Study on the Mechanism of Mesaconitine-Induced Hepatotoxicity in Rats Based on Metabonomics and Toxicology Network

Abstract
:
1. Introduction
2. Results
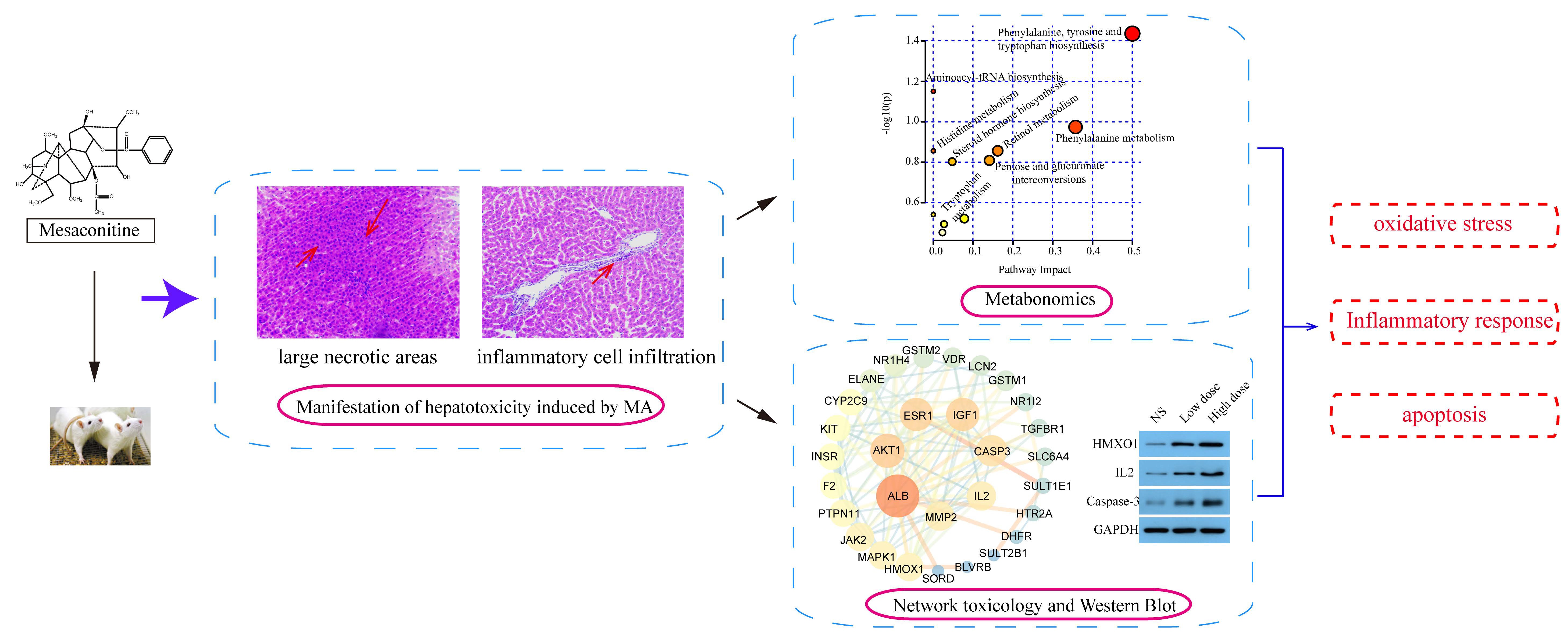
2.1. Manifestation of Hepatotoxicity Induced by MA
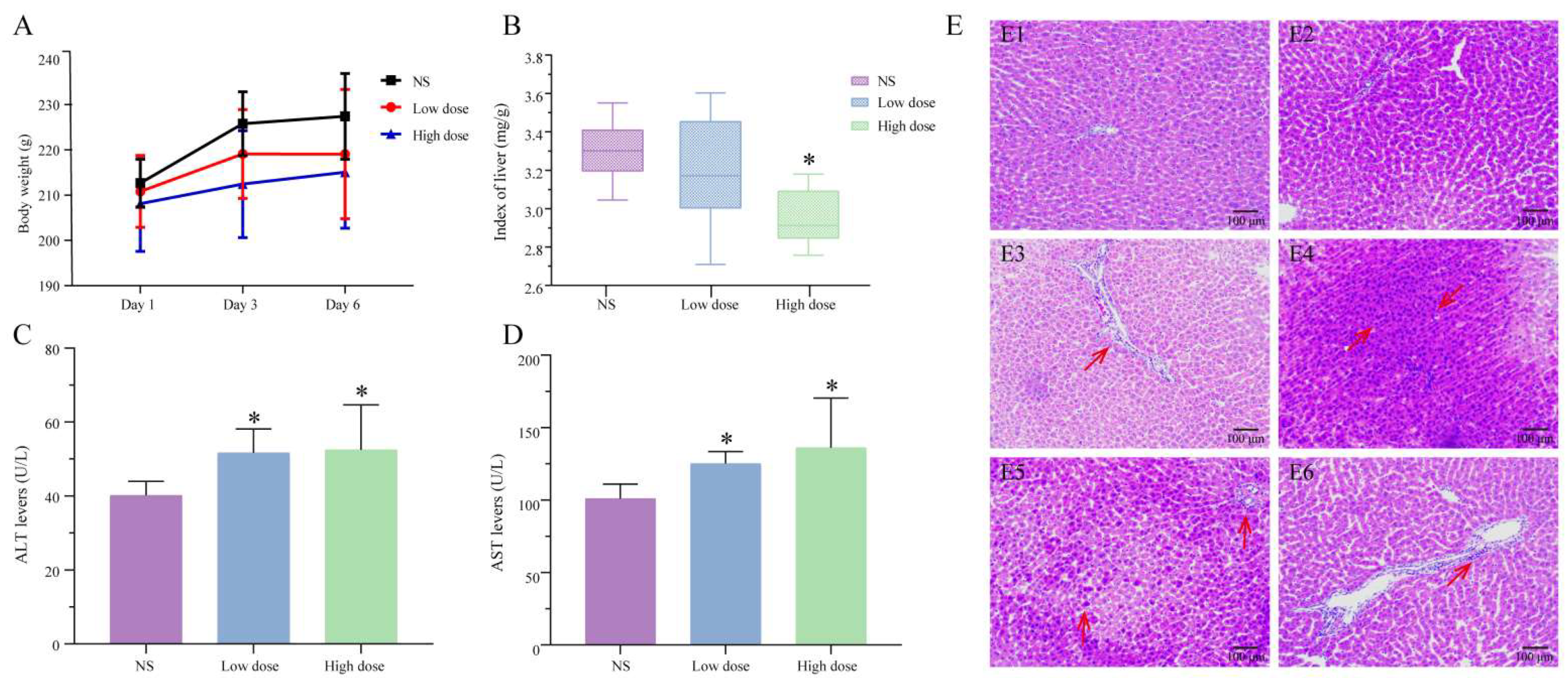
2.1.1. Analysis of Body Weight and Organ Coefficient
2.1.2. Serum Biochemical Analysis
2.1.3. Analysis of Pathological Results
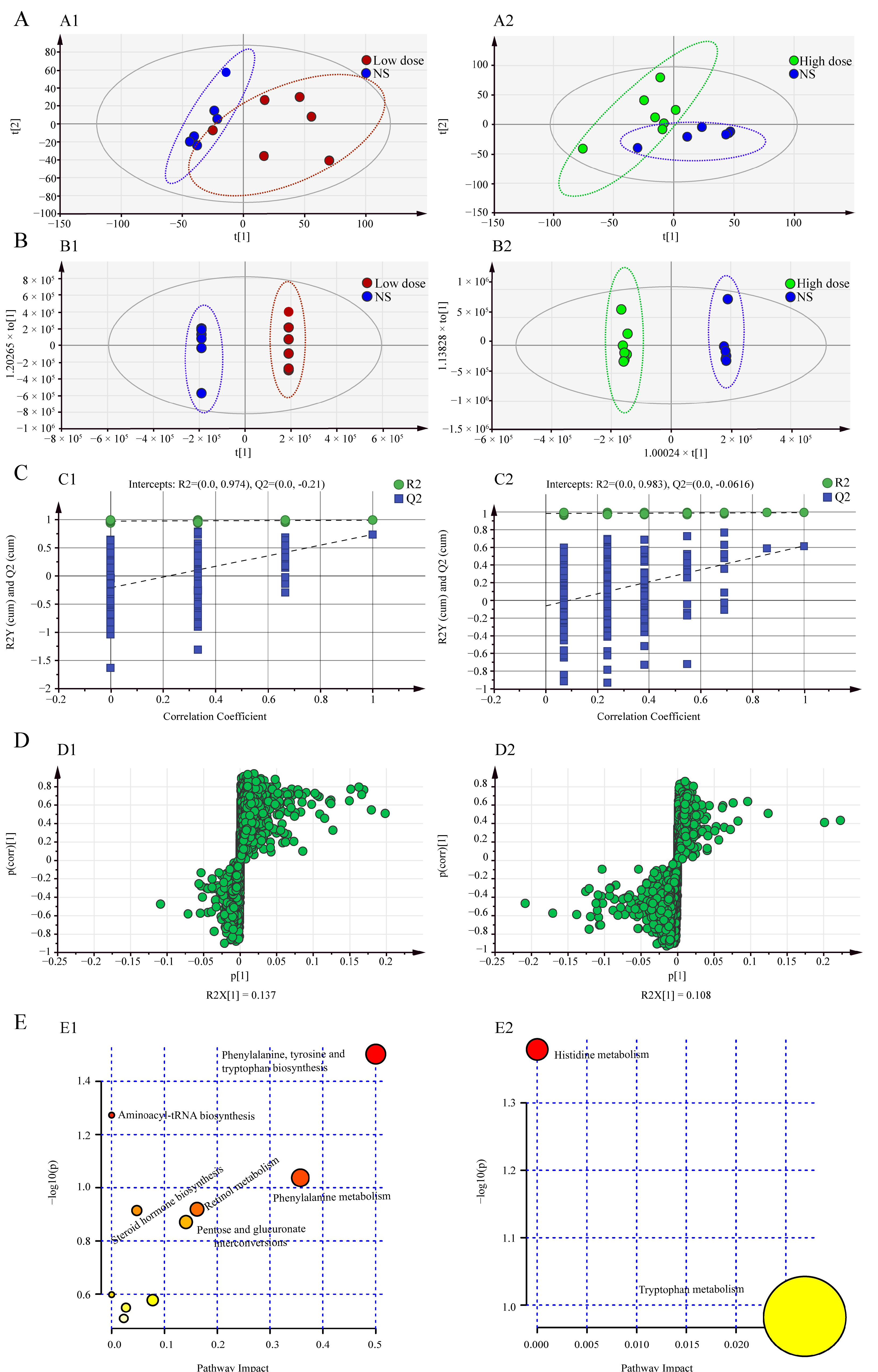
2.2. Metabonomics Study on Hepatotoxicity Induced by MA
2.2.1. Methodology Test
2.2.2. Multivariate Statistical Analysis
2.2.3. Screening of Differential Metabolites
2.2.4. Metabolic Pathway Analysis
2.3. Network Toxicological Study on Hepatotoxicity Induced by MA
2.3.1. Acquisition of “MA-Targets”
2.3.2. Acquisition of “Hepatotoxicity-Targets”
2.3.3. Construction of Common Target Based on Pathoy Language
2.3.4. Construction and Analysis of PPI Network
2.3.5. Enrichment Analysis of GO and KEGG Pathways
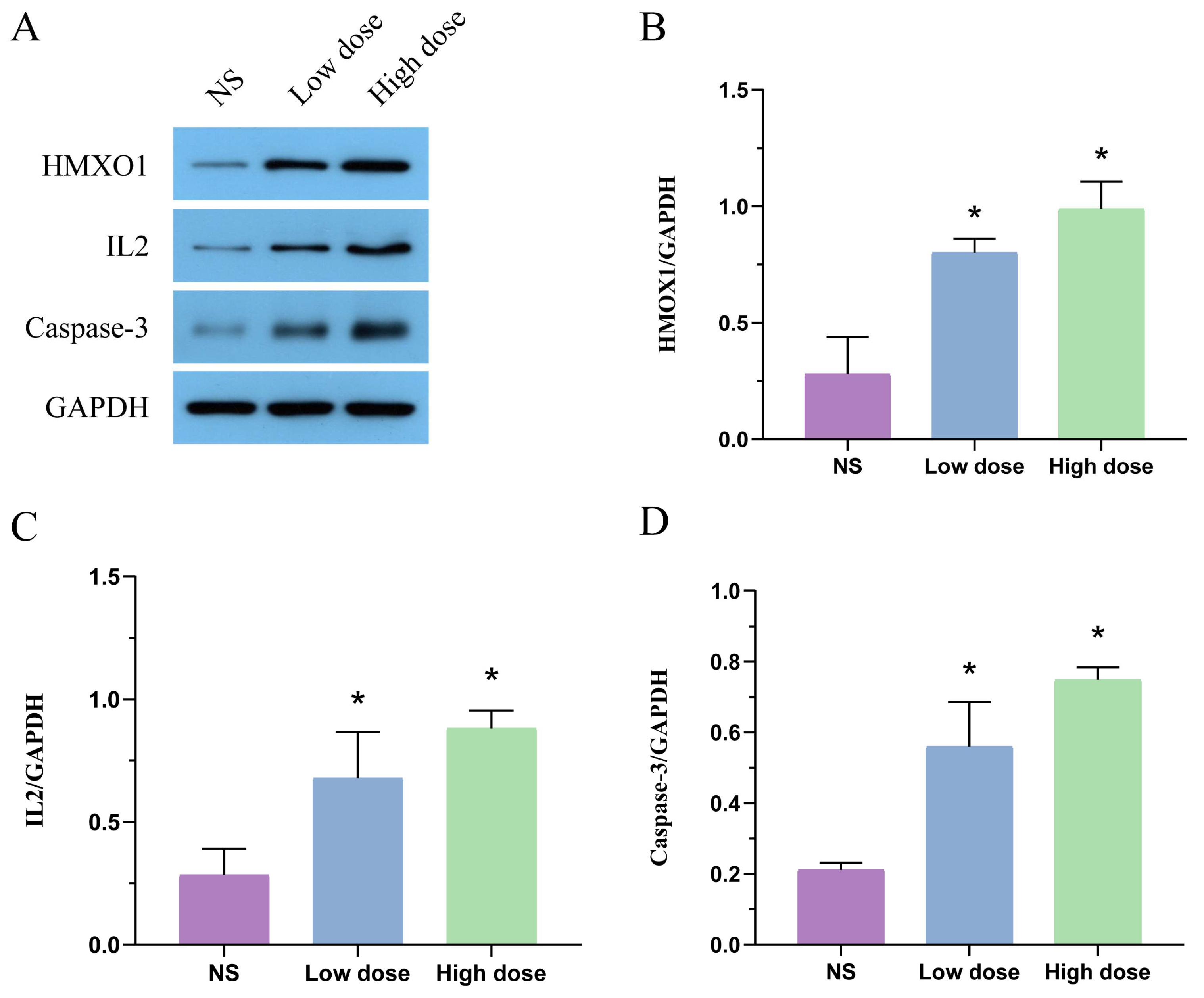
2.4. Effect of MA on Protein Expression of HMOX1, IL2 and Caspase-3 in Liver Tissue
3. Discussion
3.1. Manifestation of Hepatotoxicity Induced by MA
3.2. The Mechanism of Hepatotoxicity Induced by MA
3.3. Limitations of the Study
4. Conclusions
5. Materials and Methods
5.1. Materials and Animals
5.1.1. Instruments
5.1.2. Reference Standards and Reagents
5.1.3. Experimental Animals and Groups
5.2. Manifestation of Hepatotoxicity Induced by MA
5.2.1. Body Weight and Organ Coefficient
5.2.2. Detection of Serum Biochemical Indexes
5.2.3. Histopathological Examination
5.3. Metabonomics Study on Hepatotoxicity Induced by MA
5.3.1. Sample Preparation
Serum Sample Preparation
QC Sample Preparation
5.3.2. Analysis Conditions
Chromatographic Conditions
Mass Spectrometric Conditions
5.3.3. Methodology Test
Instrument Precision
Method Reproducibility
Sample Stability
5.3.4. Data Processing
5.4. Network Toxicological Study on Hepatotoxicity Induced by MA
5.4.1. Acquisition of “MA-Targets”
5.4.2. Acquisition of “Hepatotoxicity-Targets”
5.4.3. Construction of Common Target Based on Pathoy Language
5.4.4. Construction and Analysis of Protein–protein Interaction (PPI) Network
5.4.5. Gene Ontology (GO) Analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Enrichment Analysis
5.5. Western Blot Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tan, P.; Chen, X.; He, R.; Liu, Y. Analysis of the metabolites of mesaconitine in rat blood using ultrafast liquid chromatography and electrospray ionization mass spectrometry. Pharmacogn. Mag. 2014, 10, 101–105. [Google Scholar]
- Sun, Z.; Yang, L.; Zhao, L.; Cui, R.; Yang, W. Neuropharmacological Effects of Mesaconitine: Evidence from Molecular and Cellular Basis of Neural Circuit. Neural Plast. 2020, 2020, 8814531. [Google Scholar]
- Zhao, L.; Sun, Z.; Yang, L.; Cui, R.; Yang, W.; Li, B. Neuropharmacological effects of Aconiti Lateralis Radix Praeparata. Clin. Exp. Pharmacol. Physiol. 2020, 47, 531–542. [Google Scholar]
- Hikino, H.; Murayama, M. Mechanism of the antinociceptive action of mesaconitine: Participation of brain stem and lumbar enlargement. Br. J. Pharmacol. 1985, 85, 575–580. [Google Scholar]
- Li, X.; Ou, X.; Luo, G.; Ou, X.; Xie, Y.; Ying, M.; Qu, W.; Zuo, H.; Qi, X.; Wang, Y.; et al. Mdr1a, Bcrp and Mrp2 regulate the efficacy and toxicity of mesaconitine and hypaconitine by altering their tissue accumulation and in vivo residence. Toxicol. Appl. Pharmacol. 2020, 409, 115332. [Google Scholar]
- Ye, L.; Tang, L.; Gong, Y.; Lv, C.; Zheng, Z.; Jiang, Z.; Liu, Z. Characterization of metabolites and human P450 isoforms involved in the microsomal metabolism of mesaconitine. Xenobiotica 2011, 41, 46–58. [Google Scholar]
- Mitamura, M.; Horie, S.; Sakaguchi, M.; Someya, A.; Tsuchiya, S.; Van de Voorde, J.; Murayama, T.; Watanabe, K. Mesaconitine-induced relaxation in rat aorta: Involvement of Ca2+ influx and nitric-oxide synthase in the endothelium. Eur. J. Pharmacol. 2002, 436, 217–225. [Google Scholar]
- Nesterova, Y.V.; Povetieva, T.N.; Suslov, N.I.; Semenov, A.A.; Pushkarskiy, S.V. Antidepressant activity of diterpene alkaloids of Aconitum baicalense Turcz. Bull. Exp. Biol. Med. 2011, 151, 425–428. [Google Scholar]
- Wang, X.C.; Jia, Q.Z.; Yu, Y.L.; Wang, H.D.; Guo, H.C.; Ma, X.D.; Liu, C.T.; Chen, X.Y.; Miao, Q.F.; Guan, B.C.; et al. Inhibition of the INa/K and the activation of peak INa contribute to the arrhythmogenic effects of aconitine and mesaconitine in guinea pigs. Acta Pharmacol. Sin. 2021, 42, 218–229. [Google Scholar]
- Hsu, S.S.; Liang, W.Z. Cytotoxic Effects of Mesaconitine, the Aconitum carmichaelii Debx Bioactive Compound, on HBEC-5i Human Brain Microvascular Endothelial Cells: Role of Ca2+ Signaling-Mediated Pathway. Neurotox. Res. 2021, 39, 256–265. [Google Scholar]
- Ogura, J.; Mitamura, M.; Someya, A.; Shimamura, K.; Takayama, H.; Aimi, N.; Horie, S.; Murayama, T. Mesaconitine-induced relaxation in rat aorta: Role of Na+/Ca2+ exchangers in endothelial cells. Eur. J. Pharmacol. 2004, 483, 139–146. [Google Scholar]
- Zhou, W.; Liu, H.; Qiu, L.Z.; Yue, L.X.; Zhang, G.J.; Deng, H.F.; Ni, Y.H.; Gao, Y. Cardiac efficacy and toxicity of aconitine: A new frontier for the ancient poison. Med. Res. Rev. 2021, 41, 1798–1811. [Google Scholar]
- Qin, Y.; Wang, J.B.; Zhao, Y.L.; Shan, L.M.; Li, B.C.; Fang, F.; Jin, C.; Xiao, X.H. Establishment of a bioassay for the toxicity evaluation and quality control of Aconitum herbs. J. Hazard. Mater. 2012, 199–200, 350–357. [Google Scholar]
- Cai, Y.; Gao, Y.; Tan, G.; Wu, S.; Dong, X.; Lou, Z.; Zhu, Z.; Chai, Y. Myocardial lipidomics profiling delineate the toxicity of traditional Chinese medicine Aconiti Lateralis radix praeparata. J. Ethnopharmacol. 2013, 147, 349–356. [Google Scholar]
- Chung, J.Y.; Lee, S.J.; Lee, H.J.; Bong, J.B.; Lee, C.H.; Shin, B.S.; Kang, H.G. Aconitine Neurotoxicity According to Administration Methods. J. Clin. Med. 2021, 10, 2149. [Google Scholar]
- Xiao, F.; Qiu, J.; Zhao, Y. Exploring the Potential Toxicological Mechanisms of Vine Tea on the Liver Based on Network Toxicology and Transcriptomics. Front. Pharmacol. 2022, 13, 855926. [Google Scholar]
- Lu, X.; Huang, H.; Fu, X.; Chen, C.; Liu, H.; Wang, H.; Wu, D. The Role of Endoplasmic Reticulum Stress and NLRP3 Inflammasome in Liver Disorders. Int. J. Mol. Sci. 2022, 23, 3528. [Google Scholar]
- Fusco, R.; Gugliandolo, E.; Siracusa, R.; Scuto, M.; Cordaro, M.; D’Amico, R.; Evangelista, M.; Peli, A.; Peritore, A.F.; Impellizzeri, D.; et al. Formyl Peptide Receptor 1 Signaling in Acute Inflammation and Neural Differentiation Induced by Traumatic Brain Injury. Biology 2020, 9, 238. [Google Scholar]
- Mihajlovic, M.; Vinken, M. Mitochondria as the Target of Hepatotoxicity and Drug-Induced Liver Injury: Molecular Mechanisms and Detection Methods. Int. J. Mol. Sci. 2022, 23, 3315. [Google Scholar]
- Yang, M.; Ji, X.; Zuo, Z. Relationships between the Toxicities of Radix Aconiti Lateralis Preparata (Fuzi) and the Toxicokinetics of Its Main Diester-Diterpenoid Alkaloids. Toxins 2018, 10, 391. [Google Scholar]
- Zhou, H.; Zhang, P.; Hou, Z.; Xie, J.; Wang, Y.; Yang, B.; Xu, Y.; Li, Y. Research on the Relationships between Endogenous Biomarkers and Exogenous Toxic Substances of Acute Toxicity in Radix Aconiti. Molecules 2016, 21, 1623. [Google Scholar]
- Tan, Y.; Ko, J.; Liu, X.; Lu, C.; Li, J.; Xiao, C.; Li, L.; Niu, X.; Jiang, M.; He, X.; et al. Serum metabolomics reveals betaine and phosphatidylcholine as potential biomarkers for the toxic responses of processed Aconitum carmichaelii Debx. Mol. BioSyst. 2014, 10, 2305–2316. [Google Scholar]
- Ji, X.; Yang, M.; Or, K.H.; Yim, W.S.; Zuo, Z. Tissue Accumulations of Toxic Aconitum Alkaloids after Short-Term and Long-Term Oral Administrations of Clinically Used Radix Aconiti Lateralis Preparations in Rats. Toxins 2019, 11, 353. [Google Scholar]
- Zhang, K.; Liu, C.; Yang, T.; Li, X.; Wei, L.; Chen, D.; Zhou, J.; Yin, Y.; Yu, X.; Li, F. Systematically explore the potential hepatotoxic material basis and molecular mechanism of Radix Aconiti Lateralis based on the concept of toxicological evidence chain (TEC). Ecotoxicol. Environ. Saf. 2020, 205, 111342. [Google Scholar]
- Sun, B.; Wu, S.; Li, L.; Li, H.; Zhang, Q.; Chen, H.; Li, F.; Dong, F.; Yan, X. A metabolomic analysis of the toxicity of Aconitum sp. alkaloids in rats using gas chromatography/mass spectrometry. Rapid Commun. Mass Spectrom. 2009, 23, 1221–1228. [Google Scholar]
- Bi, C.; Zhang, T.; Li, Y.; Zhao, H.; Zhang, P.; Wang, Y.; Xu, Y.; Gu, K.; Liu, Y.; Yu, J.; et al. A Proteomics- and Metabolomics-Based Study Revealed That Disorder of Palmitic Acid Metabolism by Aconitine Induces Cardiac Injury. Chem. Res. Toxicol. 2020, 33, 3031–3040. [Google Scholar]
- Fujita, Y.; Terui, K.; Fujita, M.; Kakizaki, A.; Sato, N.; Oikawa, K.; Aoki, H.; Takahashi, K.; Endo, S. Five cases of aconite poisoning: Toxicokinetics of aconitines. J. Anal. Toxicol. 2007, 31, 132–137. [Google Scholar]
- Yan, L.; Wang, P.; Zhao, C.; Fan, S.; Lin, H.; Guo, Y.; Ma, Z.; Qiu, L. Toxic responses of liver in Lateolabrax maculatus during hypoxia and re-oxygenation. Aquat. Toxicol. 2021, 236, 105841. [Google Scholar]
- He, T.; Liu, C.; Li, M.; Wang, M.; Liu, N.; Zhang, D.; Han, S.; Li, W.; Chen, S.; Yuan, R.; et al. Integrating non-targeted metabolomics and toxicology networks to study the mechanism of Esculentoside A-induced hepatotoxicity in rats. J. Biochem. Mol. Toxicol. 2021, 35, 1–15. [Google Scholar]
- Hu, Q.; Wei, S.; Wen, J.; Zhang, W.; Jiang, Y.; Qu, C.; Xiang, J.; Zhao, Y.; Peng, X.; Ma, X. Network pharmacology reveals the multiple mechanisms of Xiaochaihu decoction in the treatment of non-alcoholic fatty liver disease. BioData Min. 2020, 13, 11. [Google Scholar]
- Yang, Z.; Mo, Y.; Cheng, F.; Zhang, H.; Shang, R.; Wang, X.; Liang, J.; Liu, Y.; Hao, B. Antioxidant Effects and Potential Molecular Mechanism of Action of Limonium aureum Extract Based on Systematic Network Pharmacology. Front. Vet. Sci. 2022, 8, 775490. [Google Scholar]
- Nishiyama, Y.; Goda, N.; Kanai, M.; Niwa, D.; Osanai, K.; Yamamoto, Y.; Senoo-Matsuda, N.; Johnson, R.S.; Miura, S.; Kabe, Y.; et al. HIF-1α induction suppresses excessive lipid accumulation in alcoholic fatty liver in mice. J. Hepatol. 2012, 56, 441–447. [Google Scholar]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline mechanisms of stress survival. Antioxid. Redox Signal. 2013, 19, 998–1011. [Google Scholar]
- Leal, J.; Teixeira-Santos, L.; Pinho, D.; Afonso, J.; Carvalho, J.; Bastos, M.d.L.; Albino-Teixeira, A.; Fraga, S.; Sousa, T. l-proline supplementation improves nitric oxide bioavailability and counteracts the blood pressure rise induced by angiotensin II in rats. Nitric Oxide Biol. Chem. 2019, 82, 1–11. [Google Scholar]
- Su, X.; Yu, W.; Liu, A.; Wang, C.; Li, X.; Gao, J.; Liu, X.; Jiang, W.; Yang, Y.; Lv, S. San-Huang-Yi-Shen Capsule Ameliorates Diabetic Nephropathy in Rats Through Modulating the Gut Microbiota and Overall Metabolism. Front. Pharmacol. 2022, 12, 808867. [Google Scholar]
- Liu, R.; Yan, X. Oxidative stress in corneal stromal cells contributes to the development of keratoconus in a rabbit model. Eur. J. Ophthalmol. 2021, 31, 3518–3524. [Google Scholar]
- Liu, Y.; Chen, X.; Liu, Y.; Chen, T.; Zhang, Q.; Zhang, H.; Zhu, Z.; Chai, Y.; Zhang, J. Metabolomic study of the protective effect of Gandi capsule for diabetic nephropathy. Chem. Biol. Interact. 2019, 314, 108815. [Google Scholar]
- Anesi, A.; Rubert, J.; Oluwagbemigun, K.; Orozco-Ruiz, X.; Nöthlings, U.; Breteler, M.; Mattivi, F. Metabolic Profiling of Human Plasma and Urine, Targeting Tryptophan, Tyrosine and Branched Chain Amino Acid Pathways. Metabolites 2019, 9, 261. [Google Scholar]
- Tu, C.; Niu, M.; Li, C.; Liu, Z.; He, Q.; Li, R.; Zhang, Y.; Xiao, X.; Wang, J. Network pharmacology oriented study reveals inflammatory state-dependent dietary supplement hepatotoxicity responses in normal and diseased rats. Food Funct. 2019, 10, 3477–3490. [Google Scholar]
- Kumar, N.; Surani, S.; Udeani, G.; Mathew, S.; John, S.; Sajan, S.; Mishra, J. Drug-induced liver injury and prospect of cytokine based therapy; A focus on IL-2 based therapies. Life Sci. 2021, 278, 119544. [Google Scholar]
- Nakagawa, K.; Miller, F.N.; Sims, D.E.; Lentsch, A.B.; Miyazaki, M.; Edwards, M.J. Mechanisms of interleukin-2-induced hepatic toxicity. Cancer Res. 1996, 56, 507–510. [Google Scholar]
- Strieter, R.M.; Remick, D.G.; Lynch, J.P.; Spengler, R.N.; Kunkel, S.L. Interleukin-2-induced tumor necrosis factor-alpha (TNF-alpha) gene expression in human alveolar macrophages and blood monocytes. Am. Rev. Respir. Dis. 1989, 139, 335–342. [Google Scholar]
- Abu-Amara, M.; Yang, S.Y.; Tapuria, N.; Fuller, B.; Davidson, B.; Seifalian, A. Liver ischemia/reperfusion injury: Processes in inflammatory networks—A review. Liver Transplant. 2010, 16, 1016–1032. [Google Scholar]
- Shuh, M.; Bohorquez, H.; Loss, G.E.; Cohen, A.J. Tumor Necrosis Factor-α: Life and Death of Hepatocytes During Liver Ischemia/Reperfusion Injury. Ochsner J. 2013, 13, 119–130. [Google Scholar]
- Guha, M.; Mackman, N. The phosphatidylinositol 3-kinase-Akt pathway limits lipopolysaccharide activation of signaling pathways and expression of inflammatory mediators in human monocytic cells. J. Biol. Chem. 2002, 277, 32124–32132. [Google Scholar]
- Díaz-Guerra, M.J.; Castrillo, A.; Martín-Sanz, P.; Boscá, L. Negative regulation by phosphatidylinositol 3-kinase of inducible nitric oxide synthase expression in macrophages. J. Immunol. 1999, 162, 6184–6190. [Google Scholar]
- Li, X.; Wu, Y.; Zhang, W.; Gong, J.; Cheng, Y. Pre-conditioning with tanshinone IIA attenuates the ischemia/reperfusion injury caused by liver grafts via regulation of HMGB1 in rat Kupffer cells. Biomed. Pharmacother. 2017, 89, 1392–1400. [Google Scholar]
- Xiao, Q.; Ye, Q.; Wang, W.; Xiao, J.; Fu, B.; Xia, Z.; Zhang, X.; Liu, Z.; Zeng, X. Mild hypothermia pretreatment protects against liver ischemia reperfusion injury via the PI3K/AKT/FOXO3a pathway. Mol. Med. Rep. 2017, 16, 7520–7526. [Google Scholar]
- Shen, Y.; Shen, X.; Cheng, Y.; Liu, Y. Myricitrin pretreatment ameliorates mouse liver ischemia reperfusion injury. Int. Immunopharmacol. 2020, 89, 107005. [Google Scholar]
- Reyes-Gordillo, K.; Shah, R.; Arellanes-Robledo, J.; Cheng, Y.; Ibrahim, J.; Tuma, P.L. Akt1 and Akt2 Isoforms Play Distinct Roles in Regulating the Development of Inflammation and Fibrosis Associated with Alcoholic Liver Disease. Cells 2019, 8, 1337. [Google Scholar]
- O’Neil, J.D.; Ammit, A.J.; Clark, A.R. MAPK p38 regulates inflammatory gene expression via tristetraprolin: Doing good by stealth. Int. J. Biochem. Cell Biol. 2018, 94, 6–9. [Google Scholar]
- Fusco, R.; Cordaro, M.; Siracusa, R.; D’Amico, R.; Genovese, T.; Gugliandolo, E.; Peritore, A.F.; Crupi, R.; Impellizzeri, D.; Cuzzocrea, S.; et al. Biochemical Evaluation of the Antioxidant Effects of Hydroxytyrosol on Pancreatitis-Associated Gut Injury. Antioxidants 2020, 9, 781. [Google Scholar]
- Yu, Y.B.; Zhuang, H.Z.; Ji, X.J.; Dong, L.; Duan, M.L. Hydroxytyrosol suppresses LPS-induced intrahepatic inflammatory responses via inhibition of ERK signaling pathway activation in acute liver injury. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 6455–6462. [Google Scholar]
- Beggs, K.M.; Maiuri, A.R.; Fullerton, A.M.; Poulsen, K.L.; Breier, A.B.; Ganey, P.E.; Roth, R.A. Trovafloxacin-induced replication stress sensitizes HepG2 cells to tumor necrosis factor-alpha-induced cytotoxicity mediated by extracellular signal-regulated kinase and ataxia telangiectasia and Rad3-related. Toxicology 2015, 331, 35–46. [Google Scholar]
- Chen, S.; Zhang, Z.; Wu, Y.; Shi, Q.; Yan, H.; Mei, N.; Tolleson, W.H.; Guo, L. Endoplasmic Reticulum Stress and Store-Operated Calcium Entry Contribute to Usnic Acid-Induced Toxicity in Hepatic Cells. Toxicol. Sci. 2015, 146, 116–126. [Google Scholar]
- Hu, J.; Ramshesh, V.K.; McGill, M.R.; Jaeschke, H.; Lemasters, J.J. Low Dose Acetaminophen Induces Reversible Mitochondrial Dysfunction Associated with Transient c-Jun N-Terminal Kinase Activation in Mouse Liver. Toxicol. Sci. 2016, 150, 204–215. [Google Scholar]
- Poulsen, K.L.; Albee, R.P.; Ganey, P.E.; Roth, R.A. Trovafloxacin potentiation of lipopolysaccharide-induced tumor necrosis factor release from RAW 264.7 cells requires extracellular signal-regulated kinase and c-Jun N-Terminal Kinase. J. Pharmacol. Exp. Ther. 2014, 349, 185–191. [Google Scholar]
- Wu, Y.; Beland, F.A.; Chen, S.; Liu, F.; Guo, L.; Fang, J.L. Mechanisms of tolvaptan-induced toxicity in HepG2 cells. Biochem. Pharmacol. 2015, 95, 324–336. [Google Scholar]
- Cheng, C.W.; Liu, M.H.; Tang, H.Y.; Cheng, M.L.; Wang, C.H. Factors associated with elevated plasma phenylalanine in patients with heart failure. Amino Acids 2021, 53, 149–157. [Google Scholar]
- Chen, W.S.; Wang, C.H.; Cheng, C.W.; Liu, M.H.; Chu, C.M.; Wu, H.P.; Huang, P.C.; Lin, Y.T.; Ko, T.; Chen, W.H.; et al. Elevated plasma phenylalanine predicts mortality in critical patients with heart failure. ESC Heart Fail. 2020, 7, 2884–2893. [Google Scholar]
- Wang, M.; Zhang, J.; Gong, N. Role of the PI3K/Akt signaling pathway in liver ischemia reperfusion injury: A narrative review. Ann. Palliat. Med. 2022, 11, 806–817. [Google Scholar]
- Wu, D.; Yang, C.; Yang, M.; Wu, Y.; Mao, Y.; Zhou, X.; Wang, J.; Yuan, Z.; Wu, J. Citrinin-Induced Hepatotoxicity in Mice Is Regulated by the Ca2+/Endoplasmic Reticulum Stress Signaling Pathway. Toxins 2022, 14, 259. [Google Scholar]
- Rehman, M.U.; Tahir, M.; Khan, A.Q.; Khan, R.; Lateef, A.; Hamiza, O.O.; Ali, F.; Sultana, S. Diosmin protects against trichloroethylene-induced renal injury in Wistar rats: Plausible role of p53, Bax and caspases. Br. J. Nutr. 2013, 110, 699–710. [Google Scholar]
- Prokhorova, E.A.; Kopeina, G.S.; Lavrik, I.N.; Zhivotovsky, B. Apoptosis regulation by subcellular relocation of caspases. Sci. Rep. 2018, 8, 12199. [Google Scholar]
- Wang, Z.; Hao, W.; Hu, J.; Mi, X.; Han, Y.; Ren, S.; Jiang, S.; Wang, Y.; Li, X.; Li, W. Maltol Improves APAP-Induced Hepatotoxicity by Inhibiting Oxidative Stress and Inflammation Response via NF-κB and PI3K/Akt Signal Pathways. Antioxidants 2019, 8, 395. [Google Scholar]
- Cui, M.; Huang, Y.; Zhao, Y.; Zheng, J. New insights for FOXO and cell-fate decision in HIV infection and HIV associated neurocognitive disorder. Adv. Exp. Med. Biol. 2009, 665, 143–159. [Google Scholar]
- Lau, C.J.; Koty, Z.; Nalbantoglu, J. Differential response of glioma cells to FOXO1-directed therapy. Cancer Res. 2009, 69, 5433–5440. [Google Scholar]
- Su, Y.W.; Hao, Z.; Hirao, A.; Yamamoto, K.; Lin, W.J.; Young, A.; Duncan, G.S.; Yoshida, H.; Wakeham, A.; Lang, P.A.; et al. 14-3-3sigma regulates B-cell homeostasis through stabilization of FOXO1. Proc. Natl. Acad. Sci. USA 2011, 108, 1555–1560. [Google Scholar]
- Sullivan, J.A.; Kim, E.H.; Plisch, E.H.; Suresh, M. FOXO3 regulates the CD8 T cell response to a chronic viral infection. J. Virol. 2012, 86, 9025–9034. [Google Scholar]
- Tan, M.; Cheng, Y.; Zhong, X.; Yang, D.; Jiang, S.; Ye, Y.; Ding, M.; Guan, G.; Yang, D.; Zhao, X. LNK promotes granulosa cell apoptosis in PCOS via negatively regulating insulin-stimulated AKT-FOXO3 pathway. Aging 2021, 13, 4617–4633. [Google Scholar]
- Sun, F.; Zhou, J.L.; Wei, S.X.; Jiang, Z.W.; Peng, H. Glucocorticoids induce osteonecrosis of the femoral head in rats via PI3K/AKT/FOXO1 signaling pathway. PeerJ 2022, 10, e13319. [Google Scholar]
- Burgering, B.M.; Kops, G.J. Cell cycle and death control: Long live Forkheads. Trends Biochem. Sci. 2002, 27, 352–360. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Rt/min | Ion Mode | Obsed m/z | Calcd m/z | Error (ppm) | Formula | Identify | HMDB | Group |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 0.931 | [M+H]+ | 130.08621 | 130.08625 | −0.307 | C6H11NO2 | D-Pipecolic acid | HMDB0005960 | High dose |
| 2 | 1.918 | [M+H]+ | 116.07054 | 116.07060 | −0.517 | C5H9NO2 | L-Proline | HMDB0000162 | Low dose |
| 3 | 2.217 | [M+H]+ | 132.10193 | 132.10190 | 0.227 | C6H13NO2 | D-Leucine | HMDB0013773 | High dose |
| 4 | 2.810 | [M+H]+ | 115.11172 | 115.11174 | −0.174 | C7H14O | 4-Heptanone | HMDB0004814 | High dose |
| 5 | 2.849 | [M+H]+ | 166.08612 | 166.08625 | −0.783 | C9H11NO2 | L-Phenylalanine | HMDB0000159 | Low dose |
| 6 | 3.373 | [M+H]+ | 340.10211 | 340.10269 | −1.705 | C15H17NO8 | 5-Hydroxy-6-methoxyindole glucuronide | HMDB0010363 | Low dose |
| 7 | 3.886 | [M+H]+ | 141.06564 | 141.06585 | −1.489 | C6H8N2O2 | Methylimidazoleacetic acid | HMDB0002820 | High dose |
| 8 | 4.757 | [M+H]+ | 514.28412 | 514.28329 | 1.614 | C26H43NO7S | Sulfolithocholylglycine | HMDB0002639 | Low dose |
| 9 | 4.901 | [M+H]+ | 176.07085 | 176.07060 | 1.420 | C10H9NO2 | 5-Hydroxyindoleacetaldehyde | HMDB0004073 | Low dose High dose |
| 10 | 5.909 | [M+H]+ | 190.08615 | 190.08625 | −0.526 | C11H11NO2 | Indole-3-methyl acetate | HMDB0029738 | Low dose High dose |
| 11 | 6.336 | [M+H]+ | 361.19955 | 361.20095 | −3.876 | C21H28O5 | Aldosterone | HMDB0000037 | Low dose |
| 12 | 7.945 | [M+H]+ | 466.31470 | 466.31631 | −3.453 | C26H43NO6 | Glycocholic acid | HMDB0000138 | Low dose |
| 13 | 8.033 | [M+H]+ | 289.21573 | 289.21620 | −1.625 | C19H28O2 | Dehydroepiandrosterone | HMDB0000077 | Low dose |
| 14 | 8.034 | [M+H]+ | 118.06486 | 118.06512 | −2.202 | C8H7N | Indole | HMDB0000738 | Low dose High dose |
| 15 | 8.536 | [M+Na]+ | 351.23093 | 351.22945 | 4.214 | C22H32O2 | Docosahexaenoic acid | HMDB0002183 | Low dose |
| 16 | 8.823 | [M+K]+ | 369.24194 | 369.24016 | 4.821 | C19H38O4 | MG (0:0/16:0/0:0) | HMDB0011533 | Low dose High dose |
| 17 | 9.351 | [M+H]+ | 314.23276 | 314.23258 | 0.573 | C17H31NO4 | 9-Decenoylcarnitine | HMDB0013205 | Low dose |
| 18 | 10.480 | [M+H]+ | 316.24741 | 316.24823 | −2.593 | C17H33NO4 | Decanoylcarnitine | HMDB0000651 | High dose |
| 19 | 12.718 | [M+H]+ | 368.27847 | 368.27953 | −2.878 | C21H37NO4 | 3,5-Tetradecadiencarnitine | HMDB0013331 | Low dose High dose |
| 20 | 12.733 | [M+H]+ | 388.30527 | 388.30574 | −1.210 | C21H41NO5 | 2-Hydroxymyristoylcarnitine | HMDB0013166 | High dose |
| 21 | 12.750 | [M+H]+ | 295.22559 | 295.22677 | −3.997 | C18H30O3 | 9-OxoODE | HMDB0004669 | Low dose |
| 22 | 13.111 | [M+H]+ | 344.28049 | 344.27953 | 2.788 | C19H37NO4 | Dodecanoylcarnitine | HMDB0002250 | High dose |
| 23 | 14.109 | [M+H]+ | 303.23233 | 303.23185 | 1.583 | C20H30O2 | Retinyl ester | HMDB0003598 | Low dose |
| 24 | 18.521 | [M+H]+ | 422.32602 | 422.32648 | −1.089 | C25H43NO4 | Gamma-linolenyl carnitine | HMDB0006318 | Low dose High dose |
| No. | Metabolic Pathway | Metabolite | p | −Log(p) | Holm p | FDR | Impact | Group |
|---|---|---|---|---|---|---|---|---|
| 1 | Phenylalanine, tyrosine and tryptophan biosynthesis | L-Phenylalanine | 0.031463 | 1.5022 | 1.0 | 1.0 | 0.5 | Low dose |
| 2 | Histidine metabolism | Methylimidazoleacetic acid | 0.041783 | 1.379 | 1.0 | 1.0 | 0.0 | High dow |
| 3 | Aminoacyl-tRNA biosynthesis | L-Phenylalanine; L-Proline | 0.05338 | 1.2726 | 1.0 | 1.0 | 0.0 | Low dose |
| 4 | Phenylalanine metabolism | L-Phenylalanine | 0.091683 | 1.0377 | 1.0 | 1.0 | 0.35714 | Low dose |
| 5 | Retinol metabolism | Retinyl ester | 0.12049 | 0.91906 | 1.0 | 1.0 | 0.16168 | Low dose |
| 6 | Steroid hormone biosynthesis | Dehydroepiandrosterone; Aldosterone | 0.12185 | 0.91416 | 1.0 | 1.0 | 0.0474 | Low dose |
| 7 | Pentose and glucuronate interconversions | beta-D-Glucuronoside | 0.13457 | 0.87104 | 1.0 | 1.0 | 0.14062 | Low dose |
| 8 | Biosynthesis of unsaturated fatty acids | (4Z,7Z,10Z,13Z,16Z,19Z)-Docosahexaenoic acid | 0.25235 | 0.59799 | 1.0 | 1.0 | 0.0 | Low dose |
| 9 | Arginine and proline metabolism | L-Proline | 0.26449 | 0.57759 | 1.0 | 1.0 | 0.0778 | Low dose |
| 10 | Tryptophan metabolism | 5-Hydroxyindoleacetaldehyde | 0.28236 0.10443 | 0.5492 0.98117 | 1.0 | 1.0 | 0.02691 | Low dose High dose |
| 11 | Primary bile acid biosynthesis | Glycocholic acid | 0.31125 | 0.50689 | 1.0 | 1.0 | 0.02285 | Low dose |
| Grouping | Number | Drug | Dose | Method of Administration | Exposure Period |
|---|---|---|---|---|---|
| NS | 10 | 0.9% NaCl | 8.0 mL/kg/day | p.o., Continuous administration | 6 days |
| Low dose | 11 | MA | 0.8 mg/kg/day | p.o., Continuous administration | 6 days |
| High dose | 16 | MA | 1.2 mg/kg/day | p.o., Continuous administration | 6 days |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Q.; Zhang, K.; Jiao, M.; Jiao, J.; Chen, D.; Yin, Y.; Zhang, J.; Li, F. Study on the Mechanism of Mesaconitine-Induced Hepatotoxicity in Rats Based on Metabonomics and Toxicology Network. Toxins 2022, 14, 486. https://doi.org/10.3390/toxins14070486
Chen Q, Zhang K, Jiao M, Jiao J, Chen D, Yin Y, Zhang J, Li F. Study on the Mechanism of Mesaconitine-Induced Hepatotoxicity in Rats Based on Metabonomics and Toxicology Network. Toxins. 2022; 14(7):486. https://doi.org/10.3390/toxins14070486
Chicago/Turabian StyleChen, Qian, Kai Zhang, Mingjie Jiao, Jiakang Jiao, Dongling Chen, Yihui Yin, Jia Zhang, and Fei Li. 2022. "Study on the Mechanism of Mesaconitine-Induced Hepatotoxicity in Rats Based on Metabonomics and Toxicology Network" Toxins 14, no. 7: 486. https://doi.org/10.3390/toxins14070486