Recent Vertebrate and Invertebrate Burrows in Lowland and Mountain Fluvial Environments (SE Poland)


Institute of Nature Conservation, Polish Academy of Sciences, 31-120 Kraków, Poland
Water 2020, 12(12), 3413; https://doi.org/10.3390/w12123413
Submission received: 28 September 2020
/
Revised: 1 December 2020
/
Accepted: 2 December 2020
/
Published: 4 December 2020
(This article belongs to the Special Issue Ecosystem Functioning in Rivers and Riparian Zones)
Abstract
:Over geological time, fauna inhabiting alluvial environment developed numerous traces of life, called bioturbation structures. However, present literature rarely offers comprehensive and comparative analyses of recent bioturbation structures between different types of environments. In this paper, the distribution of recent animal life traces in two different fluvial environments (lowland and mountain watercourse) was examined. An analysis of a set of vertical cross-sections of river bank enabled to determine physical and environmental features of the burrows, as well as degree of bioturbation in individual sections. Most of the burrows were assigned to their tracemakers and compared between two studied reaches in relation to the geomorphic zones of a stream channel. A mesocosm was conducted using glass terraria filled with river bank sediment and specimens of Lumbricus terrestris. The experiment confirmed field observations on the ability of earthworms to migrate into deep sediment layers along plant roots. The impact of floods on fauna survival was assessed on the basis of observations of large floods in 2016, 2018 and 2019. The abundance, distribution and activity of fauna in the sediment are mainly controlled by the occurrence of high and low flows (droughts and floods), which was particularly visible in lowland river reach.
1. Introduction
Riverine environment is characterized by large hydrodynamic gradients in a relatively short time, forcing special adaptations in burrowing animals, which has been recorded in bioturbation structures [1]. Vertebrate and invertebrate predators produce mainly shelter burrows, and saprofags, especially earthworms, produce locomotion and feeding structures [2]. Their behavioral (ethological) classification by Seilacher [3] is domichnia and pascichnia, respectively. Organisms with a larger body size (mainly mammals) create more complex structures that perform many living functions [4]. Fauna commonly found in an alluvial environment plays an important role in the distribution of organic matter and trophic energy in fluvial ecosystems [5].
Quick development of continental invertebrate paleoichnology observed in the last two decades [2,5,6,7,8] is not fully accompanied by neoichnological investigations but see [9,10,11,12], mostly because geologists or paleontologists are not accustomed to working on such interdisciplinary topics. However, entomological literature contains numerous descriptions of recent and fossil traces produced by soil insects [13,14,15,16,17]. The existing literature offers analogies between recent and fossil tracemakers [18,19], but these analogies are mostly based on one type of environment (a specific outcrop, place, river, or rock layer). Experience shows, however, that the same organisms living in slightly different environments produce different traces of life activity. Recent bioturbation structures in fine-grained alluvial sediments of Polish rivers have so far been studied by Mikuś and Uchman [20], but only in terms of ground beetles (Coleoptera, Carabidae) and their contribution to Macanopsis ichnospecies. Many experimental research studies, aimed at understanding biology, ecology and ethology of organisms inhabiting a variety of environments [18], are becoming key to understanding analogical traces known from the fossil record. However, there is still a need to compare recent bioturbation structures formed by the same groups of organisms among different environments.
This paper aims to explore and compare the quantity and distribution of recent bioturbation structures formed in coeval lowland and mountain fluvial environment and identify the main groups of their tracemakers. These results may make an important contribution to analogous research conducted in other contemporary environments as well as be helpful in describing and interpreting similar-looking structures known from the fossil record. The detailed study tasks were as follows: (i) identification and comparison of bioturbation structures in the sediments of the mountain and lowland river; (ii) determination of the rate of bioturbation of recently deposited fine-grained alluvial sediments in a controlled laboratory setting; (iii) examining the style and rate of fauna migration into recent overbank sediments with the comparison of field and laboratory conditions; (iv) visual observation of general reaction of river bank fauna on water-level fluctuations during floods.
2. Study Area
The studies were conducted on two Polish watercourses:
The Dunajec River originates from the conjunction of the Czarny Dunajec and the Biały Dunajec rivers at 610 m a.s.l. and is a typical Carpathian gravel-bed river with a mean annual discharge in the study area of 93 m3·s−1 and a maximum flood discharge of 4500 m3·s−1. Its total length is 247 km, and the catchment area is 6804 km2. Most of the river load constitutes fine sediments (<1 mm) transported in floatation. Gravel (2–80 mm in diameter) is transported and deposited on gravel bars during medium or large floods. Mean channel slope in the studied reach is 0.06%. Channelization works carried out within the Sandomierz Basin in the 1950s–1970s considerably narrowed the river and transformed its multi-thread channel into a single-thread, straight channel [21]. This resulted in rapid channel incision reaching up to 8 m in the study area. In its lower course, the river flows through the Sandomierz Basin in the valley composed of a set of recent alluvial terraces. The last ice-cover period took place between 730,000–430,000 BP and was succeeded by a long-term period of sedimentation and erosion [22]. Holocene alluvial terraces began to develop about 8000 BP on partially eroded Pleistocene fluvioglacial terraces formed about 40,000 BP [23]. Holocene alluvial terrace, locally called “redzina terrace” occurs from 4 to 8 m above the mean water table, incises into Pleistocene terrace [21] and is surrounded by a set of (mostly dry) oxbow lakes. This terrace is mainly composed of sand, gravel, silt and mud deposited in channels, bars, oxbow lakes and floodplain settings [22,23]. Its surface is well-drained and is rarely inundated during high-magnitude floods originating from heavy, long-lasting rainfall or rapid snow melt occurring in late spring–late autumn. The highest flood peaks registered at Żabno gauging station reach a height of up to 11 m above channel bottom [22]. The recent river channel is deeply incised into the Pleistocene terrace and bounded either by eroded scarps, or by a low floodplain with extensive gravel bars. Bioturbation structures, which are the focus of this study, occur in non-vegetated or poorly vegetated, fine- to medium-grained sand (grain size from 0.05 to 0.6 mm) or muddy sediments (grain size from 0.003 to 0.05 mm). The riparian vegetation, which grows mostly on recent alluvial soils, is dominated by sedge meadows and riparian forests, the latter being composed primarily of willows (Salix sp.) and poplars (Populus sp.). The redzina terrace is partially anthropogenically modified, mostly by agriculture. The area is located in a temperate suboceanic climate zone, with average annual precipitation of 650 mm, average annual temperature of 8 °C, and average temperature of the coldest (January) and warmest month (July) being −3 °C and 19 °C, respectively.
The Wielki Rogoźnik Stream originates at 910 m a.s.l. from a conjunction of the Cichy Stream and Raczy Stream (Figure 1B). Flowing through Orawa-Nowy Targ Basin, it ends its course in the Czarny Dunajec River at an altitude of 601 m a.s.l. Its catchment area is 125 km2 and the channel slope is 0.43%, almost seven times steeper than in the Dunajec River. Mean annual discharge is 1.87 m3·s−1 and the maximum recorded discharge was 213 m3·s−1. The stream shows a typical for gravel-bed mountain streams riffle-and-pool pattern, as well as a tight channel and narrow V-shape valley with steep slopes [24]. The channel of Wielki Rogoźnik is also incised, but not as deep as the Dunajec River, with the height of the banks between 1.5 and 3 m above the mean water table. The stream is unconstrained, which enables free channel migration and the development of a system of active meanders and paleochannels. Stream energy is considerably high, which is reflected in the sizes of the transported sediment (most of the bedload consists of gravel with a grain size from 8 to 350 mm; median: 56 mm). This mountain environment is typically scarce in fine-grained sediments; however, quite thick (ca. 50 cm) layers of sandy (<2 mm and >0.1 mm) and muddy sediments (<0.05 mm and >0.002 mm) were deposited along the channel margins during each Holocene flood event. As Polish mountain rivers transport relatively tiny portions of fine sediments in relation to gravel, these thick sandy and muddy layers come from many flood events. These layers are often partially eroded during large floods, so their total thickness is relatively constant. Contrary to the Dunajec River, this area was not glaciated in the Pleistocene, but its valley floor is made of fluvioglacial sediments washed out during the melting of Tatra Mountains glaciers. These sediments form raised terraces of different ages, constituting natural elevations surrounding valley bottom. During the Holocene, brown soils were developed on these fluvioglacial and fluvial deposits, shaping recent lithography [25]. The riparian vegetation is dominated here by alder (Alnus glutinosa and A. incana), Alnetum incanae and willows (Salix sp.). Exposed gravel bars, covered with a thin layer of sandy and silty deposits, are dominated by Petasites albus and P. hybridus. Channel banks are mostly covered with meadows and grazed from May to September. Mean annual precipitation is 950 mm, average annual temperature is 4 °C and average temperature of the coldest (January) and warmest month (July) is −7 °C and 14 °C, respectively.
3. Materials and Methods
The research subjects were recent bioturbation structures formed in the whole Holocene fluvial sediments.
The definition of “bioturbation” mainly depends on the specific environment we are currently studying. In terms of this study, bioturbation is a process of sediment displacement (as a result of mounding, mixing and burial) caused by vertebrate and invertebrate fauna permanently or temporarily inhabiting this sediment. Its sediment record was reflected in the occurrence of straight, curved or U-shaped biogenic structures (in the case of single individuals) and oval, elongated, multi-structural forms (in the case of colonial organisms).
Both studied environments are characterized by the alternation of erosion and deposition [26], which means that the thickness of the Holocene sediments does not increase at present. As a proof, the first observations of the banks of these rivers were made in 2007, and since then, the thickness of fine-grained alluvia remains almost the same despite many episodes of erosion and deposition.
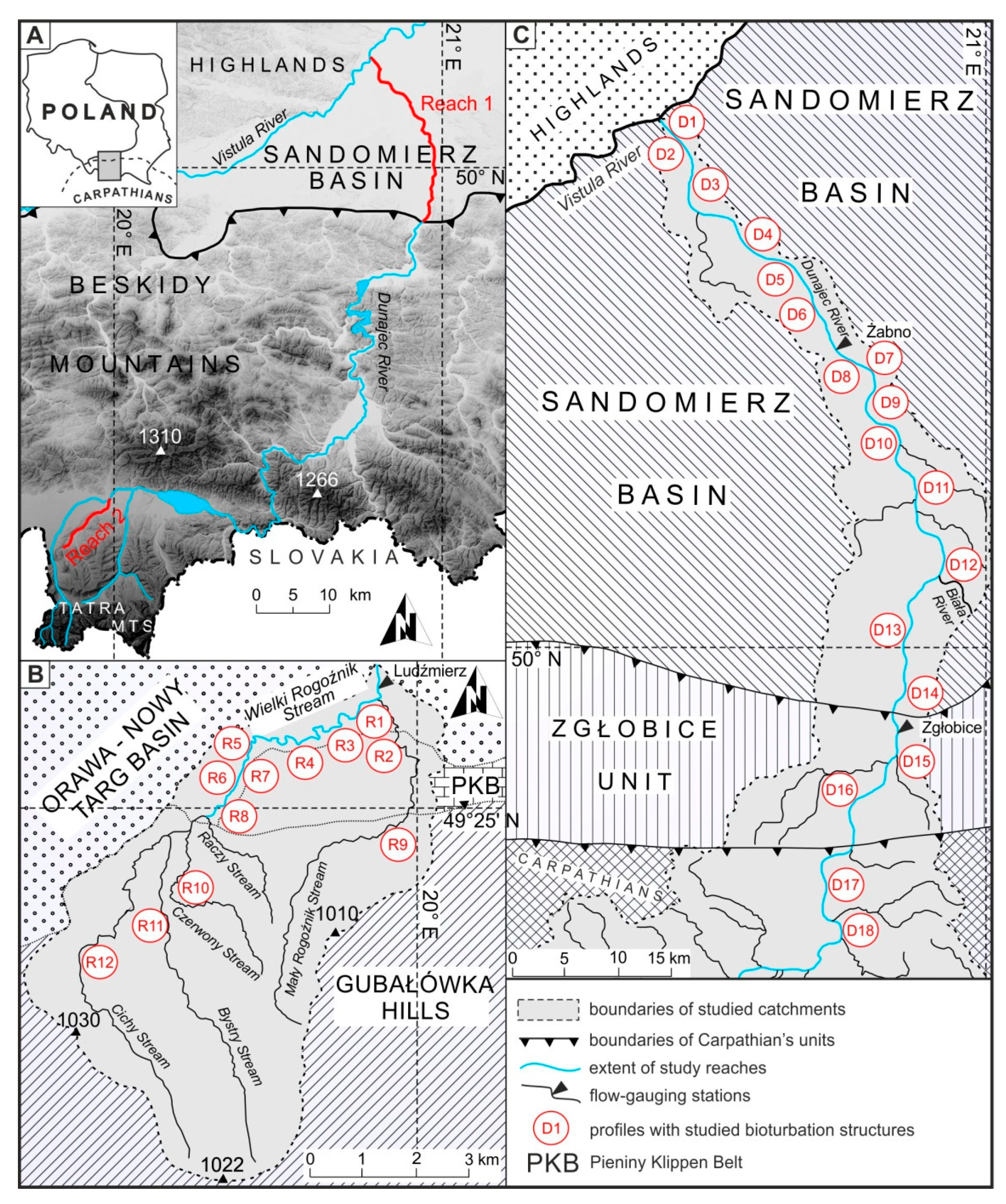
The most promising location for observations of bioturbation structures was a set of vertical sections made in steep, eroded channel banks. Eighteen profiles were made in the lower course of the Dunajec River (Figure 1C), while in the Rogoźnik Stream, 12 profiles were made (Figure 1B). Five cross-sections were located in the tributaries of the Rogoźnik Stream. The height of the studied profiles ranged from 0.99 to 4.7 m in the Dunajec River and from 0.88 to 1.75 m in Rogoźnik Stream, and the width of the field of observation of each profile ranged from 0.8 to 2.0 m. Each of the profiles started in gravel in-channel sediments, reaching water level. The upper parts represented different types of sediments, typical for natural levees and crevasses. All vertical profiles were made by using a shovel to prepare a smooth, vertical wall. Using a centimeter scale, detailed photos (Figure 2) were taken, and each profile was described bed-by-bed.
Without specific methods of dating, it is not possible to accurately determine the year of establishment of a given bioturbation structure, uninhabited at the moment of observation. Moreover, in some profiles, numerous thin layers of each depositional episodes occur, while in others, thick layers of fluvioglacial fan superimposed by one (up to 1.5 m thick) episode of fine-grained sedimentation. Luckily, most of these thick layers contained various anthropogenic waste (plastic, bottles, ash, debris) that gave information about its age with an accuracy of about 10 years.
The studies of recently deposited river bank sediments were supplemented with observations of oxbow lakes and paleochannels that are different type of environment, where burrows are not often flooded. For this purpose, seven vertical sections (three in reach 1 and four in reach 2) were made in these older, but still Holocene sediments. The size, location and density of burrows in a given profile were described, and most of the recent tracemakers were specified.
Observations of vertical profiles were conducted from April to September in the years 2016–2018. The characteristics of each studied profile included distinguishing sedimentary facies and visual definition of the degree of bioturbation within each layer. The burrows were either open or filled with sediment; the latter visually and structurally differed from the surrounding sediment due to their different color. As open burrows were better visible immediately after preparation of a given profile, and those filled with fine sediment became visible after 15–30 min, amendments to the initial descriptions were often made and photographs were repeated.
Quantification of bioturbation degree in vertical profiles was measured by applying categories with a unique linear distribution, adapting a visual method similar to that used by Knaust [27].
The degree of bioturbation was assessed by using four-step scale: 100–70%, 69–40%, 39–10% and 9–1% of bioturbation and no bioturbation structures. These values show the percentage share of bioturbation in the total area of a given layer, visible on a flat surface of vertical section. Burrows of mammals were not studied inside and their contribution to the total percentage of bioturbation was calculated using only the area of their entrance to the burrow. Burrows made by flying insects (Hymenoptera) and ground beetles (Carabidae) were numerous mainly in sections D2, D3, D5, D6 and D8. Therefore, they were not only included in the degree of bioturbation, but also their density in the number of burrows per square meter of the profile surface was assessed. Mole cricket (G. gryllotalpa) produced a system of branched corridors running along one layer towards the water surface. Its burrow system was firstly excavated with a spatula and subsequently measured using a tape.
The burrows were “paired” with their tracemakers firstly by searching for live individuals inside them, and secondly by visually comparing the shape and size of a given structure with its literature descriptions (mainly those made by Genise [28]). Mean degree of bioturbation was calculated separately for bioturbated layers, and within the entire studied profile. The first value was expressed as the sum of the mathematical products of the layers surface area and their visually determined degrees of bioturbation for each bioturbated layer. The second value was calculated as the product of the surface areas and the degrees of bioturbation of each layer in a given cross-section.
Another problem was the distinction between visually similar burrows of large earthworms and smaller beetles. In most cases, burrows of earthworms were oriented parallel to the course of the layers and their walls were smooth, slippery, often enclosed with feces, or filled with them. On the contrary, beetle burrows were mostly oriented almost perpendicular to the course of the layers and formed mainly in sediments that were easy to process—fine-to-coarse grained sand. In the case of burrows located along the border of two different layers, earthworm corridors ran in silty layer, while beetle burrows ran in sandy layer, with a silt layer forming a ceiling.
Two major floods occurred during field studies in June 2016 and August 2018. Floods can affect bioturbation structures either by erosion (common for mountain river environment) or by a temporary flooding of the burrow without destroying it and by filling the burrow with fine sediment from falling flood wave. In the case of R. riparia, their burrows are often partially eroded, but are still inhabited. After the passage of large floods, observations on the rate of fauna recolonization were made during one month, at three-day intervals in the vicinity of the 10 profiles most abundant in bioturbation structures.
The studies of recently-deposited river bank sediments have been supplemented with observations of oxbow lakes and paleochannels that are a different type of environment, where burrows are not often flooded. For this purpose, several ditches were made in these older (but still Holocene) sediments. The size, location and density of burrows in a given profile were described, and most of the recent tracemakers were specified. Plant roots were not included in these figures, but were often observed as earthworm migration routes, and thus, contributed to the increased bioturbation degree inside the deeper layers.
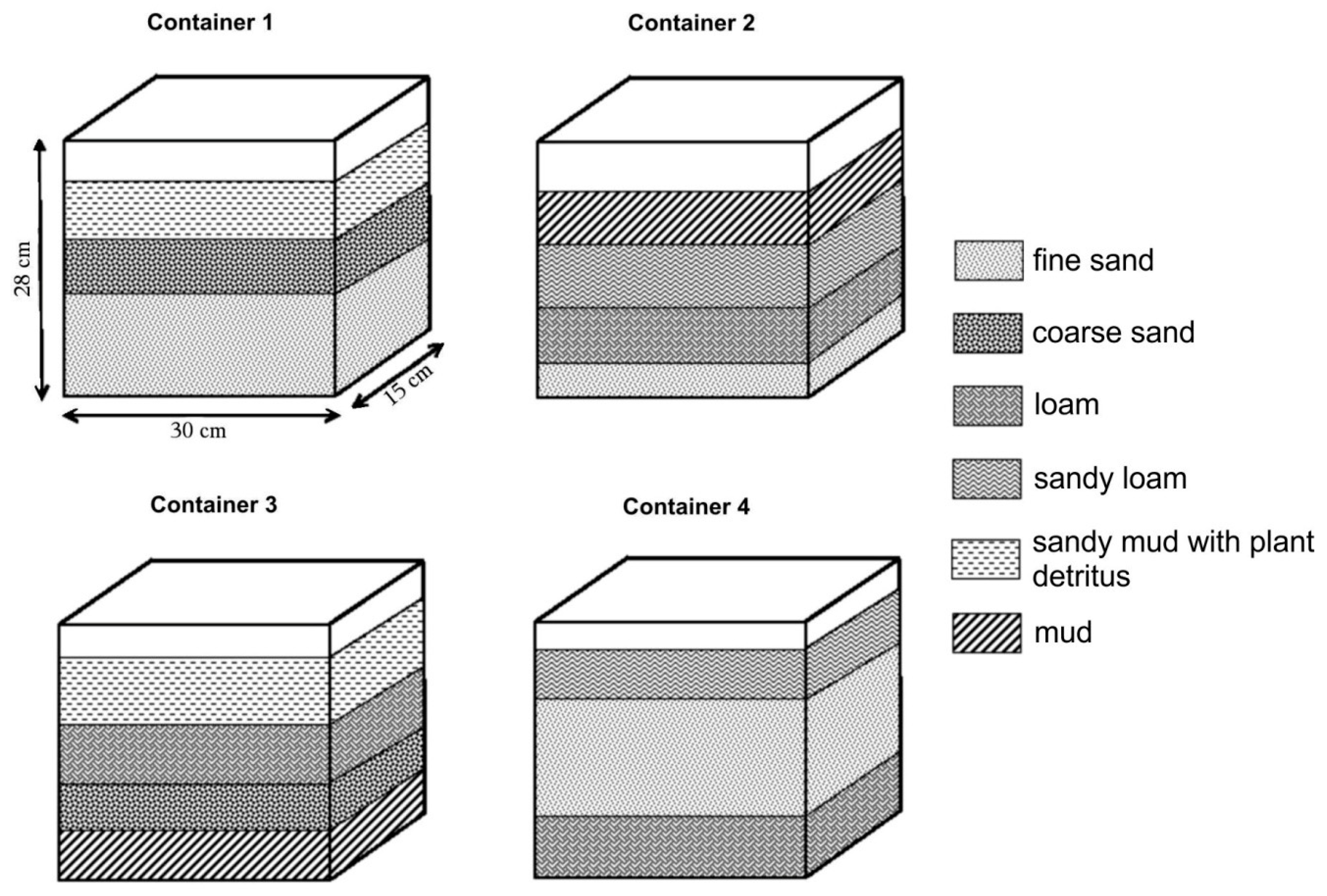
A mesocosm was performed in order to explain how earthworms migrate into deeply fertile sediment layers, separated from the top of the profile by a layer of coarser, sharp-edged sediments where the earthworms do not feed. In the spring of 2015, four equal-sized glass containers were filled with fine-grained overbank sediments collected from the Dunajec River in four different arrangements, as shown in Figure 3. The total follow-up period was 20 months. Different substrates used in containers were reflected in different grain size of the sediment. Mud is defined as a mixture of silt and clay (with grain size of 0.01 to 0.1 mm) with the addition of organic substances. Fine sand was defined as a rounded grain, sized >0.63 and <1 mm. Coarse sand was defined as sharp-edged sand with a grain size >1 mm and <2 mm. Loam was defined as a type of recent soil, developed on silt-sandy alluvia. Ten specimens of L. terrestris collected from natural river sediment were placed in each container. Earthworms were of different sizes, which made it possible to observe burrows of different diameters. The experiment lasted 20 months, and ongoing observations were limited to weekly visual inspection and photography of container walls. During the first year, earthworms were not fed, but the containers were supplied with various quantities of water (from 0.2 L per container per month to 1 L per container per week). In the second year, dead tree roots with a diameter of between 7 and 9 mm were delivered to the containers in order to observe if the roots can be used by a kind of “bridges,” enabling the entry of earthworms to lower fertile layers, separated from the higher ones by at least one layer of coarse, sharp-edged sand. The roots extended from the top to the bottom of sediment and were oriented either vertically or at an angle of approximately 45 degrees to the bottom of containers. One wall of all containers was exposed to direct sunlight every day. As this was the only surface where photosynthesis took place, algae and grass developed. After 9 month of observations, container 1 and 4 was subjected to negative temperatures: −1 °C for 8 h and on the next day −3 °C for 8 h in order to check the resistance of earthworms to freezing in weakly hydrated sediment.
The density of bioturbation structures within various geomorphological units of the studied watercourses was also compared (floodplain, natural levee, channel bar, estuary). This parameter was evaluated using the same methodology as for profile studies (% of the layer surface covered by bioturbation or a number of entrances per square meter).
4. Results
4.1. Description of the Studied Burrows
4.1.1. Earthworms
Earthworms produced rounded-shape burrows with a diameter of 7–11 mm which are open for several months. Older burrows were either cast or filled with very fine sediment mechanically rinsed by water during floods. The burrows occurred only in very fine sandy, silty or loamy substrate and often formed a branched network oriented parallel to the layer course and extended across its entire width. In older Holocene deposits, earthworm bioturbations are visible as a system of horizontal, thin, dark burrows, sharply demarcated from the surrounding sediment. A characteristic feature common for both of the studied reaches was their coexistence with burrows of small mammals in sediments of natural levees.
4.1.2. Small Mammals
The second largest group of bioturbation structures is formed by small mammals, represented in the study area by the common mole (Talpa europaea) and the bank vole (Myodes glareolus). The mole burrows are a complex net of horizontal and vertical corridors, reaching the water level through the layers of sand, silt and fine gravel of any thickness. In the vertical section of the study area, the burrows were round or oval, with a diameter of 40–70 mm. Smaller, round holes (about 30 mm in diameter) in the banks marked bank vole burrows. Their burrows were located just beneath the surface grass roots, between 8 and 25 cm below the top of vertical cross-sections. Three significant features of the bank vole burrows differentiating them from moles were noticed: (1) horizontal orientation; (2) the presence of opening of hazelnuts close to the entrance of the burrow; and (3) the lack of molehills in the proximity of the burrows.
4.1.3. Ground Beetles
Burrows produced by ground beetles were present almost exclusively either on sandy or muddy substrata. Their burrows show a characteristic morphology, with a straight or curved shaft and a horizontal to oblique terminal chamber. In the studied vertical profiles, the burrows were approximately perpendicular to the plane of the scarp, although rare horizontal orientations were also observed. The shaft diameter may vary slightly, with some burrows exhibiting a gentle tapering or widening upward [20]. The shaft ranged in length from 45 to 98 mm in and 6 to 11 mm in diameter.
4.1.4. Hymenoptera
The burrows of the mole cricket (Gryllotalpa gryllotalpa) occurred on the flatter river banks in silty and muddy substrate reaching the water table. The burrows were 18–28 mm in diameter, of rounded or slightly elongated shape and had a characteristic “herringbone” texture on the underside, formed during its digging. In spring, male mole produced a special type of the burrow entrance: a singing burrow shaped as a double exponential horn with bulb. Flying insects represented by wasps and bees produce relatively short burrows (a few cm long) consisting of a main tunnel (vertical or inclined) that ended with a cell for breeding and food provision [27]. In silty sediments near the outlet of both studied reaches, these burrows were numerous, appearing up to 200 per square meter. Ant colonies were present in vertical sections made in the vicinity of forest or meadow areas. Their burrows were characterized by irregular shape, sharp edges and disc-shaped chambers connected to each other by very thin corridors. The diameter of these structures was between 10 and 32 cm.
4.1.5. Large Mammals
Large mammals are the only group of organisms capable of digging burrows in coarse-grained sediments (coarse sand, fluvioglacial fan and point bar deposits) in the study area. The burrows of red fox (Vulpes vulpes) were oval-shaped and their diameter was between 32–45 cm. Their orientation was generally downwards (at angle 40°). Several broadens occurred inside the burrow, with numerous side tunnel branches. Their depth ranged from 0.5–2.5 m and extended nearly to the water level [29]. The burrows of common otter (Lutra lutra) have a similar characteristic, size and shape, but their main entrance is mainly under water table. It then slopes upward to a nest chamber situated above the high-water level [30]. The Eurasian beaver (Castor fiber) produces burrows similar in size and shape to the previous species, but occurring throughout the profile height, which are generally inclined at an angle of between 50 and 80°. Moreover, its presence in the sediment is often paired with migration paths towards the water cut in the uppermost muddy layer. The burrows of large mammals occurred in the lower part of the cross sections and always remained open. The flood led either to lateral erosion of the whole burrow or collapse of the ceiling and destruction of the burrow.
4.1.6. Birds
This group was represented by bank swallow (Riparia riparia), typically nesting in small to large colonies in soft sediments of eroding banks along rivers and streams [31]. Bank swallow occupied uppermost layers in the studied profiles, built of silt to fine sand (mean grain size from 0.002 to 0.05 mm). The burrows had a rounded shape, 32–51 cm in diameter, 60–145 cm long and finished with a chamber. Scratch marks were often visible on the inner walls.
4.2. Distribution of Bioturbation Structures in Studied Watercourses
Overall characteristic of each studied vertical profile was illustrated in Table 1. The analysis of vertical profiles was made in the slopes of alluvial banks that allowed a clear separation of these two types of environments:
4.2.1. Mountain River Environment
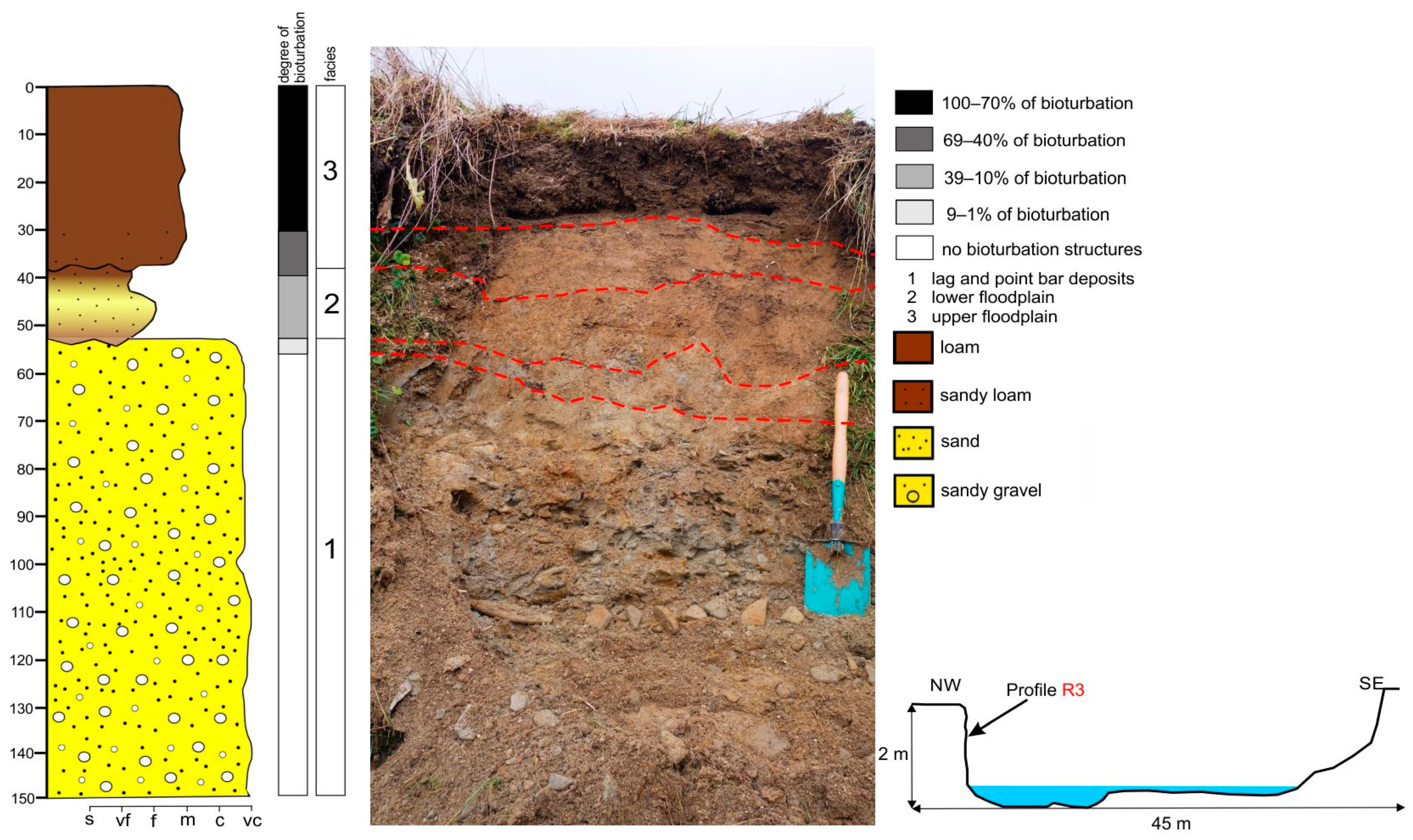
Sediments in the Wielki Rogoźnik Stream reflect a substantially higher physical energetic environment (steeper channel slope and higher flow energy) controlled by an increased lateral erosion of the banks, rather than by large fluctuations of the water level. Typical for this environment, individuals with the largest body size, represented by mammals (besides beavers), most often inhabited the top part of the banks or paleochannels and lagoon terrace. This is reflected in short-term but very rapid changes in sediment moisture in the lowermost layers. Vertical silty slopes falling towards the water table were inhabited in its upper part by insects whose burrows occurred in numbers between 5 and 15 per square meter of the profile area. A characteristic feature of the mountain river sediments was the large fluctuations in the mean bioturbation degree, from less than 10% in the case of uppermost profiles dominated by gravel layers separated with a thin layers of sand (profiles R12–R9), to nearly 24% in the lowermost profile R1. Paleochannels, which were definitely ‘hot-spots’ of fauna concentration (Figure 4 and Figure 5B), showed a much higher average degree of bioturbation, up to 36% (Table 1). The zone of open burrows reaches 60–100 cm below the ground surface. Here, the highest concentration of fauna occurred not in the uppermost part of the studied profiles, but in the fertile mud layer located across the sediment profile. The most complex bioturbation structures occurred in profiles R1–R8, where near 50-cm thick layer of fine sediments showed almost complete bioturbation in its uppermost part (Figure 4). Small mammals and earthworms were responsible for over 90% of the bioturbation structures in this reach. The relatively poor species composition on the stream banks is affected by the relative scarcity of bioturbation structures created by insects and birds. However, in the last five years, beavers have rapidly spread across the Carpathian rivers. During this study period, the number of beaver burrows in the river bank almost doubled during each year of observation.
4.2.2. Lowland River Environment
As the Dunajec River is deeply incised in its lower reach (up to 5 m)—vertical profiles D1–D4, D6 and D8–10 were substantially higher than others (Figure 5A and Figure 6). Moreover, a complex mosaic of layers in these profiles enabled to observe the occurrence of many layers with different level of bioturbation, sometimes separated by coarse-grained, unbioturbed layers. The zone of open burrows reached from 60 to 150 cm below the top of the profiles, reflecting the water level during the last flood event. Ground beetles (Coleoptera: Carabidae) were very widespread in fine-grained layers of profiles D1–D4 and D8–D10.
Both in-channel and overbank deposits in the lower course of the Dunajec River are characterized by better sediment sorting and median grain size, clearly decreasing towards the mouth. The mean degree of sediment bioturbation amounted to an average of 35% in profiles located in the uppermost part of the studied watercourse (profiles D16–D18) to 69% in the lowermost profiles. This reflects a decreasing flow energy associated with less frequent flooding and less lateral erosion of the banks. Characteristic features for this type of sediment were the presence of the burrows of the European mole (T. europaea) filled with casts of the common earthworm (L. terrestris) reaching water level in the low-flow channel in profiles D3–D14.
The uppermost layers of fertile fine-grained sediments exhibited almost complete (near 100%) bioturbation in all studied sections. However, burrows of large mammals (common fox Vulpes vulpes, European beaver (Castor fiber) and the Eurasian otter (L. lutra)) were cumulated in the lower parts of the profiles, along the border of fluvioglacial fan and natural levee facies. Steep, actively eroded scarps in profiles D2–D4 and D8–D10 built of silty to fine-sand sediments were typical nesting sites for bank swallow R. riparia, which appeared in colonies of 15–145 nests per 100 m2 of the streamside bank. Freshly deposited muddy layers, gently sloping towards the channel in profiles D6 and D12–D14, were surface-penetrated by mole cricket (G. gryllotalpa) with a highly branched corridor network with a total length of 3 to 20 m.
4.2.3. Similarities and Differences in Both Studied Reaches
In a mountain river, fertile layers are often thin (can be thinner than the length of some of adult tracemakers), which is reflected in the predominant share of horizontally oriented bioturbations in relation to the course of the layers. On the contrary, a lowland river is dominated by vertical arrangement of burrows, often having two or three corridors running to the surface, enabling individuals to adapt their location to the rapid change of water level in the river. The decreasing degree of bioturbation upstream of each of the studied rivers proves not only an increase in the stream energy, but also a significant decrease in the amount and thickness of fertile fine-grained sediment layers, as well as a smaller number of frost-free days upstream.
Beetle burrows are more or less evenly distributed in sandy-silt sediments of natural levee and crevasse splays. It contrasts with the situation in the mountain river, where almost the entire population of beetles is concentrated in gravelly ground on strongly inclined surfaces near the water surface. The density of beetle burrows ranged from 5 per square meter in the banks of Wielki Rogoźnik Stream to over 100 per square meter in the estuary of the Dunajec. The presence of nesting insects was an indicator of a sediment not flooded for at least 2 months. Moreover, their large colonies (mainly those of flying insects) were an indicator of permanently emerged sediments, being the evidence of the cutting of rivers.
Thick packages of fine-grained sand layers were low to medium bioturbated in both studied reaches, and the observed increase in the number of burrows during one season was up to 20%.
Muddy layers located at the bottom of profiles (up to 450 cm below the top) were fully penetrated by both living and dead roots of trees and shrubs, widely branching along muddy layers. Several living earthworms were noticed in these layers separated by coarse sediments.
The presence of dense, woody and shrub vegetation on the river bank played an important role in the volume of bioturbation structures, but strong inter-species competition forced by the high variability of physical habitat conditions caused disappearance of some groups (often flying insects) and the dominance of two-three species (mainly Carabidae). This was a common feature of both studied watercourses.
4.3. Burrowing Activity of L. Terrestris During Mesocosm
Physical conditions in the containers varied significantly, which strongly reflected the activity of earthworms. The largest increase in the number of bioturbation structures occurred in the first 30 days of the experiment and after supplying water or plant roots to the containers. Generally, the degree of bioturbation in fertile layers increased linearly between months 1 and 12 of observation. After 5 months, the layer boundaries became very uneven and blurred, especially on the border of very fertile and less fertile sediments. In the second year of observation, a decrease in earthworm activity was observed (practically no new burrows appeared) due to a shortage of nutrients in the containers.
In container 1 and 3, the layers that were easy to penetrate and rich in nutrients were placed at the top and at the bottom and were separated by layers of sharp-edged, coarse sand. During the first 12 months, earthworms did not enter these layers. When dead roots were delivered, the earthworms passed through a layer of coarse sand and extensive penetrations were observed in the highest and lowest layers, while the middle, coarse-grained layer was bioturbed only vertically and used as a “fast-track” between the uppermost and lowermost fertile layers. The supply of dead roots resumed the formation of new burrows, but only within the fertile layers.
One common process was noticed: filling earlier penetrations in less fertile sediment with excrements originating from lower or higher located layer. Additionally, mechanical rinsing and filling of burrows with overlying fine sediment took place in the lowermost layers, while each container was subjected to delivery of a large amount of water in a short time, simulating a rapid flood. However, the two types of burrow fillings can be distinguished both in the containers and during fieldworks by basic sedimentological analysis.
Humidity was a crucial factor controlling earthworm activity and survivability [32]. With humidity decreasing in the soil for a long time, the number of earthworms and their activity significantly decreased or many of them hibernated. In the long-lasting absence of food, this state can last up to several months. After introducing a large dose of organic matter into the containers in the form of roots, the restoration of earthworm activity and an increase in their population proceeded very quickly—in two or three days. Another important factor regulating earthworm population was soil temperature. Exposing any container to even light frost (−3 °C for 8 h) caused death of the entire population living in it.
4.4. Effects of Water-Level Fluctuations on River Bank Fauna
Three-year observations of changes in the Dunajec and Rogoźnik stream ecosystem indicate that a large part of the fauna inhabiting the alluvial environment has not developed adaptations to survive big floods. The dominant strategy of river-bank fauna was quick recolonization. The exceptions were mammals, which are able to quickly escape from a flooded area (although at the cost of destroying their burrows), as well as bank swallows (R. riparia), whose burrows have a considerable length and can be habitable despite the progressive erosion of the river bank.
After a large flood, the river bank was often completely rebuilt, which made it impossible to recover the population (after the flood from June 2016, Carabidae did not recover their population to the end of the year). On the other hand, flying insects quickly returned and were pioneer tracemakers, building a residential burrow shortly after the fresh sediment had dried. However, after another large and long-lasting flood from August 2018, even flying insects reconstructed only 30% of their burrows to the end of frost-free period.
The colonization time of freshly deposited sediments by insects was much higher in the mountain river and averaged 10 days, while in the lower course of the Dunajec River this averaged only 2 days. The sediment was subsequently settled by other insects (15 days in reach A and 5 in reach B); earthworms (21 days in reach A and 14 in reach B), small mammals (30 days in reach A and 20 in reach B) and large mammals and birds.
5. Discussion and Conclusions
Studying the activity of modern organisms allows paleontologists and sedimentary geologists to find answers to questions regarding ancient life from the behavioral and environmental side. In the sedimentary record, burrows are a major group of trace fossils, but only about 3% of the named burrows are known to be produced by vertebrates [33]. However, a strict indication of fossil both invertebrate and vertebrate burrows is often not possible because similar looking burrows may be produced by both groups. Therefore, neoichnological investigations are crucial for the interpretation of such traces. Despite enormous advances in biological and environmental sciences, details of animal burrows and their differences between different environments are still poorly understood [18]. This study improves this knowledge, showing the differences in the communities of organisms and providing the formation about their burrows in the environment of the mountain and lowland rivers of temperate regions.
The presented results provide a set of information allowing to infer about physical conditions (channel type, grain size, intense of flooding) in recently-deposited fine-grained fluvial sediment on the basis of basic sedimentological analysis coupled with a detailed description of bioturbation structures. Assessing more detailed features can also be reachable by coupling this sequence of analysis with some specific chemical properties of the sediment as well as a biological and ecological analysis of individual taxa. The decreasing degree of bioturbation upstream of each of the studied rivers proves not only an increase in the stream energy, but also a significant decrease in the amount and thickness of fertile fine-grained sediment layers, as well as a smaller number of frost-free days upstream.
The effect of tracemaker behavior on resultant burrows can also be investigated through observations during the production of individual structures. Apart from some advanced novel techniques of studying burrows presented here (i.e., three-dimensional tomography [34], X-ray radiography [35]), the mesocosm seems to be one of the most promising and accurate techniques, functioning as an eyewitness to the formation of bioturbation structures depending on variable environmental factors that can be freely controlled.
Actively eroded scarps along the river channel provide safe nesting sites for burrowing bird species, represented in the study area by R. riparia. These burrows can then, in turn, provide nest sites for species unable to create burrows themselves, especially in areas where the abundance of safe nesting holes is a limiting factor [36]. As a result, primary burrowing species create ecosystem services for secondary species, often modifying these primary burrows, which was also evidenced among large mammals with the example of otter colonization of abandoned beaver burrows [37].
Differences in the arrangement of burrows within the two studied reaches can be explained not only by different species composition, but also by the possibility of burrowing in two different substrates [38]. Thick sets of fine-grained layers in the lowland river show a very rich set of bioturbation structures and a greater proportion of vertical burrows. Thinner sediment layers in mountain river favors more mobile species, which do not build extensive systems of burrows in the proximity of the water table. Observations made by Rosenzweig [39] confirm this dependency. In the studied reaches, only mammals were able to relatively deeply process altering sand-mud-gravel sediment series.
According to the previous studies [40] the estimated number of earthworm burrows in alluvial soils of temperate region range from 100 to 800/m2, which should result in over 90% degree of bioturbation. Indeed, observations from vertical profiles confirm that fertile silty and muddy sediments are almost completely processed by earthworms. Other studies [41] reported that most burrows are located between 30 and 40 cm deep in the fine-grained surface sediment, but this can be applied only for the majority of the lowland reach with well-developed soils. Sediment maturity in the studied mountain stream was low to medium, and in most of the profiles, the mean thickness of the soil was between 10 and 20 cm—consequently, the highest density of bioturbation structures was found in this thin layer. Moreover, several profiles in the lower reach of the Dunajec River (D4, D5, D6, D8) had a thickness between 300 and 500 cm, and fertile mud layers were located at a depth of more than 300 cm. In some cases, these layers contained living earthworms, and their degree of bioturbation was comparable to the surface layers. Since earthworms are not able to migrate that deep through sharp-edged sediments, it had to do with the presence of dead roots breaking the entire set of layers. In addition, earthworm burrows provide a kind of “transport channels” facilitating the penetration of roots into the soil without losing much energy [42]. Observations from the two analyzed river reaches confirm previous findings [32,43] that water level is regarded as a main factor controlling vertical migrations of earthworms in alluvial sediments. Vertical migration of earthworms is also related to living roots, but limited to their growth range [44], which in the case of the studied rivers is rather limited to the uppermost layers in each profile.
Large floods are the most important life-disturbing factor in river and riverside ecosystems [45] and can be a cause of large, long-lasting changes in the number and diversity of fauna. Disturbances caused by floods significantly reduce the density and diversity of fauna in the alluvial environment on a regional scale [46]. Variations in body size and the position in the alluvial profile [47] determine the energy of flowing waters and the frequency of flooding by water. Generally, the closer the water table, the smaller the size of the individuals. The exceptions are mammals, namely, the European beaver (C. fiber), European otter (Lutra lutra) and European mole (T. europaea), whose dwelling burrows reach the water level (in the case of otters, the entrance to their corridors was often observed below the water surface). Most of the burrows of large mammals can survive floods practically untouched, which was observed in the study area after the passage of two major floods.
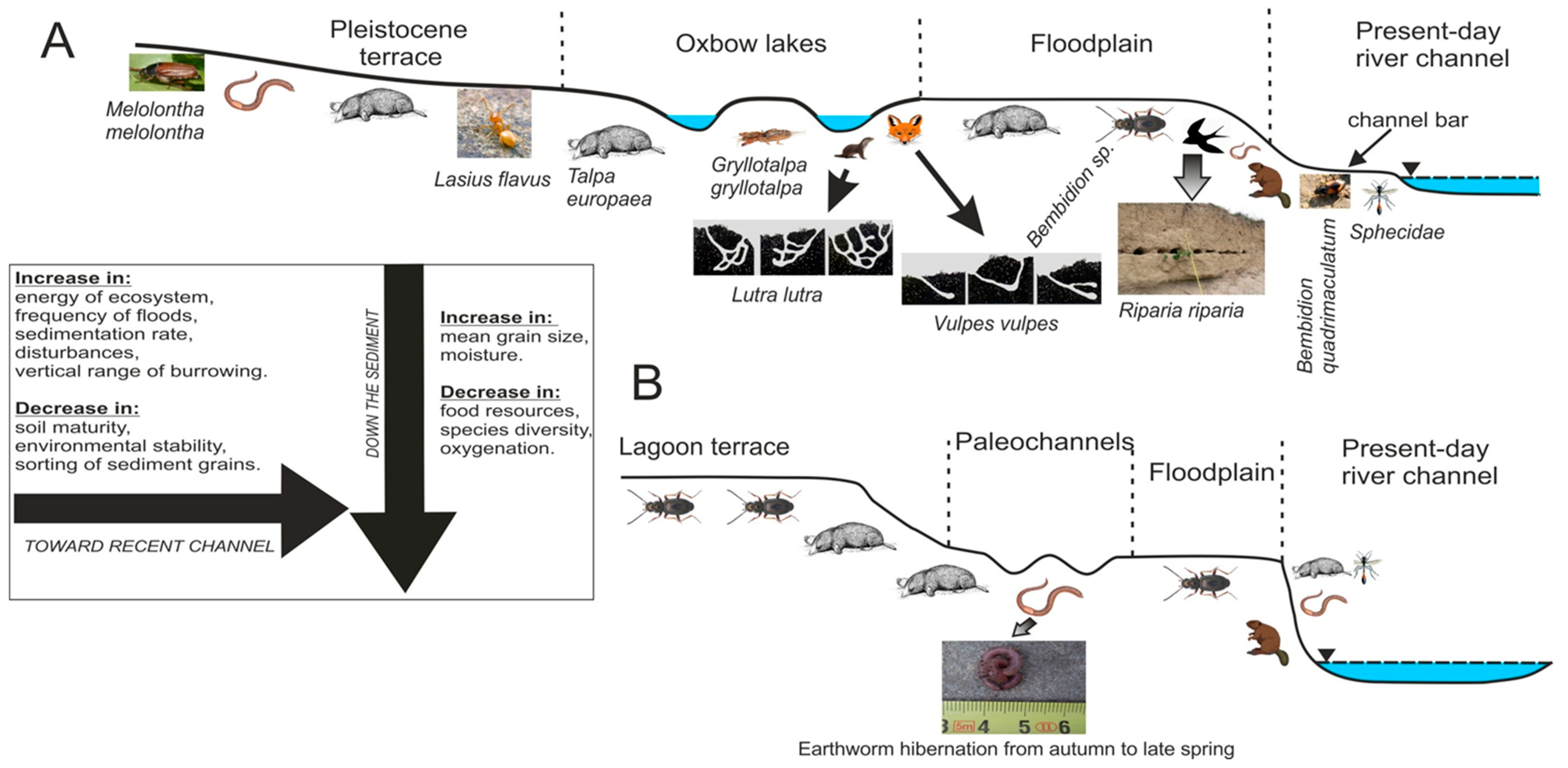
On gravel bars, the period of favorable conditions is limited and abruptly interrupted by floods, and smaller species dominate [48]. Higher in the profile, mainly on flood plains and in oxbow lakes, the period of availability of resources for organisms is longer and less frequently disturbed, which contributes to a greater concurrence of individuals of larger sizes (this applies in particular to Carabidae). The higher in the alluvial profile—the greater the interspecies competition, caused by the lower physical energy of the environment and greater possibilities of obtaining food [46], as well as the establishment of burrows out of the range of low to medium floods (Figure 5).
Studies comparing physical, biological and other aspects in different types of aquatic environments give an idea of their modern functioning. Their combination with the knowledge known from the fossil record is a valuable contribution to understanding the adaptations of the entire river ecosystem to the conditions of variable flow as well as sediment erosion and accumulation. Future directions focusing on the river environment should be scoped to more comprehensive studies using data from different altitude zones as well as seasonal rivers, freezing up of rivers and rivers that are highly affected by human impact. As bioturbation is the main factor of developing recent soils in freshly deposited alluvial sediments, human interventions to natural, wild river channels substantially disturb these processes by breaking the continuity of the river corridor and overuse of channel regulation, which often leads to the cutting of channel from permanent surfacing and drying of the flood plain, and a lack of freshly deposited alluvia. In some regulated reaches, no alluvial soils develop while the existing ones are subject to erosion and deterioration of quality [49]. The ideal situation for the development of alluvial soils is the presence of natural and semi-natural rivers that are “fenced off” from local buildings by some kind of “erodible corridor,” enabling the river to freely migrate in a given space. However, in some situations (e.g., drying up of rivers), the development of soils will be difficult, even in the vicinity of a completely natural channel.
Linking the degree and complexity of sediment bioturbation with geomorphological features of the channel and historical and current use of the floodplains is a useful tool for studying the continuity between given river zones and groups of organisms. Despite some limitations coming from the facts that the sediment layers are not laterally continuous over a large area and that most of the organisms studied in the vertical profile do not migrate over long distances, certain quantitative and qualitative patterns can be developed regarding the degree of bioturbation of the sediment and the tracemakers diversity.
Funding
The study was performed within the scope of the Research Project 2014/13/N/ST10/04902 financed by the National Science Centre.
Conflicts of Interest
The author declares no conflict of interest.
References
- Kristensen, E.; Penha-Lopes, G.; Delefoss, M.; Valdemarsen, T.; Quintana, C.O.; Banta, G.T. What is bioturbation? The need for a precise definition for fauna in aquatic sciences. Mar. Ecol. Prog. Ser. 2012, 446, 285–302. [Google Scholar] [CrossRef] [Green Version]
- Buatois, L.A.; Mángano, M.G.; Genise, J.F.; Taylor, T.N. The ichnologic record of the continental invertebrate invasion: Evolutionary trends in environmental expansion, ecospace utilization, and behavioral complexity. Palaios 1998, 13, 217–240. [Google Scholar] [CrossRef]
- Seilacher, A. Studien zur Palichnologie. Über die Methoden der Palichnologie. Neues Jahrb. Geol. Palaontol. Abh. 1953, 96, 421–452. [Google Scholar]
- Mermillod-Blondin, F.; Gerino, M.; Sauvage, S.; Creuzé Des Châtelliers, M. Influence of nontrophic interactions between benthic invertebrates on river sediment processes: A microcosm study. Can. J. Fish. Aquat. Sci. 2011, 61, 1817–1831. [Google Scholar] [CrossRef] [Green Version]
- Buatois, L.A.; Mángano, M.G. Animal-substrate interactions in freshwater environments: Applications of ichnology in facies and sequence stratigraphic analysis of fluvio-lacustrine successions. In The Application of Ichnology to Palaeoenvironmental and Stratigraphic Analysis; McIlroy, D., Ed.; Geological Society of London: London, UK, 2004; pp. 311–335. ISBN 978-1862391543. [Google Scholar]
- Hasiotis, S.T. Continental Trace Fossils: SEPM Short Course Notes; Sepm Society for Sedimentary: New York, NY, USA, 2011; ISBN 978-1565761247. [Google Scholar]
- Hasiotis, S.T. Continental ichnology: Fundamental processes and controls on trace fossil distribution. In Trace Fossils Concepts, Problems, Prospects; Miller, W., Ed.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 268–284. ISBN 978-1565761247. [Google Scholar]
- Genise, J.F.; Bellosi, E.S.; Gonzalez, M.G. An approach to the description and interpretation of ichnofabrics in palaeosols. In The Application of Ichnology to Palaeoenvironmental and Stratigraphic Analysis; McIlroy, D., Ed.; Geological Society of London: London, UK, 2004; pp. 355–383. ISBN 978-1429413220. [Google Scholar]
- Metz, R. Tunnels formed by mole cricket (Orthoptera: Gryllotalpidae): Paleoecological implications. Ichnos 1990, 1, 139–141. [Google Scholar] [CrossRef]
- Uchman, A. Treptichnus-like traces made by insect larvae (Diptera: Chironomidae, Tipulidae). In Pennsylvanian Footprints in the Black Warrior Basin of Alabama: Alabama Paleontological Society Monograph; Buta, R.J., Rindsberg, A.K., Kopaska-Merkel, D.C., Eds.; University of Alabama Printing Services: Birmingham, UK, 2005; pp. 143–146. ISBN 978-0976930402. [Google Scholar]
- Davis, R.B.; Minter, N.J.; Braddy, S.J. The neoichnology of terrestrial arthropods. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2007, 255, 284–307. [Google Scholar] [CrossRef]
- Hembree, D.I. Neoichnology of burrowing millipedes: Linking modern burrow morphology, organism behaviour, and sediment properties to interpret continental ichnofossils. Palaios 2009, 24, 425–439. [Google Scholar] [CrossRef]
- Rau, P. The biology and behaviour of mining bees, Antiotophora abrupta and Entechnia taurea. Psyche 1929, 36, 155–181. [Google Scholar] [CrossRef] [Green Version]
- Iwata, K. Evolution of Instinct. Comparative Ethology of Hymenoptera; Amerind Publishing Co: New Delhi, India, 1976; ISBN 978-0862494216. [Google Scholar]
- Halffter, G.; Edmonds, W.D. The Nesting Behavior of Dung Beetles: Publication 10; Instituto de Ecologia: Mexico City, México, 1982. [Google Scholar]
- Grasse, P. Termitologia, Tome II; Mason, Z., Ed.; Springer: Paris, France, 1984; ISBN 978-3540459095. [Google Scholar]
- Tschinkel, W.R. Subterranean ant nests: Trace fossils past and future? Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 192, 321–333. [Google Scholar] [CrossRef] [Green Version]
- Brandt, D.; McCoy, V. Modern Analogs for the Study of Eurypterid Paleobiology. In Experimental Approaches to Understanding Fossil Organisms. Topics in Geobiology; Hembree, D., Platt, B., Smith, J., Eds.; Springer: Dordrecht, The Netherlands, 2014; ISBN 978-9401787208. [Google Scholar]
- Genise, J. Ichnoentomology. Topics in Geobiology; Springer Nature: Cham, Switzerland, 2017; ISBN 978-3319282084. [Google Scholar]
- Mikuś, P.; Uchman, A. Beetle burrows with a terminal chamber: A contribution to the knowledge of the trace fossil Macanopsis in continental sediments. Palaios 2013, 28, 403–413. [Google Scholar] [CrossRef]
- Zawiejska, J.; Wyżga, B. Twentieth-century channel change on the Dunajec River, southern Poland: Patterns, causes and controls. Geomorphology 2010, 117, 234–246. [Google Scholar] [CrossRef]
- Robinson, D.T.; Tockner, K.; Ward, J.V. The fauna of dynamic riverine landscapes. Freshw. Biol. 2002, 47, 661–677. [Google Scholar] [CrossRef]
- Starkel, L. Evolution of the Vistula river valley during the last 15 000 years, part III. Geogr. Stud. 1990, 5, 11–184. [Google Scholar]
- Sokołowski, T. Uwagi o terasach doliny Dunajca koło Tarnowa (Remarks on the terraces in Dunajec valley near Tarnów (Southern Poland). Ann. Soc. Geol. Pol. 1981, 51, 579–594. [Google Scholar]
- Sokołowski, T. Development of the lover reach of the Dunajec river in the Vistulian and Holocene. In Evolution of the Vistula River Valley during the Last 15 000 Years. Geographical Studies, Special Issue; Starkel, L., Ed.; Continuo: Wrocław, Poland, 1995; ISBN 83-86682035. [Google Scholar]
- Zuchiewicz, W. Możliwość zastosowania teoretycznego profilu podłużnego rzeki w badaniach nad młodymi ruchami tektonicznymi. Ann. Soc. Geol. Pol. 1979, 49, 327–342. [Google Scholar]
- Knaust, D. Atlas of Trace Fossils in Well Core. Appearance, Taxonomy and Interpretation; Springer Nature: Cham, Switzerland, 2017; ISBN 978-3319498362. [Google Scholar]
- Genise, J.F.; Edwards, N. Ichnotaxonomy, origin and palaeoenvironment of Quarternary insect cells from Fuerteventura, canary Islands, Spain. J. Kansas Entomol. Soc. 2003, 76, 320–327. [Google Scholar]
- Statham, M.J.; Murdoch, J.; Janecka, J.; Aubry, K.B.; Edwards, C.J.; Soulsbury, C.D.; Berry, O.; Wang, Z.; Harrison, D.; Pearch, M.; et al. Range-wide multilocus phylogeography of the red fox reveals ancient continental divergence, minimal genomic exchange and distinct demographic histories. Mol. Ecol. 2014, 23, 4813–4830. [Google Scholar] [CrossRef]
- Noordhuis, R. The river otter in Clarke County (Georgia, USA): Survey, food habits, and environmental factors. IUCN Otter Spec. Group Bull. 2002, 19, 75–86. [Google Scholar]
- Garrison, B. Riparia riparia. Birds N. Am. 1999, 414, 1–20. [Google Scholar]
- Lavelle, P. Earthworm activities and the soil system. Biol. Fertil. Soils 1988, 6, 237–251. [Google Scholar] [CrossRef]
- Knaust, D. Trace fossil systematics. In Trace Fossils as Indicators of Sedimentary Environments. Developments in Sedimentology; Knaust, D., Bromley, R.G., Eds.; Elsevier: Amsterdam, The Netherlands, 2012; ISBN 978-0444538130. [Google Scholar]
- Hale, R.; Boardman, R.; Mavrogordato, M.N.; Sinclair, I.; Tolhurst, T.J.; Solan, M. High-resolution computed tomography reconstructions of invertebrate burrow systems. Sci. Data 2015, 2, 150052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gingras, M.K.; Zonneveld, J.P.; Konhauser, K.O. Using X-ray Radiography to Observe Fe Distributions in Bioturbated Sediment. In Experimental Approaches to Understanding Fossil Organisms. Topics in Geobiology; Hembree, D.I., Platt, B.F., Smith, J.J., Eds.; Springer: Dordrecht, The Netherlands, 2014; ISBN 978-9402403251. [Google Scholar]
- Rohrer, Z.; Rebollo, S.; Andivia, E.; Rodríguez, C.; Bird, J.F. Services applicable to mine restoration: A case study of Sand Martin (Riparia riparia) burrow construction. J. Ornithol. 2020, 161, 243–255. [Google Scholar] [CrossRef]
- Jenkins, D.; Burrows, G.O. Ecology of Otters in Northern Scotland. III. The Use of Faeces as Indicators of Otter (Lutra lutra) Density and Distribution. J. Anim. Ecol. 1980, 49, 755–774. [Google Scholar] [CrossRef]
- Dashtgard, S.D.; Gingras, M.K.; Pemberton, G. Grain-size controls on the occurrence of bioturbation. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008, 257, 224–243. [Google Scholar] [CrossRef]
- Rosenzweig, M.L. Species Diversity in Space and Time; Cambridge University Press: Cambridge, UK, 2010; ISBN 978-0511623387. [Google Scholar]
- Curry, J. Factors affecting the abundance of earthworms in soils. In Earthworm Ecology; Edwards, C.A., Ed.; CRC Press: New York, NY, USA, 2004; pp. 91–115. ISBN 978-0849318191. [Google Scholar]
- Kretzschmar, A. The burrow system of earthworms in grassland—Seasonal variations of field observations. Rev. Ecol. Biol. Sol. 1982, 19, 579–591. [Google Scholar]
- Le Bayon, R.; Bullinger-Weber, G.; Schomburg, A.; Turberg, P.; Schlaepfer, R.; Guenat, C. Earthworms as ecosystem engineers: A review. In Insects and Other Terrestrial Arthropods: Biology, Chemistry and Behavior; Horton, C.G., Ed.; Nova Science Publishers: London, UK, 2017; pp. 129–178. ISBN 978-1617614668. [Google Scholar]
- Jiménez, J.J.; Decaëns, T. Vertical distribution of earthworms in grassland soils of the Colombian Llanos. Biol. Fertil. Soils 2000, 32, 463–473. [Google Scholar] [CrossRef] [Green Version]
- Lacoste, M.; Ruiz, S.; Or, D. Listening to earthworms burrowing and roots growing—Acoustic signatures of soil biological activity. Sci. Rep. 2018, 8, 10236. [Google Scholar] [CrossRef] [PubMed]
- Hershey, A.E.; Lamberti, G.A. Stream macroinvertebrate communities. In River Ecology and Management; Naiman, R.J., Bilby, R.E., Kantor, S., Eds.; Springer: New York, NY, USA, 2001; pp. 169–192. ISBN 978-0387952468. [Google Scholar]
- Skalski, T.; Kędzior, R.; Radecki-Pawlik, A. Czynniki kształtujące strukturę zgrupowań biegaczowatych (Coleoptera: Carabidae) na terenach zalewowych rzek górskich. Infrastrukt. Ekol. Teren. Wiej. 2008, 3, 13–21. [Google Scholar]
- Paetzold, A.; Schubert, C.J.; Tockner, K. Aquatic-terrestrial linkages across a braided river: Riparian arthropods feeding on aquatic insects. Ecosystems 2005, 8, 748–759. [Google Scholar] [CrossRef]
- Van Looy, K.; Vanacker, S.; Jochems, H.; De Blust, G.; Dufrêne, M. Ground beetle habitat templets and riverbank integrity. River Res. Appl. 2005, 21, 1133–1146. [Google Scholar] [CrossRef]
- Rinaldi, M.; Wyżga, B.; Surian, N. Sediment mining in alluvial channels: Physical effects and management perspectives. River Res. Appl. 2005, 21, 805–828. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
(A) Location of the studied reaches in relation to physiogeographic regions of southern Poland. Drainage network of Wielki Rogoźnik Stream, (B) and of the lower reach of the Dunajec River (C) with detailed setting of studied profiles.
Figure 1.
(A) Location of the studied reaches in relation to physiogeographic regions of southern Poland. Drainage network of Wielki Rogoźnik Stream, (B) and of the lower reach of the Dunajec River (C) with detailed setting of studied profiles.

Figure 2.
Sample photos taken during detailed descriptions of vertical profiles in the studied river sections showing ant colony (A); burrow of Talpa europaea filled with earthworm castings (B); burrow of Gryllotalpa gryllotalpa and its branching structure (C,D); burrow with the terminal chamber of Carabidae (E); vertical profile of Pleistocene terrace superimposed by the oxbow lake of the Dunajec River (F).
Figure 2.
Sample photos taken during detailed descriptions of vertical profiles in the studied river sections showing ant colony (A); burrow of Talpa europaea filled with earthworm castings (B); burrow of Gryllotalpa gryllotalpa and its branching structure (C,D); burrow with the terminal chamber of Carabidae (E); vertical profile of Pleistocene terrace superimposed by the oxbow lake of the Dunajec River (F).

Figure 3.
Parameters of containers used for mesocosm.

Figure 4.
Characteristic of the vertical profile R3 in the Rogoźnik Stream—sediment features and degree of recent bioturbation.
Figure 4.
Characteristic of the vertical profile R3 in the Rogoźnik Stream—sediment features and degree of recent bioturbation.

Figure 5.
Distribution of the studied burrows with the most common geomorphic units of their occurrence and the main factors controlling burrowing activity within the studied reach of the Dunajec River (A) and the Wielki Rogoźnik Stream (B).
Figure 5.
Distribution of the studied burrows with the most common geomorphic units of their occurrence and the main factors controlling burrowing activity within the studied reach of the Dunajec River (A) and the Wielki Rogoźnik Stream (B).

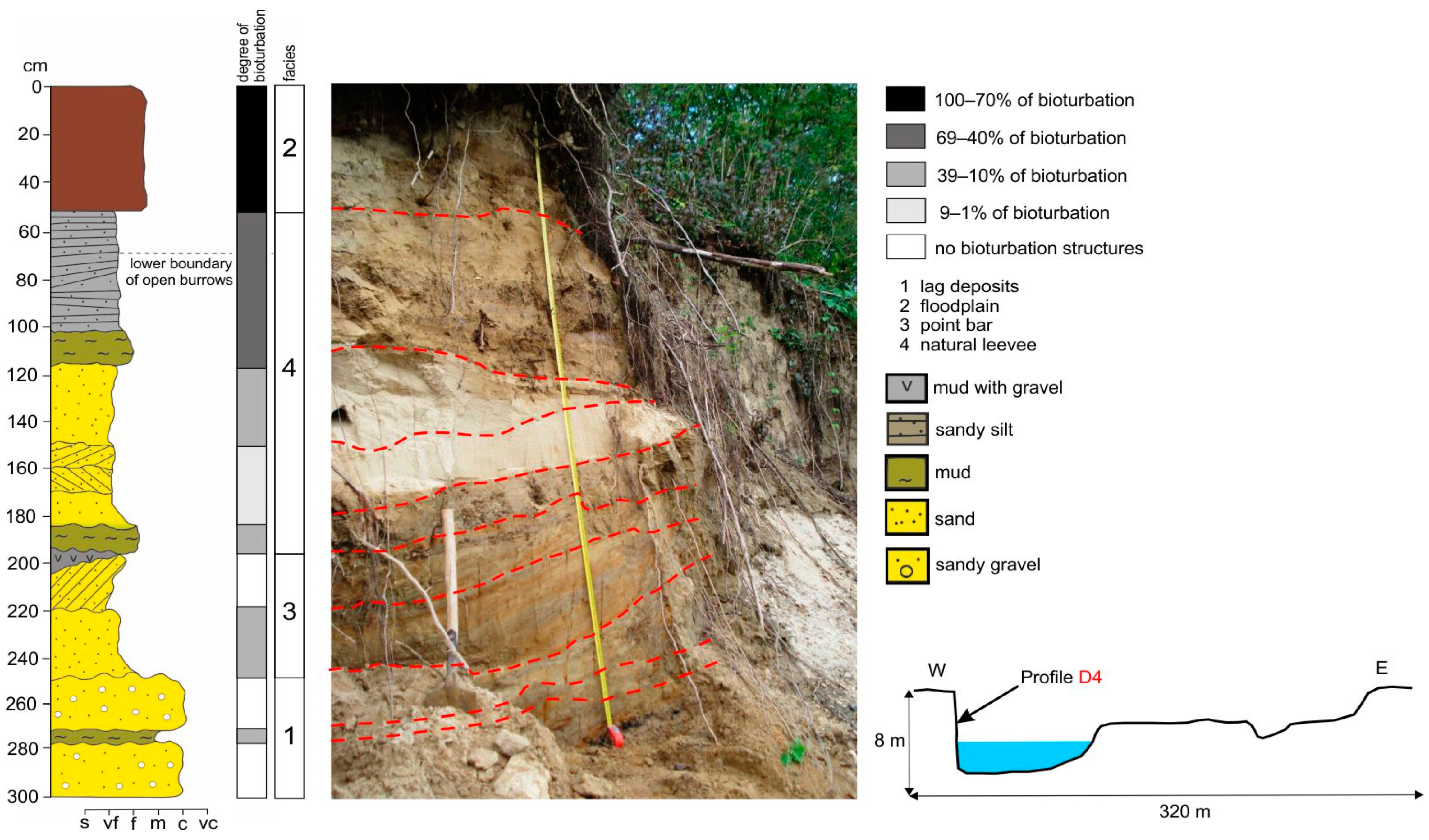
Figure 6.
Characteristic of the vertical profile D4 in the Dunajec River—sediment features and degree of recent bioturbation.
Figure 6.
Characteristic of the vertical profile D4 in the Dunajec River—sediment features and degree of recent bioturbation.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
General characteristic of the studied vertical profiles in the lower reach of the Dunajec River and in the Rogoźnik Stream.
Table 1.
General characteristic of the studied vertical profiles in the lower reach of the Dunajec River and in the Rogoźnik Stream.
| Profile nr | Longitude | Latitude | Elevation [m a.s.l.] | Channel Width [m] | Number of Bioturb./ Unbioturb. Layers | Mean Degree of Bioturbation [%] | Total Thickness of | Geomorphic Unit of the Stream | ||
|---|---|---|---|---|---|---|---|---|---|---|
| In Bioturb. Layers | In Entire Profile | Bioturb. Layers | Unbioturb. Layers | |||||||
| D1 | 50°14′18.99′’ N | 20°44′’02.88′’ E | 170.3 | 71 | 6/2 | 82.2 | 68.1 | 1.88 | 0.39 | natural levee |
| D2 | 50°14′10.13′’ N | 20°44′05.99′’ E | 170.8 | 120 | 6/3 | 79.7 | 69.3 | 2.21 | 0.25 | natural levee |
| D3 | 50°12′00.63′’ N | 20°45′11.51′’ E | 171.3 | 131 | 5/3 | 74.5 | 63.7 | 3.14 | 0.36 | natural levee |
| D4 | 50°10′46.57′’ N | 20°47′54.37′’ E | 172.5 | 112 | 6/3 | 72.1 | 63.3 | 3.55 | 0.35 | natural levee |
| D5 | 50°10′19.05′’ N | 20°49′15.72′’ E | 172.7 | 102 | 5/2 | 87.6 | 76.9 | 1.16 | 0.30 | oxbow lake |
| D6 | 50°10′04.94′’ N | 20°49′31.49′’ E | 172.9 | 111 | 6/2 | 79.7 | 68.4 | 3.05 | 0.34 | natural levee |
| D7 | 50°07′39.68′’ N | 20°52′18.81′’ E | 174.8 | 120 | 5/3 | 73.3 | 65.4 | 1.83 | 0.22 | oxbow lake |
| D8 | 50°06′35.60′’ N | 20°52′13.37′’ E | 175.8 | 153 | 5/2 | 70.0 | 58.6 | 1.70 | 0.33 | natural levee |
| D9 | 50°06′16.97′’ N | 20°52′30.04′’ E | 176.5 | 116 | 4/1 | 74.4 | 61.6 | 2.50 | 0.31 | natural levee |
| D10 | 50°05′07.67′’ N | 20°53′22.85′’ E | 178.2 | 102 | 5/2 | 71.2 | 52.4 | 2.45 | 0.52 | natural levee |
| D11 | 50°04′17.29′’ N | 20°53′50.31′’ E | 180.2 | 122 | 5/2 | 80.9 | 69.8 | 1.33 | 0.21 | oxbow lake |
| D12 | 50°02′16.05′’ N | 20°54′51.61′’ E | 183.3 | 115 | 3/1 | 77.5 | 39.7 | 1.05 | 1.00 | natural levee |
| D13 | 50°00′46.51′’ N | 20°53′23.05′’ E | 187.7 | 130 | 4/1 | 68.4 | 33.8 | 0.83 | 0.85 | natural levee |
| D14 | 49°59′14.19′’ N | 20°53′26.15′’ E | 189.7 | 109 | 3/2 | 66.8 | 47.4 | 1.08 | 0.44 | natural levee |
| D15 | 49°57′16.46′’ N | 20°52′34.69′’ E | 192.9 | 75 | 4/2 | 89.5 | 71.0 | 0.77 | 0.22 | oxbow lake |
| D16 | 49°57′07.47′’ N | 20°49′14.12′’ E | 209.5 | 95 | 3/1 | 56.6 | 35.7 | 0.70 | 0.41 | natural levee |
| D17 | 49°54′57.30′’ N | 20°50′04.02′’ E | 200.2 | 117 | 4/2 | 64.4 | 32.7 | 0.71 | 0.68 | natural levee |
| D18 | 49°53′19.26′’ N | 20°49′41.94′’ E | 203.6 | 115 | 3/2 | 61.5 | 35.2 | 0.63 | 0.47 | natural levee |
| R1 | 49°27′10.57′’ N | 19°59′07.60′’ E | 598.0 | 43 | 3/2 | 58.4 | 23.9 | 0.61 | 0.88 | natural levee |
| R2 | 49°26′56.60′’ N | 19°59′22.36′’ E | 604.4 | 35 | 2/2 | 55.1 | 19.4 | 0.53 | 0.97 | natural levee |
| R3 | 49°26′41.61′’ N | 19°58′09.53′’ E | 605.6 | 41 | 2/2 | 67.0 | 26.8 | 0.70 | 1.05 | paleochannel |
| R4 | 49°26′25.64′’ N | 19°56′24.20′’ E | 614.2 | 42 | 2/2 | 45.2 | 13.3 | 0.44 | 1.05 | natural levee |
| R5 | 49°26′26.81′’ N | 19°55′41.22′’ E | 617.9 | 31 | 2/1 | 48.8 | 20.7 | 0.71 | 0.96 | paleochannel |
| R6 | 49°26′16.20′’ N | 19°55′35.36′’ E | 619.5 | 25 | 1/3 | 60.0 | 11.2 | 0.38 | 1.01 | natural levee |
| R7 | 49°25′20.66′’ N | 19°54′56.19′’ E | 627.7 | 17 | 2/2 | 50.0 | 16.4 | 0.35 | 1.16 | paleochannel |
| R8 | 49°24′54.13′’ N | 19°54′00.68′’ E | 638.4 | 15 | 2/2 | 51.2 | 8.3 | 0.15 | 0.77 | natural levee |
| R9 | 49°24′48.70′’ N | 19°59′19.77′’ E | 645.3 | 6 | 1/3 | 50.0 | 9.3 | 0.21 | 0.58 | natural levee |
| R10 | 49°24′38.72′’ N | 19°57′53.21′’ E | 670.4 | 5 | 1/2 | 60.0 | 8.0 | 0.16 | 0.71 | natural levee |
| R11 | 49°23′02.87′’ N | 19°53′24.39′’ E | 687.5 | 4 | 2/2 | 90.0 | 5.5 | 0.11 | 0.88 | natural levee |
| R12 | 49°23′01.83′’ N | 19°51′49.81′’ E | 680.5 | 5 | 1/2 | 50.0 | 7.5 | 0.13 | 0.76 | natural levee |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mikuś, P. Recent Vertebrate and Invertebrate Burrows in Lowland and Mountain Fluvial Environments (SE Poland). Water 2020, 12, 3413. https://doi.org/10.3390/w12123413
AMA Style
Mikuś P. Recent Vertebrate and Invertebrate Burrows in Lowland and Mountain Fluvial Environments (SE Poland). Water. 2020; 12(12):3413. https://doi.org/10.3390/w12123413
Chicago/Turabian StyleMikuś, Paweł. 2020. "Recent Vertebrate and Invertebrate Burrows in Lowland and Mountain Fluvial Environments (SE Poland)" Water 12, no. 12: 3413. https://doi.org/10.3390/w12123413
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.