
Characterization of the In Vitro and In Vivo Efficacy of Baloxavir Marboxil against H5 Highly Pathogenic Avian Influenza Virus Infection

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Compounds
2.2. Cells and Viruses
2.3. Viral Yield Reduction Assay
2.4. Animal Experiments
2.4.1. Experiment 1
2.4.2. Experiment 2
2.5. Sequence Analysis of the PA Region in Experiment 1
2.6. Quantitative Analysis of Proinflammatory Cytokines and Chemokines
2.7. Histopathological Experiments
2.8. Assessment of Lung Index
2.9. Combined Effects of BXA and NAIs In Vitro
2.10. Statistical Analysis
2.11. Ethics Statement
3. Results
3.1. Inhibitory Effects of BXA on H5 HPAIV Replication In Vitro
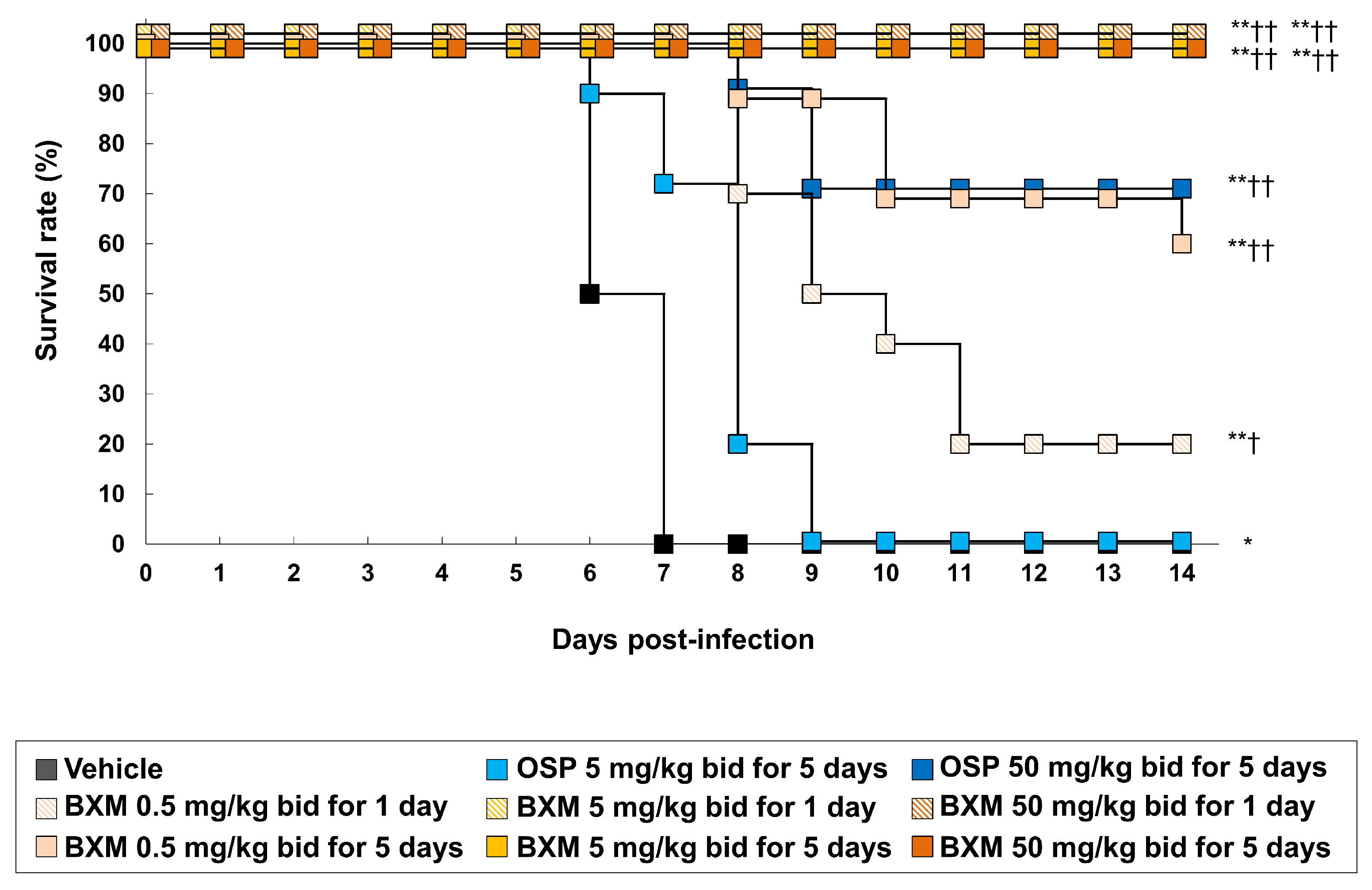
3.2. The Protective Efficacy of BXM on Lethal H5N1 HPAIV Infections In Vivo
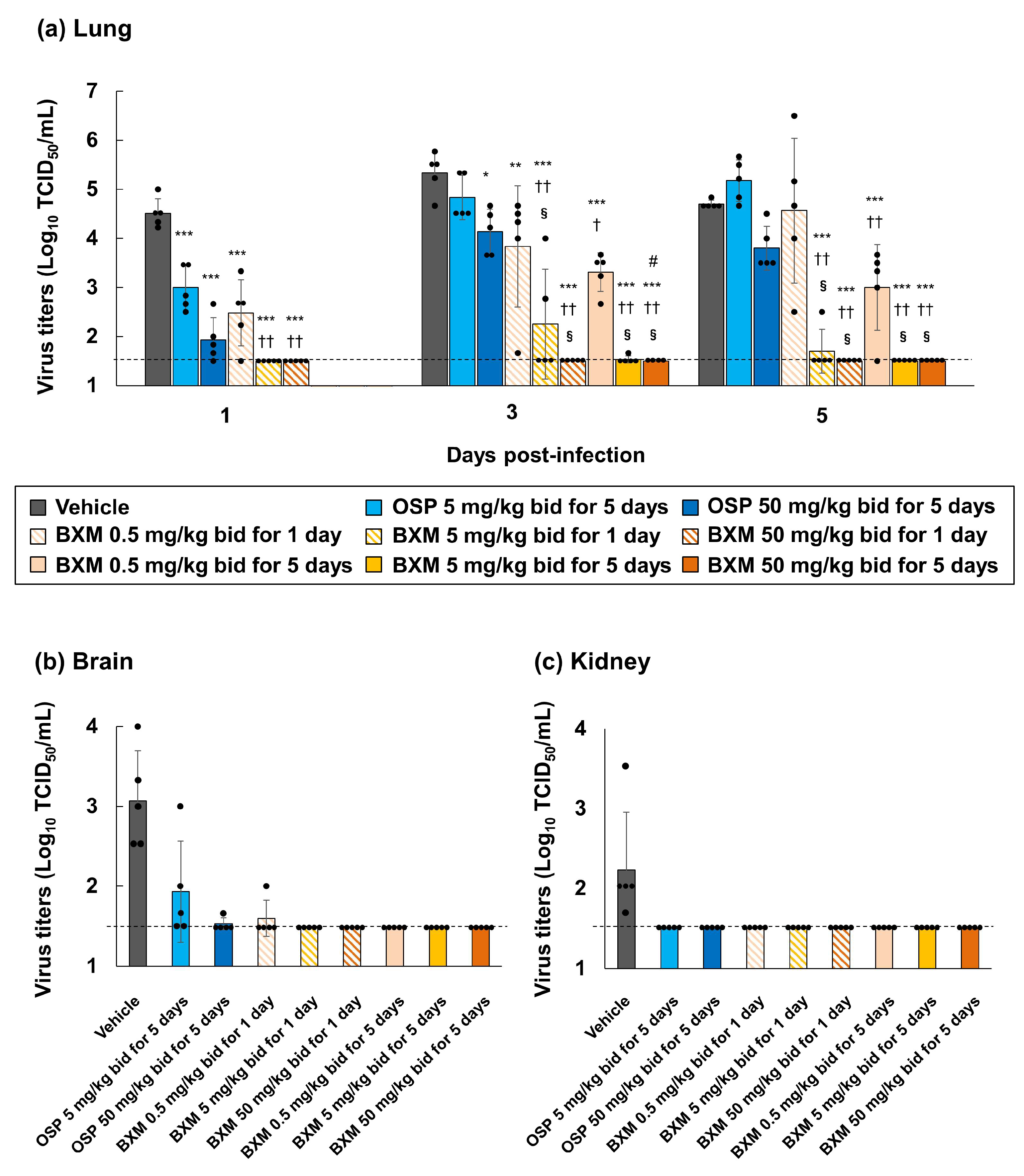
3.3. Effects of BXM on Viral Titers in Mice Infected with H5N1 HPAIVs
3.4. Prevention of Inflammation in Mice Lungs following BXM Treatment
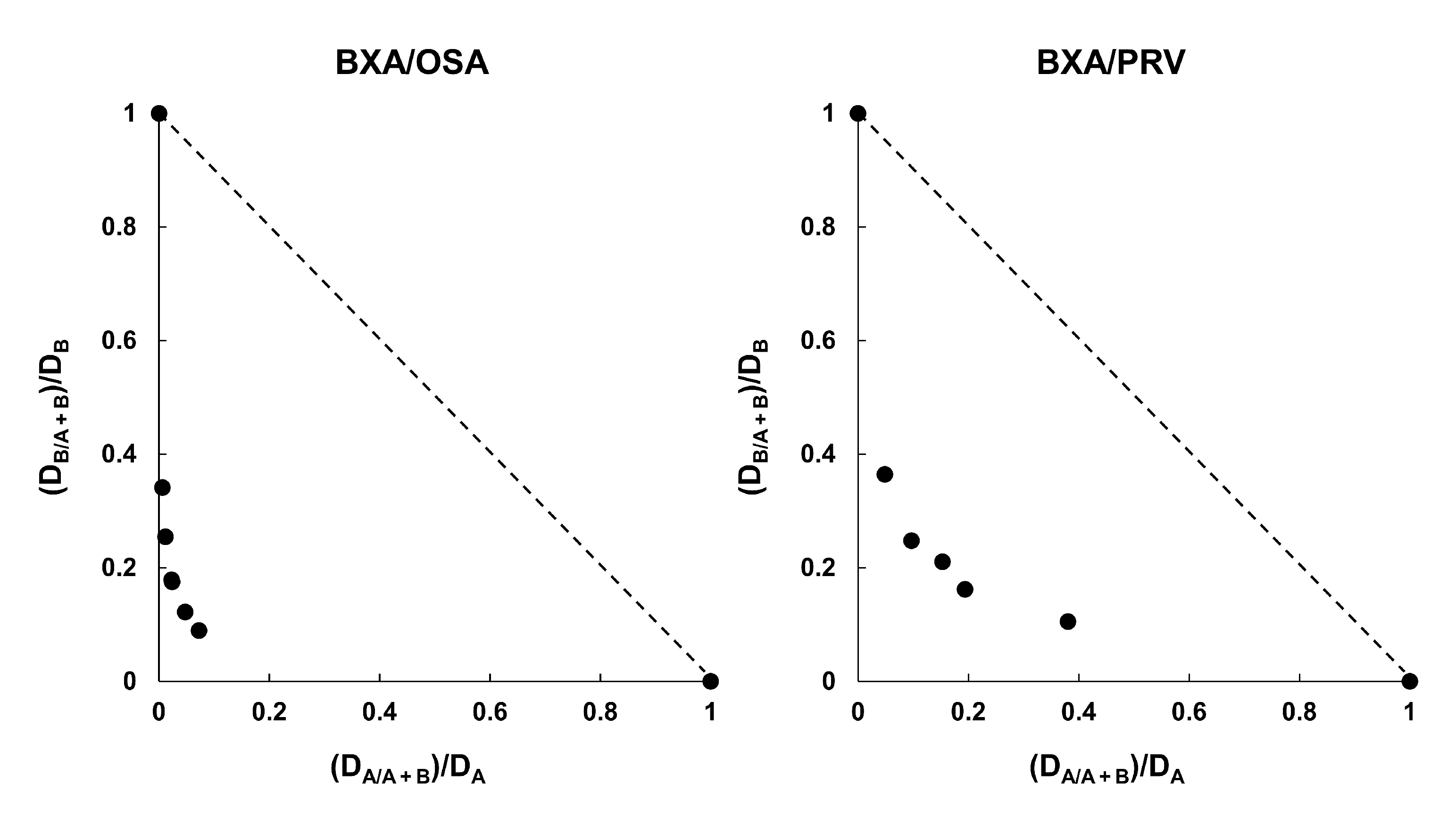
3.5. Combination Efficacy of BXM and NAIs on H5N1 Virus Infections
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Centers for Disease Control and Prevention. Highly Pathogenic Asian Avian Influenza A (H5N1) Virus. Available online: https://www.cdc.gov/flu/avianflu/h5n1-virus.htm (accessed on 14 August 2021).
- Lai, S.; Qin, Y.; Cowling, B.J.; Ren, X.; Wardrop, N.A.; Gilbert, M.; Tsang, T.K.; Wu, P.; Feng, L.; Jiang, H.; et al. Global epidemiology of avian influenza A H5N1 virus infection in humans, 1997–2015: A systematic review of individual case data. Lancet Infect. Dis. 2016, 16, e108–e118. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhang, Z.; Yu, A.; Ho, S.Y.W.; Carr, M.; Zheng, W.; Zhang, Y.; Zhu, C.-D.; Lei, F.; Shi, W. Global and local persistence of influenza A(H5N1) Virus. Emerg. Infect. Dis. 2014, 20, 1287–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Zhao, X.; Li, X.; Bo, H.; Li, D.; Liu, J.; Wang, D. Case report for human infection with a highly pathogenic avian influenza A(H5N6) virus in Beijing, China 2019. Biosaf. Health 2020, 2, 49–52. [Google Scholar] [CrossRef]
- World Health Organization Human Infection with Avian Influenza A (H5N8)—The Russian Federation. Available online: https://www.who.int/csr/don/26-feb-2021-influenza-a-russian-federation/en/ (accessed on 14 August 2021).
- McKenzie, D.K.; Butler, J.E.; Gandevia, S.C.; Lagerquist, O.; Walsh, L.D.; Blouin, J.-S.; Collins, D.F. Respiratory muscle function and activation in chronic obstructive pulmonary disease. J. Appl. Physiol. 2009, 107, 621–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peiris, J.S.M.; de Jong, M.D.; Guan, Y. Avian Influenza Virus (H5N1): A threat to human health. Clin. Microbiol. Rev. 2007, 20, 243–267. [Google Scholar] [CrossRef] [Green Version]
- Hui, K.P.Y.; Lee, S.M.Y.; Cheung, C.-Y.; Ng, I.H.Y.; Poon, L.; Guan, Y.; Ip, N.Y.-Y.; Lau, A.S.Y.; Peiris, J.S.M. Induction of proinflammatory cytokines in primary human macrophages by influenza a virus (H5N1) is selectively regulated by IFN regulatory factor 3 and p38 MAPK. J. Immunol. 2009, 182, 1088–1098. [Google Scholar] [CrossRef] [Green Version]
- Wonderlich, E.R.; Swan, Z.D.; Bissel, S.J.; Hartman, A.L.; Carney, J.P.; O’Malley, K.J.; Obadan, A.O.; Santos, J.; Walker, R.; Sturgeon, T.J.; et al. Widespread virus replication in alveoli drives acute respiratory distress syndrome in aerosolized H5N1 influenza infection of macaques. J. Immunol. 2017, 198, 1616–1626. [Google Scholar] [CrossRef] [Green Version]
- de Jong, M.D.; Simmons, C.P.; Thanh, T.T.; Hien, V.M.; Smith, G.J.D.; Chau, T.N.B.; Hoang, D.M.; Chau, N.V.V.; Khanh, T.H.; Dong, V.C.; et al. Fatal outcome of human influenza A (H5N1) is associated with high viral load and hypercytokinemia. Nat. Med. 2006, 12, 1203–1207. [Google Scholar] [CrossRef]
- Kobayashi, M.; Kodama, M.; Noshi, T.; Yoshida, R.; Kanazu, T.; Nomura, N.; Soda, K.; Isoda, N.; Okamatsu, M.; Sakoda, Y.; et al. Therapeutic efficacy of peramivir against H5N1 highly pathogenic avian influenza viruses harboring the neuraminidase H275Y mutation. Antivir. Res. 2017, 139, 41–48. [Google Scholar] [CrossRef]
- Shionogi announces European Commission Approval of XOFLUZA® (Baloxavir Marboxil) for the Treatment and Post-Exposure Prophylaxis of Influenza Virus Infection. Available online: https://www.shionogi.com/global/en/news/2021/01/e-210115_2.html (accessed on 26 December 2021).
- Noshi, T.; Kitano, M.; Taniguchi, K.; Yamamoto, A.; Omoto, S.; Baba, K.; Hashimoto, T.; Ishida, K.; Kushima, Y.; Hattori, K.; et al. In vitro characterization of baloxavir acid, a first-in-class cap-dependent endonuclease inhibitor of the influenza virus polymerase PA subunit. Antivir. Res. 2018, 160, 109–117. [Google Scholar] [CrossRef]
- Omoto, S.; Speranzini, V.; Hashimoto, T.; Noshi, T.; Yamaguchi, H.; Kawai, M.; Kawaguchi, K.; Uehara, T.; Shishido, T.; Naito, A.; et al. Characterization of influenza virus variants induced by treatment with the endonuclease inhibitor baloxavir marboxil. Sci. Rep. 2018, 8, 9633. [Google Scholar] [CrossRef]
- Mishin, V.P.; Patel, M.C.; Chesnokov, A.; De La Cruz, J.; Nguyen, H.T.; Lollis, L.; Hodges, E.; Jang, Y.; Barnes, J.; Uyeki, T.; et al. Susceptibility of influenza A, B, C, and D viruses to baloxavir1. Emerg. Infect. Dis. 2019, 25, 1969–1972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayden, F.G.; Sugaya, N.; Hirotsu, N.; Lee, N.; De Jong, M.D.; Hurt, A.C.; Ishida, T.; Sekino, H.; Yamada, K.; Portsmouth, S.; et al. Baloxavir Marboxil for Uncomplicated Influenza in Adults and Adolescents. N. Engl. J. Med. 2018, 379, 913–923. [Google Scholar] [CrossRef] [PubMed]
- Fukao, K.; Noshi, T.; Yamamoto, A.; Kitano, M.; Ando, Y.; Noda, T.; Baba, K.; Matsumoto, K.; Higuchi, N.; Ikeda, M.; et al. Combination treatment with the cap-dependent endonuclease inhibitor baloxavir marboxil and a neuraminidase inhibitor in a mouse model of influenza A virus infection. J. Antimicrob. Chemother. 2018, 74, 654–662. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, A.; Ishida, T.; Hirotsu, N.; Kawaguchi, K.; Ishibashi, T.; Shishido, T.; Sato, C.; Portsmouth, S.; Tsuchiya, K.; Uehara, T. Baloxavir marboxil in Japanese patients with seasonal influenza: Dose response and virus type/subtype outcomes from a randomized phase 2 study. Antivir. Res. 2019, 163, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Fukao, K.; Ando, Y.; Noshi, T.; Kitano, M.; Noda, T.; Kawai, M.; Yoshida, R.; Sato, A.; Shishido, T.; Naito, A. Baloxavir marboxil, a novel cap-dependent endonuclease inhibitor potently suppresses influenza virus replication and represents therapeutic effects in both immunocompetent and immunocompromised mouse models. PLoS ONE 2019, 14, e0217307. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, K.; Ando, Y.; Nobori, H.; Toba, S.; Noshi, T.; Kobayashi, M.; Kawai, M.; Yoshida, R.; Sato, A.; Shishido, T.; et al. Inhibition of avian-origin influenza A(H7N9) virus by the novel cap-dependent endonuclease inhibitor baloxavir marboxil. Sci. Rep. 2019, 9, 3466. [Google Scholar] [CrossRef] [Green Version]
- Ikematsu, H.; Hayden, F.G.; Kawaguchi, K.; Kinoshita, M.; De Jong, M.D.; Lee, N.; Takashima, S.; Noshi, T.; Tsuchiya, K.; Uehara, T. Baloxavir Marboxil for Prophylaxis against Influenza in Household Contacts. N. Engl. J. Med. 2020, 383, 309–320. [Google Scholar] [CrossRef]
- Wang, Y.; Fan, G.; Salam, A.; Horby, P.; Hayden, F.G.; Chen, C.; Pan, J.; Zheng, J.; Lu, B.; Guo, L.; et al. Comparative effectiveness of combined favipiravir and oseltamivir therapy versus oseltamivir monotherapy in critically Ill patients with influenza virus infection. J. Infect. Dis. 2019, 221, 1688–1698. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, Y.; Sasaki, H.; Horiuchi, H.; Miyata, N.; Kawakami, C.; Usuku, S.; Tachikawa, N. Early combination treatment with baloxavir and peramivir for hospitalized adults with influenza A in Yokohama, Japan. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1637–1640. [Google Scholar] [CrossRef] [PubMed]
- Smee, D.F.; Hurst, B.L.; Wong, M.-H.; Bailey, K.W.; Tarbet, E.B.; Morrey, J.D.; Furuta, Y. Effects of the combination of favipiravir (t-705) and oseltamivir on influenza a virus infections in mice. Antimicrob. Agents Chemother. 2010, 54, 126–133. [Google Scholar] [CrossRef] [Green Version]
- Marathe, B.M.; Wong, S.-S.; Vogel, P.; Garcia-Alcalde, F.; Webster, R.G.; Webby, R.J.; Najera, I.; Govorkova, E.A. Combinations of oseltamivir and T-705 extend the treatment window for highly pathogenic influenza A(H5N1) virus infection in mice. Sci. Rep. 2016, 6, 26742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilyushina, N.A.; Hoffmann, E.; Salomon, R.; Webster, R.G.; Govorkova, E.A. Amantadine-oseltamivir combination therapy for H5N1 influenza virus infection in mice. Antivir. Ther. 2007, 12, 363–370. [Google Scholar]
- Govorkova, E.; Leneva, I.A.; Goloubeva, O.G.; Bush, K.; Webster, R.G. Comparison of efficacies of RWJ-270201, zanamivir, and oseltamivir against H5N1, H9N2, and other avian influenza viruses. Antimicrob. Agents Chemother. 2001, 45, 2723–2732. [Google Scholar] [CrossRef] [Green Version]
- Liem, N.T.; Tung, C.V.; Hien, N.T.; Hien, T.T.; Chau, N.Q.; Long, H.T.; Mai, L.Q.; Taylor, W.R.J.; Wertheim, H.; Farrar, J.; et al. Clinical features of human influenza a (H5N1) infection in Vietnam: 2004–2006. Clin. Infect. Dis. 2009, 48, 1639–1646. [Google Scholar] [CrossRef] [Green Version]
- De Jong, M.D.; Thanh, T.T.; Khanh, T.H.; Hien, V.M.; Smith, G.J.; Chau, N.V.; Van Cam, B.; Qui, P.T.; Ha, D.Q.; Guan, Y.; et al. Oseltamivir resistance during treatment of influenza a (H5N1) infection. N. Engl. J. Med. 2005, 353, 2667–2672. [Google Scholar] [CrossRef] [Green Version]
- Karthick, V.; Ramanathan, K. Insight into the Oseltamivir Resistance R292K Mutation in H5N1 Influenza Virus: A Molecular Docking and Molecular Dynamics Approach. Cell Biophys. 2013, 68, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Govorkova, E.; Baranovich, T.; Seiler, P.; Armstrong, J.; Burnham, A.; Guan, Y.; Peiris, J.S.M.; Webby, R.J.; Webster, R.G. Antiviral resistance among highly pathogenic influenza A (H5N1) viruses isolated worldwide in 2002–2012 shows need for continued monitoring. Antivir. Res. 2013, 98, 297–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaduangrat, N.; Phanich, J.; Rungrotmongkol, T.; Lerdsamran, H.; Puthavathana, P.; Ubol, S. The significance of naturally occurring neuraminidase quasispecies of H5N1 avian influenza virus on resistance to oseltamivir: A point of concern. J. Gen. Virol. 2016, 97, 1311–1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uehara, T.; Hayden, F.G.; Kawaguchi, K.; Omoto, S.; Hurt, A.C.; De Jong, M.D.; Hirotsu, N.; Sugaya, N.; Lee, N.; Baba, K.; et al. Treatment-emergent influenza variant viruses with reduced baloxavir susceptibility: Impact on clinical and virologic outcomes in uncomplicated influenza. J. Infect. Dis. 2019, 221, 346–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takashita, E.; Morita, H.; Ogawa, R.; Nakamura, K.; Fujisaki, S.; Shirakura, M.; Kuwahara, T.; Kishida, N.; Watanabe, S.; Odagiri, T. Susceptibility of influenza viruses to the novel cap-dependent endonuclease inhibitor baloxavir marboxil. Front. Microbiol. 2018, 9, 3026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takashita, E.; Abe, T.; Morita, H.; Nagata, S.; Fujisaki, S.; Miura, H.; Shirakura, M.; Kishida, N.; Nakamura, K.; Kuwahara, T.; et al. Influenza A(H1N1)pdm09 virus exhibiting reduced susceptibility to baloxavir due to a PA E23K substitution detected from a child without baloxavir treatment. Antiviral Res. 2020, 180, 104828. [Google Scholar] [CrossRef]
- Nishimura, H.; Itamura, S.; Iwasaki, T.; Kurata, T.; Tashiro, M. Characterization of human influenza A (H5N1) virus infection in mice: Neuro-, pneumo- and adipotropic infection. J. Gen. Virol. 2000, 81, 2503–2510. [Google Scholar] [CrossRef] [Green Version]
- Tumpey, T.M.; Lu, X.; Morken, T.; Zaki, S.R.; Katz, J.M. Depletion of lymphocytes and diminished cytokine production in mice infected with a highly virulent influenza A (H5N1) virus isolated from humans. J. Virol. 2000, 74, 6105–6116. [Google Scholar] [CrossRef] [Green Version]
- Shichinohe, S.; Okamatsu, M.; Yamamoto, N.; Noda, Y.; Nomoto, Y.; Honda, T.; Takikawa, N.; Sakoda, Y.; Kida, H. Potency of an inactivated influenza vaccine prepared from a non-pathogenic H5N1 virus against a challenge with antigenically drifted highly pathogenic avian influenza viruses in chickens. Veter Microbiol. 2013, 164, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Sakoda, Y.; Sugar, S.; Batchluun, D.; Erdene-Ochir, T.-O.; Okamatsu, M.; Isoda, N.; Soda, K.; Takakuwa, H.; Tsuda, Y.; Yamamoto, N.; et al. Characterization of H5N1 highly pathogenic avian influenza virus strains isolated from migratory waterfowl in Mongolia on the way back from the southern Asia to their northern territory. Virology 2010, 406, 88–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiono, T.; Okamatsu, M.; Matsuno, K.; Haga, A.; Iwata, R.; Nguyen, L.T.; Suzuki, M.; Kikutani, Y.; Kida, H.; Onuma, M.; et al. Characterization of H5N6 highly pathogenic avian influenza viruses isolated from wild and captive birds in the winter season of 2016-2017 in Northern Japan. Microbiol. Immunol. 2017, 61, 387–397. [Google Scholar] [CrossRef]
- Isoda, N.; Twabela, A.T.; Bazarragchaa, E.; Ogasawara, K.; Hayashi, H.; Wang, Z.-J.; Kobayashi, D.; Watanabe, Y.; Saito, K.; Kida, H.; et al. Re-Invasion of H5N8 High pathogenicity avian influenza virus clade 2.3.4.4b in Hokkaido, Japan, 2020. Viruses 2020, 12, 1439. [Google Scholar] [CrossRef]
- Hoffmann, E.; Neumann, G.; Kawaoka, Y.; Hobom, G.; Webster, R.G. A DNA transfection system for generation of influenza A virus from eight plasmids. Proc. Natl. Acad. Sci. USA 2000, 97, 6108–6113. [Google Scholar] [CrossRef] [Green Version]
- Ward, P.; Small, I.; Smith, J.; Suter, P.; Dutkowski, R. Oseltamivir (Tamiflu®) and its potential for use in the event of an influenza pandemic. J. Antimicrob. Chemother. 2005, 55, i5–i21. [Google Scholar] [CrossRef] [Green Version]
- Qiao, J.; Zhang, M.; Bi, J.; Wang, X.; Deng, G.; He, G.; Luan, Z.; Lv, N.; Xun, W.; Zhao, L. Pulmonary fibrosis induced by H5N1 viral infection in mice. Respir. Res. 2009, 10, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobori, H.; Toba, S.; Yoshida, R.; Hall, W.W.; Orba, Y.; Sawa, H.; Sato, A. Identification of compound-B, a novel anti-dengue virus agent targeting the non-structural protein 4A. Antivir. Res. 2018, 155, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Earhart, K.C.; Elsayed, N.M.; Saad, M.D.; Gubareva, L.V.; Nayel, A.; Deyde, V.M.; Abdelsattar, A.; AbdelGhani, A.S.; Boynton, B.R.; Mansour, M.M.; et al. Oseltamivir resistance mutation N294S in human influenza A(H5N1) virus in Egypt. J. Infect. Public Health 2009, 2, 74–80. [Google Scholar] [CrossRef] [Green Version]
- Park, C.H.; Ishinaka, M.; Takada, A.; Kida, H.; Kimura, T.; Ochiai, K.; Umemura, T. The invasion routes of neurovirulent A/Hong Kong/483/97 (H5N1) influenza virus into the central nervous system after respiratory infection in mice. Arch. Virol. 2002, 147, 1425–1436. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Tumpey, T.M.; Morken, T.; Zaki, S.R.; Cox, N.J.; Katz, J.M. A mouse model for the evaluation of pathogenesis and immunity to influenza A (H5N1) viruses isolated from humans. J. Virol. 1999, 73, 5903–5911. [Google Scholar] [CrossRef] [Green Version]
- Maines, T.R.; Lu, X.H.; Erb, S.M.; Edwards, L.; Guarner, J.; Greer, P.W.; Nguyen, D.C.; Szretter, K.J.; Chen, L.-M.; Thawatsupha, P.; et al. Avian influenza (H5N1) viruses isolated from humans in Asia in 2004 exhibit increased virulence in mammals. J. Virol. 2005, 79, 11788–11800. [Google Scholar] [CrossRef] [Green Version]
- Checkmahomed, L.; Padey, B.; Pizzorno, A.; Terrier, O.; Rosa-Calatrava, M.; Abed, Y.; Baz, M.; Boivin, G. In Vitro combinations of baloxavir acid and other inhibitors against seasonal influenza a viruses. Viruses 2020, 12, 1139. [Google Scholar] [CrossRef]
- Flannery, A.H.; Bastin, M.L.T. Oseltamivir dosing in critically Ill patients with severe influenza. Ann. Pharmacother. 2014, 48, 1011–1018. [Google Scholar] [CrossRef]
- Westenius, V.; Mäkelä, S.M.; Julkunen, I.; Österlund, P. Highly pathogenic H5N1 influenza A virus spreads efficiently in human primary monocyte-derived macrophages and dendritic cells. Front. Immunol. 2018, 9, 1664. [Google Scholar] [CrossRef]
- Plourde, J.R.; Pyles, J.A.; Layton, R.C.; Vaughan, S.E.; Tipper, J.L.; Harrod, K.S. Neurovirulence of H5N1 Infection in ferrets is mediated by multifocal replication in distinct permissive neuronal cell regions. PLoS ONE 2012, 7, e46605. [Google Scholar] [CrossRef] [Green Version]
- Zitzow, L.A.; Rowe, T.; Morken, T.; Shieh, W.-J.; Zaki, S.; Katz, J.M. Pathogenesis of avian influenza a (H5N1) viruses in ferrets. J. Virol. 2002, 76, 4420–4429. [Google Scholar] [CrossRef] [Green Version]
- Rimmelzwaan, G.F.; Kuiken, T.; van Amerongen, G.; Bestebroer, T.M.; Fouchier, R.; Osterhaus, A.D.M.E. Pathogenesis of influenza a (H5N1) virus infection in a primate model. J. Virol. 2001, 75, 6687–6691. [Google Scholar] [CrossRef] [Green Version]
- Szretter, K.J.; Gangappa, S.; Lu, X.; Smith, C.; Shieh, W.-J.; Zaki, S.R.; Sambhara, S.; Tumpey, T.M.; Katz, J.M. Role of host cytokine responses in the pathogenesis of avian H5N1 influenza viruses in mice. J. Virol. 2007, 81, 2736–2744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spesock, A.; Malur, M.; Hossain, M.J.; Chen, L.-M.; Njaa, B.L.; Davis, C.T.; Lipatov, A.S.; York, I.A.; Krug, R.M.; Donis, R.O. The virulence of 1997 H5N1 influenza viruses in the mouse model is increased by correcting a defect in their NS1 proteins. J. Virol. 2011, 85, 7048–7058. [Google Scholar] [CrossRef] [Green Version]
- Perrone, L.A.; Plowden, J.K.; Garcia-Sastre, A.; Katz, J.M.; Tumpey, T.M. H5N1 and 1918 pandemic influenza virus infection results in early and excessive infiltration of macrophages and neutrophils in the lungs of mice. PLoS Pathog. 2008, 4, e1000115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, H.; Wang, N.; Che, H.-L.; Zhao, Y.; Jin, H. Pathological observations of lung inflammation after administration of IP-10 in influenza virus- and respiratory syncytial virus-infected mice. Exp. Ther. Med. 2011, 3, 76–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, T.; Qiao, J.; Zhao, L.; Wang, G.; He, G.; Li, K.; Tian, Y.; Gao, M.; Wang, J.; Wang, H.; et al. Acute Respiratory distress syndrome induced by avian influenza a (H5N1) virus in mice. Am. J. Respir. Crit. Care Med. 2006, 174, 1011–1017. [Google Scholar] [CrossRef] [PubMed]
- Ogiwara, H.; Yasui, F.; Munekata, K.; Takagi-Kamiya, A.; Munakata, T.; Nomura, N.; Shibasaki, F.; Kuwahara, K.; Sakaguchi, N.; Sakoda, Y.; et al. Histopathological evaluation of the diversity of cells susceptible to H5N1 virulent avian influenza virus. Am. J. Pathol. 2014, 184, 171–183. [Google Scholar] [CrossRef]
- Bird, N.L.; Olson, M.R.; Hurt, A.; Oshansky, C.M.; Oh, D.Y.; Reading, P.; Chua, B.; Sun, Y.; Tang, L.; Handel, A.; et al. Oseltamivir prophylaxis reduces inflammation and facilitates establishment of cross-strain protective T cell memory to influenza viruses. PLoS ONE 2015, 10, e0129768. [Google Scholar] [CrossRef]
- Salomon, R.; Hoffmann, E.; Webster, R.G. Inhibition of the cytokine response does not protect against lethal H5N1 influenza infection. Proc. Natl. Acad. Sci. USA 2007, 104, 12479–12481. [Google Scholar] [CrossRef] [Green Version]
- Salomon, R.; Franks, J.; Govorkova, E.; Ilyushina, N.A.; Yen, H.-L.; Hulse-Post, D.J.; Humberd, J.; Trichet, M.; Rehg, J.E.; Webby, R.J.; et al. The polymerase complex genes contribute to the high virulence of the human H5N1 influenza virus isolate A/Vietnam/1203/04. J. Exp. Med. 2006, 203, 689–697. [Google Scholar] [CrossRef] [Green Version]
- Byrn, R.A.; Jones, S.M.; Bennett, H.B.; Bral, C.; Clark, M.P.; Jacobs, M.D.; Kwong, A.D.; Ledeboer, M.W.; Leeman, J.R.; McNeil, C.F.; et al. Preclinical activity of VX-787, a first-in-class, orally bioavailable inhibitor of the influenza virus polymerase PB2 subunit. Antimicrob. Agents Chemother. 2015, 59, 1569–1582. [Google Scholar] [CrossRef] [Green Version]
- Kiso, M.; Takahashi, K.; Sakai-Tagawa, Y.; Shinya, K.; Sakabe, S.; Le, Q.M.; Ozawa, M.; Furuta, Y.; Kawaoka, Y. T-705 (favipiravir) activity against lethal H5N1 influenza A viruses. Proc. Natl. Acad. Sci. USA 2010, 107, 882–887. [Google Scholar] [CrossRef] [Green Version]
- Nogales, A.; Villamayor, L.; Utrilla-Trigo, S.; Ortego, J.; Martinez-Sobrido, L.; DeDiego, M.L. Natural selection of H5N1 avian influenza A Viruses with increased PA-X and NS1 shutoff activity. Viruses 2021, 13, 1760. [Google Scholar] [CrossRef] [PubMed]
- Svyatchenko, S.V.; Goncharova, N.I.; Marchenko, V.Y.; Kolosova, N.P.; Shvalov, A.N.; Kovrizhkina, V.L.; Durymanov, A.G.; Onkhonova, G.S.; Tregubchak, T.V.; Susloparov, I.M.; et al. An influenza A(H5N8) virus isolated during an outbreak at a poultry farm in Russia in 2017 has an N294S substitution in the neuraminidase and shows reduced susceptibility to oseltamivir. Antivir. Res. 2021, 191, 105079. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-H.; Kim, B.; Antigua, K.J.C.; Jeong, J.H.; Kim, C.I.; Choi, W.-S.; Oh, S.; Kim, C.H.; Kim, E.-G.; Choi, Y.K.; et al. Baloxavir-oseltamivir combination therapy inhibits the emergence of resistant substitutions in influenza A virus PA gene in a mouse model. Antivir. Res. 2021, 193, 105126. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Influenza Virus Strains | Mean EC90 (nmol/L) ± SD | ||
|---|---|---|---|
| Baloxavir Acid | Oseltamivir Acid | Favipiravir | |
| A/ruddy turnstone/Delaware/103/2007 (H5N1) | 1.4 ± 1.3 | 12.8 ± 5.7 | 16,927.4 ± 12,375.0 |
| A/muscovy duck/Vietnam/OIE-559/2011 (H5N1) | 1.5 ± 0.3 | 20.7 ± 10.9 | 11,689.6 ± 7333.8 |
| A/whooper swan/Mongolia/2/2006 (H5N1) | 0.9 ± 0.5 | 14.9 ± 1.6 | 13,812 ± 11,056.8 |
| A/black swan/Akita/1/2016 (H5N6) | 0.8 ± 0.5 | 20 ± 9.7 | 50,156.1 ± 69,943.5 |
| A/northern pintail/Hokkaido/M13/2020 (H5N8) | 1.3 ± 1.0 | 19.5 ± 11.9 | 16,025.5 ± 10,164.5 |
| A/whooper swan/Fukushima/0701B002/2021 (H5N8) | 1.1 ± 1.0 | 19.2 ± 13.9 | 16,052.1 ± 12,507.3 |
| A/whooper swan/Miyagi/0402B001/2021 (H5N8) | 0.7 ± 0.3 | 11.3 ± 2.8 | 6992.6 ± 1370.8 |
| rg-A/Hong Kong/483/1997 (H5N1) * | 1.6 ± 1.0 | 16.4 ± 11.4 | 26,948.7 ± 5081.5 |
| rg-A/Hong Kong/483/1997 NA-H274Y * | 3.2 ± 1.2 | 4054.9 ± 1295.7 | 31,129.5 ± 11,788.5 |
| rg-A/Hong Kong/483/1997 NA-N294S * | 1.7 ± 0.4 | 1291.2 ± 482.6 | 77,002.6 ± 2190.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taniguchi, K.; Ando, Y.; Kobayashi, M.; Toba, S.; Nobori, H.; Sanaki, T.; Noshi, T.; Kawai, M.; Yoshida, R.; Sato, A.; et al. Characterization of the In Vitro and In Vivo Efficacy of Baloxavir Marboxil against H5 Highly Pathogenic Avian Influenza Virus Infection. Viruses 2022, 14, 111. https://doi.org/10.3390/v14010111
Taniguchi K, Ando Y, Kobayashi M, Toba S, Nobori H, Sanaki T, Noshi T, Kawai M, Yoshida R, Sato A, et al. Characterization of the In Vitro and In Vivo Efficacy of Baloxavir Marboxil against H5 Highly Pathogenic Avian Influenza Virus Infection. Viruses. 2022; 14(1):111. https://doi.org/10.3390/v14010111
Chicago/Turabian StyleTaniguchi, Keiichi, Yoshinori Ando, Masanori Kobayashi, Shinsuke Toba, Haruaki Nobori, Takao Sanaki, Takeshi Noshi, Makoto Kawai, Ryu Yoshida, Akihiko Sato, and et al. 2022. "Characterization of the In Vitro and In Vivo Efficacy of Baloxavir Marboxil against H5 Highly Pathogenic Avian Influenza Virus Infection" Viruses 14, no. 1: 111. https://doi.org/10.3390/v14010111