Protective Immunity Induced by Virus-Like Particle Containing Merozoite Surface Protein 9 of Plasmodium berghei


 ,
, 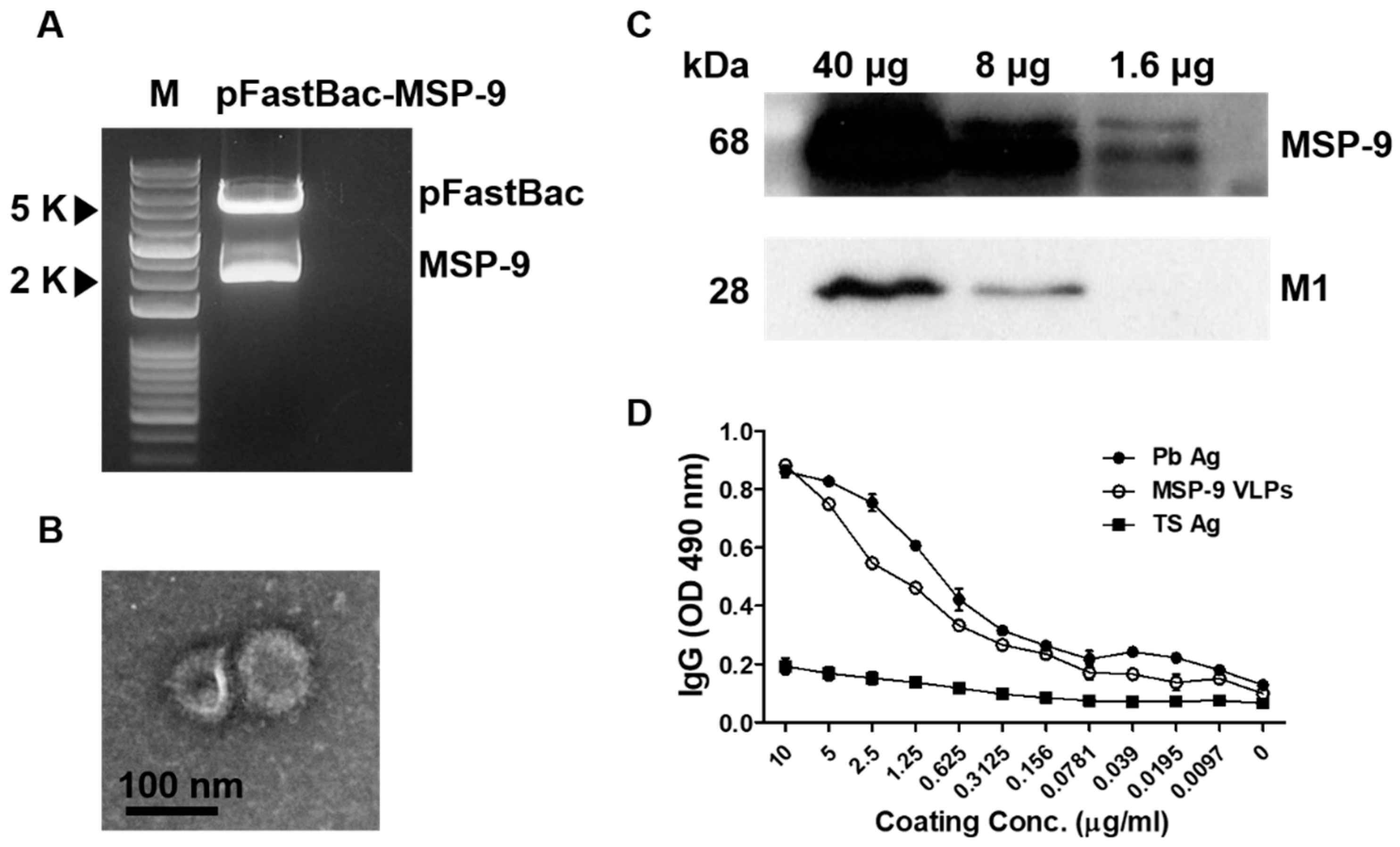{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals, Parasites, Cells, and Antibodies
2.3. Plasmodium Berghei Antigen Preparation
2.4. Generation of Plasmids, Recombinant Baculovirus, and Virus-Like Particles
2.5. Characterization of VLPs
2.6. Mice Immunization and Challenge
2.7. Antibody Responses in Sera
2.8. Immune Cell Responses by Flow Cytometry
2.9. Inflammatory Cytokine Analysis
2.10. Parasitemia
2.11. Statistics
3. Results
3.1. Generation of MSP-9 Virus-Like Particle Vaccines
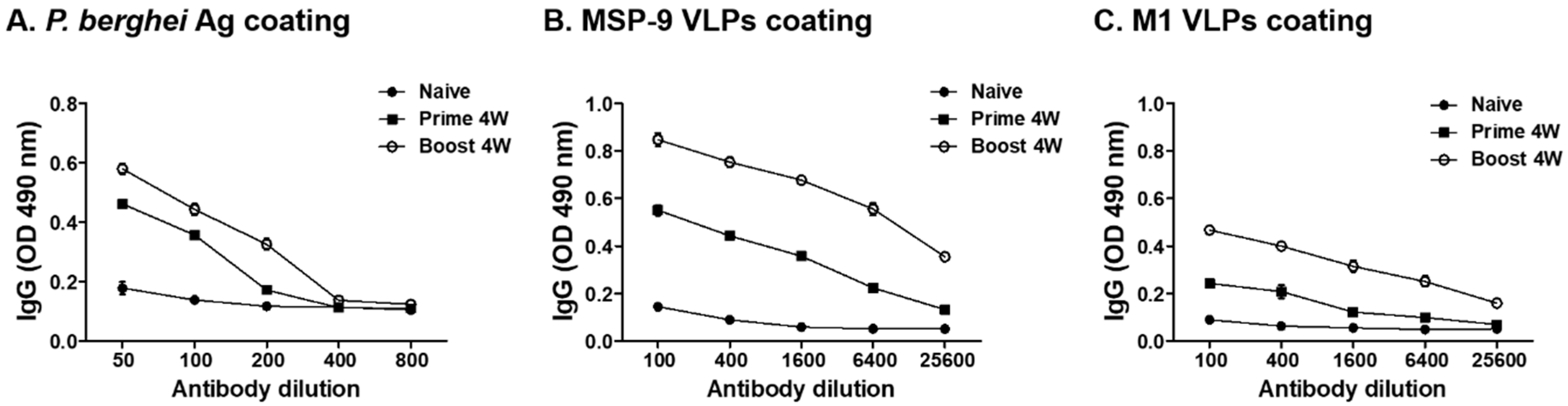
3.2. P. berghei-Specific IgG Antibody Response
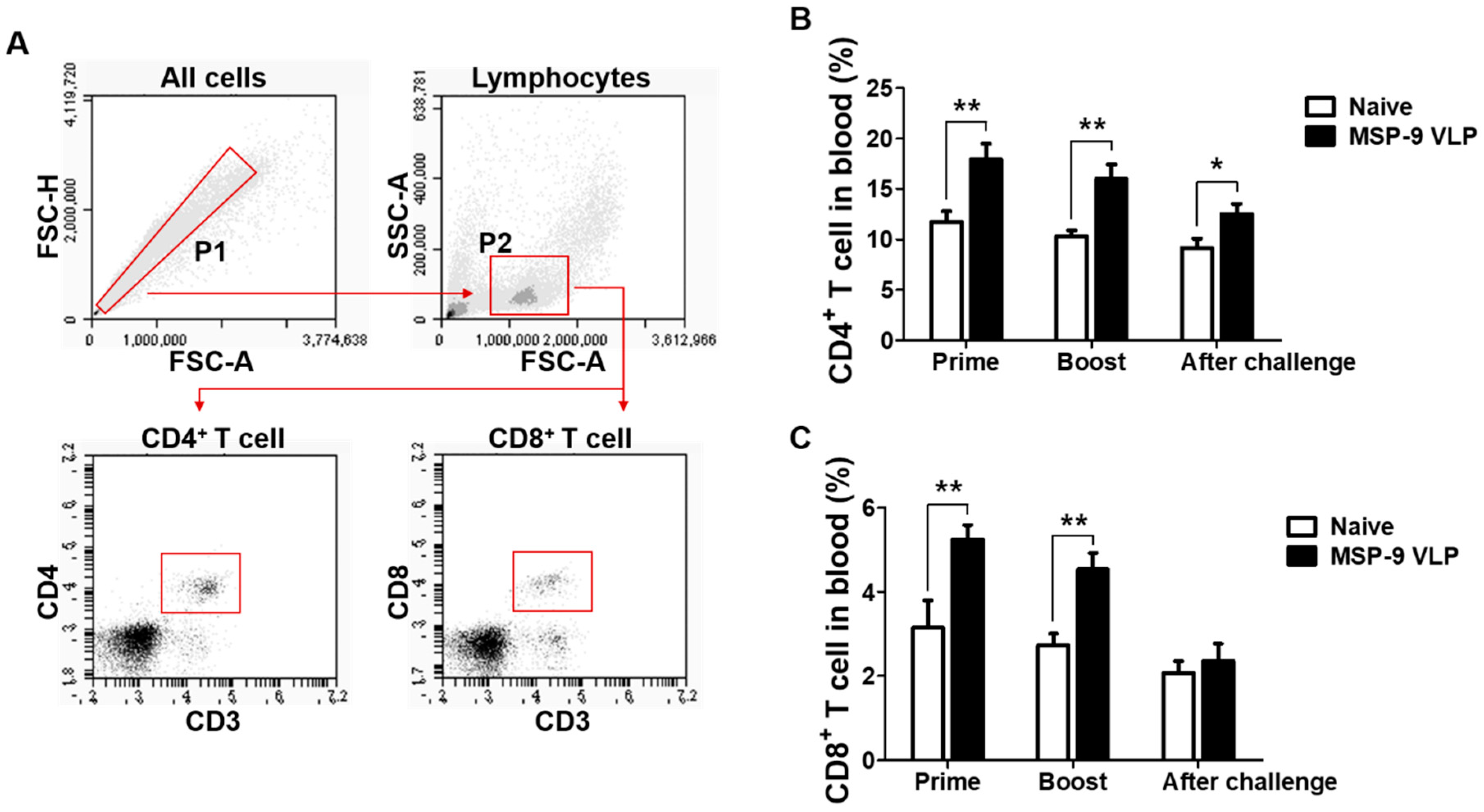
3.3. T Cell Responses in Blood
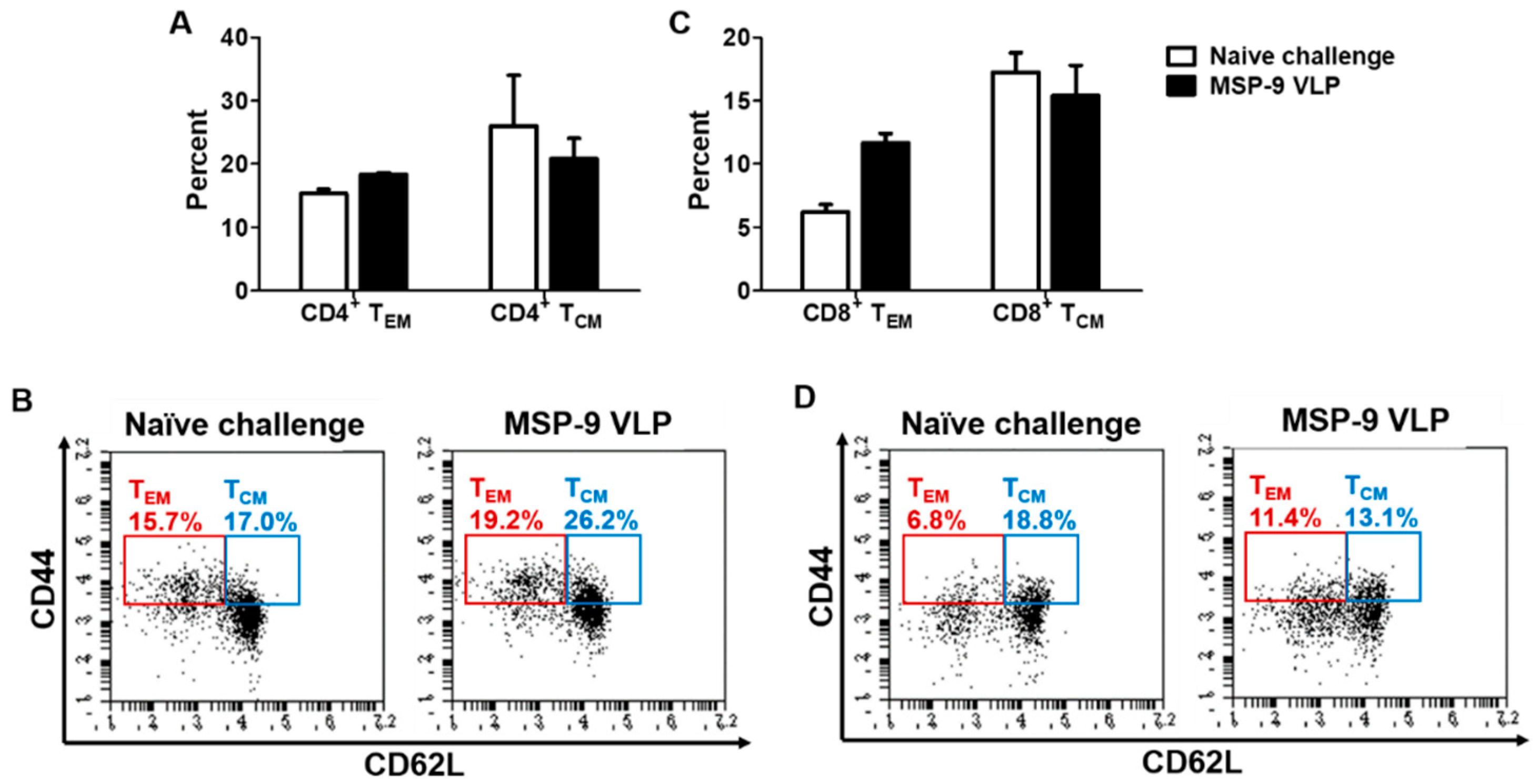
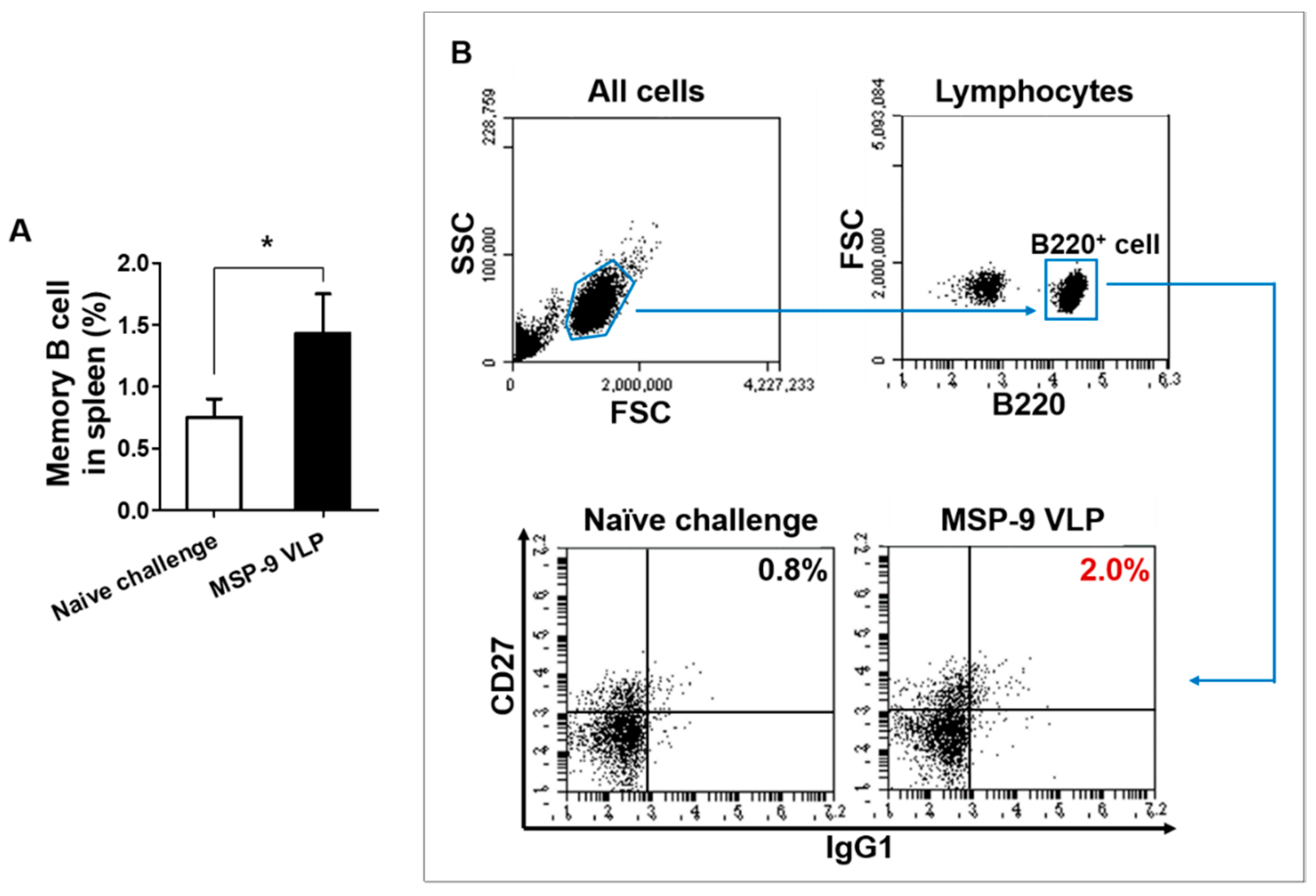
3.4. Memory T and B Cell Responses
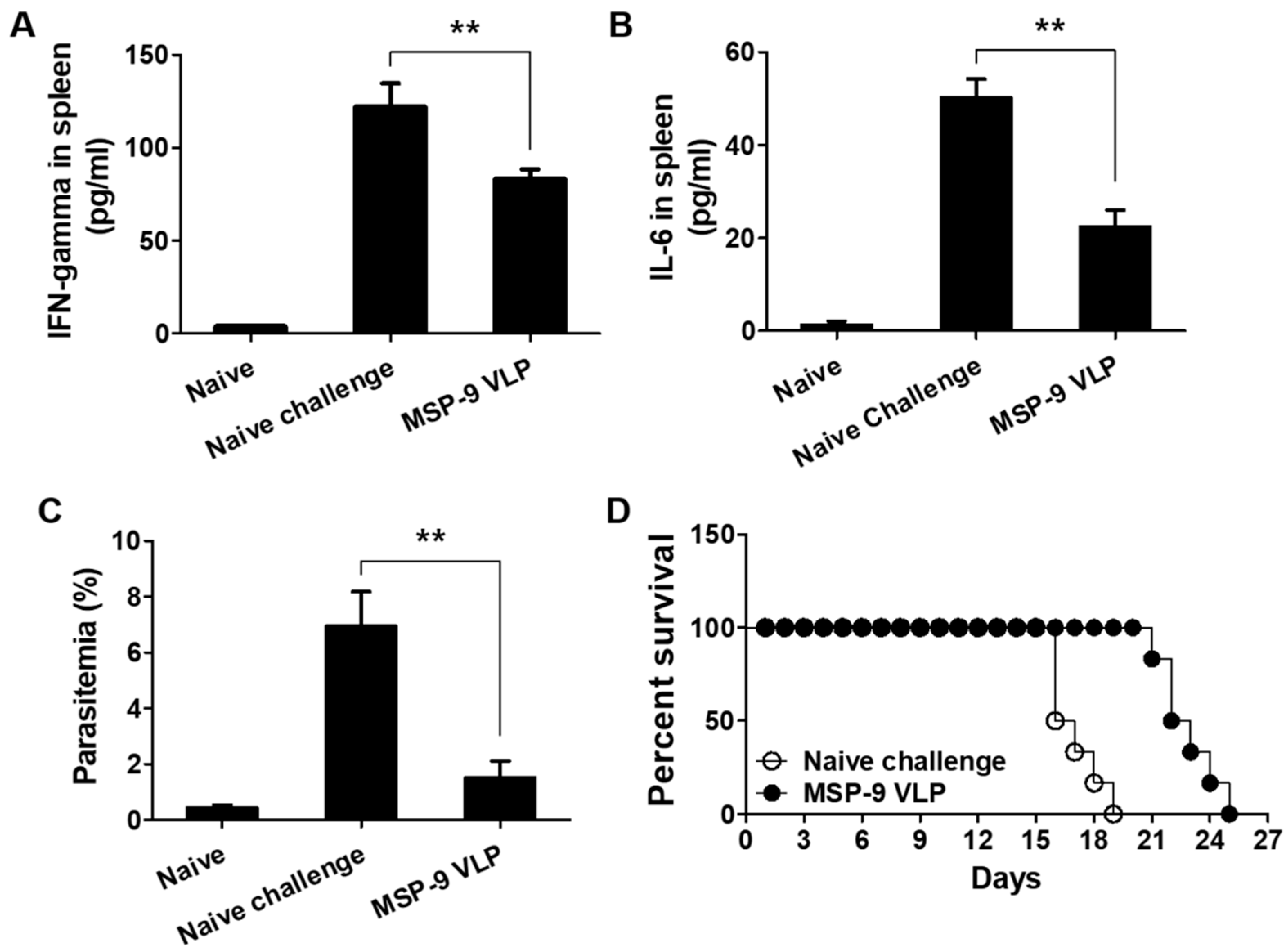
3.5. Pro-Inflammatory Responses and Parasitemia against P. berghei Challenge Infection
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- World Health Organization. World Malaria Report 2019; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- RTS, S Clinical Trials Partnership. Efficacy and safety of RTS, S/AS01 malaria vaccine with or without a booster dose in infants and children in Africa: Final results of a phase 3, individually randomised, controlled trial. Lancet 2015, 386, 31–45. [Google Scholar]
- RTS, S Clinical Trials Partnership. Efficacy and safety of the RTS, S/AS01 malaria vaccine during 18 months after vaccination: A phase 3 randomized, controlled trial in children and young infants at 11 African sites. PLoS Med. 2014, 11, e1001685. [Google Scholar] [CrossRef]
- Rodrigues-da-Silva, R.N.; Correa-Moreira, D.; Soares, I.F.; de-Luca, P.M.; Totino, P.R.R.; Morgado, F.N.; de Oliveira Henriques, M.d.G.; Candea, A.L.P.; Singh, B.; Galinski, M.R. Immunogenicity of synthetic peptide constructs based on PvMSP9E795-A808, a linear B-cell epitope of the P. vivax merozoite surface protein-9. Vaccine 2019, 37, 306–313. [Google Scholar] [CrossRef] [Green Version]
- Stahl, H.D.; Bianco, A.E.; Crewther, P.E.; Anders, R.F.; Kyne, A.P.; Coppel, R.L.; Mitchell, G.F.; Kemp, D.J.; Brown, G.V. Sorting large numbers of clones expressing Plasmodium falciparum antigens in Escherichia coli by differential antibody screening. Mol. Biol. Med. 1986, 3, 351–368. [Google Scholar] [PubMed]
- Weber, J.L.; Lyon, J.A.; Wolff, R.H.; Hall, T.; Lowell, G.H.; Chulay, J.D. Primary structure of a Plasmodium falciparum malaria antigen located at the merozoite surface and within the parasitophorous vacuole. J. Biol. Chem. 1988, 263, 11421–11425. [Google Scholar] [PubMed]
- Lyon, J.A.; Haynes, J.D.; Diggs, C.L.; Chulay, J.D.; Pratt-Rossiter, J.M. Plasmodium falciparum antigens synthesized by schizonts and stabilized at the merozoite surface by antibodies when schizonts mature in the presence of growth inhibitory immune serum. J. Immunol. 1986, 136, 2252–2258. [Google Scholar] [PubMed]
- Nwagwu, M.; Haynes, J.D.; Orlandi, P.A.; Chulay, J.D. Plasmodium falciparum: Chymotryptic-like proteolysis associated with a 101-kDa acidic-basic repeat antigen. Exp. Parasitol 1992, 75, 399–414. [Google Scholar] [CrossRef] [Green Version]
- Curtidor, H.; Urquiza, M.; Suarez, J.; Rodriguez, L.; Ocampo, M.; Puentes, A.; Garcia, J.; Vera, R.; Lopez, R.; Ramirez, L. Plasmodium falciparum acid basic repeat antigen (ABRA) peptides: Erythrocyte binding and biological activity. Vaccine 2001, 19, 4496–4504. [Google Scholar] [CrossRef]
- Sharma, P.; Kumar, A.; Singh, B.; Bharadwaj, A.; Sailaja, V.N.; Adak, T.; Kushwaha, A.; Malhotra, P.; Chauhan, V.S. Characterization of protective epitopes in a highly conserved Plasmodium falciparum antigenic protein containing repeats of acidic and basic residues. Infect. Immun. 1998, 66, 2895–2904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pimtanothai, N.; Parra, M.; Johnson, A.H.; David, C.S.; Katovich Hurley, C. Assessing the binding of four Plasmodium falciparum T helper cell epitopes to HLA-DQ and induction of T-cell responses in HLA-DQ transgenic mice. Infect. Immun. 2000, 68, 1366–1373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira-Ferreira, J.; Vargas-Serrato, E.; Barnwell, J.W.; Moreno, A.; Galinski, M.R. Immunogenicity of Plasmodium vivax merozoite surface protein-9 recombinant proteins expressed in E. coli. Vaccine 2004, 22, 2023–2030. [Google Scholar] [CrossRef] [PubMed]
- Lima-Junior, J.; Banic, D.; Tran, T.; Meyer, V.; De-Simone, S.; Santos, F.; Porto, L.; Marques, M.; Moreno, A.; Barnwell, J. Promiscuous T-cell epitopes of Plasmodium Merozoite Surface Protein 9 (PvMSP9) induces IFN-γ and IL-4 responses in individuals naturally exposed to malaria in the Brazilian Amazon. Vaccine 2010, 28, 3185–3191. [Google Scholar] [CrossRef] [Green Version]
- Jennings, G.T.; Bachmann, M.F. The coming of age of virus-like particle vaccines. Biol. Chem. 2008, 389, 521–536. [Google Scholar] [CrossRef] [PubMed]
- Noad, R.; Roy, P. Virus-like particles as immunogens. Trends Microbiol. 2003, 11, 438–444. [Google Scholar] [CrossRef]
- Buonaguro, L.; Racioppi, L.; Tornesello, M.; Arra, C.; Visciano, M.; Biryahwaho, B.; Sempala, S.; Giraldo, G.; Buonaguro, F. Induction of neutralizing antibodies and cytotoxic T lymphocytes in Balb/c mice immunized with virus-like particles presenting a gp120 molecule from a HIV-1 isolate of clade A. Antivir. Res. 2002, 54, 189–201. [Google Scholar] [CrossRef]
- Buonaguro, L.; Tornesello, M.L.; Tagliamonte, M.; Gallo, R.C.; Wang, L.X.; Kamin-Lewis, R.; Abdelwahab, S.; Lewis, G.K.; Buonaguro, F.M. Baculovirus-derived human immunodeficiency virus type 1 virus-like particles activate dendritic cells and induce ex vivo T-cell responses. J. Virol. 2006, 80, 9134–9143. [Google Scholar] [CrossRef] [Green Version]
- Gatto, D.; Ruedl, C.; Odermatt, B.; Bachmann, M.F. Rapid response of marginal zone B cells to viral particles. J. Immunol. 2004, 173, 4308–4316. [Google Scholar] [CrossRef] [Green Version]
- Lechmann, M.; Murata, K.; Satoi, J.; Vergalla, J.; Baumert, T.F.; Liang, T.J. Hepatitis C virus–like particles induce virus-specific humoral and cellular immune responses in mice. Hepatology 2001, 34, 417–423. [Google Scholar] [CrossRef]
- Sedlik, C.; Dridi, A.; Deriaud, E.; Saron, M.F.; Rueda, P.; Sarraseca, J.; Casal, J.I.; Leclerc, C. Intranasal delivery of recombinant parvovirus-like particles elicits cytotoxic T-cell and neutralizing antibody responses. J. Virol. 1999, 73, 2739–2744. [Google Scholar] [CrossRef] [Green Version]
- Swenson, D.L.; Warfield, K.L.; Negley, D.L.; Schmaljohn, A.; Aman, M.J.; Bavari, S. Virus-like particles exhibit potential as a pan-filovirus vaccine for both Ebola and Marburg viral infections. Vaccine 2005, 23, 3033–3042. [Google Scholar] [CrossRef]
- Lenz, P.; Thompson, C.D.; Day, P.M.; Bacot, S.M.; Lowy, D.R.; Schiller, J.T. Interaction of papillomavirus virus-like particles with human myeloid antigen-presenting cells. Clin. Immunol. 2003, 106, 231–237. [Google Scholar] [CrossRef]
- Lee, D.; Chu, K.; Kang, H.; Lee, S.; Chopra, M.; Choi, H.; Moon, E.; Inn, K.; Quan, F. Protection induced by malaria virus-like particles containing codon-optimized AMA-1 of Plasmodium berghei. Malar. J. 2019, 18, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wykes, M.N.; Good, M.F. What have we learnt from mouse models for the study of malaria? Eur. J. Immunol. 2009, 39, 2004–2007. [Google Scholar] [CrossRef] [PubMed]
- Carter, R.; Diggs, C.L. Plasmodia of rodents. Parasit. Protozoa 1977, 3, 359–465. [Google Scholar]
- Fairlie-Clarke, K.J.; Lamb, T.J.; Langhorne, J.; Graham, A.L.; Allen, J.E. Antibody isotype analysis of malaria-nematode co-infection: Problems and solutions associated with cross-reactivity. BMC Immunol. 2010, 11, 6. [Google Scholar] [CrossRef]
- Cao, Y.; Zhang, D.; Pan, W. Construction of transgenic Plasmodium berghei as a model for evaluation of blood-stage vaccine candidate of Plasmodium falciparum chimeric protein 2.9. PLoS ONE 2009, 4, e6894. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Kim, S.; Lee, D.; Kim, A.; Quan, F. Evaluation of protective efficacy induced by virus-like particles containing a Trichinella spiralis excretory-secretory (ES) protein in mice. Parasites Vectors 2016, 9, 384. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.H.; Lee, S.H.; Kim, A.R.; Quan, F.S. Virus-Like Nanoparticle Vaccine Confers Protection against Toxoplasma gondii. PLoS ONE 2016, 11, e0161231. [Google Scholar] [CrossRef]
- Lee, S.; Kim, A.; Lee, D.; Rubino, I.; Choi, H.; Quan, F. Protection induced by virus-like particles containing Toxoplasma gondii microneme protein 8 against highly virulent RH strain of Toxoplasma gondii infection. PLoS ONE 2017, 12, e0175644. [Google Scholar] [CrossRef]
- Choi, H.; Yoo, D.; Bondy, B.J.; Quan, F.; Compans, R.W.; Kang, S.; Prausnitz, M.R. Stability of influenza vaccine coated onto microneedles. Biomaterials 2012, 33, 3756–3769. [Google Scholar] [CrossRef] [Green Version]
- Somsak, V.; Srichairatanakool, S.; Yuthavong, Y.; Kamchonwongpaisan, S.; Uthaipibull, C. Flow cytometric enumeration of Plasmodium berghei-infected red blood cells stained with SYBR Green I. Acta Trop. 2012, 122, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Chulay, J.D.; Lyon, J.A.; Haynes, J.D.; Meierovics, A.I.; Atkinson, C.T.; Aikawa, M. Monoclonal antibody characterization of Plasmodium falciparum antigens in immune complexes formed when schizonts rupture in the presence of immune serum. J. Immunol. 1987, 139, 2768–2774. [Google Scholar] [PubMed]
- Welsh, J.P. Production of Complex Heterologous Proteins and Protein Assemblies Using E. coli-Based Cell-Free Protein Synthesis; Stanford University: Stanford, CA, USA, 2011. [Google Scholar]
- Amanna, I.J.; Slifka, M.K. Contributions of humoral and cellular immunity to vaccine-induced protection in humans. Virology 2011, 411, 206–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odendahl, M.; Mei, H.; Hoyer, B.F.; Jacobi, A.M.; Hansen, A.; Muehlinghaus, G.; Berek, C.; Hiepe, F.; Manz, R.; Radbruch, A. Generation of migratory antigen-specific plasma blasts and mobilization of resident plasma cells in a secondary immune response. Blood 2005, 105, 1614–1621. [Google Scholar] [CrossRef]
- Cohen, S.; McGregor, I.; Carrington, S. Gamma-globulin and acquired immunity to human malaria. Nature 1961, 192, 733–737. [Google Scholar] [CrossRef]
- Wykes, M.N.; Horne-Debets, J.M.; Leow, C.; Karunarathne, D.S. Malaria drives T cells to exhaustion. Front. Microbiol. 2014, 5, 249. [Google Scholar] [CrossRef] [Green Version]
- Campos, M.; Godson, D.L. The effectiveness and limitations of immune memory: Understanding protective immune responses. Int. J. Parasitol. 2003, 33, 655–661. [Google Scholar] [CrossRef]
- Zielinski, C.E.; Corti, D.; Mele, F.; Pinto, D.; Lanzavecchia, A.; Sallusto, F. Dissecting the human immunologic memory for pathogens. Immunol. Rev. 2011, 240, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Farber, D.L.; Yudanin, N.A.; Restifo, N.P. Human memory T cells: Generation, compartmentalization and homeostasis. Nat. Rev. Immunol. 2014, 14, 24. [Google Scholar] [CrossRef]
- Slifka, M.K.; Antia, R.; Whitmire, J.K.; Ahmed, R. Humoral immunity due to long-lived plasma cells. Immunity 1998, 8, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Sallusto, F. Heterogeneity of human CD4 T cells against microbes. Annu. Rev. Immunol. 2016, 34, 317–334. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kang, H.; Lee, D.; Quan, F. Protective Immunity Induced by Incorporating Multiple Antigenic Proteins of Toxoplasma gondii Into Influenza Virus-Like Particles. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Cunnington, A.J.; Walther, M.; Riley, E.M. Piecing together the puzzle of severe malaria. Sci. Transl. Med. 2013, 5, 211. [Google Scholar] [CrossRef]
- Barber, B.E.; William, T.; Grigg, M.J.; Parameswaran, U.; Piera, K.A.; Price, R.N.; Yeo, T.W.; Anstey, N.M. Parasite biomass-related inflammation, endothelial activation, microvascular dysfunction and disease severity in vivax malaria. PLoS Pathog. 2015, 11, e1004558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.; Chu, K.; Lee, S.; Kim, M.; Park, H.; Jin, H.; Moon, E.; Quan, F. Toxoplasma gondii virus-like particle vaccination alleviates inflammatory response in the brain upon T gondii infection. Parasite Immunol. 2020, 42, e12716. [Google Scholar] [CrossRef] [PubMed]
- Cabral-Miranda, G.; Heath, M.D.; Mohsen, M.O.; Gomes, A.C.; Engeroff, P.; Flaxman, A.; Leoratti, F.; El-Turabi, A.; Reyes-Sandoval, A.; Skinner, M.A. Virus-like particle (VLP) plus microcrystalline tyrosine (MCT) adjuvants enhance vaccine efficacy improving T and B cell immunogenicity and protection against Plasmodium berghei/vivax. Vaccines 2017, 5, 10. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-H.; Kang, H.-J.; Chu, K.-B.; Basak, S.; Lee, D.-H.; Moon, E.-K.; Quan, F.-S. Protective Immunity Induced by Virus-Like Particle Containing Merozoite Surface Protein 9 of Plasmodium berghei. Vaccines 2020, 8, 428. https://doi.org/10.3390/vaccines8030428
Lee S-H, Kang H-J, Chu K-B, Basak S, Lee D-H, Moon E-K, Quan F-S. Protective Immunity Induced by Virus-Like Particle Containing Merozoite Surface Protein 9 of Plasmodium berghei. Vaccines. 2020; 8(3):428. https://doi.org/10.3390/vaccines8030428
Chicago/Turabian StyleLee, Su-Hwa, Hae-Ji Kang, Ki-Back Chu, Swarnendu Basak, Dong-Hun Lee, Eun-Kyung Moon, and Fu-Shi Quan. 2020. "Protective Immunity Induced by Virus-Like Particle Containing Merozoite Surface Protein 9 of Plasmodium berghei" Vaccines 8, no. 3: 428. https://doi.org/10.3390/vaccines8030428