
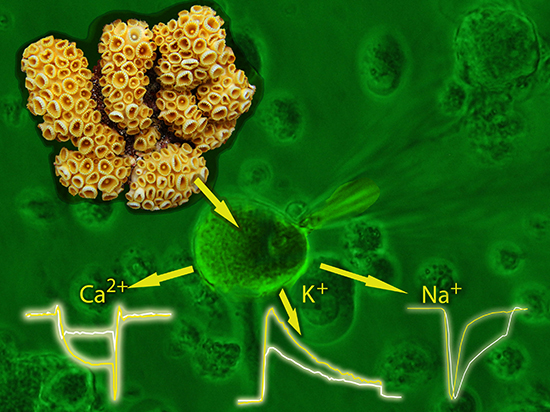
Activity of Palythoa caribaeorum Venom on Voltage-Gated Ion Channels in Mammalian Superior Cervical Ganglion Neurons


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
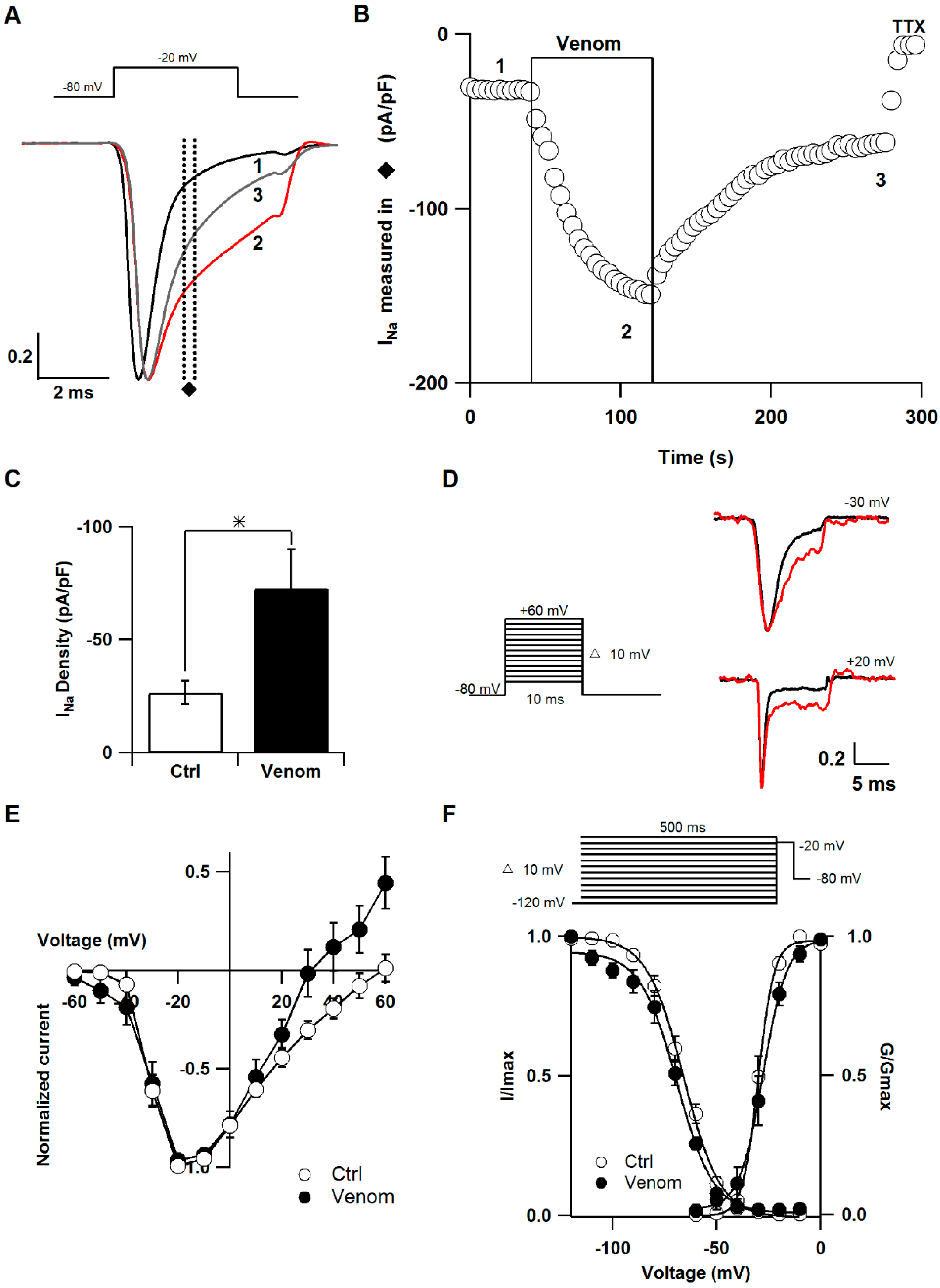
2.1. Effect of P. caribaeorum Venom on NaV1.7 Sodium Channel Current
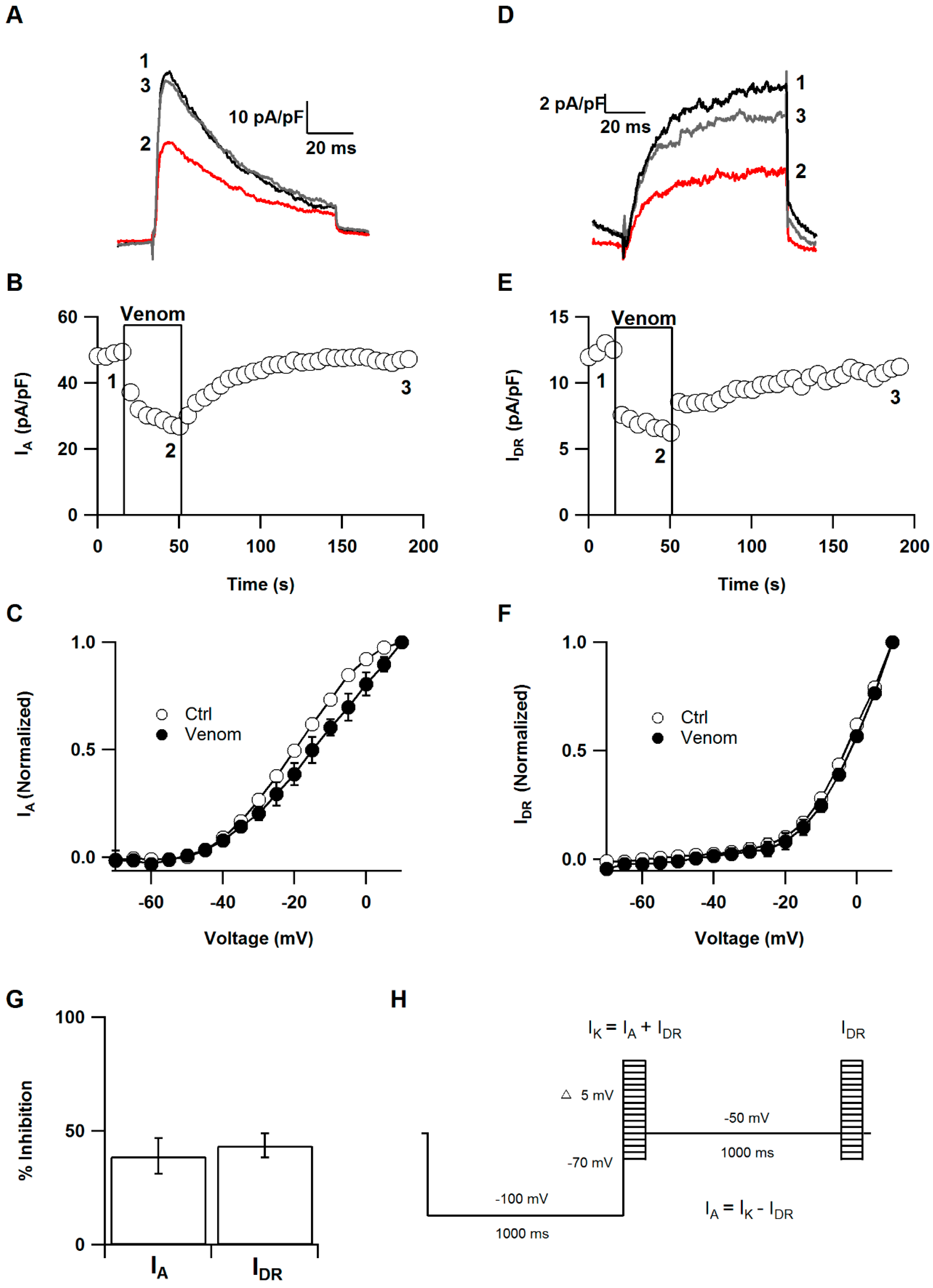
2.2. Effect of P. caribaeorum Venom on IA and IDR Potassium Currents
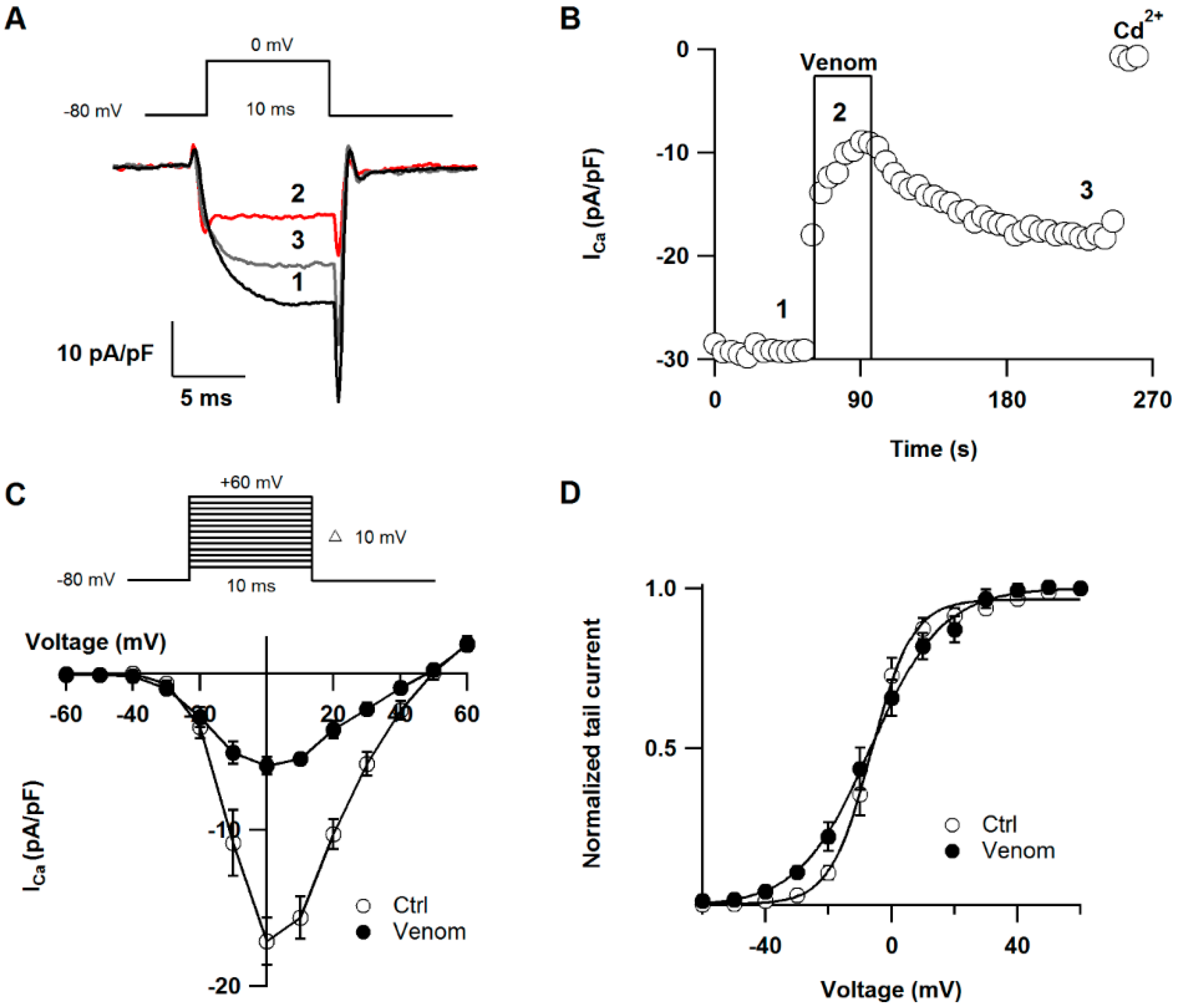
2.3. Effect of P. caribaeorum Venom on CaV2.2 Calcium Channel Current
3. Conclusions
4. Experimental Section
4.1. Venom Extraction
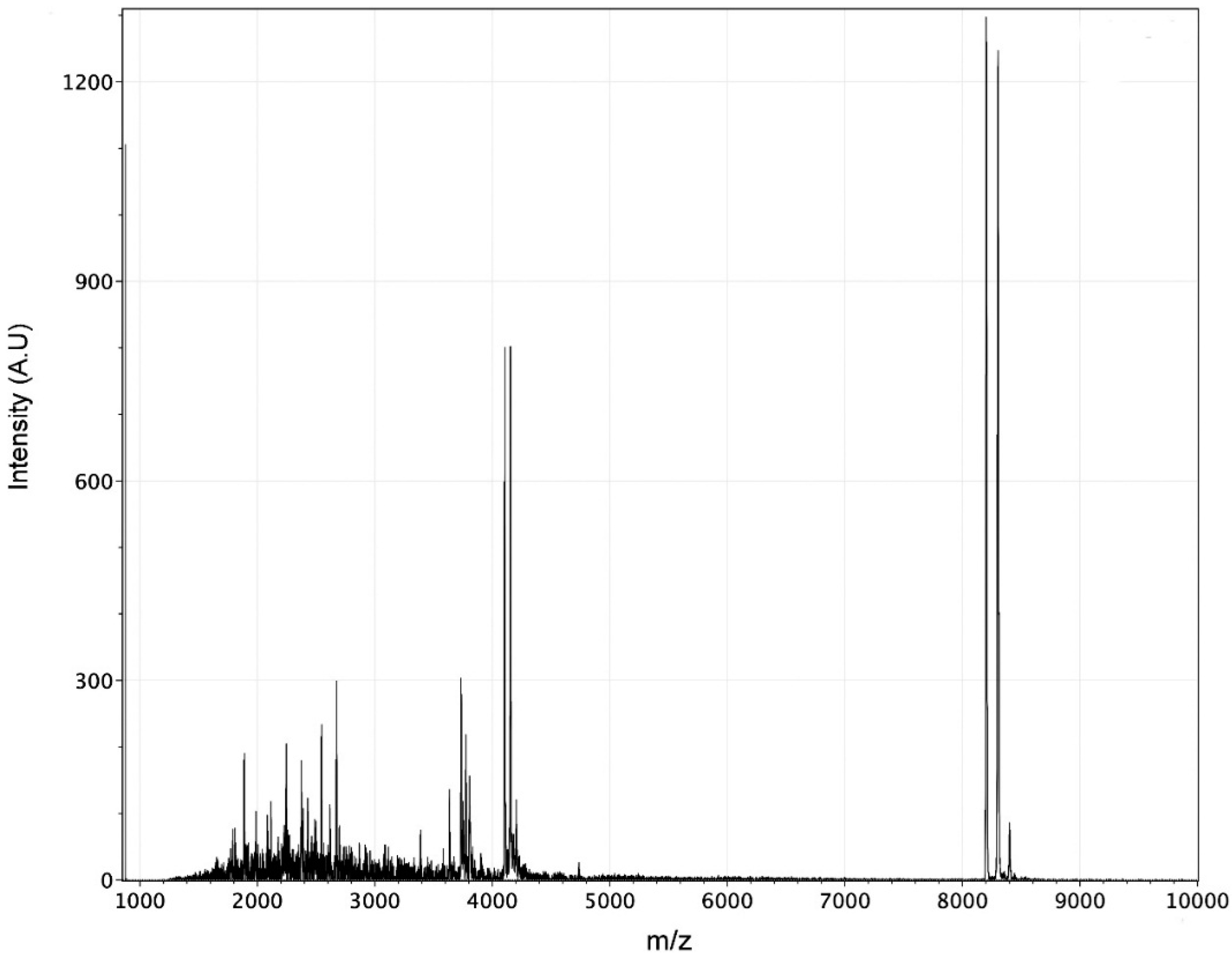
4.2. Mass Spectrometry Analysis
4.3. Superior Cervical Ganglion Neuron Culture
4.4. Electrophysiology Recording and Data Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Daly, M.; Brugler, M.R.; Cartwright, P.; Collins, A.G.; Dawson, M.N.; Fautin, D.G.; France, S.C.; McFadden, C.S.; Opresko, D.M.; Rodriguez, E.; et al. The phylum Cnidaria: A review of phylogenetic patterns and diversity 300 years after Linnaeus. Zootaxa 2007, 1668, 127–182. [Google Scholar]
- Parker, S.P. Synopsis and Classification of Living Organisms; McGraw-Hill: New York, NY, USA, 1982. [Google Scholar]
- Watters, M.R. Tropical marine neurotoxins: Venoms to drugs. Semin. Neurol. 2005, 25, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Shiomi, K. Novel peptide toxins recently isolated from sea anemones. Toxicon 2009, 54, 1112–1118. [Google Scholar] [CrossRef] [PubMed]
- Frazão, B.; Vasconcelos, V.; Antunes, A. Sea anemone (Cnidaria, Anthozoa, Actiniaria) toxins: An overview. Mar. Drugs 2012, 10, 1812–1851. [Google Scholar] [CrossRef] [PubMed]
- Mariottini, G.; Pane, L. The role of Cnidaria in drug discovery. A review on CNS implications and new perspectives. Recent Pat. CNS Drug Discov. 2013, 8, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Honma, T.; Shiomi, K. Peptide toxins in sea anemones: Structural and functional aspects. Mar. Biotechnol. 2006, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lazcano-Pérez, F.; Hernández-Guzman, U.; Sánchez-Rodríguez, J.; Arreguín-Espinosa, R. Cnidarian neurotoxic peptides affecting central nervous system targets. In Cent. Nerv. Syst. Agents Med. Chem.; 2015. Available online: http://www.ncbi.nlm.nih.gov/pubmed/26201531 (accessed on 2 May 2016). [Google Scholar]
- Sinniger, F.; Montoya-Burgos, J.I.; Chevaldonné, P.; Pawlowski, J. Phylogeny of the order Zoantharia (Anthozoa, Hexacorallia) based on the mitochondrial ribosomal genes. Mar. Biol. 2005, 147, 1121–1128. [Google Scholar] [CrossRef]
- Lakshmi, V.; Saxena, A.; Pandey, K.; Bajpai, P.; Misra-Bhattacharya, S. Antifilarial activity of Zoanthus species (Phylum Coelenterata, Class Anthzoa) against human lymphatic filaria, Brugia malayi. Parasitol. Res. 2004, 93, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Alencar, D.B.; Melo, A.A.; Silva, G.C.; Lima, R.L.; Pires-Cavalcante, K.M.S.; Carneiro, R.F.; Rabelo, A.S.; Sousa, O.V.; Vieira, R.H.S.F.; Viana, F.A.; et al. Antioxidant, hemolytic, antimicrobial, and cytotoxic activities of the tropical Atlantic marine zoanthid Palythoa caribaeorum. An. Acad. Bras. Ciênc. 2015, 87, 1113–1123. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Pérez, D.; Diaz-Garcia, C.M.; García-Delgado, N.; Sierra-Gómez, Y.; Castañeda, O.; Antunes, A. Insights into the toxicological properties of a low molecular weight fraction from Zoanthus sociatus (Cnidaria). Mar. Drugs 2013, 11, 2873–2881. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Garcia, C.M.; Sanchez-Soto, C.; Fuentes-Silva, D.; Leon-Pinzon, C.; Dominguez-Perez, D.; Varela, C.; Rodriguez-Romero, A.; Castañeda, O.; Hiriart, M. Low molecular weight compounds from Zoanthus sociatus impair insulin secretion via Ca+2 influx blockade and cause glucose intolerance in vivo. Toxicon 2012, 59, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Lazcano-Pérez, F.; Vivas, O.; Román-González, S.A.; Rodríguez-Bustamante, E.; Castro, H.; Arenas, I.; García, D.E.; Sánchez-Puig, N.; Arreguín-Espinosa, R. A purified Palythoa venom fraction delays sodium current inactivation in sympathetic neurons. Toxicon 2014, 82, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Béress, L.; Béress, R.; Wunderer, G. Purification of three polypeptides with neuroand cardiotoxic activity from the sea anemone Anemonia sulcata. Toxicon 1975, 13, 359–364. [Google Scholar] [CrossRef]
- Toledo-Aral, J.J.; Moss, B.L.; He, Z.J.; Koszowski, A.G.; Whisenand, T.; Levinson, S.R.; Wolf, J.J.; Silos-Santiago, I.; Halegoua, S.; Mandel, G. Identification of PN1, a predominant voltage-dependent sodium channel expressed principally in peripheral neurons. Proc. Natl. Acad. Sci. USA 1997, 94, 1527–1532. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A.; Goldin, A.L.; Waxman, S.G. International union of pharmacology. XLVII. Nomenclature and structure-function relationships of voltage-gated sodium channels. Pharmacol. Rev. 2005, 57, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Jia, Y.; Liu, B.; Zhao, Z.; Jia, Q.; Liang, H.; Zhang, H. Genistein inhibits voltage-gated sodium currents in SCG neurons through protein tyrosine kinase-dependent and kinase-independent mechanisms. Pflug. Arch. Eur. J. Physiol. 2008, 456, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Norton, R.S. Structure and structure-function relationships of sea anemone proteins that interact with the sodium channel. Toxicon 1991, 29, 1051–1084. [Google Scholar] [CrossRef]
- Kehl, S.J. Voltage-clamp analysis of the voltage-gated sodium current of the rat pituitary melanotroph. Neurosci. Lett. 1994, 165, 67–70. [Google Scholar] [CrossRef]
- Hernández-Ochoa, E.O.; García-Ferreiro, R.E.; García, D.E. G protein activation inhibits gating charge movement in rat sympathetic neurons. Am. J. Physiol. Cell Physiol. 2007, 292, C2226–C2238. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A.; Cestèle, S.; Yarov-Yarovoy, V.; Yu, F.H.; Konoki, K.; Scheuer, T. Voltage-gated ion channels and gating modifier toxins. Toxicon 2007, 49, 124–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schofield, G.G.; Ikeda, S.R. Potassium currents of acutely isolated adult rat superior cervical ganglion neurons. Brain Res. 1989, 485, 205–214. [Google Scholar] [CrossRef]
- Cai, S.-Q.; Li, W.; Sesti, F. Multiple modes of A-type potassium current regulation. Curr. Pharm. Des. 2007, 13, 3178–3184. [Google Scholar] [CrossRef] [PubMed]
- Blaine, J.T.; Ribera, A.B. Kv2 channels form delayed-rectifier potassium channels In Situ. J. Neurosci. 2001, 21, 1473–1480. [Google Scholar] [PubMed]
- Malin, S.A.; Nerbonne, J.M. Elimination of the fast transient in superior cervical ganglion neurons with expression of KV4.2W362F: Molecular dissection of/A. J. Neurosci. 2000, 20, 5191–5199. [Google Scholar] [PubMed]
- Liu, P.W.; Bean, B.P. Kv2 channel regulation of action potential repolarization and firing patterns in superior cervical ganglion neurons and hippocampal CA1 pyramidal neurons. J. Neurosci. 2014, 34, 4991–5002. [Google Scholar] [CrossRef] [PubMed]
- Diochot, S.; Schweitz, H.; Béress, L.; Lazdunski, M. Sea anemone peptides with a specific blocking activity against the fast inactivating potassium channel Kv3.4. J. Biol. Chem. 1998, 273, 6744–6749. [Google Scholar] [CrossRef] [PubMed]
- Yeung, S.Y.M.; Thompson, D.; Wang, Z.; Fedida, D.; Robertson, B. Modulation of Kv3 subfamily potassium currents by the sea anemone toxin BDS: Significance for CNS and biophysical studies. J. Neurosci. 2005, 25, 8735–8745. [Google Scholar] [CrossRef] [PubMed]
- Castañeda, O.; Harvey, A.L. Discovery and characterization of cnidarian peptide toxins that affect neuronal potassium ion channels. Toxicon 2009, 54, 1119–1124. [Google Scholar] [CrossRef] [PubMed]
- Kalman, K.; Pennington, M.W.; Lanigan, M.D.; Nguyen, A.; Rauer, H.; Mahnir, V.; Paschetto, K.; Kem, W.R.; Grissmer, S.; Gutman, G.A.; et al. ShK-Dap22, a Potent Kv1.3-specific Immunosuppressive Polypeptide. J. Biol. Chem. 1998, 273, 32697–32707. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.H.; Catterall, W.A. The VGL-chanome: A protein superfamily specialized for electrical signaling and ionic homeostasis. Sci. STKE Signal Transduct. Knowl. Environ. 2004, 2004. [Google Scholar] [CrossRef] [PubMed]
- Plummer, M.R.; Logothetis, D.E.; Hess, P. Elementary properties and pharmacological sensitivities of calcium channels in mammalian peripheral neurons. Neuron 1989, 2, 1453–1463. [Google Scholar] [CrossRef]
- McDonough, S.I. Gating modifier toxins of voltage-gated calcium channels. Toxicon 2007, 49, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Sousa, S.R.; Vetter, I.; Lewis, R.J. Venom peptides as a rich source of Cav2.2 channel blockers. Toxins 2013, 5, 286–314. [Google Scholar] [CrossRef] [PubMed]





© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lazcano-Pérez, F.; Castro, H.; Arenas, I.; García, D.E.; González-Muñoz, R.; Arreguín-Espinosa, R. Activity of Palythoa caribaeorum Venom on Voltage-Gated Ion Channels in Mammalian Superior Cervical Ganglion Neurons. Toxins 2016, 8, 135. https://doi.org/10.3390/toxins8050135
Lazcano-Pérez F, Castro H, Arenas I, García DE, González-Muñoz R, Arreguín-Espinosa R. Activity of Palythoa caribaeorum Venom on Voltage-Gated Ion Channels in Mammalian Superior Cervical Ganglion Neurons. Toxins. 2016; 8(5):135. https://doi.org/10.3390/toxins8050135
Chicago/Turabian StyleLazcano-Pérez, Fernando, Héctor Castro, Isabel Arenas, David E. García, Ricardo González-Muñoz, and Roberto Arreguín-Espinosa. 2016. "Activity of Palythoa caribaeorum Venom on Voltage-Gated Ion Channels in Mammalian Superior Cervical Ganglion Neurons" Toxins 8, no. 5: 135. https://doi.org/10.3390/toxins8050135