DEF Cell-Derived Exosomal miR-148a-5p Promotes DTMUV Replication by Negative Regulating TLR3 Expression

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Approval and Consent to Participate
2.2. Reagents
2.3. Cell Lines, Birds, Virus, and Infection
2.4. RNA and cDNA Preparation
2.5. Quantitative Real-Time PCR (qRT-PCR)
2.6. Recombinant Plasmid Construction
2.7. TLR3-shRNA, MDA5-shRNA, and Generation of shRNA-Based Knockdown Cell Lines
2.8. Western Blot Analysis of TLR3, MDA5, and Exosome Proteins
2.9. Exosome isolation
2.10. PBMC Preparation
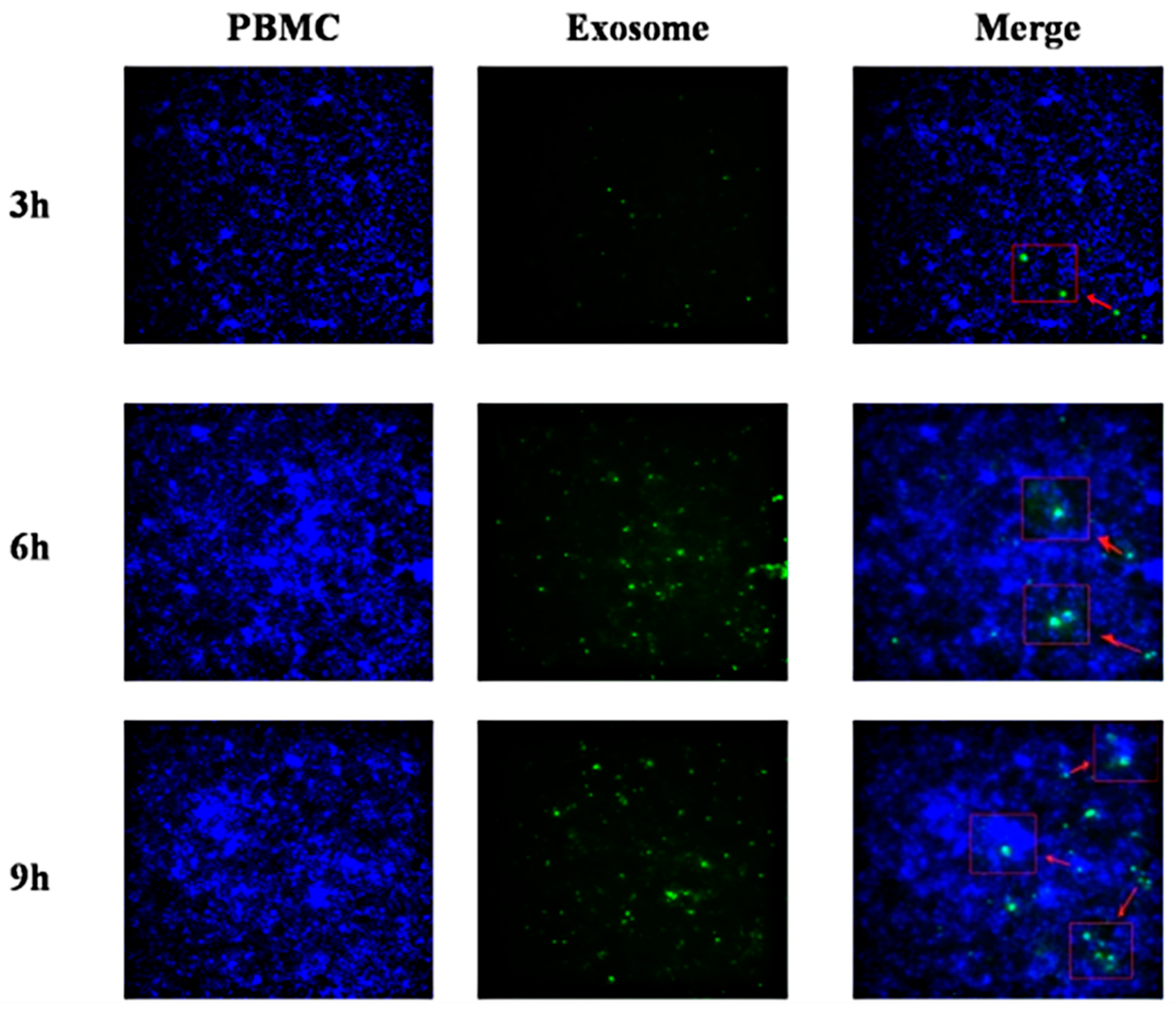
2.11. Interaction of Exosomes with PBMCs
2.12. Enzyme-Linked Immunosorbent Assay (ELISA) to Detect Duck TLR3 Protein
2.13. Dual-Luciferase Reporter Assay
2.14. Statistical Analysis
3. Results
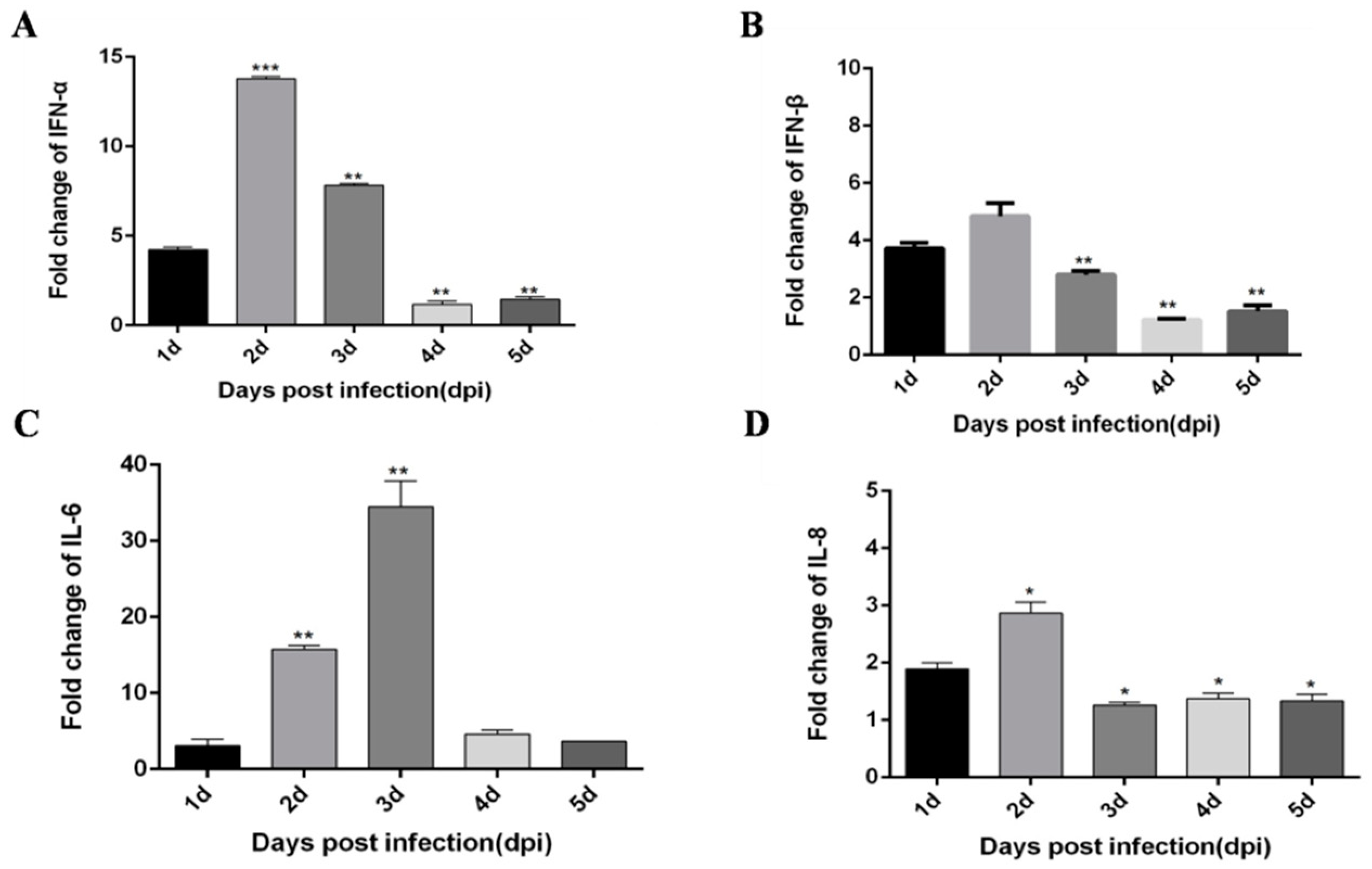
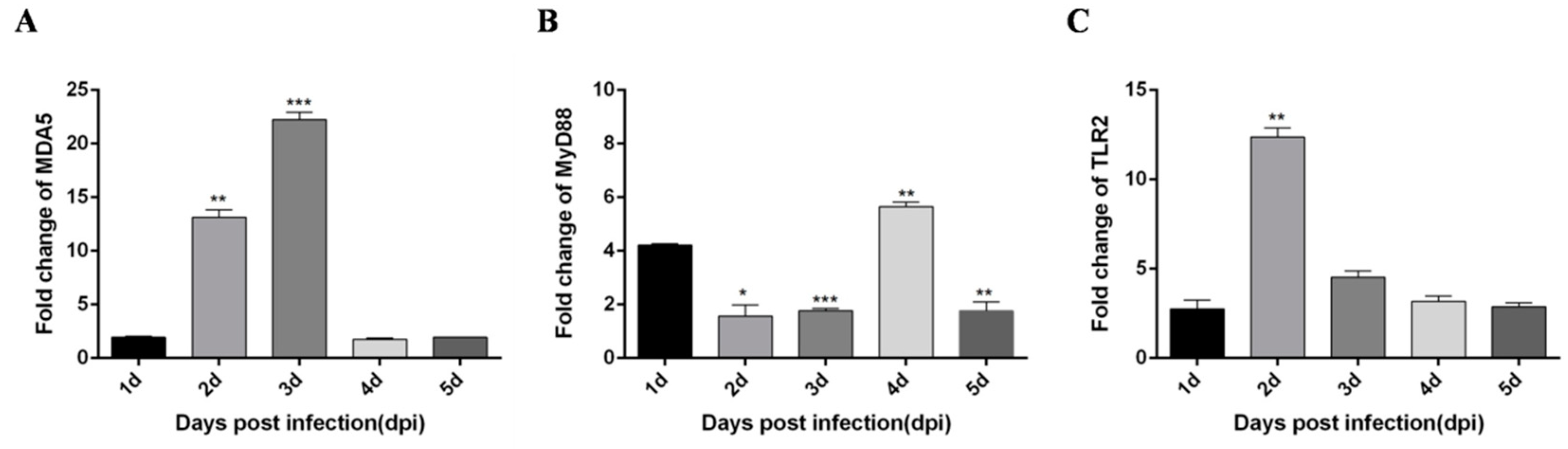
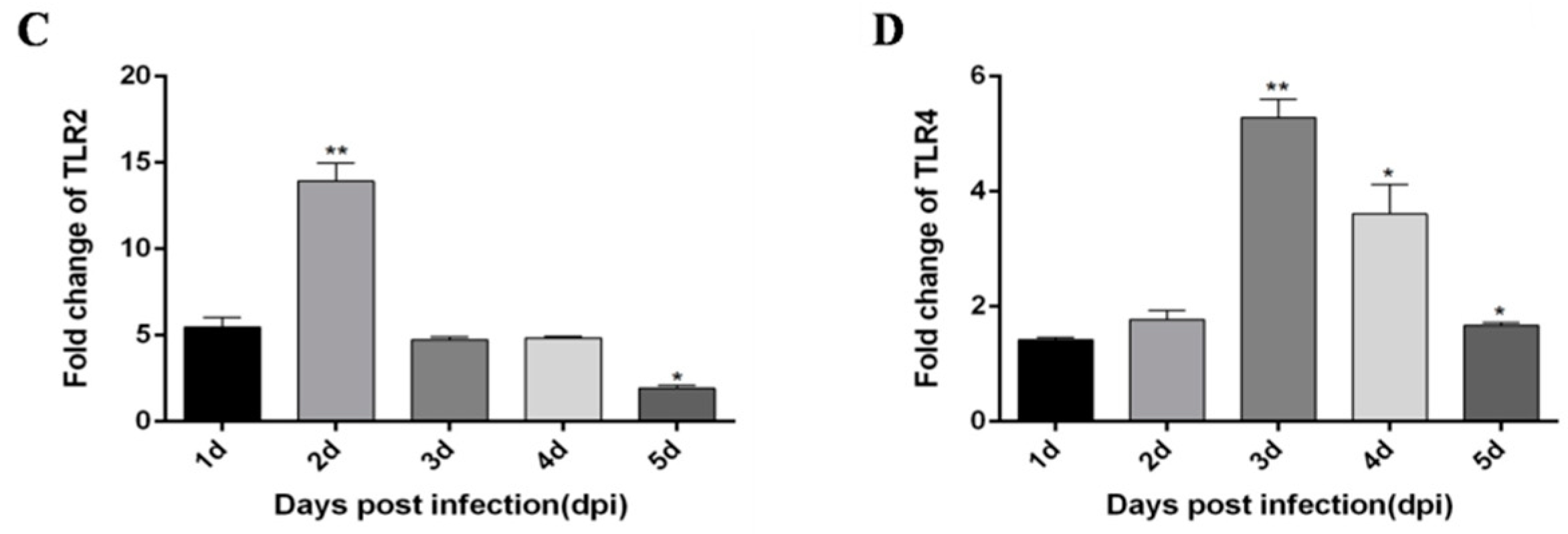
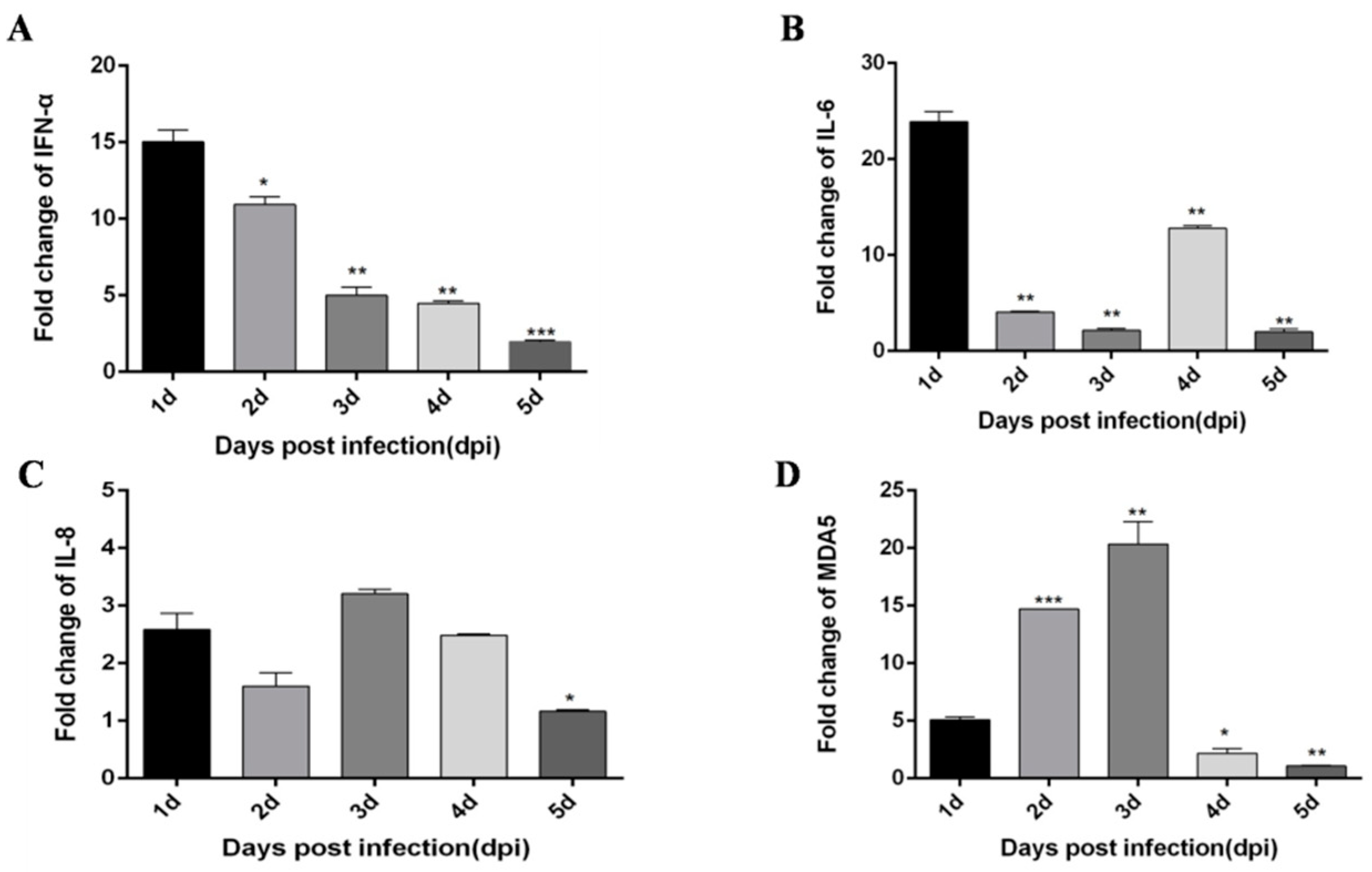
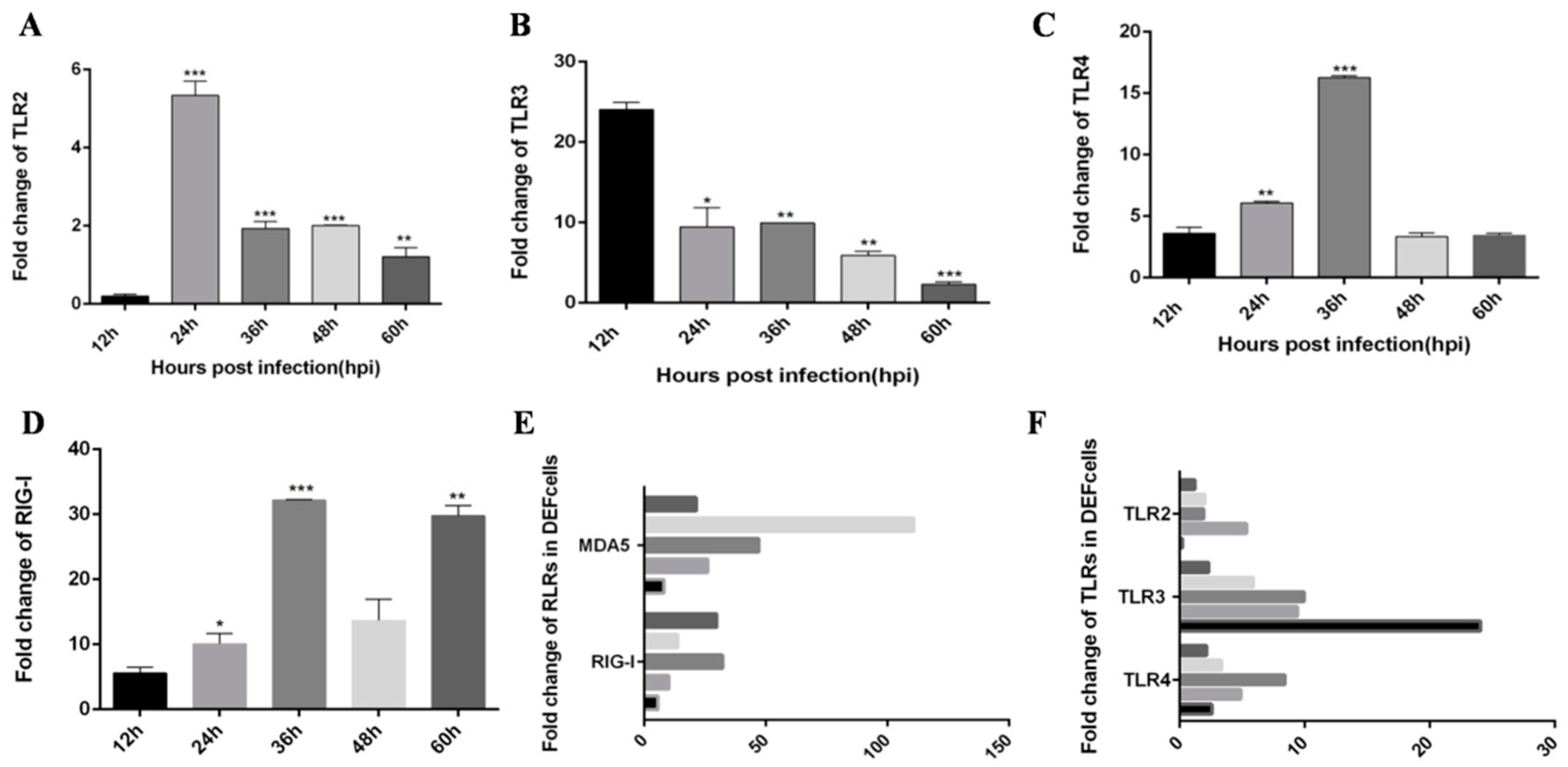
3.1. DTMUV-Infected Ducks Show Up-Regulated the Expression of TLR3, MDA5, and Inflammatory Cytokines
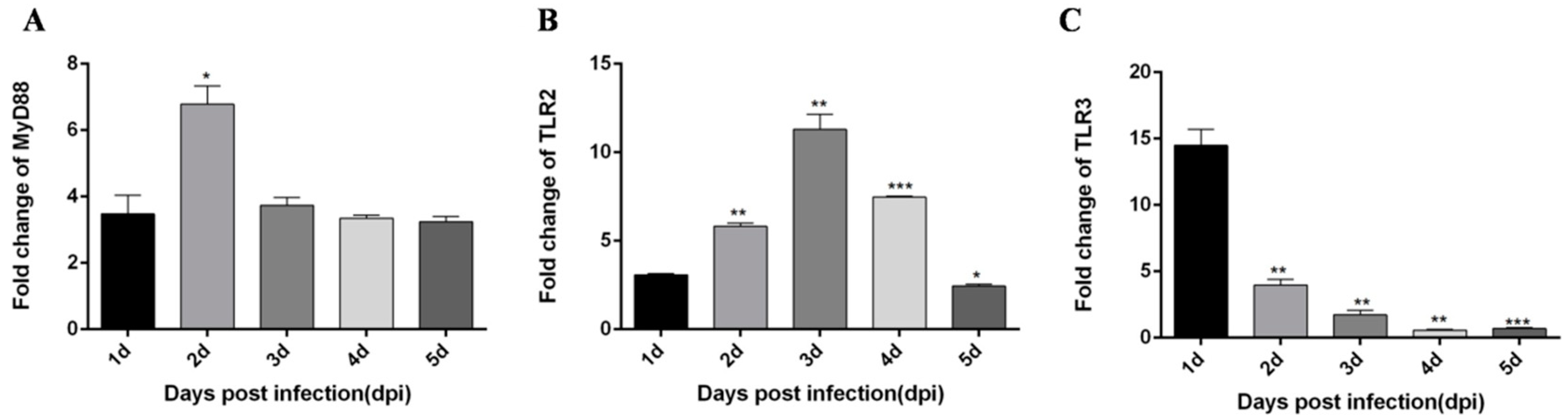
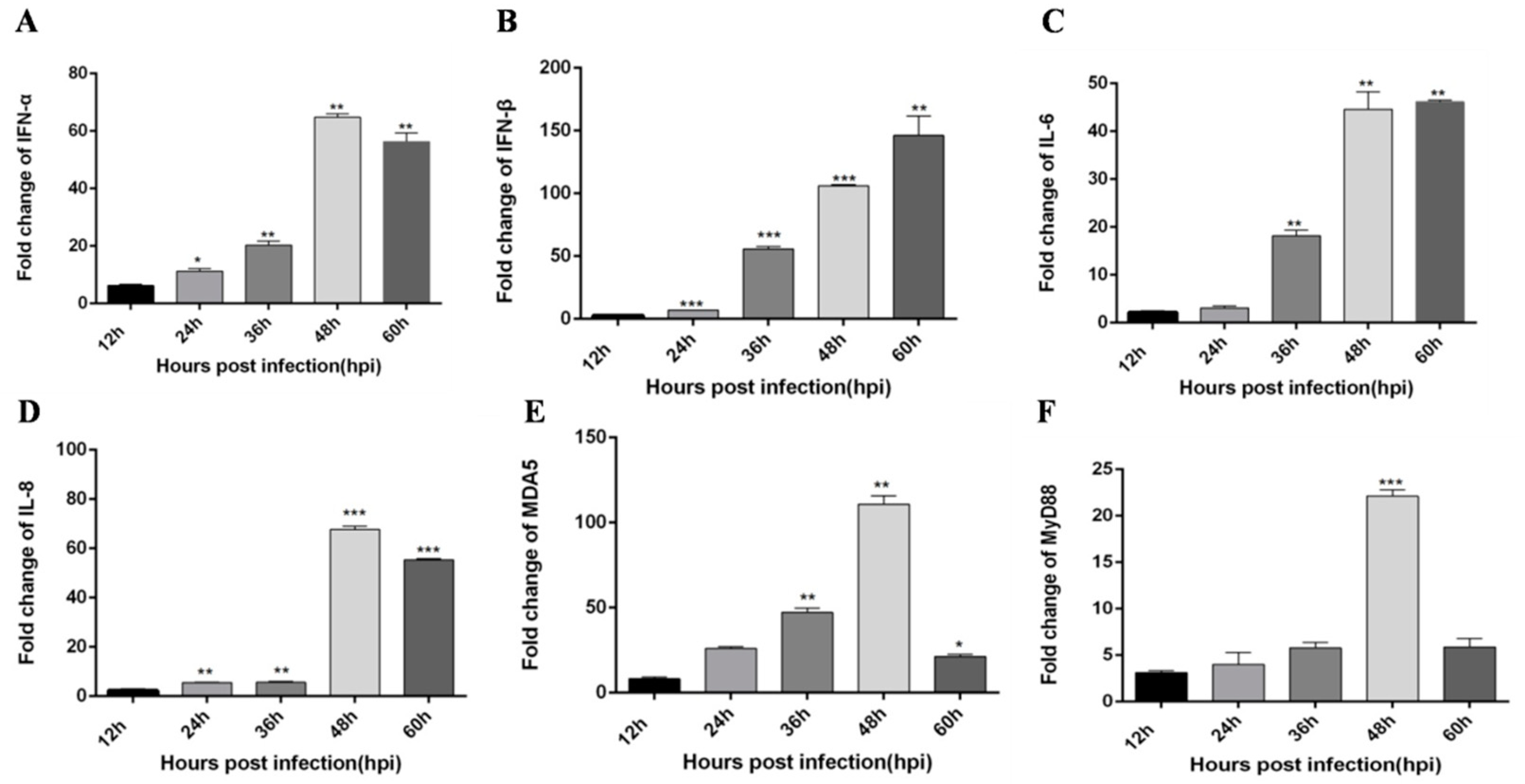
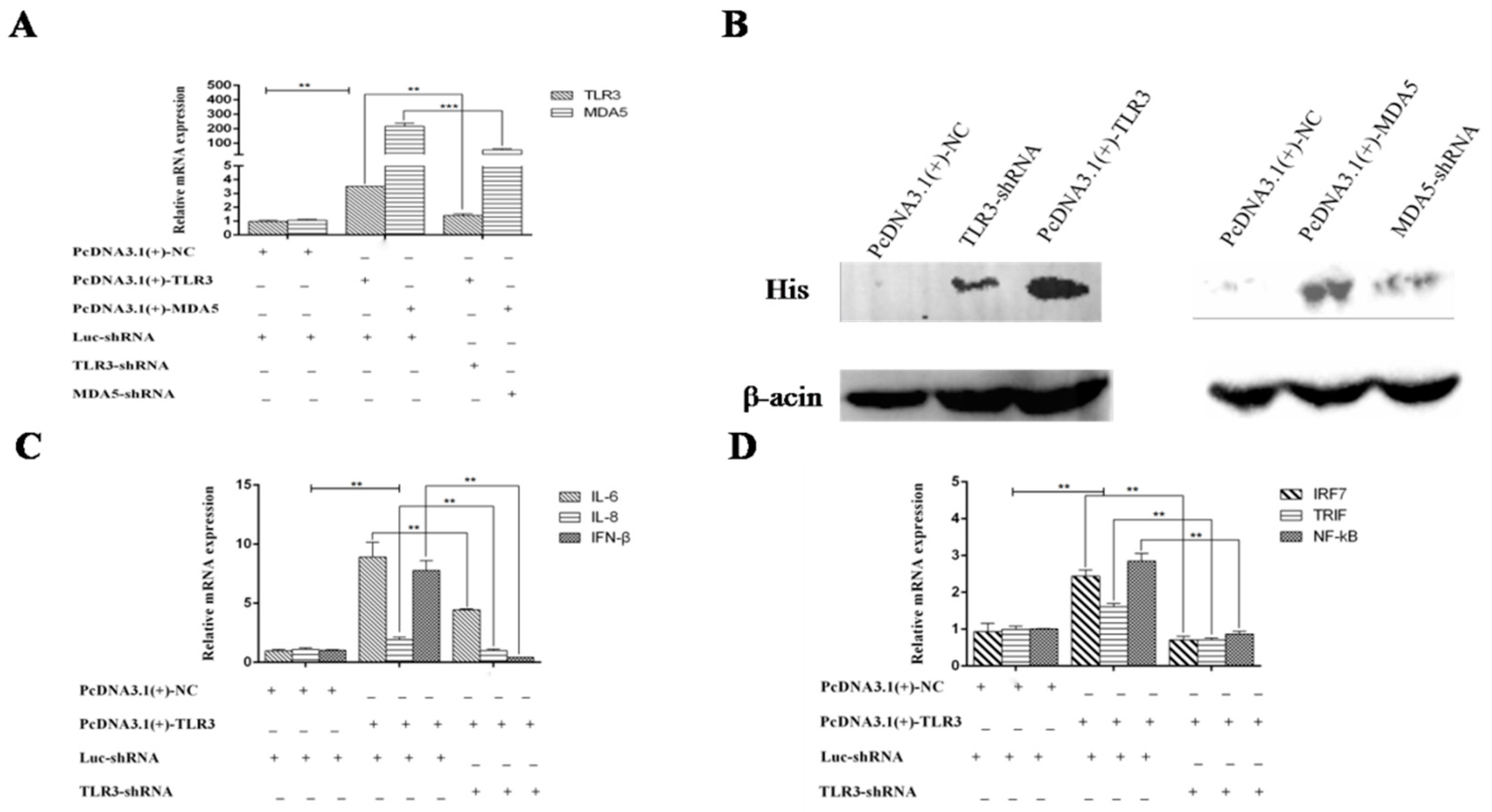
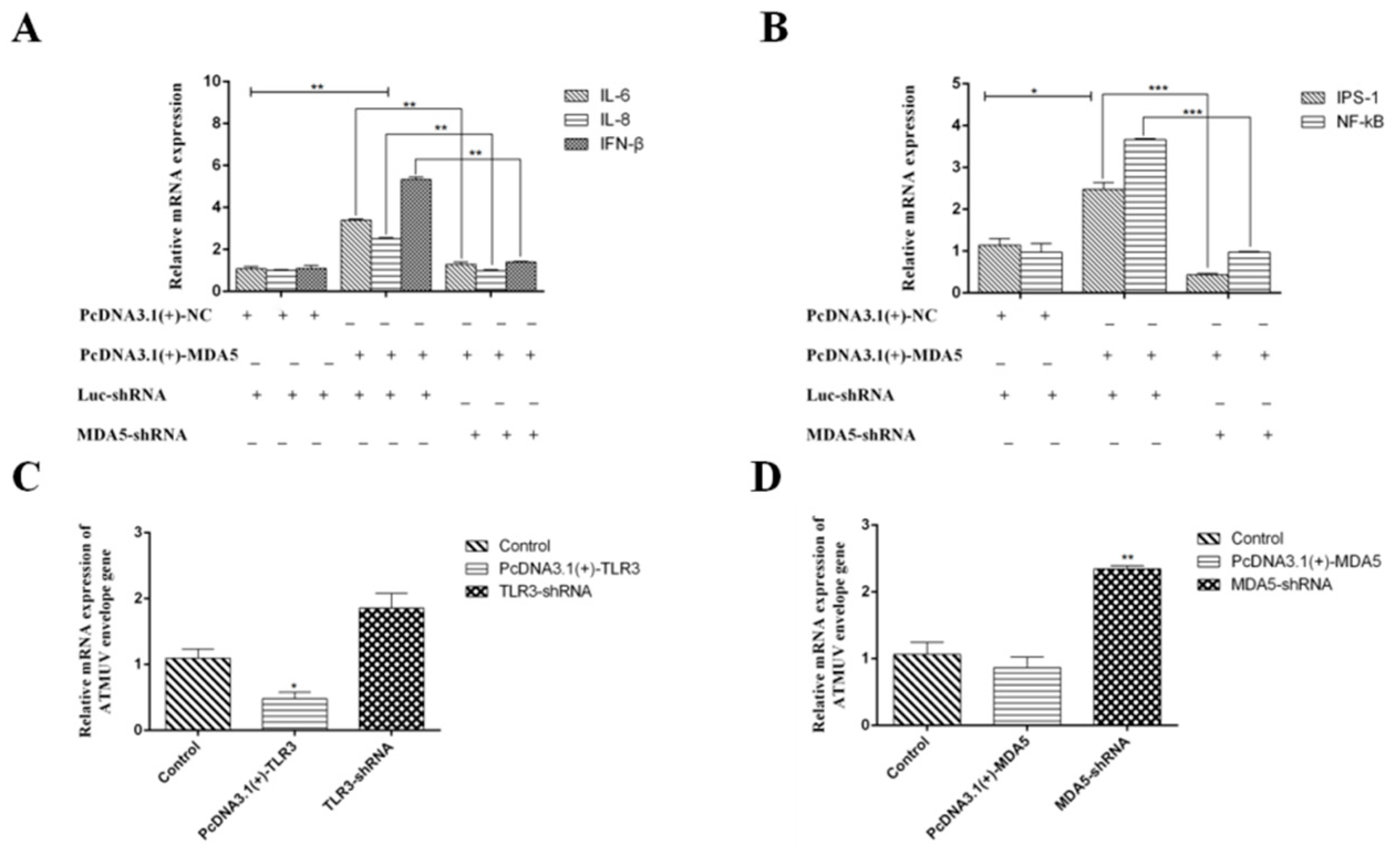
3.2. TLR3 and MDA5 Play Major Immunity Roles in DTMUV-Infected DEF Cells
3.3. MDA5 and TLR3-Dependent Signaling Pathways have Important Roles in Immunity During DTMUV Infection
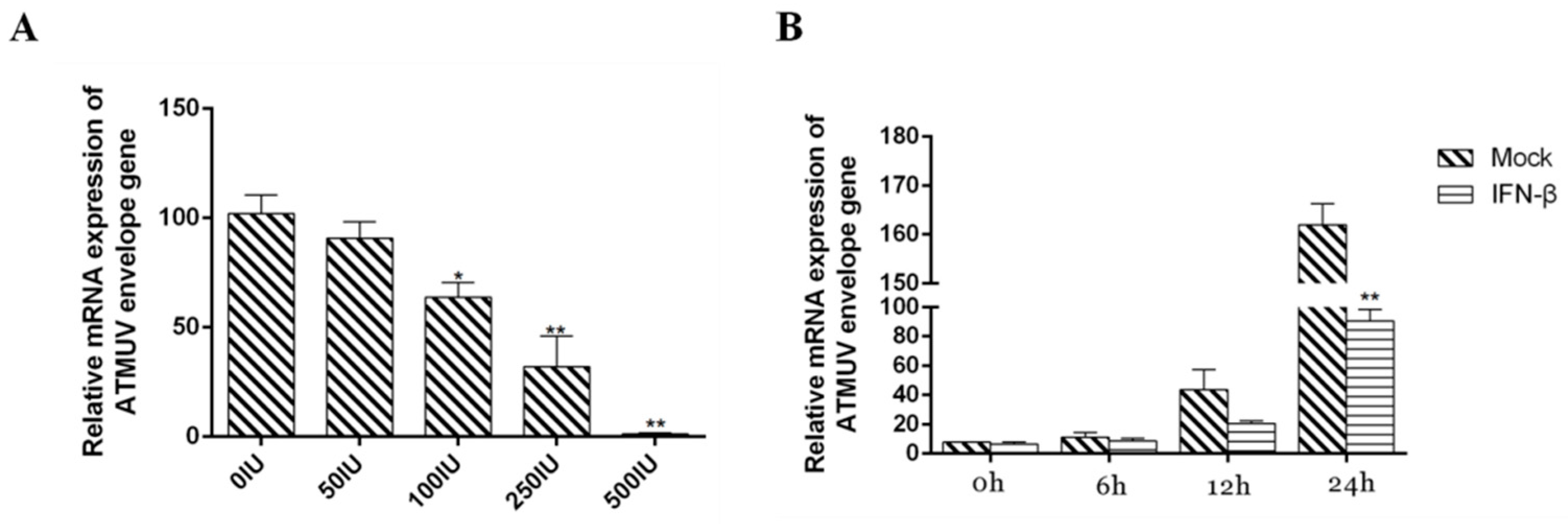
3.4. Type I IFN Significantly Impairs DTMUV Replication in Dose- and Time-Dependent Manners
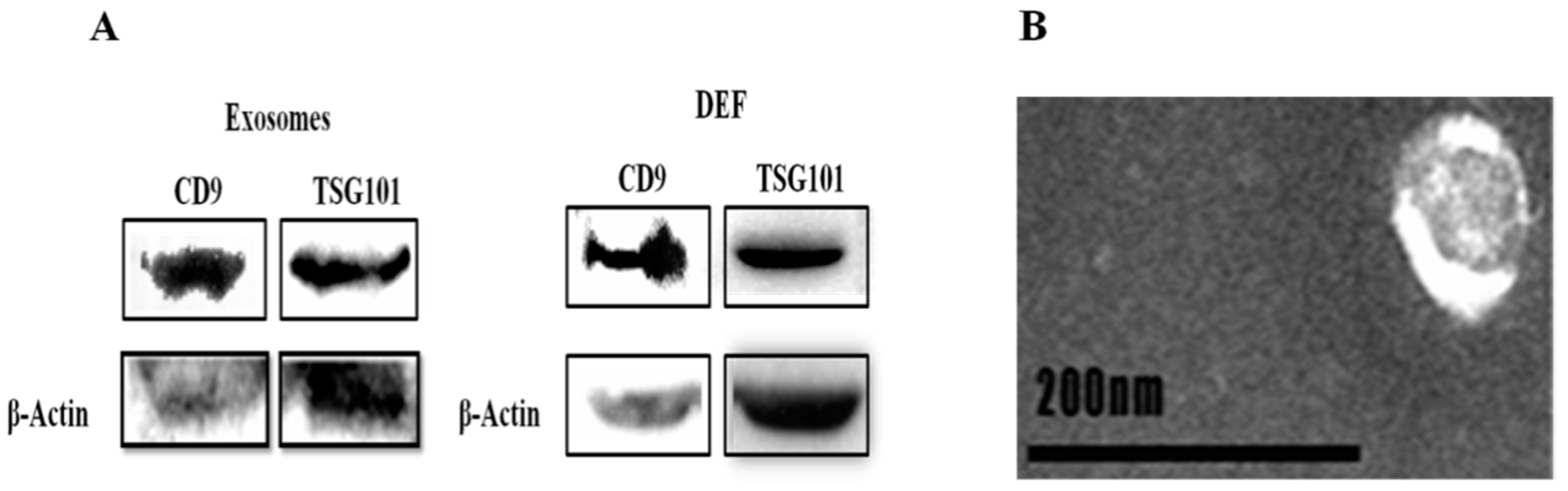
3.5. Morphological Characterization and Identification of Exosomes
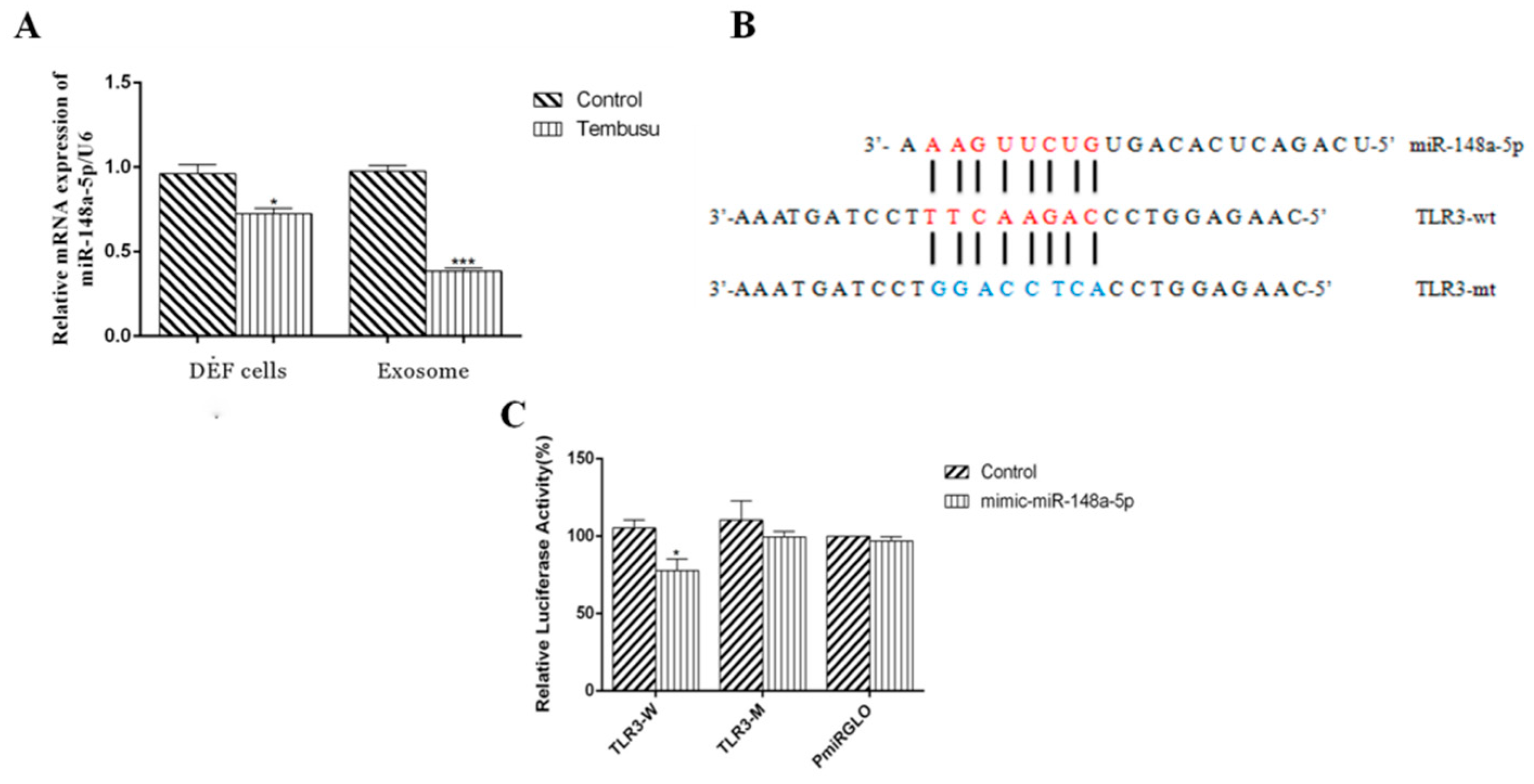
3.6. DTMUV-Infected DEF Cells and DEF-Derived Exosomes Show Decreased Levels of miR-148a-5p
3.7. miR-148a-5p Targets TLR3
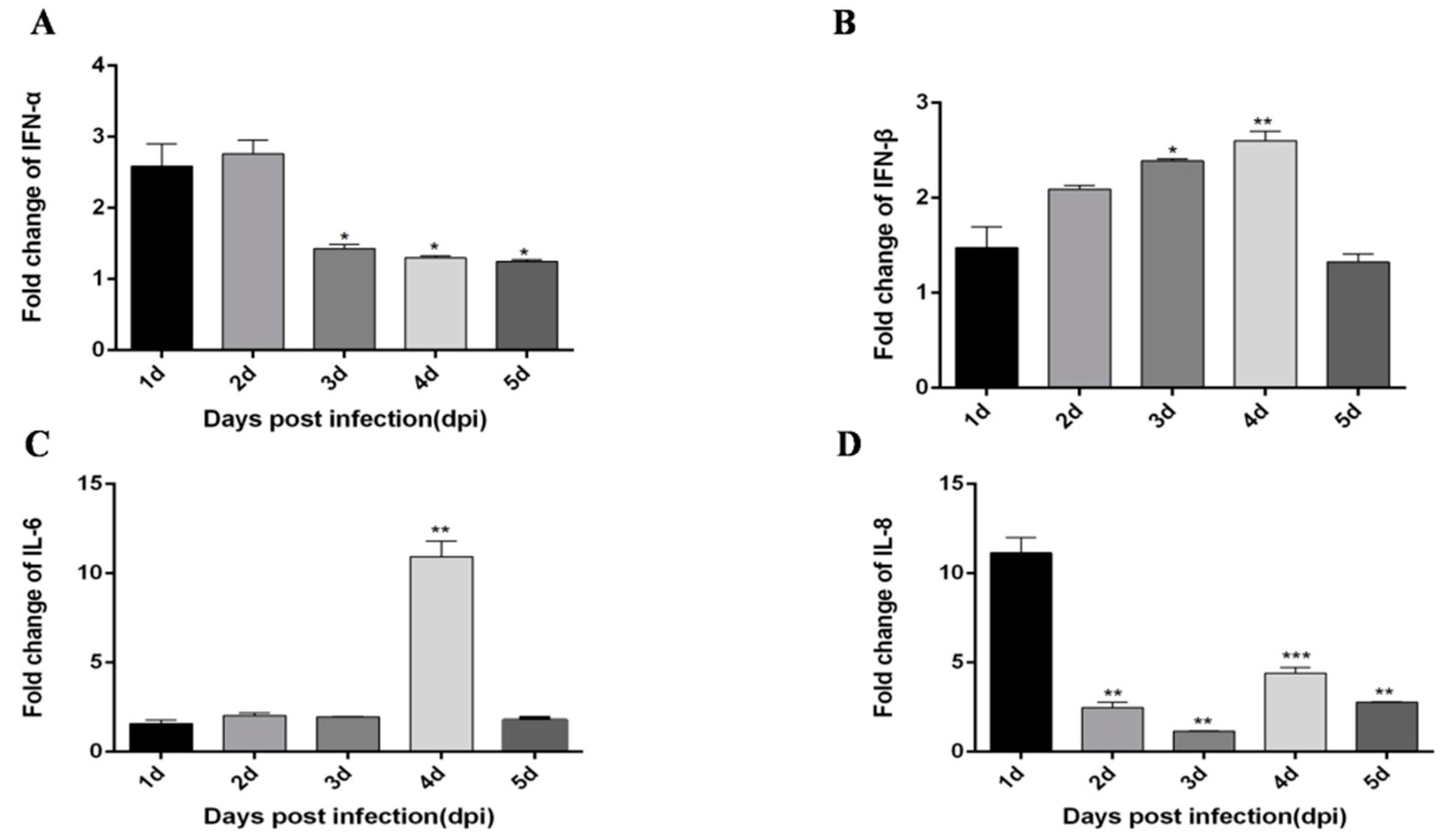
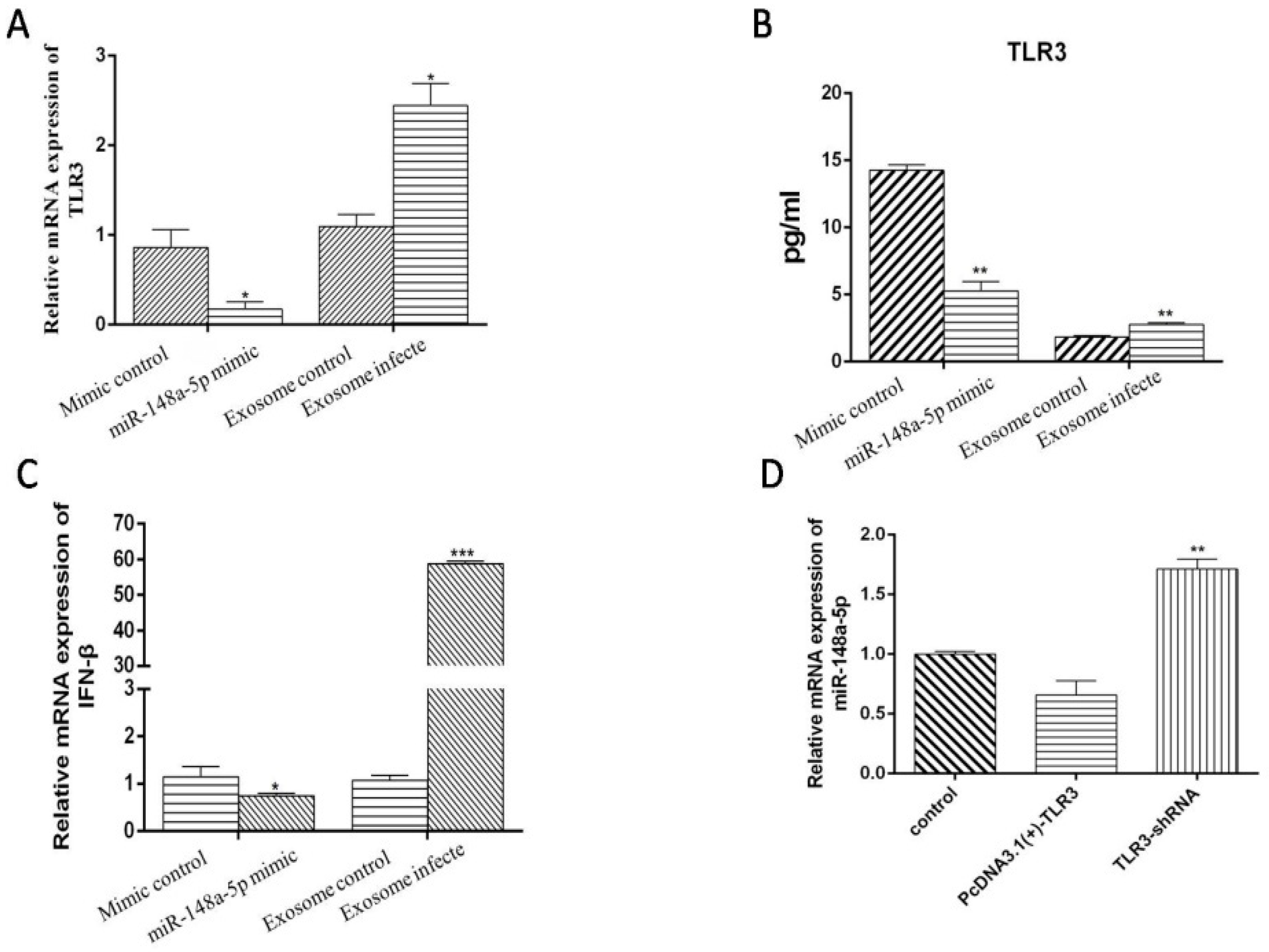
3.8. miR-148a-5p and Exo-miR-148a-5p Negatively Regulate TLR3 and IFN-β Expression in PBMCs
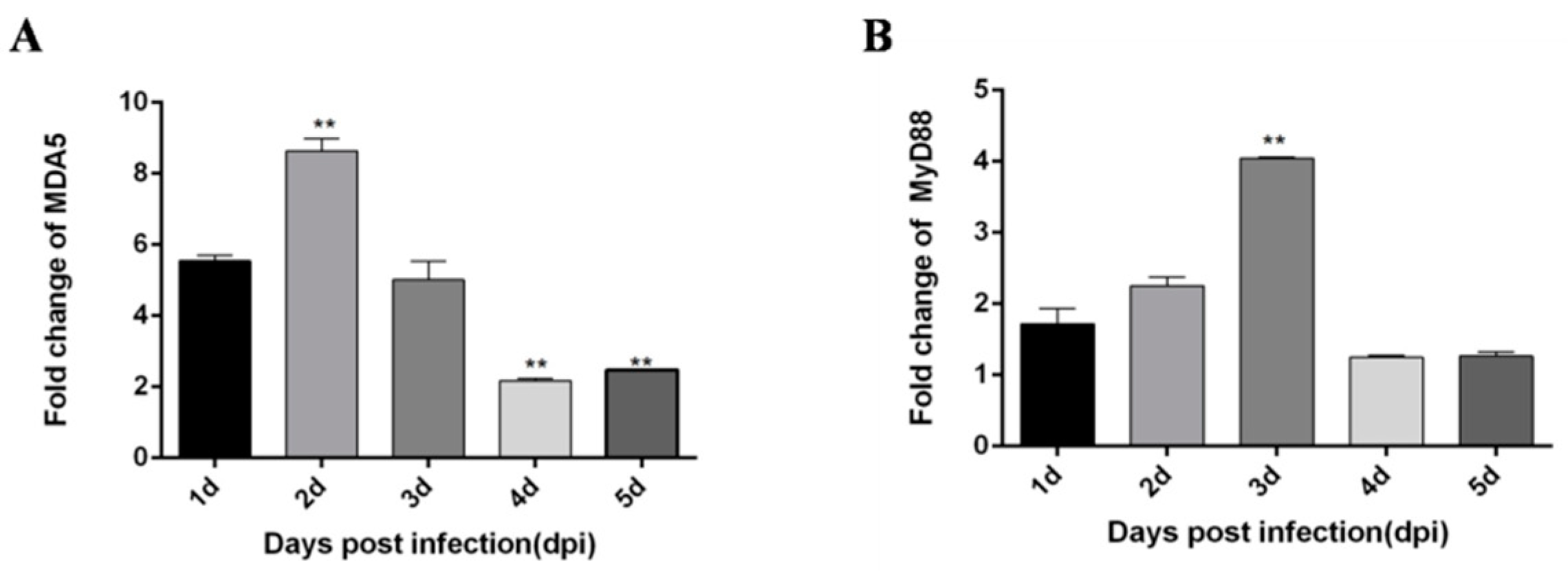
3.9. TLR3 May Be One of the Reasons for the Decrease of miR-148a-5p
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Liu, P.; Lu, H.; Li, S.; Wu, Y.; Gao, G.F.; Su, J. Duck egg drop syndrome virus: An emerging Tembusu-related flavivirus in China. China Life Sci. 2013, 56, 701–710. [Google Scholar] [CrossRef] [Green Version]
- Cao, Z.; Zhang, C.; Liu, Y.; Liu, Y.; Ye, W.; Han, J.; Ma, G.; Zhang, D.; Xu, F.; Gao, X.; et al. Tembusu virus in ducks, China. Emerg. Infect. Dis. 2011, 17, 1873–1875. [Google Scholar] [CrossRef]
- Su, J.; Li, S.; Hu, X.; Yu, X.; Wang, Y.; Liu, P.; Lu, X.; Zhang, G.; Hu, X.; Liu, D.; et al. Duck egg-drop syndrome caused by BYD virus, a new Tembusu-related flavivirus. PLoS ONE 2011, 6, e18106. [Google Scholar]
- Yan, P.; Zhao, Y.; Zhang, X.; Xu, D.; Dai, X.; Teng, Q.; Yan, L.; Zhou, J.; Ji, X.; Zhang, S.; et al. An infectious disease of ducks caused by a newly emerged Tembusu virus strain in mainland China. Virology 2011, 417, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Wang, A.; Chen, S.; Wu, Z.; Zhang, J.; Wang, M.; Jia, R.; Zhu, D.; Liu, M.; Yang, Q.; et al. Differential immune-related gene expression in the spleens of duck Tembusu virus-infected goslings. Vet. Microbiol. 2017, 212, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Fu, G.; Huang, Y.; Cheng, L.; Wan, C.; Shi, S.; Fu, Q.; Chen, H.; Lin, J.; Lin, F. Genome sequence and phylogenetic analysis of Tembusu viruses isolated from chicken. Chin. J. Vet. Sci. 2014, 34, 1418–1441. [Google Scholar]
- Tang, Y.; Diao, Y.; Yu, C.; Gao, X.; Ju, X.; Xue, C.; Liu, X.; Ge, P.; Qu, J.; Zhang, D.; et al. Characterization of a Tembusu virus isolated from naturally infected house sparrows (Passer domesticus) in Northern China. Transbound. Emerg. Dis. 2013, 60, 152–158. [Google Scholar] [CrossRef]
- Liu, P.; Lu, H.; Li, S.; Moureau, G.; Deng, Y.Q.; Wang, Y.; Zhang, L.; Jiang, T.; de Lamballerie, X.; Qin, C.F.; et al. Genomic and antigenic characterization of the newly emerging Chinese duck egg-drop syndrome flavivirus: Genomic comparison with Tembusu and Sitiawan viruses. J. Gen. Virol. 2012, 93, 2158–2170. [Google Scholar] [CrossRef] [Green Version]
- Pétrilli, V.; Dostert, C.; Muruve, D.A.; Tschopp, J. The inflammasome: A danger sensing complex triggering innate immunity. Curr. Opin. Immunol. 2007, 19, 615–622. [Google Scholar] [CrossRef]
- Brownlie, R.; Allan, B. Avian toll-like receptors. Cell Tissue Res. 2011, 343, 121–130. [Google Scholar] [CrossRef]
- Nasirudeen, A.M.; Wong, H.H.; Thien, P.; Xu, S.; Lam, K.P.; Liu, D.X. RIG-I, MDA5, and TLR3 synergistically play an important role in the restriction of dengue virus infection. PLoS Negl. Trop. Dis. 2011, 5, e926. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.W.; Choi, J.Y.; Uyangaa, E.; Kim, S.B.; Kim, J.H.; Kim, B.S.; Kim, K.; Eo, S.K. Distinct dictation of Japanese encephalitis virus-induced neuroinflammation and lethality via triggering TLR3 and TLR4 signal pathways. Cytokine 2014, 70, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, J.; Tiwari, S.K.; Lichinchi, G.; Yue, Q.; Patil, V.; Eroshkin, A.; Rana, T.M. Zikavirus depletes neural progenitors in human cerebral organoids through activation of the innate immune receptor TLR3. Cell Stem Cell 2016, 19, 258–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef]
- Temperley, N.D.; Berlin, S.; Paton, I.R.; Griffin, D.K.; Burt, D.W. Evolution of the chicken Toll-like receptor gene family: A story of gene gain and gene loss. BMC Genom. 2008, 9, 62. [Google Scholar] [CrossRef] [Green Version]
- Häcker, H.; Karin, M. Regulation and function of IKK and IKK-related kinases. Sci. STKE 2006, 357, 13. [Google Scholar] [CrossRef]
- Häcker, H.; Redecke, V.; Blagoev, B.; Kratchmarova, I.; Hsu, L.C.; Wang, G.G.; Kamps, M.P.; Raz, E.; Wagner, H.; Häcker, G.; et al. Specificity in Toll-like receptor signaling through distinct effector functions of TRAF3 and TRAF6. Nature 2006, 439, 204–207. [Google Scholar] [CrossRef] [Green Version]
- Bhoj, V.G.; Chen, Z.J. Ubiquitylation in innate and adaptive immunity. Nature 2009, 458, 430–437. [Google Scholar] [CrossRef]
- Kawagoe, T.; Sato, S.; Matsushita, K.; Kato, H.; Matsui, K.; Kumagai, Y.; Saitoh, T.; Kawai, T.; Takeuchi, O.; Akira, S. Sequential control of Toll-like receptor-dependent responses by IRAK1 and IRAK2. Nat. Immunol. 2008, 9, 684–691. [Google Scholar] [CrossRef]
- Kawai, T.; Takahashi, K.; Sato, S.; Coban, C.; Kumar, H.; Kato, H.; Ishii, K.J.; Takeuchi, O.; Akira, S. IPS-1, an adaptor triggering RIG-I- and Mda5-mediated type I interferon induction. Nat. Immunol. 2005, 6, 981–988. [Google Scholar] [CrossRef]
- Chen, S.; Luo, G.; Yang, Z.; Lin, S.; Chen, S.; Wang, S.; Goraya, M.U.; Chi, X.; Zeng, X.; Chen, J.-L. Avian Tembusu virus infection effectively triggers host innate immune response through MDA5 and TLR3-dependent signaling pathways. Vet. Res. 2016, 47, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, G.; Lin, Y.; Tang, Y.; Diao, Y. Comparative transcriptomic analysis of immune-related gene expression in duck embryo fibroblasts following duck Tembusu virus infection. Int. J. Mol. Sci. 2018, 19, 2328. [Google Scholar] [CrossRef] [Green Version]
- Vlassov, A.V.; Magdaleno, S.; Setterquist, R.; Conrad, R. Exosomes: Current knowledge of their composition, biological functions, and diagnostic and therapeutic potentials. Biochim. Biophys. Acta 2012, 1820, 940–948. [Google Scholar] [CrossRef] [PubMed]
- Brinton, L.T.; Sloane, H.S.; Kester, M.; Kelly, K.A. Formation and role of exosomes in cancer. Cell. Mol. Life Sci. 2015, 72, 659–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Chopp, M. Exosome therapy for stroke. Stroke 2018, 49, 1083–1090. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Yang, F.; Jiang, L.; Chen, Y.; Wang, K.; Xu, F.; Wei, Y.; Cao, X.; Wang, J.; Cai, Z. Exosomes with membrane-associated TGF-β1 from gene-modified dendritic cells inhibit murine EAE independently of MHC restriction. Eur. J. Immunol. 2013, 43, 2461–2472. [Google Scholar] [CrossRef]
- Cai, J.; Han, Y.; Ren, H.; Chen, C.; He, D.; Zhou, L.; Eisner, G.M.; Asico, L.D.; Jose, P.A.; Zeng, C. Extracellular vesicle-mediated transfer of donor genomic DNA to recipient cells is a novel mechanism for genetic influence between cells. J. Mol. Cell. Biol. 2013, 5, 227–238. [Google Scholar] [CrossRef] [Green Version]
- Valencia, K.; Luis-Ravelo, D.; Bovy, N.; Antón, I.; Martínez-Canarias, S.; Zandueta, C.; Ormazábal, C.; Struman, I.; Tabruyn, S.; Rebmann, V.; et al. miRNA cargo within exosome-like vesicle transfer influences metastatic bone colonization. Mol. Oncol. 2014, 8, 689–703. [Google Scholar] [CrossRef]
- Andre, F.; Schartz, N.E.; Movassagh, M.; Flament, C.; Pautier, P.; Morice, P.; Pomel, C.; Lhomme, C.; Escudier, B.; Le Chevalier, T.; et al. Malignant effusions and immunogenic tumour-derived exosomes. Lancet 2002, 360, 295–305. [Google Scholar] [CrossRef]
- Théry, C.; Zitvogel, L.; Amigorena, S. Exosomes: Composition, biogenesis, and function. Nat. Rev. Immunol. 2002, 2, 569–579. [Google Scholar] [CrossRef]
- Wang, K.; Liu, H.; He, Y.; Chen, T.; Yang, Y.; Niu, Y.; Chen, H.; Chen, Y.; Liu, J.; Ye, F.; et al. Correlation of TLR1-10 expression in peripheral blood mononuclear cells with chronic hepatitis B and chronic hepatitis B-related liver failure. Hum. Immunol. 2010, 71, 950–956. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, G.; Yao, Z.Q.; Moorman, J.P.; Ning, S. MicroRNA regulation of viral immunity, latency, and carcinogenesis of selected tumor viruses and HIV. Rev. Med. Virol. 2015, 25, 320–341. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Huang, J.; Jia, R.; Zhang, B.; Wang, M.; Zhu, D.; Chen, S.; Liu, M.; Yin, Z.; Cheng, A. Identification and molecular characterization of a novel duck Tembusu virus isolate from Southwest China. Arch. Virol. 2015, 160, 2781–2790. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Li, N.; Zhang, J.; Wang, Y.; Liu, J.; Cai, Y.; Chai, T.; Wei, L. Expression of immune-related genes of ducks infected with Avian Pathogenic Escherichia coli (APEC). Front. Microbiol. 2016, 7, 637. [Google Scholar] [CrossRef] [Green Version]
- Song, K.; Chen, Z.; Huang, Y.; Li, C.; Meng, C.; Li, D.; Zhang, M.; Liu, G. Development of real-time fluorescence quantitative PCR assay for detection of duck Toll-like receptor 3. Chin. J. Anim. Infect. Dis. 2015, 3, 42–49. [Google Scholar]
- Li, N.; Wang, Y.; Li, R.; Liu, J.; Zhang, J.; Cai, Y.; Liu, S.; Chai, T.; Wei, L. Immune responses of ducks infected with duck Tembusu virus. Front. Microbiol. 2015, 6, 425. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Wang, T.; Liu, P.; Yang, C.; Wang, M.; Jia, R.; Zhu, D.; Liu, M.; Yang, Q.; Wu, Y.; et al. Duck interferon regulatory factor 7 (IRF7) can control duck Tembusu virus (DTMUV) infection by triggering type I interferon production and its signal transduction pathway. Cytokine 2019, 113, 31–38. [Google Scholar] [CrossRef]
- Cui, M.; Jia, R.; Huang, J.; Wu, X.; Hu, Z.; Zhang, X.; Wang, M.; Zhu, D.; Chen, S.; Liu, M.; et al. Analysis of the microRNA expression profiles in DEF cells infected with duck Tembusu virus. Infect. Genet. Evol. 2018, 63, 126–134. [Google Scholar] [CrossRef]
- Palma-Ocampo, H.K.; Flores-Alonso, J.C.; Vallejo-Ruiz, V.; Reyes-Leyva, J.; Flores-Mendoza, L.; Herrera-Camacho, I.; Rosas-Murrieta, N.H.; Santos-López, G. Interferon lambda inhibits dengue virus replication in epithelial cells. Virol. J. 2015, 12, 150. [Google Scholar] [CrossRef] [Green Version]
- Pinto, S.; Cunha, C.; Barbosa, M.; Vaz, A.R.; Brites, D. Exosomes from NSC-34 cells transfected with hSOD1-G93A are enriched in miR-124 and drive alterations in microglia phenotype. Front. Neurosci. 2017, 11, 273. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Wang, A.; Xia, C.; Lin, Q.; Chen, C. A single nucleotide polymorphism in primary-microRNA-146a reduces the expression of mature microRNA-146a in patients with Alzheimer’s disease and is associated with the pathogenesis of Alzheimer’s disease. Mol. Med. Rep. 2015, 12, 4037–4042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.W.; King, K.; Tu, J.; Luster, A.D.; Shresta, S. The roles of IRF-3 and IRF-7 in innate antiviral immunity against dengue virus. J. Immunol. 2013, 191, 4194–4201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, Y.T.; Chang, S.Y.; Lee, C.N.; Kao, C.L. Human TLR3 recognizes dengue virus and modulates viral replication in vitro. Cell. Microbiol. 2009, 11, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Winkelmann, E.R.; Gorder, S.R.; Mason, P.W.; Milligan, G.N. TLR3-and MyD88-dependent signaling differentially influences the development of West Nile virus-specific B cell responses in mice following immunization with RepliVAX WN, a single-cycle flavivirus vaccine candidate. J. Virol. 2013, 87, 12090–12101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalrymple, N.A.; Cimica, V.; Mackow, E.R. Dengue virus NS proteins inhibit RIG-I/MAVS signaling by blocking TBK1/IRF3 phosphorylation: Dengue virus serotype 1 NS4A Is a unique interferon-regulating virulence determinant. mBio 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Wu, Z.; Zhang, J.; Wang, M.; Jia, R.; Zhu, D.; Liu, M.; Sun, K.; Yang, Q.; Wu, Y.; et al. Duck stimulator of interferon genes plays an important role in host anti-duck plague virus infection through an IFN-dependent signalling pathway. Cytokine 2017, 102, 191–199. [Google Scholar] [CrossRef]
- Zhou, M.; Chen, J.; Zhou, L.; Chen, W.; Ding, G.; Cao, L. Pancreatic cancer derived exosomes regulate the expression of TLR4 in dendritic cells via miR-203. Cell. Immunol. 2014, 292, 65–69. [Google Scholar] [CrossRef]
- Paschon, V.; Takada, S.H.; Sousa, E.; Raeisossadati, R.; Ulrich, H.; Kihara, A.H. Interplay between exosomes, microRNAs and Toll-like receptors in brain disorders. Mol. Neurobiol. 2016, 53, 1–13. [Google Scholar] [CrossRef]
- Li, X.; Wang, S.; Zhu, R.; Li, H.; Han, Q.; Zhao, R.C. Lung tumor exosomes induce a pro-inflammatory phenotype in mesenchymal stem cells via NFκB-TLR signaling pathway. J. Hematol. Oncol. 2016, 9, 42. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Lei, C.Q.; Ji, Y.; Zhou, H.; Ren, Y.; Peng, Q.; Zeng, Y.; Jia, Y.; Ge, J.; Zhong, B.; et al. Duck Tembusu Virus Nonstructural Protein 1 Antagonizes IFN-β Signaling Pathways by Targeting VISA. J. Immunol. 2016, 197, 4704–4713. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Zhang, W.; Wu, Y.Y.; Wang, T.; Wu, S.; Wang, M.; Jia, R.; Zhu, D.; Liu, M.; Zhao, X.; et al. Binding of the Duck Tembusu Virus Protease to STING Is Mediated by NS2B and Is Crucial for STING Cleavage and for Impaired Induction of IFN-β. J. Immunol. 2019, 203, 3374–3385. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, S.; Tang, Y.; Diao, Y. Screening of Duck Tembusu Virus NS3 Interacting Host Proteins and Identification of Its Specific Interplay Domains. Viruses 2019, 11, 740. [Google Scholar] [CrossRef] [PubMed] [Green Version]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Oligonucleotide Sequence (5′→3′) | Length | References |
|---|---|---|---|
| TLR2 | F1: TCCTTCATTCAGCACCAGGC R2: GAAAAACACAGCGCAGATCA | 171 bp | [34] |
| TLR3 | F2: ATGTCATGCAAACCTGACCA R2: CCAGGGTCTTGAAAGGATCA | 239 bp | [35] |
| TLR4 | F3: ATCTTTCAAGGTGCCACATCC R3: ACTGACCTACCGATTGGACAC | 194 bp | |
| MyD88 | F4: TTTACAGCATGAATCCCTTGGC R4: TGGGAGTGTAAAATCCTGGTGT | 184 bp | |
| IFNα | F5: TCAGCACCACATCCACCACCTT R5: GGTTCTGGAGGAAGTGTTGGAT | 131 bp | |
| IFNβ | F6: TCAGCAGTCCAAGCATCCCT R6: GGAAGTGTTGGATGCTCCTGAAGTA | 186 bp | |
| IL-6 | F7: GGTCCAGAACAACCTCAACCTCC R7: CGTTGCCAGATGCTTTGTGC | 202 bp | |
| IL-8 | F8: CGGCATCGGTGTTCTTATCT R8: CTGTCCAGTGCCTTCAGTTT | 147 bp | |
| MDA5 | F9: GCTACAGAAGATAGAAGTGTCA R9: CAGGATCAGATCTGGTTCAG | 120 bp | [36] |
| IPS-1 | F10: TGCGACCGCCTACAAATTCTAT | 139 bp | |
| R10: AGGGGTTTGGTAGAGGTCGTAG | |||
| NF-κB | F11: ATCAACCCTTTTAACGTGCCT | 142 bp | |
| R11: GGTTGGAAATCAAAGGAGGC | |||
| TRIF | F12: TCTACTCACTGCTGGCAAAGG | 129 bp | |
| R12: CAGCCAGGACGCAGTTTTGTG | |||
| IRF7 | F13: ACAACGCCAGGAAGGATGT | 120 bp | |
| R13: AGCGAAAGTTGGTCTTCCACT | |||
| RIG-I | F14: GCGGATAGAGGCAACAAT | 133 bp | |
| R14: AGTTATGCCTGCTGCTTT | |||
| DTMUV envelope | F15: AATGGCTGTGGCTTGTTTGG | 207 bp | [37] |
| R15: GGGCGTTATCACGAATCTA | |||
| U6 | F16: CTCGCTTCGGCACGACA | 73 bp | [38] |
| R16: GCGTGTCATCCTTGCGC | |||
| β-action | F17: CCCCATTGAACACGGTATTGTC R17: GGCTACATACATGGCTGGGG | 199 bp | [35] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, H.; Cheng, A.; Zhang, X.; Pan, Y.; Wang, M.; Huang, J.; Zhu, D.; Chen, S.; Liu, M.; Zhao, X.; et al. DEF Cell-Derived Exosomal miR-148a-5p Promotes DTMUV Replication by Negative Regulating TLR3 Expression. Viruses 2020, 12, 94. https://doi.org/10.3390/v12010094
Guo H, Cheng A, Zhang X, Pan Y, Wang M, Huang J, Zhu D, Chen S, Liu M, Zhao X, et al. DEF Cell-Derived Exosomal miR-148a-5p Promotes DTMUV Replication by Negative Regulating TLR3 Expression. Viruses. 2020; 12(1):94. https://doi.org/10.3390/v12010094
Chicago/Turabian StyleGuo, Hongyan, Anchun Cheng, Xingcui Zhang, YuHong Pan, Mingshu Wang, Juan Huang, Dekang Zhu, Shun Chen, Mafeng Liu, Xinxin Zhao, and et al. 2020. "DEF Cell-Derived Exosomal miR-148a-5p Promotes DTMUV Replication by Negative Regulating TLR3 Expression" Viruses 12, no. 1: 94. https://doi.org/10.3390/v12010094