Passive and Active Vaccination Strategies to Prevent Ricin Poisoning

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
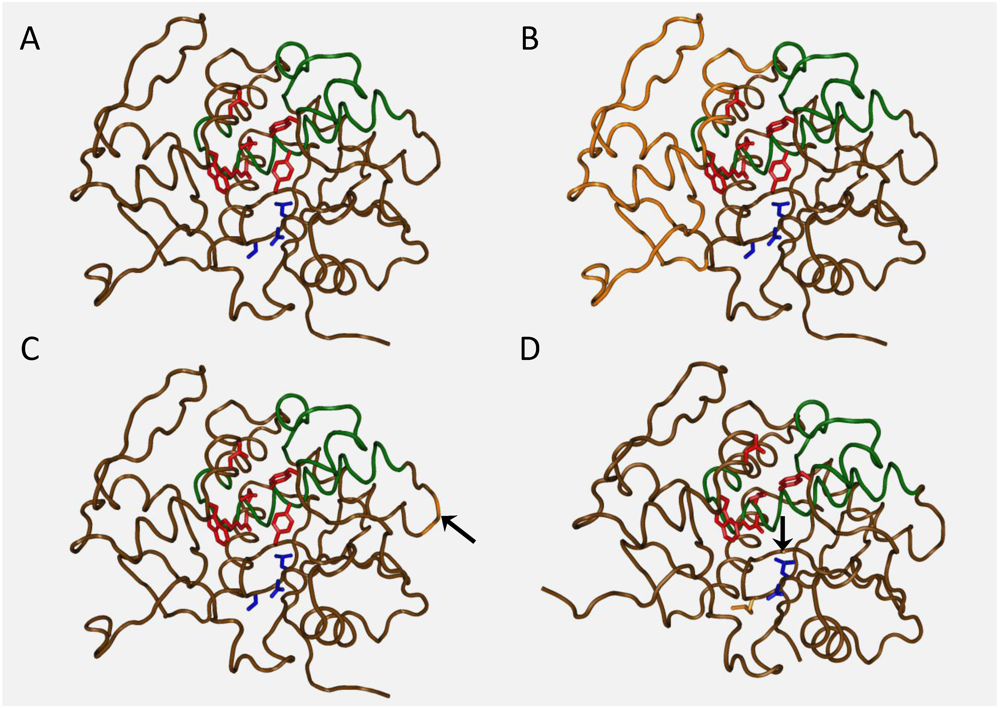
:1. Ricin Toxin
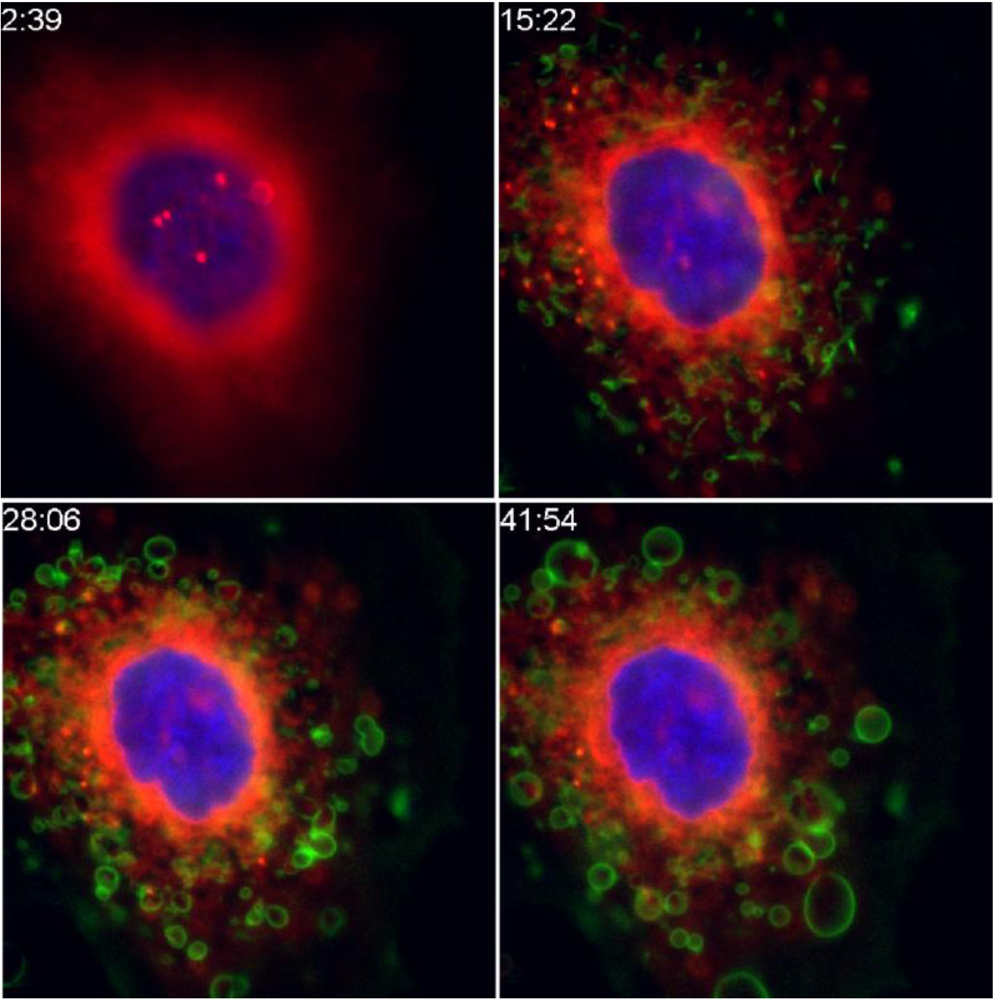
2. Cellular Toxicity of RT

3. Beneficial Uses of Ricin

4. Ricin as a Weapon of Bioterrorism
5. Routes of Exposure
5.1. Oral Exposure
5.2. Inhalation Exposure
5.3. Injection of RT
6. Protective Measures
6.1. Public Health Approaches to Containing Risk
6.2. Post-Exposure Therapies
6.3. Active and Passive Immunization
6.3.1. Passive Administration of Antibodies


6.3.2. Active Immunization
7. Future Considerations
Acknowledgements
References
- Pastan, I.; Hassan, R.; FitzGerald, D.J.; Kreitman, R.J. Immunotoxin treatment of cancer. Annu. Rev. Med. 2007, 58, 221–237. [Google Scholar]
- Audi, J.; Belson, M.; Patel, M.; Schier, J.; Osterloh, J. Ricin poisoning: A comprehensive review. J. Am. Med. Assoc. 2005, 294, 2342–2351. [Google Scholar]
- Holtermann, K. Response to a ricin incident: Guidelines for federal, state, and local public health and medical officials. Available online: http://www.bt.cdc.gov/agent/ricin/hp.asp (accessed on 7 September 2011).
- Bradberry, S.M.; Dickers, K.J.; Rice, P.; Griffiths, G.D.; Vale, J.A. Ricin poisoning. Toxicol. Rev. 2003, 22, 55–70. [Google Scholar]
- Franz, D.R.; Jaax, N.K. Chapter 32: Ricin Toxin. In Medical Aspects of Chemical and Biological Warfare; Sidell, F.R., Takafuji, E.T., Franz, D.R., Eds.; Office of the Surgeon General of the Army: Washington, DC, USA, 1997; Volume 3, pp. 631–642. [Google Scholar]
- Smallshaw, J.E.; Richardson, J.A.; Vitetta, E.S. Rivax, a recombinant ricin subunit vaccine, protects mice against ricin delivered by gavage or aerosol. Vaccine 2007, 25, 7459–7469. [Google Scholar] [PubMed]
- Schep, L.J.; Temple, W.A.; Butt, G.A.; Beasley, M.D. Ricin as a weapon of mass terror—Separating fact from fiction. Environ. Int. 2009, 35, 1267–1271. [Google Scholar]
- Stechmann, B.; Bai, S.-K.; Gobbo, E.; Lopez, R.; Merer, G.; Pinchard, S.; Panigai, L.; Tenza, D.; Raposo, G.; Beaumelle, B.; et al. Inhibition of retrograde transport protects mice from lethal ricin challenge. Cell 2010, 141, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Sandvig, K.; Torgersen, M.L.; Engedal, N.; Skotland, T.; Iversen, T.-G. Protein toxins from plants and bacteria: Probes for intracellular transport and tools in medicine. FEBS Lett. 2010, 584, 2626–2634. [Google Scholar]
- Johannes, L.; Popoff, V. Tracing the retrograde route in protein trafficking. Cell 2008, 135, 1175–1187. [Google Scholar]
- Lord, M.; Jolliffe, N.; Marsden, C.; Pateman, C.; Smith, D.; Spooner, R.; Watson, P.; Roberts, L. Ricin mechanisms of cytotoxicity. Toxicol. Rev. 2003, 22, 53–64. [Google Scholar]
- Leppla, S.H. Domain Structure of Bacterial Toxins. In Bacterial Protein Toxins; Burns, D.L., Barbieri, J.T., Iglewski, B.H., Rappuoli, R., Eds.; ASM Press: Washington, DC, USA, 2003; pp. 132–133. [Google Scholar]
- Tsai, B.; Ye, Y.; Rapoport, T.A. Retro-translocation of proteins from the endoplasmic reticulum into the cytosol. Nat. Rev. Mol. Cell Biol. 2002, 3, 246–255. [Google Scholar]
- Thomas, R.J. Receptor mimicry as novel therapeutic treatment for biothreat agents. Bioeng. Bugs 2010, 1, 17–30. [Google Scholar]
- Mayerhofer, P.; Cook, J.; Wahlman, J.; Pinheiro, T.; Moore, K.; Lord, J.; Johnson, A.; Roberts, L. Ricin a-chain insertion into ER membranes is triggered by a temperature increase to 37 °C. J. Biol. Chem. 2009, 284, 10232–10242. [Google Scholar]
- Spooner, R.; Hart, P.; Cook, J.; Pietroni, P.; Rogon, C.; Höhfeld, J.; Roberts, L.; Lord, J. Cytosolic chaperones influence the fate of a toxin dislocated from the endoplasmic reticulum. Proc. Natl. Acad. Sci. USA 2008, 105, 17408–17413. [Google Scholar]
- Qin, S.; Zhou, H. Dissection of the high rate constant for the binding of a ribotoxin to the ribosome. Proc. Natl. Acad. Sci. USA 2009, 106, 6974–6979. [Google Scholar]
- Dai, J.; Zhao, L.; Yang, H.; Guo, H.; Fan, K.; Wang, H.; Qian, W.; Zhang, D.; Li, B.; Wang, H.; et al. Identification of a novel functional domain of ricin responsible for its potent toxicity. J. Biol. Chem. 2011, 286, 12166–12171. [Google Scholar] [PubMed]
- Ho, M.-C.; Sturm, M.B.; Almo, S.C.; Schramm, V.L. Transition state analogues in structures of ricin and saporin ribosome-inactivating proteins. Proc. Natl. Acad. Sci. USA 2009, 106, 20276–20281. [Google Scholar]
- Miller, D.J.; Ravikumar, K.; Shen, H.; Suh, J.-K.; Kerwin, S.M.; Robertus, J.D. Structure-based design and characterization of novel platforms for ricin and shiga toxin inhibition. J. Med. Chem. 2002, 45, 90–98. [Google Scholar]
- Wahome, P.G.; Bai, Y.; Neal, L.M.; Robertus, J.D.; Mantis, N.J. Identification of small-molecule inhibitors of ricin and shiga toxin using a cell-based high-throughput screen. Toxicon 2010, 56, 313–323. [Google Scholar]
- Bai, Y.; Watt, B.; Wahome, P.G.; Mantis, N.J.; Robertus, J.D. Identification of new classes of ricin toxin inhibitors by virtual screening. Toxicon 2010, 56, 526–534. [Google Scholar]
- Van Deurs, B.; Sandvig, K.; Peterson, O.W.; Olsnes, S.; Simons, K.; Griffiths, G. Estimation of the amount of internalised ricin that reaches the trans-golgi network. J. Cell Biol. 1988, 106, 253–267. [Google Scholar]
- Schnell, R.; Borchmann, P.; Staak, J.O.; Schindler, J.; Ghetie, V.; Vitetta, E.S.; Engert, A. Clinical evaluation of ricin a-chain immunotoxins in patients with hodgkin’s lymphoma. Ann. Oncol. 2003, 14, 729–736. [Google Scholar]
- Herrera, L.; Bostrom, B.; Gore, L.; Sandler, E.; Lew, G.; Schlegel, P.; Aquino, V.; Ghetie, V.; Vitetta, E.; Schindler, J. A phase 1 study of combotox in pediatric patients with refractory B-lineage acute lymphoblastic leukemia. J. Pediatr. Hematol. Oncol. 2009, 12, 936–941. [Google Scholar]
- Schindler, J.; Gajavelli, S.; Ravandi, F.; Shen, Y.; Parekh, S.; Braunchweig, I.; Barta, S.; Ghetie, V.; Vitetta, E.; Verma, A. A phase i study of a combination of anti-CD19 and anti-CD22 immunotoxins (combotox) in adult patients with refractory B-lineage acute lymphoblastic leukaemia. Br. J. Haematol. 2011, 154, 471–476. [Google Scholar]
- Martin, P.J.; Pei, J.; Gooley, T.; Anasetti, C.; Appelbaum, F.R.; Deeg, J.; Hansen, J.A.; Nash, R.A.; Petersdorf, E.W.; Storb, R.; et al. Evaluation of a CD25-specific immunotoxin for prevention of graft-versus-host disease after unrelated marrow transplantation. Biol. Blood Marrow Transplant. 2004, 10, 552–560. [Google Scholar] [CrossRef] [PubMed]
- Saavedra-Lozano, J.; Cao, Y.; Callison, J.; Sarode, R.; Sodora, D.; Edgar, J.; Hatfield, J.; Picker, L.; Peterson, D.; Ramilo, O.; et al. An anti-CD45Ro immunotoxin kills hiv-latently infected cells from individuals on HAART with little effect on CD8 memory. Proc. Natl. Acad. Sci. USA 2004, 101, 2494–2499. [Google Scholar]
- Pincus, S.H.; Fang, H.; Wilkinson, R.A.; Marcotte, T.K.; Robinson, J.E.; Olson, W.C. In vivo efficacy of anti-gp41, but not anti-gp120, immunotoxins in a mouse model of HIV infection. J. Immunol. 2003, 170, 2236–2241. [Google Scholar] [PubMed]
- Webb, S. Pharma interest surges in antibody drug conjugates. Nat. Biotechnol. 2011, 29, 297–298. [Google Scholar]
- LeMaistre, C.F.; Rosen, S.; Frankel, A.; Kornfeld, S.; Saria, E.; Meneghetti, C.; Drajesk, J.; Fishwild, D.; Scannon, P.; Byers, V. Phase i trial of h65-rta immunoconjugate in patients with cutaneous T-cell lymphoma. Blood 1991, 78, 1173–1182. [Google Scholar]
- Amlot, P.L.; Stone, M.J.; Cunningham, D.; Fay, J.; Newman, J.; Collins, R.; May, R.; McCarthy, M.; Richardson, J.; Ghetie, V.; et al. A phase i study of an anti-CD22-deglycosylated ricin A chain immunotoxin in the treatment of B-cell lymphomas resistant to conventional therapy. Blood 1993, 82, 2624–2633. [Google Scholar] [PubMed]
- Strand, V.; Lipsky, P.E.; Cannon, G.W.; Calabrese, L.H.; Weisenhutter, C.; Cohen, S.B.; Olsen, N.J.; Lee, M.L.; Lorenz, T.J.; Nelson, B. Effects of administration of an anti-CD5 plus immunoconjugate in rheumatoid arthritis: Results of two phase two studies. Arthritis Rheum. 1993, 36, 620–630. [Google Scholar]
- Thorpe, P.E.; Wallace, P.M.; Knowles, P.P.; Relf, M.G.; Brown, A.N.; Watson, G.J.; Blakey, D.C.; Newell, D.R. Improved antitumor effects of immunotoxins prepared with deglycosylated ricin A-chain and hindered disulfide linkages. Cancer Res. 1988, 48, 6396–6403. [Google Scholar]
- Baluna, R.; Rizo, J.; Gordon, B.E.; Ghetie, V.; Vitetta, E.S. Evidence for a structural motif in toxins and interleukin-2 that may be responsible for binding to endothelial cells and initiating vascular leak syndrome. Proc. Natl. Acad. Sci. USA 1999, 96, 3957–3962. [Google Scholar]
- Smallshaw, J.E.; Ghetie, V.; Rizo, J.; Fulmer, J.R.; Trahan, L.L.; Ghetie, M.A.; Vitetta, E.S. Genetic engineering of an immunotoxin to eliminate pulmonary vascular leak in mice. Nat. Biotechnol. 2003, 21, 387–391. [Google Scholar]
- Siegall, C.B.; Liggitt, D.; Chace, D.; Tepper, M.A.; Fell, H.P. Prevention of immunotoxin-mediated vascular leak syndrome in rats with retention of antitumor activity. Proc. Natl. Acad. Sci. USA 1994, 91, 9514–9518. [Google Scholar]
- Pai, L.H.; FitzGerald, D.J.; Tepper, M.; Schacter, B.; Spitalny, G.; Pastan, I. Inhibition of antibody response to pseudomonas exotoxin and an immunotoxin containing pseudomonas exotoxin by 15-deoxyspergualin in mice. Cancer Res. 1990, 50, 7750–7753. [Google Scholar] [PubMed]
- Pietersz, G.A.; Rowland, A.; Smyth, M.J.; McKenzie, I.F.C. Chemoimmunoconjugates for the treatment of cancer. Adv. Immunol. 1994, 56, 301–387. [Google Scholar]
- Jin, F.-S.; Youle, R.J.; Johnson, V.G.; Shiloach, J.; Fass, R.; Longo, D.L.; Bridges, S.H. Suppression of the immune response to immunotoxins with anti-CD4 monoclonal antibodies. J. Immunol. 1991, 146, 1806–1811. [Google Scholar]
- Siegall, C.B.; Haggerty, H.G.; Warner, G.L.; Chace, D.; Mixan, B.; Linsley, P.S.; Davidson, T. Prevention of immunotoxin-induced immunogenicity by coadministration with CTLA4-Ig enhances antitumor efficacy. J. Immunol. 1997, 159, 5168–5173. [Google Scholar]
- Thomas, J.M.; Contreras, J.L.; Smyth, C.A.; Lobashevsky, A.; Jenkins, S.; Hubbard, W.J.; Eckhoff, D.E.; Stavrou, S.; Neville, D.M.; Thomas, F.T. Successful reversal of streptozotocin-induced diabetes with stable allogeneic islet function in a preclinical model of type 1 diabetes. Diabetes 2001, 50, 1227–1236. [Google Scholar]
- Hubbard, W.J.; Moore, J.K.; Contreras, J.L.; Smyth, C.A.; Chen, Z.W.; Lobashevsky, A.L.; Nagata, K.; Neville, D.M.; Thomas, J.M. Phenotypic and functional analysis of T-cell recovery after anti-CD3 immunotoxin treatment for tolerance induction in rhesus macaques. Hum. Immunol. 2001, 62, 479–487. [Google Scholar]
- Gelber, E.E.; Vitetta, E.S. Effect of immunosuppressive agents on the immunogenicity and efficacy of an immunotoxin in mice. Clin. Cancer Res. 1998, 4, 1297–1304. [Google Scholar]
- Takata, M.; Maiti, P.K.; Kubo, R.T.; Chen, Y.; Holford-Strevens, V.; Rector, E.S.; Sehon, A.H. Cloned suppressor T cells derived from mice tolerized with conjugates of antigen and monomethoxypolyethylene glycol. Relationship between monoclonal t suppressor factor and the T cell receptor. J. Immunol. 1990, 145, 2846–2853. [Google Scholar]
- Wang, Q.-C.; Pai, L.H.; Debinski, W.; FitzGerald, D.J.; Pastan, I. Polyethylene glycol-modified chimeric toxin composed of transforming growth factor a and pseudomonas exotoxin. Cancer Res. 1993, 53, 4588–4594. [Google Scholar] [PubMed]
- Tsutsumi, Y.; Onda, M.; Nagata, S.; Lee, B.; Kreitman, R.J.; Pastan, I. Site-specific chemical modification with polyethylene glycol of recombinant immunotoxin anti-tac(fv)-pe38 (lmb-2) improves antitumor activity and reduces animal toxicity and immunogenicity. Proc. Natl. Acad. Sci. USA 2000, 97, 8548–8553. [Google Scholar]
- Roy, C.J.; Hale, M.; Hartings, J.M.; Pitt, L.; Duniho, S. Impact of inhalation exposure modality and particle size on the respiratory deposition of ricin in balb/c mice. Inhal. Toxicol. 2003, 15, 619–638. [Google Scholar]
- Smallshaw, J.; Firan, A.; Fulmer, J.; Ruback, S.L.; Ghetie, V.; Vitetta, E. A novel recombinant vaccine which protects mice against ricin intoxication. Vaccine 2002, 20, 3422–3427. [Google Scholar]
- Smallshaw, J.E.; Richardson, J.A.; Pincus, S.; Schindler, J.; Vitetta, E.S. Preclinical toxicity and efficacy testing of Rivax, a recombinant protein vaccine against ricin. Vaccine 2005, 23, 4775–4784. [Google Scholar]
- Vitetta, E.S.; Smallshaw, J.E.; Coleman, E.; Jafri, H.; Foster, C.; Munford, R.; Schindler, J. A pilot clinical trial of a recombinant ricin vaccine in normal humans. Proc. Natl. Acad. Sci. USA 2006, 103, 2268–2273. [Google Scholar]
- Marconescu, P.S.; Smallshaw, J.E.; Pop, L.M.; Ruback, S.L.; Vitetta, E.S. Intradermal administration of Rivax protects mice from mucosal and systemic ricin intoxication. Vaccine 2010, 28, 5315–5322. [Google Scholar]
- Ishiguro, M.; Harada, H.; Ichiki, O.; Sekine, I.; Nishimori, N.; Kikutani, M. Effects of ricin, a protein toxin, on glucose absorption by the rat small intestine. (biochemical studies on oral toxicity of ricin. Ii). Chem. Pharmacol. Bull. 1984, 32, 3141–3147. [Google Scholar]
- Ishiguro, M.; Mitarai, M.; Harada, H.; Sekine, I.; Nishimori, N.; Kikutani, M. Biochemical studies on oral toxicity of ricin. I. Ricin administered orally can impair sugar absorption by rat small intestine. Chem. Pharmacol. Bull. 1983, 31, 3222–3227. [Google Scholar]
- Jackson, J.H. Tissue changes in alimentary canal of mouse induced by ricin poisoning. J. Physiol. 1957, 135, 30–31. [Google Scholar]
- Yoder, J.M.; Aslam, R.U.; Mantis, N.J. Evidence for widespread epithelial damage and coincident production of monocyte chemotactic protein 1 in a murine model of intestinal ricin intoxication. Infect. Immun. 2007, 75, 1745–1750. [Google Scholar]
- David, J.; Wilkinson, L.J.; Griffiths, G.D. Inflammatory gene expression in response to sub-lethal ricin exposure in BALB/c mice. Toxicology 2009, 264, 119–130. [Google Scholar]
- Wong, J.; Korcheva, V.; Jacoby, D.B.; Magun, B.E. Proinflammatory responses of human airway cells to ricin involve stress-activated protein kinases and NF-kappa b. Am. J. Physiol. Lung Cell Mol. Physiol. 2007, 293, L1385–L1394. [Google Scholar]
- Wong, J.; Korcheva, V.; Jacoby, D.B.; Magun, B. Intrapulmonary delivery of ricin at high dosage triggers a systemic inflammatory response and glomerular damage. Am. J. Pathol. 2007, 170, 1497–1510. [Google Scholar]
- Pincus, S.H.; Eng, L.; Cooke, C.L.; Maddaloni, M. Identification of hypoglycemia in mice as a surrogate marker of ricin toxicosis. Comp. Med. 2002, 52, 530–533. [Google Scholar]
- O’Connor, M.J.; Buckeridge, D.L.; Choy, M.; Crubezy, M.; Pincus, Z.; Musen, M.A. Biostorm: A system for automated surveillance of diverse data sources. AMIA Annu. Symp. Proc. 2003, 2003, 1071. [Google Scholar]
- Wilhelmsen, C.L.; Pitt, M.L. Lesions of acute inhaled lethal ricin intoxication in rhesus monkeys. Vet. Pathol. 1996, 33, 296–302. [Google Scholar]
- Pierce, M.; Kahn, J.N.; Chiou, J.; Tumer, N.E. Development of a quantitative rt-pcr assay to examine the kinetics of ribosome depurination by ribosome inactivating proteins using saccharomyces cerevisiae as a model. RNA 2011, 17, 201–210. [Google Scholar]
- Roche, J.; Stone, M.; Gross, L.; Lindner, M.; Seaner, R.; Pincus, S.; Obrig, T. Post-exposure targeting of specific epitopes on ricin toxin abrogates toxin-induced hypoglycemia, hepatic injury, and lethality in a mouse model. Lab. Invest. 2008, 88, 1178–1191. [Google Scholar] [CrossRef] [PubMed]
- Pratt, T.S.; Pincus, S.H.; Hale, M.L.; Moreira, A.L.; Roy, C.J.; Tchou-Wong, K.-M. Oropharyngeal aspiration of ricin as a lung challenge model for evaluation of the therapeutic index of antibodies against ricin a-chain post-exposure treatment. Exp. Lung Res. 2008, 33, 459–471. [Google Scholar]
- Maddaloni, M.; Cooke, C.; Wilkinson, R.; Stout, A.V.; Eng, L.; Pincus, S.H. Immunological characteristics associated with protective efficacy of antibodies to ricin. J. Immunol. 2004, 172, 6221–6228. [Google Scholar]
- Liu, X.-Y.; Pop, L.M.; Vitetta, E.S. Engineering therapeutic monoclonal antibodies. Immunol. Rev. 2008, 222, 9–27. [Google Scholar]
- Vitetta, E.S.; Ghetie, V.F. Immunology. Considering therapeutic antibodies. Science 2006, 313, 308–309. [Google Scholar] [PubMed]
- Hansel, T.T.; Kropshofer, H.; Singer, T.; Mitchell, J.A.; George, A.J.T. The safety and side effects of monoclonal antibodies. Nat. Rev. Drug Discov. 2010, 9, 325–338. [Google Scholar]
- Lowy, I.; Molrine, D.C.; Leav, B.A.; Blair, B.M.; Baxter, R.; Gerding, D.N.; Nichol, G.; Thomas, W.D.; Leney, M.; Sloan, S.; et al. Treatment with monoclonal antibodies against clostridium difficile toxins. N. Engl. J. Med. 2010, 362, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Migone, T.-S.; Subramanian, G.M.; Zhong, J.; Healey, L.M.; Corey, A.; Devalaraja, M.; Lo, L.; Ullrich, S.; Zimmerman, J.; Chen, A.; et al. Raxibacumab for the treatment of inhalational anthrax. N. Engl. J. Med. 2009, 361, 135–144. [Google Scholar] [PubMed]
- Lemley, P.V.; Amanatides, P.; Wright, D.C. Identification and characterization of a monoclonal antibody that neutralizes ricin toxicity in vitro and in vivo. Hybridoma 1994, 13, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Lemley, P.V.; Wright, D.C. Mice are actively immunized after passive monoclonal antibody prophylaxis and ricin toxin challenge. Immunology 1992, 76, 511–513. [Google Scholar]
- Furukawa-Stoffer, T.L.; Mah, D.C.W.; Cherwonogrodzky, J.W.; Weselake, R.J. A novel biological-based assay for the screening of neutralizing antibodies to ricin. Hybridoma 1999, 18, 505–511. [Google Scholar]
- Mantis, N.J.; McGuinness, C.R.; Sonuyi, O.; Edwards, G.; Farrant, S.A. Immunoglobulin a antibodies against ricin A and B subunits protect epithelial cells from ricin intoxication. Infect. Immun. 2006, 74, 3455–3462. [Google Scholar]
- McGuinness, C.R.; Mantis, N.J. Characterization of a novel high-affinity monoclonal immunoglobulin g antibody against the ricin B subunit. Infect. Immun. 2006, 74, 3463–3470. [Google Scholar]
- Pelat, T.; Hust, M.; Hale, M.; Lefranc, M.; Dubel, S.; Thullier, P. Isolation of a human-like antibody fragment (scFv) that neutralizes ricin biological activity. BMC Biotechnol. 2009, 9, 60. [Google Scholar] [CrossRef] [PubMed]
- Neal, L.M.; O’Hara, J.; Brey, R.N.; Mantis, N.J. A monoclonal immunoglobulin G antibody directed against an immunodominant linear epitope on the ricin A chain confers systemic and mucosal immunity to ricin. Infect. Immun. 2010, 78, 552–561. [Google Scholar]
- O’Hara, J.M.; Neal, L.M.; Mccarthy, E.A.; Kasten-Jolly, J.A.; Brey, R.N.; Mantis, N.J. Folding domains within the ricin toxin a subunit as targets of protective antibodies. Vaccine 2010, 28, 7035–7046. [Google Scholar]
- Abboud, N.; Chow, S.-K.; Saylor, C.; Janda, A.; Ravetch, J.V.; Scharff, M.D.; Casadevall, A. A requirement for Fc{gamma}r in antibody-mediated bacterial toxin neutralization. J. Exp. Med. 2010, 207, 2395–2405. [Google Scholar]
- Richalet-Sécordel, P.M.; Rauffer-Bruyère, N.; Christensen, L.L.; Ofenloch-Haehnle, B.; Seidel, C.; van Regenmortel, M.H. Concentration measurement of unpurified proteins using biosensor technology under conditions of partial mass transport limitation. Anal. Biochem. 1997, 249, 165–173. [Google Scholar]
- Griffiths, G.D.; Bailey, S.C.; Hambrook, J.L.; Keyte, M.; Jayasekera, P.; Miles, J.; Williamson, E. Liposomally-encapsulated ricin toxoid vaccine delivered intratracheally elicits a good immune response and protects against a lethal pulmonary dose of ricin toxin. Vaccine 1997, 15, 1933–1939. [Google Scholar]
- Griffiths, G.D.; Lindsay, C.D.; Allenby, A.C.; Bailey, S.C.; Scawin, J.W.; Rice, P.; Upshall, D.G. Protection against inhalation toxicity of ricin and abrin by immunisation. Hum. Exp. Toxicol. 1995, 14, 155–164. [Google Scholar]
- Kende, M.; Yan, C.; Hewetson, J.; Frick, M.A.; Rill, W.L.; Tammariello, R. Oral immunization of mice with ricin toxoid vaccine encapsulated in polymeric microspheres against aerosol challenge. Vaccine 2002, 20, 1681–1691. [Google Scholar]
- Yan, C.; Rill, W.L.; Malli, R.; Hewetson, J.; Naseem, H.; Tammariello, R.; Kende, M. Intranasal stimulation of long-lasting immunity against aerosol ricin challenge with ricin toxoid vaccine encapsulated in polymeric microspheres. Vaccine 1996, 14, 1031–1038. [Google Scholar]
- Blakey, D.C.; Thorpe, P.E. Effect of chemical deglycosylation on the in vivo fate of ricin A-chain. Cancer Drug Deliv. 1986, 3, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Foxwell, B.M.; Blakey, D.C.; Brown, A.N.; Donovan, T.A.; Thorpe, P.E. The preparation of deglycosylated ricin by recombination of glycosidase-treated A- and B-chains: Effects of deglycosylation on toxicity and in vivo distribution. Biochim. Biophys. Acta 1987, 923, 59–65. [Google Scholar] [PubMed]
- Griffiths, G.D.; Bailey, S.C.; Hambrook, J.L.; Keyte, M.P. Local and systemic responses against ricin toxin promoted by toxoid or peptide vaccines alone or in liposomal formulations. Vaccine 1998, 16, 530–535. [Google Scholar]
- Griffiths, G.D.; Phillips, G.J.; Bailey, S.C. Comparison of the quality of protection elicited by toxoid and peptide liposomal vaccine formulations against ricin as assessed by markers of inflammation. Vaccine 1999, 17, 2562–2568. [Google Scholar]
- Marsden, C.J.; Knight, S.; Smith, D.C.; Day, P.J.; Roberts, L.M.; Phillips, G.J.; Lord, J.M. Insertional mutagenesis of ricin a chain: A novel route to an anti-ricin vaccine. Vaccine 2004, 22, 2800–2805. [Google Scholar]
- Olson, M.A.; Carra, J.H.; Roxas-Duncan, V.; Wannemacher, R.W.; Smith, L.A.; Millard, C.B. Finding a new vaccine in the ricin protein fold. Protein Eng. Des. Sel. 2004, 17, 391–397. [Google Scholar]
- Porter, A.; Phillips, G.; Smith, L.; Erwin-Cohen, R.; Tammariello, R.; Hale, M.; Dasilva, L. Evaluation of a ricin vaccine candidate (rvec) for human toxicity using an in vitro vascular leak assay. Toxicon 2011, 58, 68–75. [Google Scholar] [CrossRef] [PubMed]
- McLain, D.E.; Horn, T.L.; Detrisac, C.J.; Lindsey, C.Y.; Smith, L.A. Progress in biological threat agent vaccine development: A repeat-dose toxicity study of a recombinant ricin toxin A-chain 1-33/44-198 vaccine (rvec) in male and female New Zealand White rabbits. Int. J. Toxicol. 2011, 30, 143–152. [Google Scholar]
- Legler, P.M.; Brey, R.N.; Smallshaw, J.; Vitetta, E.; Millard, C.B. Structure of rivax, a recombinant ricin vaccine. Acta Cryst. 2011, D67, 834–838. [Google Scholar]
- Neal, L.M.; Mccarthy, E.A.; Morris, C.R.; Mantis, N.J. Vaccine-induced intestinal immunity to ricin toxin in the absence of secretory IgA. Vaccine 2010, 29, 681–689. [Google Scholar]
- Mantis, N.J.; Farrant, S.A.; Mehta, S. Oligosaccharide side chains on human secretory IgA serve as receptors for ricin. J. Immunol. 2004, 11, 6838–6845. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Pincus, S.H.; Smallshaw, J.E.; Song, K.; Berry, J.; Vitetta, E.S. Passive and Active Vaccination Strategies to Prevent Ricin Poisoning. Toxins 2011, 3, 1163-1184. https://doi.org/10.3390/toxins3091163
Pincus SH, Smallshaw JE, Song K, Berry J, Vitetta ES. Passive and Active Vaccination Strategies to Prevent Ricin Poisoning. Toxins. 2011; 3(9):1163-1184. https://doi.org/10.3390/toxins3091163
Chicago/Turabian StylePincus, Seth H., Joan E. Smallshaw, Kejing Song, Jody Berry, and Ellen S. Vitetta. 2011. "Passive and Active Vaccination Strategies to Prevent Ricin Poisoning" Toxins 3, no. 9: 1163-1184. https://doi.org/10.3390/toxins3091163