in the caprellid assemblages collected in Sancti Petri marina after three months, where it was significantly more abundant in collectors than in local basibionts (Table 2 and Table S2). In connection with the cryptogenic species Phtisica marina, its dominance in the caprellid assemblages surveyed in Sancti Petri marina for one month was based on the significant interaction between the factors ‘locality’, ‘substrate’, and ‘time’ (F = 0.0012; p < 0.01), being significantly more abundant in the collectors than in the local basibionts (Table 2 and Table S2).
The PERMANOVA analysis (Table 3) showed a significant interaction between the factors ‘locality’, ‘substrate’ and ‘time’, determining some differences in the community structure between collectors and local basibionts. These differences where not constant neither spatially nor temporally. Specifically, they were found in Sancti Petri and Rota marinas for the one-month study period and remained in Sancti Petri marina in the three-months study period.
3.2. Population Structure of the Introduced Caprellid Species
The most abundant populations of the introduced species Caprella scaura and Paracaprella pusilla were recorded in Puerto América marina and Sancti Petri marina, respectively, being these population structures analyzed.
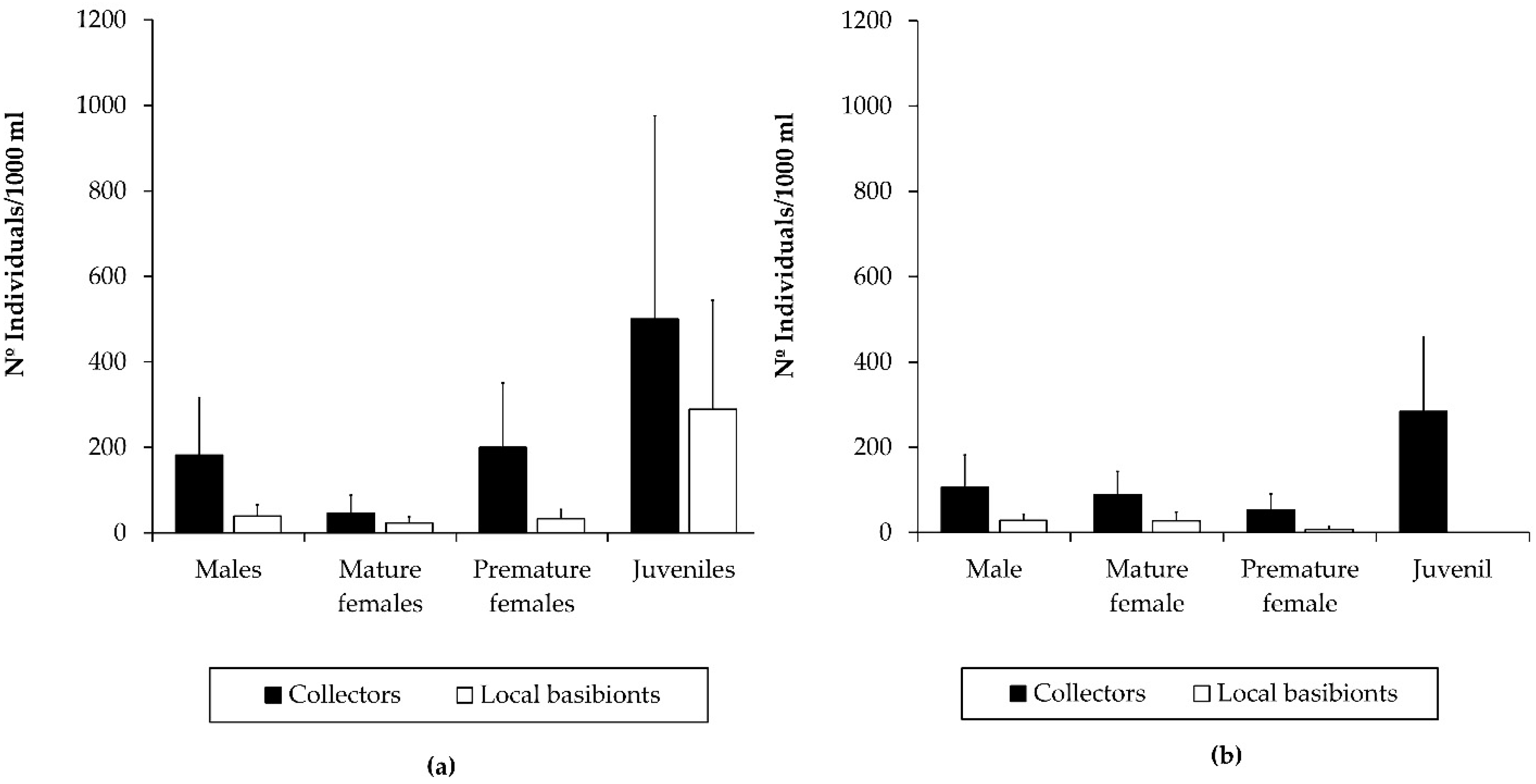
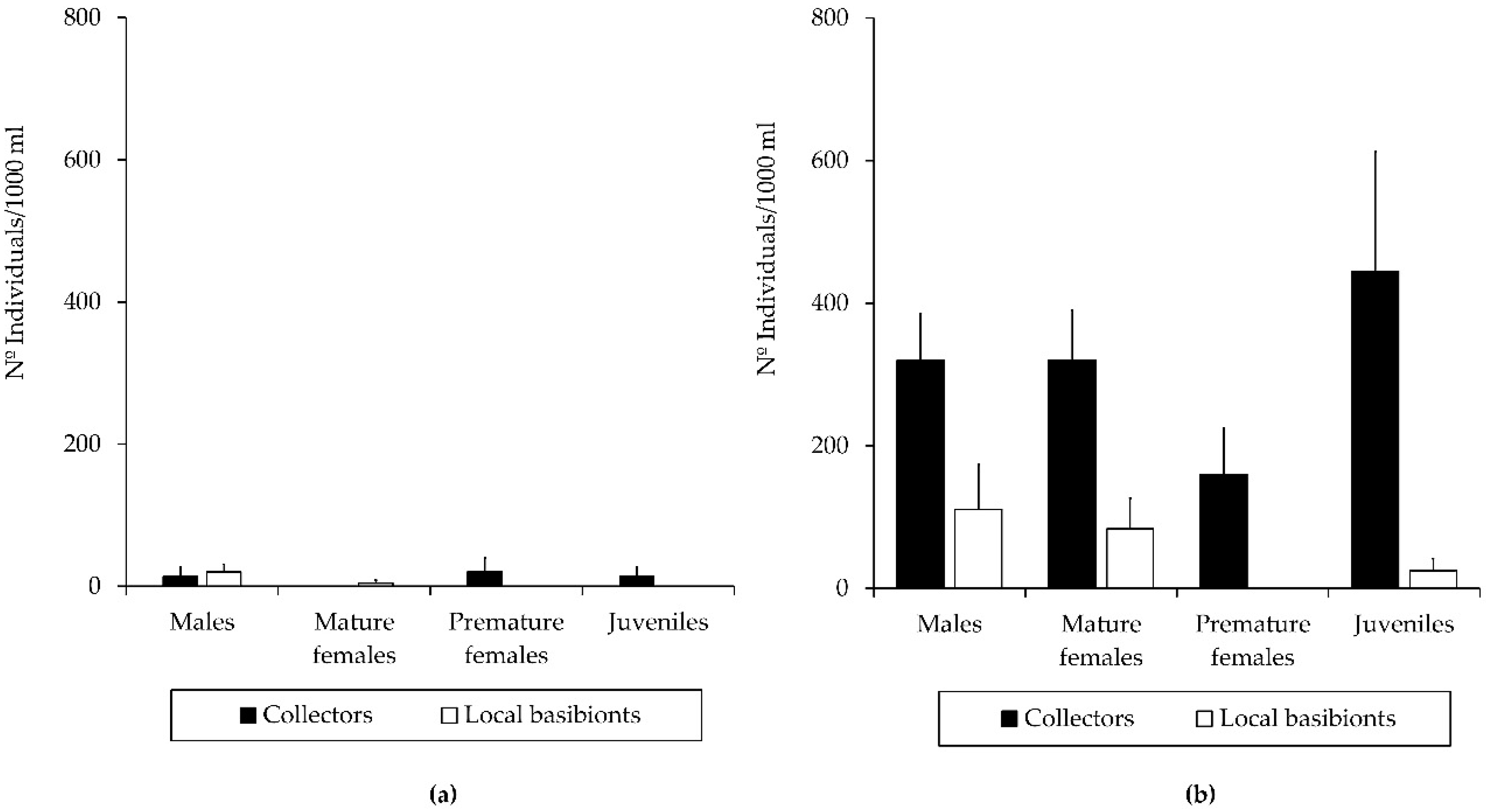
C. scaura was the dominant caprellid species in the collectors deployed in Puerto América marina for both study periods (one and three months), showing the highest densities after one month (Table 2 and Table S1). The abundance of C. scaura was higher in the collectors than in the local basibionts for both study periods, which also revealed a greater representation of all of the demographic categories, as well as a better population structure in the collectors than in the local basibionts (Figure 4). The population structure of C. scaura showed that premature classes (juveniles and premature females) predominated throughout the whole study period, followed by males and mature females in the collectors. This pattern was consistent in the population associated with the local basibionts after one month. After three months, however, no juveniles were found, and premature females were scarcely represented in the population inhabiting local basibionts. Despite population declining in local basibionts after three months, the abundance in collectors were high together with a good population structure (Figure 4).
In the case of P. pusilla, this introduced caprellid species was detected in the marina of Sancti Petri after one month, representing the first record of the species in the locality. After its first reported occurrence, the species became the most abundant among the caprellids found in the Sancti Petri marina after three months (Table 2 and Table S1). Regarding the population structure of P. pusilla, all of the sex/age groups considered were far better represented in the collectors than in the local basibionts (Figure 5), determining the dominance of juveniles followed by males, mature females, and premature females in populations colonizing collectors.
4. Discussion
We found that caprellid species inhabiting marina structures have, in general, a high ability to colonize new artificial substrates. Thus, collectors reflected the caprellid community assemblages inhabiting marina structures after just one month of deployment. Moreover, the two NIS found, Caprella scaura and Paracaprella pusilla, successfully established abundant and well-structured populations in the collectors. Therefore, our results suggest that the hull of a boat which is moored to the marina structures for at least one month may be colonized by the resident caprellid community and inadvertently transport them outside the marina.
4.1. Factors Affecting the Colonization Patterns of Caprellid Community Assemblages
Locality was the most important factor in determining the abundance and the number of species in caprellid community assemblages. We found that both the total abundance, as well as the number of species were significantly different among marinas, with Sancti Petri marina being the one characterized by the highest abundance and number of species, followed by Puerto América and Rota marina. This may be due to the fact that the Sancti Petri marina, being located in a tidal channel, receives a greater amount of sediment rich in organic matter. Considering that caprellids are mainly detritivorous species [37], this could have affected the density of individuals in the locality. Regarding the substrate factor, no significant differences were found between the abundance and number of species in the local basibionts and the collectors. At specific level, we found that some caprellid species inhabiting marina structures showed a high propensity to disperse from their source population (i.e., the local basibionts) and colonize the collectors, where reached higher densities (e.g., Paracaprella pusilla, Caprella scaura or Phtisica marina). These caprellid species with a high affinity for colonizing collectors were probably responsible for the differences in the community structure between assemblages inhabiting local basibionts and collectors. The absence of some caprellid species in the local basibionts that were nevertheless found in the collectors could be an artifact due to the impossibility of sampling all possible basibionts [14]. This potential capacity of caprellids to colonize new artificial substrates in human transport hubs, such as marinas, may play a fundamental role at the beginning of the stowaway pathway. Therefore, considering that all caprellid species found in the collectors (except to the native species Caprella acanthifera) are characterized by having invasive potential (based on the presence of documented history of anthropogenic transport), we discuss below the potential interplay between colonization capacity and invasive potential in this group of crustaceans. In the case of C. acanthifera, although it was a good colonizer of collectors, its densities were noticeably lower than the rest of caprellid species, being absent in several marinas surveyed. This suggests that the native C. acanthifera can disperse and colonize new artificial substrates (such as the hull of a boat). However, it seems that C. acanthifera fail to spread at later stages of the stowaway pathway (e.g., the post-establishment dispersal or during the anthropogenic transport) considering its restricted distribution [38].
Our results support our hypothesis that the new artificial substrates may largely mirror the resident caprellid community after just one month of deployment; and in turn that caprellid species inhabiting marina structures are good colonizers of new artificial substrates. Thus, the hull of a boat which is moored to the marina structures for at least one month may be colonized by the resident caprellid community and inadvertently transport them outside the marina. This probably also occurs in other groups of mobile epibionts inhabiting transport hubs. However, little information exists about the role that colonization ability plays at the beginning of the stowaway pathway, especially in small marine invertebrates with direct development. Interestingly, unlike sessile fouling communities, which generally need deployment periods of at least three months of PVC settlement panels (e.g., [39,40]), the collectors deployed for one month were efficient for early detection of non-indigenous species of mobile epibionts (specifically caprellid crustaceans) and their monitoring in relation to the resident community.
4.2. Colonization Abilities of Potential Stowaways Inhabiting Marinas
Using the colonization patterns of collectors as a quantitative proxy for colonization capacity, we found that the caprellid species inhabiting marina structures have, in general, a high ability to colonize new artificial substrates. However, we also found that some caprellid species showed a greater colonization capacity than others.
In the case of Sancti Petri marina, caprellid community assemblages were characterized by the presence of cryptogenic species after one month, being NIS virtually absent. As we mentioned before, the Sancti Petri marina is situated in a tidal channel, with high hydrodynamic conditions, which could have affected the colonization rates and, therefore, the settlement of new NIS [41,42]. The long-established species C. equilibra, Caprella dilatata, and Phtisica marina have been traditionally considered native to the region; however, they are widely distributed and have a long history of documented anthropogenic transport [20]. Thus, we considered here as cryptogenic species. For example, P. marina has been established in the Mediterranean and the East Atlantic coast for hundreds of years [43]. However, this caprellid species has a cosmopolitan distribution [44] and has been reported from fouling communities including floating plastic debris, aquaculture facilities and numerous harbors around the globe [45,46,47,48], being clearly associated with human activities (e.g., anthropogenic transport). Focusing on the collectors, P. marina was the dominant species, showing a high ability to colonize new artificial substrates.
After three months, however, we found that the introduced species Paracaprella pusilla was the most abundant species in the collectors deployed in Sancti Petri marina. It is worth noting that P. pusilla was detected in collectors after one month, representing its first record in the locality. It is most likely that P. pusilla was introduced via hull fouling of recreational boats travelling from San Fernando marina (Cádiz), where an established population was recently detected [49]. This, combined with suitable environmental conditions may have favored their establishment success. Once an exotic species is introduced into a marina, there is a high spreading probability via recreational boating [50,51,52]. Regarding the local basibionts, P. pusilla exhibited similar abundances than other resident caprellid species, such as the cryptogenic C. dilatata, which suggest a greater colonization ability.
Lastly, it should be noted that, the cryptogenic C. equilibra was the dominant species in the local basibionts surveyed in Puerto América marina after one month. However, it was scarcely represented in the collectors, where the introduced C. scaura dominated in both study periods. The cosmopolitan species C. equilibra was one of the most found caprellid species in southern Spain [53], reaching high densities associated with fouling communities in marinas [54] and harbour areas [55]. However, after the detection of its congener C. scaura in marinas of southern Spain, a decline in C. equilibra populations was observed when the introduced species was present. The high abundance of C. equilibra noticed in collectors deployed in another nearby marina (Sancti Petri), where C. scaura was absent, supports that C. equilibra and C. scaura compete for the occupation of substrate. It also supports the notion that C. scaura is capable of displacing their ecologically similar congener [22,32,56].
Therefore, we found that the introduced species C. scaura and P. pusilla, and the cryptogenic species P. marina showed a higher ability to colonize new artificial substrates than the rest of resident caprellid species. This potential variation in colonization abilities of marine organisms may increase the chances of human vector uptake (or human vector colonization), the frequency of transport and, in turn, invasion success. Considering that these species are characterized by having a high invasive potential (considered here as the ability for successful anthropogenic transportation), our results suggest a potential relationship between the colonization ability and the invasive potential in this group of crustaceans.
4.3. The Colonization Dynamics of the Introduced Species
The introduced species Caprella scaura, originally described from Mauritius by Templeton (1836), is considered one of the most widespread invaders in the last 10 years [57] with records in all oceans of the world except for the Arctic [52]. In the northern hemisphere, there is sound evidence of its successful spread as stowaway on human transport vectors (i.e., via hull fouling and ballast water) [52,58,59,60]. According to these studies, most of their populations in European Seas have an Indo-Pacific origin via the Suez Canal, most likely with Mediterranean populations acting as stepping-stone events. In the Iberian Peninsula C. scaura was first recorded by Martínez and Adarraga [61] on the northeastern coast of Spain (Roses Bay, Girona) in 2005. A short time later, an important post-introduction range expansion of C. scaura was reported along marinas in the Mediterranean Sea and the South Atlantic coast of the Iberian Peninsula [59], showing its high potential for invasion via recreational boating.
C. scaura was the dominant species in the collectors deployed in Puerto América marina during the whole study period. The population structure was consistent for both study periods, with all of the demographic categories (i.e., males, mature females, premature females, and juveniles) represented in the collectors. Premature classes (i.e., juveniles and premature females) dominated in collectors followed by males and mature females. This pattern was consistent with the assemblages inhabiting the local basibionts after one month. Nevertheless, the introduced species exhibited a better structure and a greater representation of demographic classes in the collectors than in the local basibionts for both study periods, showing a high tendency to colonize new empty niches. This suggests that C. scaura can efficiently colonize new artificial substrates (i.e., the collectors) in less than a month, with reproductive success and survival of its offspring. Thus, this high ability to colonize new substrates could increase the chances of human-vector colonization (or human-vector uptake) and, in turn, promote success in the early stages of the invasion process (i.e., leading to unintentional introductions), as well as in the subsequent stages (i.e., establishment of self-sustaining populations, and spread) [14].
The introduced species Paracaprella pusilla, first described by Mayer (1890) from Brazil, is thought to originate in the Atlantic coast of Central and South America. Since its original description, P. pusilla has been widely reported in both tropical and subtropical areas around the world, mainly on fouling communities colonizing human-made infrastructures (i.e., marinas, harbors, or desalination plants) ([49,51,62,63] and references therein).
In European Seas, its first recorded occurrence was in the marina of Puerto América (Cádiz) by Ros and Guerra-García [51], one of the marinas surveyed in the present study. Soon afterwards, this tropical caprellid species was reported in several locations of the Mediterranean Sea (the Balearic Islands, Tunisia, and Israel) [63,64,65,66]. Cabezas et al. [49] suggested the Strait of Gibraltar as its main introduction pathway in temperate European waters with native populations of the East Atlantic coast acting as potential source [38,51,65]. Thus, P. pusilla could have spread from Puerto America marina to San Fernando marina [49], and from there to Sancti Petri marina, where the species was detected for the first time in the present study. Therefore, P. pusilla has demonstrated a high potential to spread as stowaway on human transport vectors (i.e., recreational boating).
Regarding its population characteristics, the flourishing population found in collectors deployed in Sancti Petri after three months, showed the predominance of juveniles followed by males, mature females, and premature females. This suggests that P. pusilla can efficiently colonize novel empty niches (i.e., the collectors) with reproduction success and survival of its offspring, as well as their potential adaptation to novel habitat conditions. Furthermore, the assemblages colonizing collectors exhibited a better structure and a greater representation of demographic classes compared to the local basibionts. This high tendency to successfully colonize new empty substrates could increase the likelihood of human-vector uptake (or human-vector colonization), the frequency of transport and, in turn, the invasion success [14]. Considering each successive establishment of self-sustaining populations may increase the variety of locations connected by vectors in the invasion pathway [6], further research is needed to determine the status of P. pusilla populations in southwest Spain.
5. Conclusions
We found that caprellid species inhabiting marinas structures typically have, despite their direct development, a high capacity to colonize novel artificial substrates. This high colonization ability could increase the likelihood of human vector uptake (or human vector colonization), as well as their subsequent establishment in the recipient location. We suggest a potential interplay between colonization ability and invasion success of stowaway organisms in this group of crustaceans. Further research is needed to understand the potential variation in colonization abilities between marine invertebrate species with direct development and fully integrate into research on managing the risk of the stowaway pathway. Additionally, the collectors were efficient in detecting changes in the population dynamics of colonizing species and, therefore, they are suggested as a useful tool in the early detection of NIS and their monitoring in relation to resident species.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w14172659/s1. Table S1: Mean abundance ± standard error (individuals/1000 mL) of caprellids species collected in the local basibionts and the collectors for the two study periods at each marina surveyed; Table S2: Results of three-way analysis of variance (ANOVA) for the effects of substrate type, time, and locality of deployment on the abundance of caprellid species collected.
Author Contributions
Study conception and design: T.R., M.R. and J.M.G.-G.; data collection: T.R., M.R. and J.M.G.-G.; data analysis: T.R.; investigation: T.R., M.R. and J.M.G.-G.; draft manuscript preparation: T.R.; manuscript review and editing: M.R. and J.M.G.-G.; supervision: M.R. and J.M.G.-G.; project administration: M.R.; funding acquisition: M.R. All authors have read and agreed to the published version of the manuscript.
Funding
Financial support for this study was provided by the Andalusian Government (Project I+D+I, US-1265621, co-financed by the Fondo Europeo de Desarrollo Regional-FEDER).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are contained in the paper and Supplementary Materials. Further data are available upon request to the corresponding author.
Acknowledgments
We would like to thank the research groups team of the Marine Biology Laboratory (University of Seville) and the Nudilab Laboratory (University of Cádiz) that assisted the field and laboratory work; We are extremely grateful to Victor López de la Rosa for his technical support, and to the editor and the anonymous referees for their constructive and very helpful comments and suggestions. We sincerely thank the marinas staff for allowing us to carry out the field experiments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hulme, P.E.; Bacher, S.; Kenis, M.; Klotz, S.; Kühn, I.; Minchin, D.; Nentwig, W.; Olenin, S.; Panov, J.; Pergl, J.; et al. Grasping at the routes of biological invasions: A framework for integrating pathways into policy. J. App. Ecol. 2008, 45, 403–414. [Google Scholar] [CrossRef]
- Banks, N.C.; Paini, D.R.; Bayliss, K.L.; Hodda, M. The role of global trade and transport network topology in the human-mediated dispersal of alien species. Ecol. Lett. 2015, 18, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Tingley, R.; García-Díaz, P.; Arantes, C.R.R.; Cassey, P. Integrating transport pressure data and species distribution models to estimate invasion risk for alien stowaways. Ecography 2018, 41, 635–646. [Google Scholar] [CrossRef]
- Fofonoff, P.W.; Ruiz, G.M.; Hines, A.H.; Steves, B.D.; Carlton, J.T. Four centuries of biological invasions in tidal waters of the Chesapeake Bay region. In Biological Invasions in Marine Ecosystems; Rilov, G., Crooks, A., Eds.; Springer: Heidelberg, Germany, 2009; pp. 479–506. ISBN 978-3-5407-9235-2. [Google Scholar]
- Carlton, J.T. Bioinvasion ecology: Assessing invasion impact and scale. In Invasive Aquatic Species of Europe. Distribution, impacts, and management; Leppäkoski, E., Gollasch, S., Olenin, S., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 7–19. ISBN 978-90-481-6111-9. [Google Scholar]
- Floerl, O.; Inglis, G.J. Starting the invasion pathway: The interaction between source populations and human transport vectors. Biol. Invasions. 2005, 7, 589–606. [Google Scholar] [CrossRef]
- Carlton, J.T. Man’s role in changing the face of the ocean: Biological invasions and implications for conservation of near-shore environments. Conserv. Biol. 1989, 3, 265–273. [Google Scholar] [CrossRef]
- Williamson, M. Biological Invasions; Springer: Dordrecht, The Netherlands, 1996; ISBN 978-0-412-59190-7. [Google Scholar]
- Ruiz, G.M.; Carlton, J.T.; Grosholz, E.D.; Hines, A.H. Global invasions of marine and estuarine habitats by non-indigenous species: Mechanisms, extent, and consequences. Am. Zool. 1997, 37, 621–632. [Google Scholar] [CrossRef]
- Puth, L.M.; Post, D.M. Studying invasion: Have we missed the boat? Ecol. Lett. 2005, 8, 715–721. [Google Scholar] [CrossRef]
- Floerl, O.; Inglis, G.J.; Dey, K.; Smith, A. The importance of transport hubs in stepping-stone invasions. J. App. Ecol. 2009, 46, 7–45. [Google Scholar] [CrossRef]
- Minchin, D.; Floerl, O.; Savini, D.; Occhipinti-Ambrogi, A. Small craft and the spread of exotic species. In The Ecology of Transportation: Managing Mobility for the Environment; Davenport, J., Davenport, J.L., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 99–118. ISBN 978-1-4020-4504-2. [Google Scholar]
- Murray, C.C.; Pakhomov, E.A.; Therriault, T.W. Recreational boating: A large unregulated vector transporting marine invasive species. Divers. Distrib. 2011, 17, 1161–1172. [Google Scholar] [CrossRef]
- Ros, M.; Navarro-Barranco, C.; González-Sánchez, M.; Ostalé-Valriberas, E.; Cervera-Currado, L.; Guerra-García, J.M. Starting the stowaway pathway: The role of dispersal behaviour in the invasion success of low-mobile marine species. Biol. Invasions. 2020, 22, 2797–2812. [Google Scholar] [CrossRef]
- Chapple, D.G.; Simmonds, S.M.; Wong, B.B. Can behavioral and personality traits influence the success of unintentional species introductions? Trends. Ecol. Evol. 2012, 27, 57–64. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967; ISBN 978-0-6910-8836-5. [Google Scholar]
- Ebenhard, T. Colonization in metapopulations: A review of theory and observations. Biol. J. Linn. Soc. 1991, 42, 105–121. [Google Scholar] [CrossRef]
- Cáceres, C.E.; Soluk, D.A. Blowing in the wind: A field test of overland dispersal and colonization by aquatic invertebrates. Oecologia 2002, 131, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Buschbaum, C.; Gutow, L. Mass occurrence of an introduced crustacean (Caprella mutica) in south-eastern North Sea. Helgol. Mar. Res. 2005, 59, 252–253. [Google Scholar] [CrossRef]
- Ros, M. Invasion Dynamic of Introduced Caprellids (Crustacea: Amphipoda) in the Mediterranean Sea and Adjacent Waters. Global Distribution, Ecology, and Vectors of Spread. Ph.D. Dissertation, University of Seville, Seville, Spain, 2015. [Google Scholar]
- Cabezas, M.P.; Navarro-Barranco, C.; Ros, M.; Guerra-García, J.M. Long-distance dispersal, low connectivity, and molecular evidence of a new cryptic species in the obligate rafter Caprella andreae Mayer, 1890 (Crustacea: Amphipoda: Caprellidae). Helgol. Mar. Res. 2013, 67, 483–497. [Google Scholar] [CrossRef]
- Ros, M.; Vázquez-Luis, M.; Guerra-Garcia, J.M. Environmental factors modulating the extent of impact in coastal invasions: The case of a widespread invasive caprellid (Crustacea: Amphipoda) in the Iberian Peninsula. Mar. Poll. Bull. 2015, 98, 247–258. [Google Scholar] [CrossRef]
- Drake, J.M.; Lodge, D.M. Global hot spots of biological invasions: Evaluating options for ballast–water management. Proc. R. Soc. Lond. B. Biol. Sci. 2004, 271, 575–580. [Google Scholar] [CrossRef]
- Ros, M.; López De La Rosa, V.L.; Revanales, T.; Spinelli, A.; Fernández-Romero, A.; Cervera-Currado, L.; Guerra-García, J.M. Towards a standardized methodology for monitoring and assessing marine mobile epibenthic communities across spatio-temporal scales. In Proceedings of the XX Iberian Symposium on Marine Biology Studies (SIEBM XX), Braga, Portugal, 9–12 September 2019. [Google Scholar] [CrossRef]
- Scribano, G.; Marchini, A.; Ros, M. Population dynamics and life history traits of the non-indigenous Paranthura japonica Richardson (1909) in a recently invaded Mediterranean site. Reg. Stud. Mar. Sci. 2021, 46, 101883. [Google Scholar] [CrossRef]
- Dafforn, K.A.; Johnston, E.L.; Glasby, T.M. Shallow moving structures promote marine invader dominance. Biofouling 2009, 25, 277–287. [Google Scholar] [CrossRef]
- Thiel, M.; Guerra-García, J.M.; Lancellotti, D.A.; Vásquez, N. The distribution of littoral caprellids (Crustacea: Amphipoda: Caprellidae) along the Pacific coast of continental Chile. Rev. Chil. Hist. Nat. 2003, 76, 203–218. [Google Scholar] [CrossRef]
- Guerra-Garcia, J.M.; Cabezas, P.; Baeza-Rojano, E.; Espinosa, F.; García-Gómez, J.C. Is the north side of the Strait of Gibraltar more diverse than the south side? A case study using the intertidal peracarids (Crustacea: Malacostraca) associated to the seaweed Corallina elongata. J. Mar. Biolog. 2009, 89, 387–397. [Google Scholar] [CrossRef]
- Guerra-García, J.M.; Ganesh, T.; Jaikumr, M.; Rama, A.V. Caprellids (Crustacea: Amphipoda) from India. Helgol. Mar. Res. 2010, 64, 297–310. [Google Scholar] [CrossRef]
- Pereira, S.G.; Lima, F.P.; Queiroz, N.C.; Ribeiro, P.A.; Santos, A.M. Biogeographic patterns of intertidal macroinvertebrates and their association with macroalgae distribution along the Portuguese coast. Hydrobiologia 2006, 555, 185–192. [Google Scholar] [CrossRef]
- Carlton, J.T. Biological Invasions and Cryptogenic Species. Ecology 1996, 77, 1653–1655. [Google Scholar] [CrossRef]
- Ros, M.; Guerra-García, J.M.; González-Macías, M.; Saavedra, Á.; López-Fe, C.M. Influence of fouling communities on the establishment success of alien caprellids (Crustacea: Amphipoda) in Southern Spain. Mar. Biol. Res. 2013, 9, 261–273. [Google Scholar] [CrossRef]
- Underwood, A.J. Experiments in Ecology: Their Logical Design and Interpretation Using Analysis of Variance; Cambridge University Press: Cambridge, UK, 1997; ISBN 978-0-5118-0640-7. [Google Scholar]
- Underwood, A.J.; Chapman, M.G.; Richards, S. GMAV-5 for Windows. An Analysis of Variance Programme; University of Sydney: Sydney, Australia, 2002. [Google Scholar]
- Anderson, M.J.; Legendre, P. An empirical comparison of permutation methods for tests of partial regression coefficients in a linear model. J. Stat. Comput. Simul. 1999, 62, 271–303. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Guerra-García, J.M.; Tierno de Figueroa, J.T. What do caprellids (Crustacea: Amphipoda) feed on? Mar. Biol. 2009, 156, 1881–1890. [Google Scholar] [CrossRef]
- Ros, M.; Vázquez-Luis, M.; Guerra-García, J.M. The role of marinas and recreational boating in the occurrence and distribution of exotic caprellids (Crustacea: Amphipoda) in the Western Mediterranean: Mallorca Island as a case study. J. Sea Res. 2013, 83, 94–103. [Google Scholar] [CrossRef]
- De Rivera, C.E.; Ruiz, G.; Crooks, J.; Wasson, K.; Lonhart, S. Broadscale non-indigenous species monitoring along the west coast in national marine sanctuaries and national estuarine research reserves. Environ. Sci. Manag. 2005, 76, 1–56. [Google Scholar]
- Tamburini, M.; Keppel, E.; Marchini, A.; Repetto, M.F.; Ruiz, G.M.; Ferrario, J.; Occhipinti-Ambrogi, A. Monitoring non-indigenous species in port habitats: First application of a standardized North American protocol in the Mediterranean Sea. Front. Mar. Sci. 2021, 8, 700730. [Google Scholar] [CrossRef]
- Floerl, O.; Inglis, G.J. Boat harbour design can exacerbate hull fouling. Austral. Ecol. 2003, 28, 116–127. [Google Scholar] [CrossRef]
- Koehl, M.R.A. Mini review: Hydrodynamics of larval settlement into fouling communities. Biofouling 2007, 23, 357–368. [Google Scholar] [CrossRef] [PubMed]
- McCain, J.C.; Steinberg, J.E. Amphipoda-I, Caprellidea-I. In Crustaceorum Catalogus; Gruner, H.E., Holthuis, L.B., Eds.; W. Junk: The Hague, The Netherlands, 1970; Volume 2, pp. 1–78. [Google Scholar]
- Bellan-Santini, D.; Ruffo, S. Faunistics and Zoogeography. In The Amphipoda of the Mediterranean, Part 4. Mémoires de l’Institut Océanographique (Monaco); Ruffo, S., Ed.; Institut Océanographique: Monaco, 1998; Volume 13, pp. 895–911. ISBN 2-7260-0201-3. [Google Scholar]
- Katsanevakis, S.; Crocetta, F. Pathways of introduction of marine alien species in European waters and the Mediterranean—A possible undermined role of marine litter. In Proceedings of the Marine Litter in the Mediterranean and Black Seas, CIESM Workshop, Tirana, Albania, 18–21 June 2014; CIESM Publisher: Monaco, Monaco, 2014; Volume 46, pp. 6–68. [Google Scholar]
- Fernandez-Gonzalez, V.; Sanchez-Jerez, P. Fouling assemblages associated with off-coast aquaculture facilities: An overall assessment of the Mediterranean Sea. Mediterr. Mar. Sci. 2017, 18, 87–96. [Google Scholar] [CrossRef]
- Leclerc, J.C.; Viard, F. Habitat formation prevails over predation in influencing fouling communities. Ecol. Evol. 2018, 8, 477–492. [Google Scholar] [CrossRef] [PubMed]
- Srinivas, T.; Sukumaran, S.; Dias, H.Q. Extended distribution of Phtisica marina Slabber, 1769 (Crustacea: Amphipoda): First observation of alien Caprellid in the coastal waters of Indian subcontinent. Bioinvasions Rec. 2019, 8, 96–107. [Google Scholar] [CrossRef]
- Cabezas, M.P.; Ros, M.; Dos Santos, A.M.; Martinez-Laiz, G.; Xavier, R.; Montelli, L.; Hoffman, R.; Fersi, A.; Claude-Davin, J.; Guerra-Garcia, J.M. Unravelling the origin and introduction pattern of the tropical species Paracaprella pusilla Mayer, 1890 (Crustacea, Amphipoda, Caprellidae) in temperate European waters: First molecular insights from a spatial and temporal perspective. NeoBiota 2019, 47, 43–80. [Google Scholar] [CrossRef]
- Ashton, G.V. Distribution and Dispersal of the Non-Native Caprellid Amphipod, Caprella mutica Schurin, 1935. Ph.D. Thesis, University of Aberdeen, Aberdeen, UK, 2006. [Google Scholar]
- Ros, M.; Guerra-García, J.M. On the occurrence of the tropical caprellid Paracaprella pusilla Mayer, 1890 (Crustacea: Amphipoda) in Europe. Mediterr. Mar. Sci. 2012, 13, 134–139. [Google Scholar] [CrossRef]
- Martínez-Laiz, G.; Guerra-García, J.M.; Ros, M.; Fenwick, D.; Bishop, J.D.; Horton, T.; Faase, M.A.; Cabezas, M.P. Hitchhiking northwards: On the presence of the invasive skeleton shrimp Caprella scaura in the UK. Mar. Biodiv. 2021, 51, 78. [Google Scholar] [CrossRef]
- Guerra-García, J.M.; Ros, M.; Gordillo, I.; Cabezas, M.D.P.; Baeza-Rojano, E.; Izquierdo, D.; Corzo, J.; Domínguez, J.; Varona, S. Distribution patterns of intertidal and shallow water caprellids associated with macroalgae along the Iberian Peninsula. Zool. Baetica. 2011, 21, 101–129. [Google Scholar]
- Baeza-Rojano, E.; Guerra-García, J.M. Ciclo de vida bajo condiciones de laboratorio de los caprélidos (Crustacea: Amphipoda) del sur de la Península Ibérica: Caprella equilibra y Caprella dilatata (Caprellidae) y Phtisica marina (Phtisicidae). Zool. Baetica. 2013, 24, 155–186. [Google Scholar]
- Conradi, M.; López-González, P.J.; Cervera, J.L.; García-Gómez, J.C. Seasonality and spatial distribution of peracarids associated with the bryozoan Bugula neritina in Algeciras Bay, Spain. J. Crust. Biol. 2000, 20, 334–349. [Google Scholar] [CrossRef]
- Guerra-García, J.M.; Ros, M.; Dugo-Cota, A.; Burgos, V.; Flores-León, A.M.; Baeza-Rojano, E.; Núñez, J. Geographical expansion of the invader Caprella scaura (Crustacea: Amphipoda: Caprellidae) to the East Atlantic coast. Marine Biol. 2011, 158, 2617–2622. [Google Scholar] [CrossRef]
- Galil, B.S.; Marchini, A.; Occhipinti-Ambrogi, A.; Minchin, D.; Narščius, A.; Ojaveer, H.; Olenin, S. International arrivals: Widespread bioinvasions in European Seas. Ethol. Ecol. Evol. 2014, 26, 152–171. [Google Scholar] [CrossRef]
- Cabezas, M.P.; Xaiver, R.; Branco, M.; Santos, A.M.; Guerra-García, J.M. Invasion history of Caprella scaura Templeton, 1836 (Amphipoda: Caprellidae) in the Iberian Peninsula: Multiple introductions revealed by mitochondrial sequence data. Biol. Invasions 2014, 16, 2221–2245. [Google Scholar] [CrossRef]
- Ros, M.; Guerra-García, J.M.; Navarro-Barranco, C.; Cabezas, M.P.; Vázquez-Luis, M.P. The spreading of the non-native caprellid (Crustacea: Amphipoda) Caprella scaura Templeton, 1836 into southern Europe and northen Africa: A complicated taxonomic history. Mediterr. Mar. Sci. 2014, 15, 145–155. [Google Scholar] [CrossRef]
- Martínez-Laiz, G.; Ros, M.; Guerra-García, J.; Faasse, M.; Santos, A.; Cabezas, M. Using molecular data to monitor the post establishment evolution of the invasive skeleton shrimp Caprella scaura. Mar. Environ. Res. 2021, 166, 105266. [Google Scholar] [CrossRef] [PubMed]
- Martínez, J.; Adarraga, I. First record of invasive caprellid Caprella scaura Templeton, 1836 sensu lato (Crustacea: Amphipoda: Caprellidae) from the Iberian Peninsula. Aquat. Invasions 2008, 3, 165–171. [Google Scholar] [CrossRef]
- Fofonoff, P.W.; Ruiz, G.M.; Steves, B.; Simkanin, C.; Carlton, J.T. National Exotic Marine and Estuarine Species Information System. Available online: http://invasions.si.edu/nemesis (accessed on 12 December 2021).
- Lo Brutto, S.; Iaciofano, D.; Guerra-García, J.M.; Lubinevsky, H.; Galil, B.S. Desalination effluents and the establishment of the non-indigenous skeleton shrimp Paracaprella pusilla Mayer, 1890 in the south-eastern Mediterranean. BioInvasions Rec. 2019, 8, 661–669. [Google Scholar] [CrossRef]
- Fersi, A.; Davin, J.; Pezy, J.; Neifar, L. Amphipods from tidal channels of the Gulf of Gabès (central Mediterranean Sea). Mediterra. Mar. Sci. 2018, 19, 430–443. [Google Scholar] [CrossRef]
- Ros, M.; Vázquez-Luis, M.; Guerra-García, J.M. The tropical caprellid amphipod Paracaprella pusilla: A new alien crustacean in the Mediterranean Sea. Helgol. Mar. Res. 2013, 67, 675–685. [Google Scholar] [CrossRef]
- Ros, M.; Guerra-García, J.M.; Hoffman, R. First record of the exotic caprellid amphipod Paracaprella pusilla Mayer, 1890 in the eastern Mediterranean. Mar. Biodivers. 2016, 46, 281–284. [Google Scholar] [CrossRef]
Figure 1.
Map of the study area showing the three sampling marinas, Puerto de Sancti Petri (SP), Puerto de Rota (RO) and Puerto América (PA), in the Bay of Cádiz (Spain).
Figure 1.
Map of the study area showing the three sampling marinas, Puerto de Sancti Petri (SP), Puerto de Rota (RO) and Puerto América (PA), in the Bay of Cádiz (Spain).

Figure 2.
Lateral view figures of the caprellid species found during the present study. Scale bars: 1 mm.
Figure 2.
Lateral view figures of the caprellid species found during the present study. Scale bars: 1 mm.

Figure 3.
Total abundances of caprellids (mean individuals/1000 mL ± standard error; n = 9) in collectors and local basibionts in each locality surveyed (L1: Sancti Petri marina, L2: Rota marina, L3: Puerto América marina) after one and three months. Number of species per replicate (mean ± standard error; n = 9) in collectors and local basibionts in each locality surveyed after one and three months. Significant differences (p < 0.05) were detected among the three localities (see Table 1 for details of the Student–Newman–Keul (SNK) tests).
Figure 3.
Total abundances of caprellids (mean individuals/1000 mL ± standard error; n = 9) in collectors and local basibionts in each locality surveyed (L1: Sancti Petri marina, L2: Rota marina, L3: Puerto América marina) after one and three months. Number of species per replicate (mean ± standard error; n = 9) in collectors and local basibionts in each locality surveyed after one and three months. Significant differences (p < 0.05) were detected among the three localities (see Table 1 for details of the Student–Newman–Keul (SNK) tests).

Figure 4.
Population structure of Caprella scaura colonizing collectors versus inhabiting local basibionts in Puerto América marina in: (a) the one-month study period (b) and the three-months study period.
Figure 4.
Population structure of Caprella scaura colonizing collectors versus inhabiting local basibionts in Puerto América marina in: (a) the one-month study period (b) and the three-months study period.

Figure 5.
Population structure of Paracaprella pusilla colonizing collectors versus inhabiting local basibionts in Sancti Petri marina in (a) the one-month study period (b) and the three-months study period.
Figure 5.
Population structure of Paracaprella pusilla colonizing collectors versus inhabiting local basibionts in Sancti Petri marina in (a) the one-month study period (b) and the three-months study period.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Results of three-way analysis of variance (ANOVA) for the effect of the type of host substrate (SU, CO = collectors and LB = local basibionts), time (TI, T1 = one month, and T3 = three months) and locality (LO, L1 = Sancti Petri, L2 = Rota and L3 = Puerto América) on the total abundance and number of caprellid species collected. MS = mean square. F = Fisher’s statistic. p = level of significance. df = degrees of freedom.
Table 1.
Results of three-way analysis of variance (ANOVA) for the effect of the type of host substrate (SU, CO = collectors and LB = local basibionts), time (TI, T1 = one month, and T3 = three months) and locality (LO, L1 = Sancti Petri, L2 = Rota and L3 = Puerto América) on the total abundance and number of caprellid species collected. MS = mean square. F = Fisher’s statistic. p = level of significance. df = degrees of freedom.
| Source of Variation | df | MS | F | p | Source of Variation | df | MS | F | p |
|---|---|---|---|---|---|---|---|---|---|
| Total Abundance | Number of Species | ||||||||
| Time = TI | 1 | 23.3316 | 1.67 | 0.3258 | Time = TI | 1 | 4.0833 | 2.41 | 0.2608 |
| Locality = LO | 2 | 181.3196 | 33.82 | 0.0000 *** | Locality = LO | 2 | 17.1944 | 26.15 | 0.0000 *** |
| Substrate = SU | 1 | 62.3382 | 5.50 | 0.1437 | Substrate = SU | 1 | 7.7870 | 9.24 | 0.0933 |
| TI × LO | 2 | 14.0017 | 2.61 | 0.0786 | TI × LO | 2 | 1.6944 | 2.58 | 0.0812 |
| TI × SU | 1 | 9.7162 | 0.76 | 0.4757 | TI × SU | 1 | 2.0833 | 1.74 | 0.3175 |
| LO × SU | 2 | 11.3397 | 2.12 | 0.1262 | LO × SU | 2 | 0.8426 | 1.28 | 0.2823 |
| TI × LO × SU | 2 | 12.8171 | 2.39 | 0.0970 | TI × LO × SU | 2 | 1.1944 | 1.82 | 0.1681 |
| Residual | 96 | 5.3607 | Residual | 96 | 0.6574 | ||||
| Total | 107 | Total | 107 | ||||||
| Cochran’s C-test | C = 0.2018 | Cochran’s C-test | C = 0.1585 | ||||||
| Transformation | Ln (X + 1) | Transformation | None | ||||||
| SNK | LO: L1 > L3 > L2 | SNK | LO: L1 > L3 > L2 | ||||||
*** p < 0.001.
Table 2.
Population density (individuals/1000 mL) of caprellid species found on the local basibionts and collectors for the two study periods (one month and three months, respectively, from the deployment of collectors) at each marina surveyed (Sancti Petri marina (L1), Rota marina (L2) and Puerto América marina (L3)). N: native species for the study area. I: introduced species in the study area. C: cryptogenic species.
Table 2.
Population density (individuals/1000 mL) of caprellid species found on the local basibionts and collectors for the two study periods (one month and three months, respectively, from the deployment of collectors) at each marina surveyed (Sancti Petri marina (L1), Rota marina (L2) and Puerto América marina (L3)). N: native species for the study area. I: introduced species in the study area. C: cryptogenic species.
 |

Table 3.
Results of PERMANOVA examining the abundance of caprellid community assemblages found in each substrata type (SU, CO = experimental collectors and LB = local basibionts), time of deployment (TI, T1 = one month, and T3 = three months) and each locality (LO, L1 = Sancti Petri, L2 = Rota and L3 = Puerto América). MS = mean square; df = degrees of freedom; P(MC) = Monte Carlo p values.
Table 3.
Results of PERMANOVA examining the abundance of caprellid community assemblages found in each substrata type (SU, CO = experimental collectors and LB = local basibionts), time of deployment (TI, T1 = one month, and T3 = three months) and each locality (LO, L1 = Sancti Petri, L2 = Rota and L3 = Puerto América). MS = mean square; df = degrees of freedom; P(MC) = Monte Carlo p values.
| Source of Variation | df | MS | Pseudo-F | P(perm) | P(MC) |
|---|---|---|---|---|---|
| Abundance (Non-Transformed) | |||||
| Substrata = SU | 1 | 11,273 | 1.3592 | 0.2146 | 0.1639 |
| Time = TI | 1 | 17,597 | 1.8889 | 0.0488 | 0.0225 * |
| Locality = LO | 2 | 18,252 | 4.5744 | 0.0001 | 0.0001 *** |
| SU × TI | 1 | 9403.5 | 1.5467 | 0.112 | 0.0856 |
| SU × LO | 2 | 8293.7 | 2.0786 | 0.0001 | 0.0003 *** |
| TI × LO | 2 | 9315.8 | 2.3347 | 0.0001 | 0.0001 *** |
| SU × TI × LO | 2 | 6079.5 | 1.5237 | 0.0069 | 0.0141 * |
| Residual | 96 | 3990.1 | |||
| Total | 107 | ||||
| Pair-Wise tests a | SU × TI × LO | ||||
| T1 × L1, L2: CO ≠ LB; T1 × L3: CO = LB. | |||||
| T3 × L1: CO ≠ LB; T3 × L2, L3: CO = LB. | |||||
* p < 0.05; *** p < 0.001. a Monte Carlo p values were used in pair-wise tests due to low number of unique permutations [36].
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Revanales, T.; Guerra-García, J.M.; Ros, M. Colonization Dynamics of Potential Stowaways Inhabiting Marinas: Lessons from Caprellid Crustaceans. Water 2022, 14, 2659. https://doi.org/10.3390/w14172659
AMA Style
Revanales T, Guerra-García JM, Ros M. Colonization Dynamics of Potential Stowaways Inhabiting Marinas: Lessons from Caprellid Crustaceans. Water. 2022; 14(17):2659. https://doi.org/10.3390/w14172659
Chicago/Turabian StyleRevanales, Triana, José M. Guerra-García, and Macarena Ros. 2022. "Colonization Dynamics of Potential Stowaways Inhabiting Marinas: Lessons from Caprellid Crustaceans" Water 14, no. 17: 2659. https://doi.org/10.3390/w14172659
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.