The Effects of Ferric Sulfate (Fe2(SO4)3) on the Removal of Cyanobacteria and Cyanotoxins: A Mesocosm Experiment

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Results and Discussions
2.1. Impact of Ferric Sulfate on Cyanobacterial Cell Counts
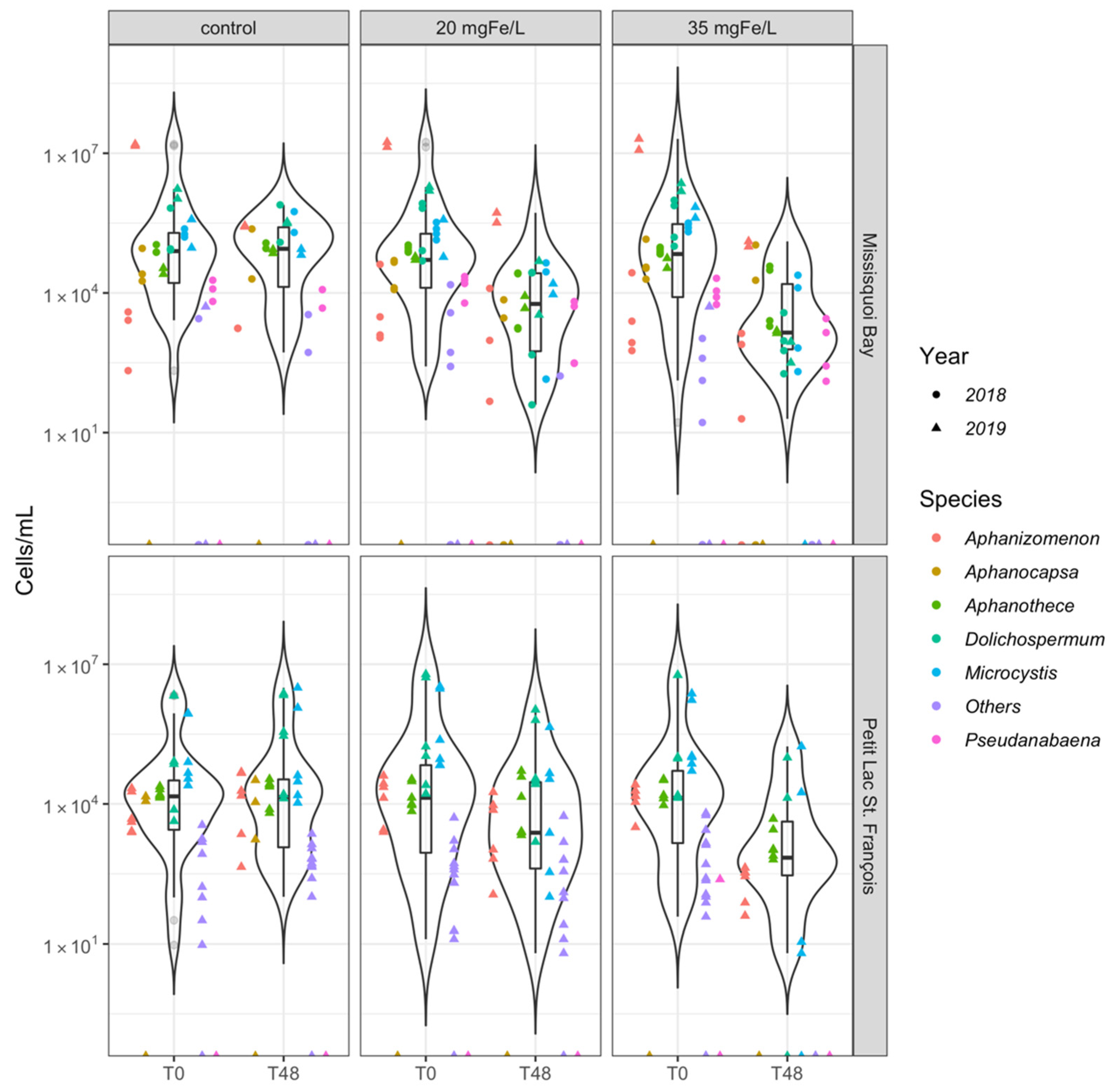
2.1.1. Taxonomic Cyanobacterial Cell Counts and Composition
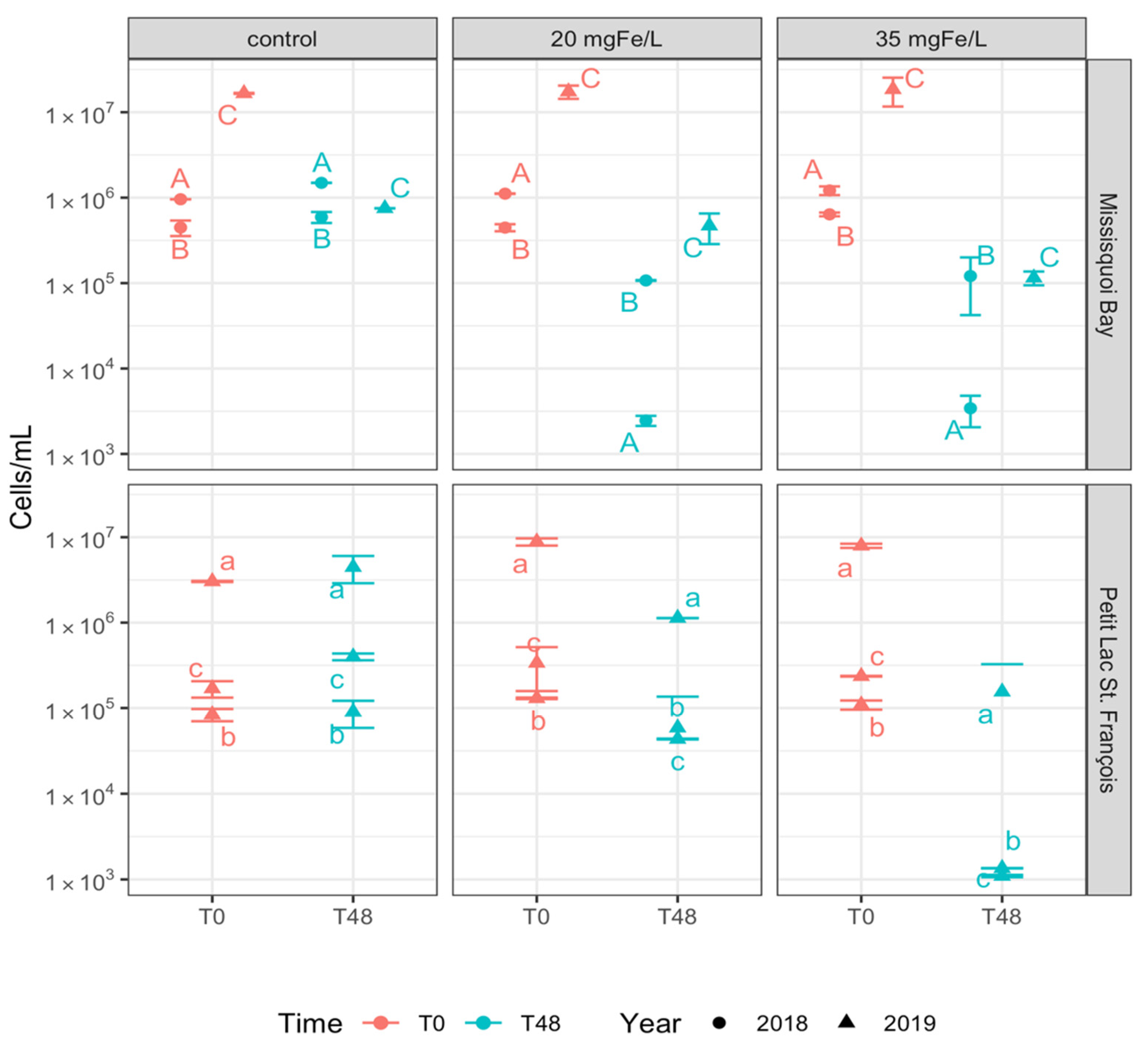
2.1.2. Removal of Cyanobacterial Cells
2.2. Impact of Ferric Sulfate on Intracellular Cyanotoxins
2.2.1. Intracellular Cyanotoxin Concentration and Composition
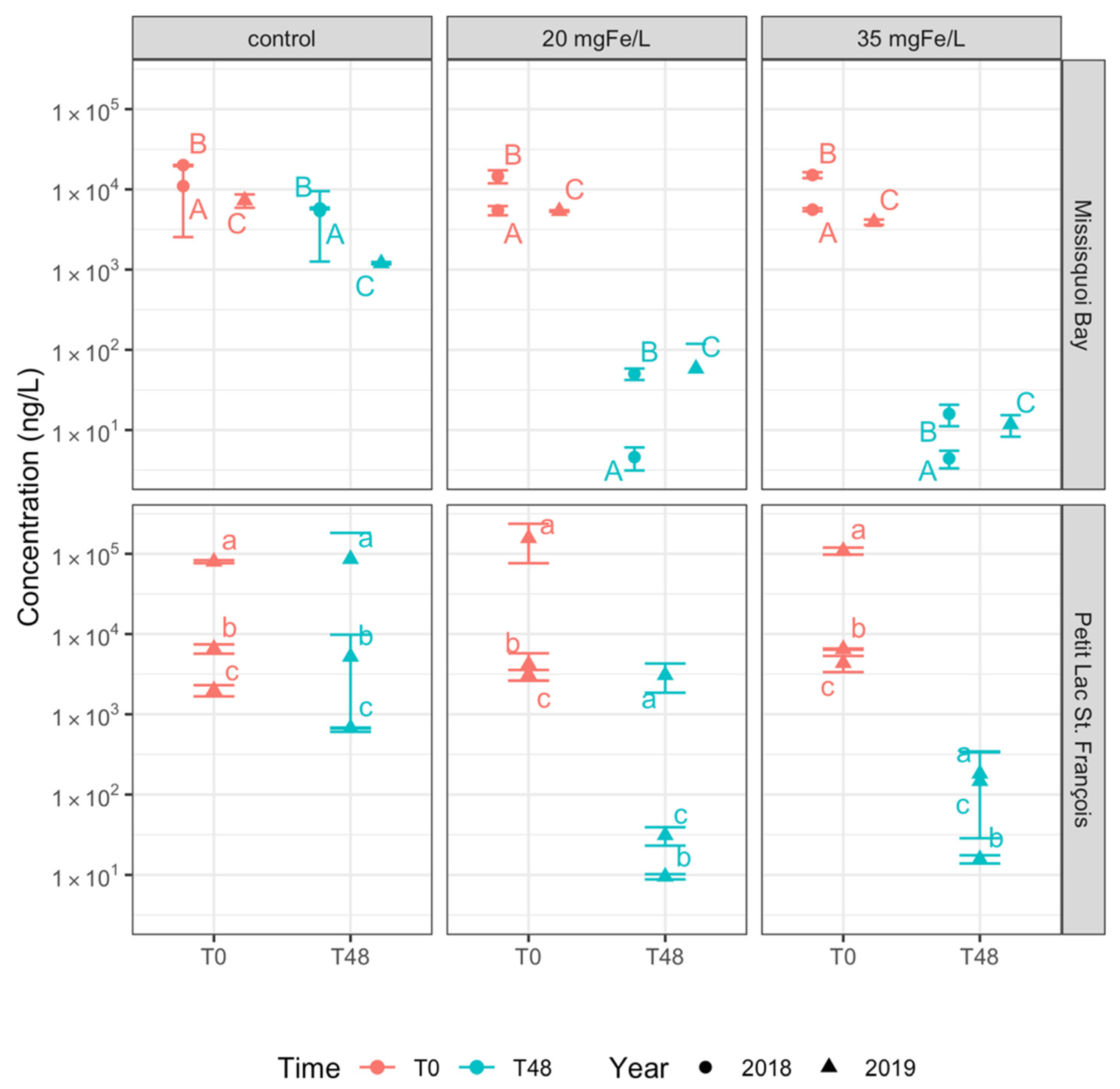
2.2.2. Intracellular Cyanotoxin Removal
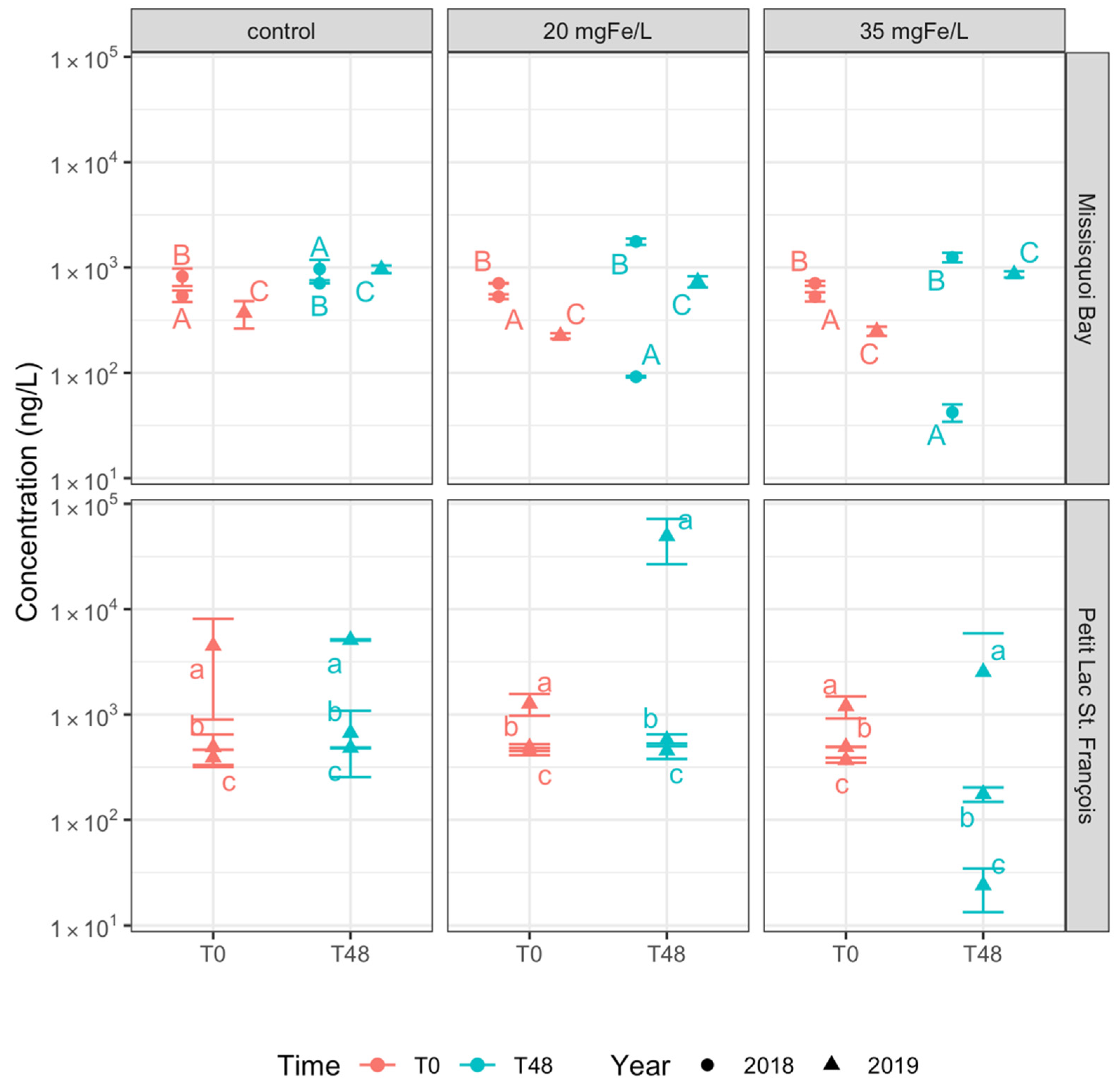
2.3. Effect of Ferric Sulfate on Extracellular Cyanotoxins
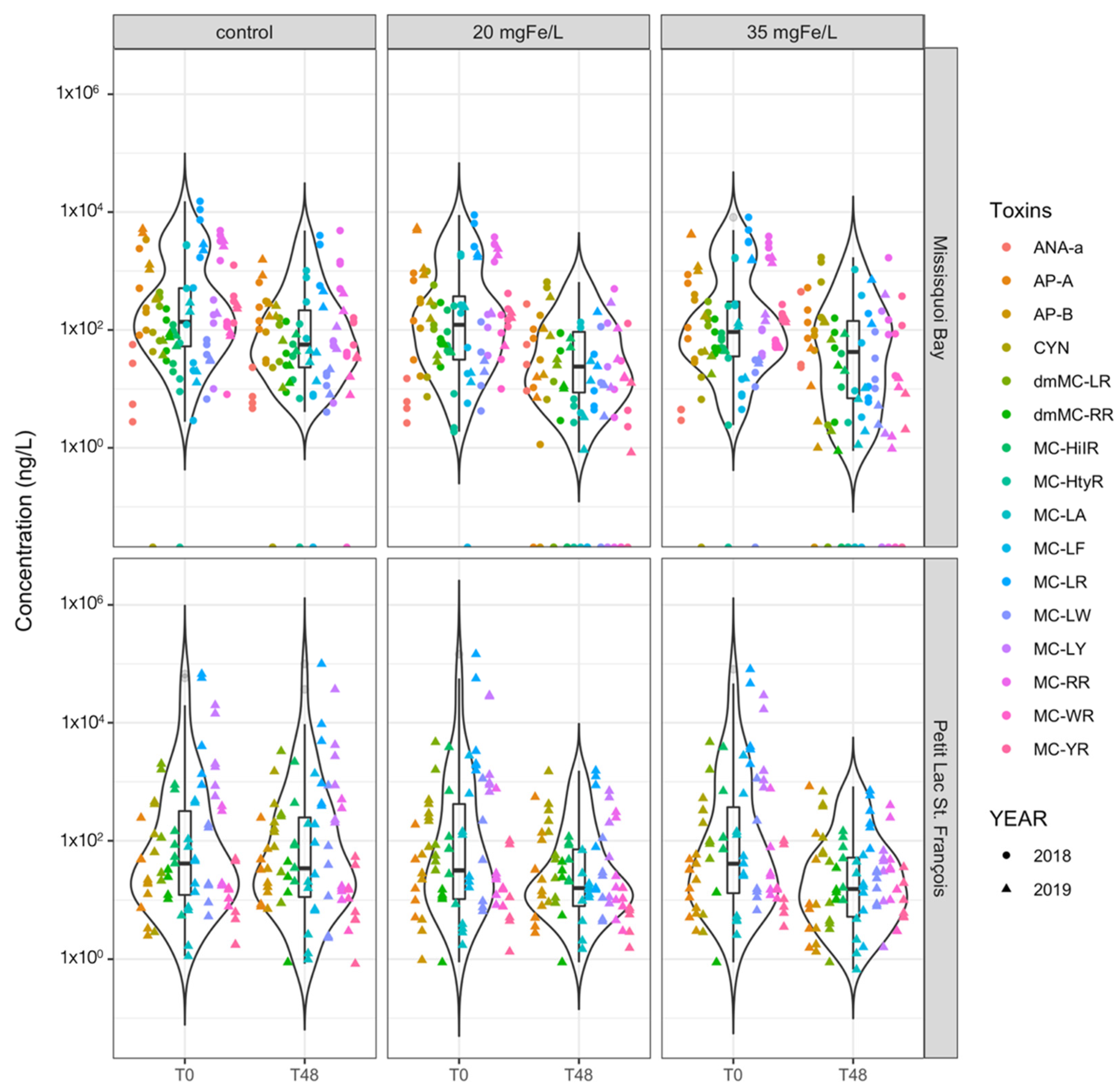
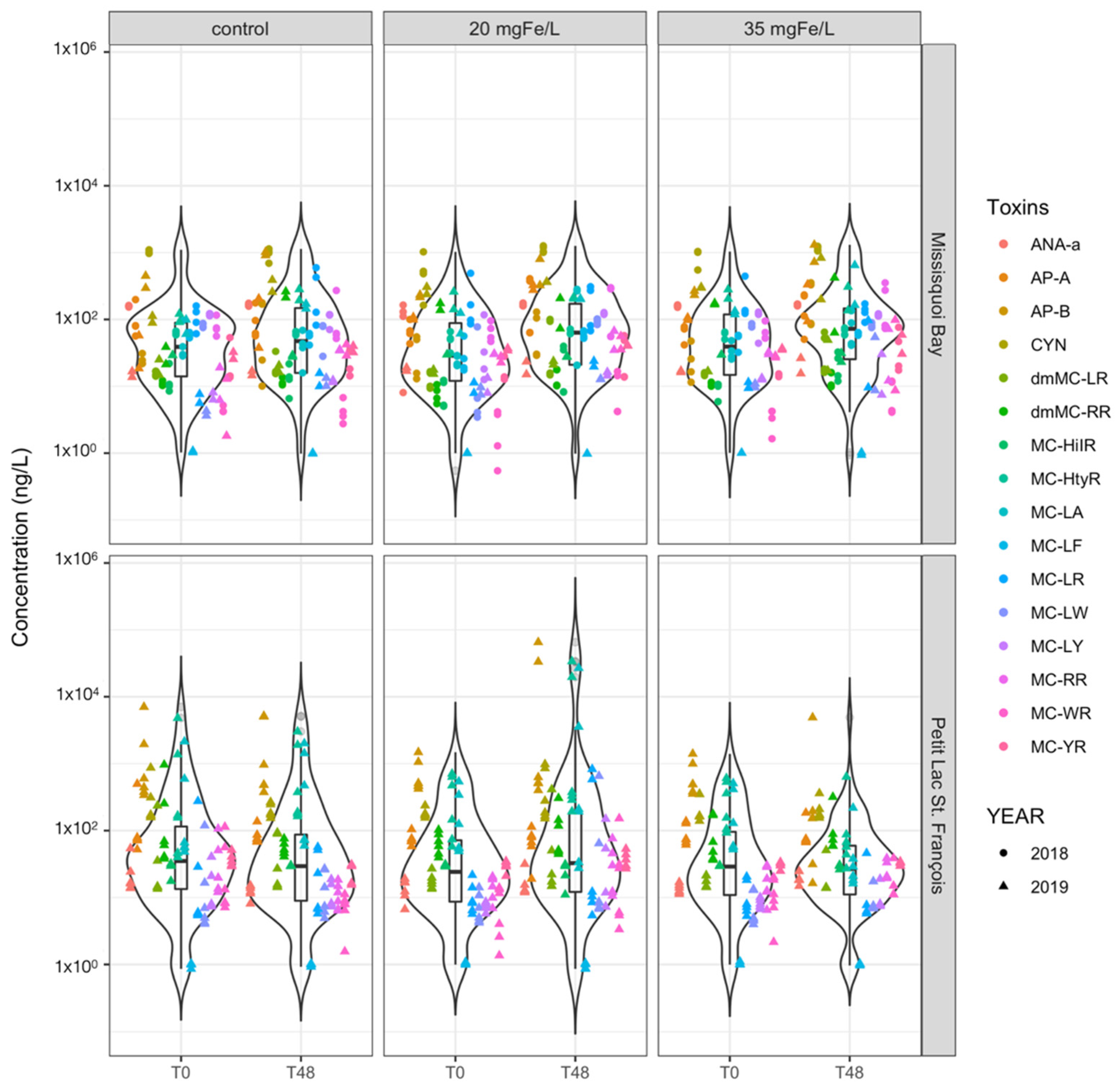
2.3.1. Extracellular Cyanotoxin Concentration and Composition
2.3.2. Extracellular Cyanotoxins Removal
3. Conclusions
- Dolichospermum, Microcystis, and Aphanizomenon were the dominant genera throughout the experimental period. Aphanothece spp. and Coelosphaerium spp. were also documented at some events;
- Total cyanobacterial cells were efficiently removed, with more than 96% and 51% removal in mesocosms with applied dose of 35 mgFe/L and 20 mgFe/L, respectively. Significant differences in removing total cyanobacterial cells and several dominant cyanobacterial genera were observed between applied doses;
- Both applied doses of ferric sulfate help to efficiently remove Dolichospermum, Microcystis, and Aphanizomenon. Aphanothece and Coelosphaerium had a lower removal rate in mesocosms with a dose of 20 mgFe/L in PLSF;
- Intracellular microcystins (MC-RR, MC-YR, MC-HtyR, MC-LR, MC-HilR, MC-WR, MC-LA, MC-LY, MC-LW, MC-LF, [Asp3]MC-RR and [Asp3]MC-LR), anatoxin-a (ANA-a), cylindrospermopsin (CYN), anabaenopeptin A (AP-A) and anabaenopeptin B (AP-B) were detected throughout the experiment;
- Ferric sulfate effectively removes detected intracellular cyanotoxins but not extracellular cyanotoxins. More than 97% removal of total intracellular microcystins were achieved for both applied doses;
- Different dosages of ferric sulfate have almost the same effectiveness in removing intra- and extra-cellular cyanotoxins meaning that source water treatment will not be highly sensitive to suboptimal dosing.
4. Material and Methods
4.1. Study Site Description
4.2. Mesocosms Experiments Description
4.3. Preparation and Application of Chemical Coagulant
4.4. Sampling and Filtration Procedure
4.5. Analysis Methods
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paerl, H.W.; Paul, V.J. Climate change: Links to global expansion of harmful cyanobacteria. Water Res. 2012, 46, 1349–1363. [Google Scholar] [CrossRef] [PubMed]
- Merel, S.; Walker, D.; Chicana, R.; Snyder, S.; Baurès, E.; Thomas, O. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ. Int. 2013, 59, 303–327. [Google Scholar] [CrossRef]
- Svirčev, Z.; Lalić, D.; Savić, G.B.; Tokodi, N.; Backović, D.D.; Chen, L.; Meriluoto, J.; Codd, G.A. Global geographical and historical overview of cyanotoxin distribution and cyanobacterial poisonings. Arch. Toxicol. 2019, 93, 2429–2481. [Google Scholar] [CrossRef]
- Carmichael, W. Cyanobacteria secondary metabolites-the cyanotoxins. J. Appl. Bacteriol. 1992, 72, 445–459. [Google Scholar] [CrossRef]
- Harke, M.J.; Steffen, M.M.; Gobler, C.J.; Otten, T.G.; Wilhelm, S.; Wood, S.A.; Paerl, H.W. A review of the global ecology, genomics, and biogeography of the toxic cyanobacterium, Microcystis spp. Harmful Algae 2016, 54, 4–20. [Google Scholar] [CrossRef] [Green Version]
- Kotak, B.G.; Zurawell, R. Cyanobacterial toxins in Canadian freshwaters: A review. Lake Reserv. Manag. 2007, 23, 109–122. [Google Scholar] [CrossRef]
- Lévesque, B.; Gervais, M.-C.; Chevalier, P.; Gauvin, D.; Anassour-Laouan-Sidi, E.; Gingras, S.; Fortin, N.; Brisson, G.; Greer, C.; Bird, D. Prospective study of acute health effects in relation to exposure to cyanobacteria. Sci. Total Environ. 2014, 466, 397–403. [Google Scholar] [CrossRef]
- Havens, K. Cyanobacteria blooms: Effects on aquatic ecosystems. Cyanobacterial Harmful Algal Bloom. State Sci. Res. Needs 2008, 619, 733–747. [Google Scholar]
- Zamyadi, A.; MacLeod, S.L.; Fan, Y.; McQuaid, N.; Dorner, S.; Sauvé, S.; Prévost, M. Toxic cyanobacterial breakthrough and accumulation in a drinking water plant: A monitoring and treatment challenge. Water Res. 2012, 46, 1511–1523. [Google Scholar] [CrossRef] [PubMed]
- Cooke, G.D.; Welch, E.B.; Peterson, S.; Nichols, S.A. Restoration and Management of Lakes and Reservoirs; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Lürling, M.; van Oosterhout, F. Controlling eutrophication by combined bloom precipitation and sediment phosphorus inactivation. Water Res. 2013, 47, 6527–6537. [Google Scholar] [CrossRef] [PubMed]
- Lelkova, E.; Rulík, M.; Hekera, P.; Dobias, P.; Dolejs, P.; Borovickova, M.; Poulickova, A. The influence of the coagulant PAX-18 on Planktothrix agardhii blow in a shallow eutrophic fishpond. Fottea 2008, 8, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Sun, F.; Hu, W.; Pei, H.; Li, X.; Xu, X.; Ma, C. Evaluation on the dewatering process of cyanobacteria-containing AlCl3 and PACl drinking water sludge. Sep. Purif. Technol. 2015, 150, 52–62. [Google Scholar] [CrossRef]
- Sun, F.; Pei, H.-Y.; Hu, W.-R.; Li, X.-Q.; Ma, C.-X.; Pei, R.-T. The cell damage of Microcystis aeruginosa in PACl coagulation and floc storage processes. Sep. Purif. Technol. 2013, 115, 123–128. [Google Scholar] [CrossRef]
- Sun, F.; Pei, H.-Y.; Hu, W.-R.; Ma, C.-X. The lysis of Microcystis aeruginosa in AlCl3 coagulation and sedimentation processes. Chem. Eng. J. 2012, 193–194, 196–202. [Google Scholar] [CrossRef]
- Jančula, D.; Maršálek, B. Critical review of actually available chemical compounds for prevention and management of cyanobacterial blooms. Chemosphere 2011, 85, 1415–1422. [Google Scholar] [CrossRef]
- Orihel, D.; Schindler, D.W.; Ballard, N.C.; Wilson, L.R.; Vinebrooke, R. Experimental iron amendment suppresses toxic cyanobacteria in a hypereutrophic lake. Ecol. Appl. 2016, 26, 1517–1534. [Google Scholar] [CrossRef] [PubMed]
- Molot, L.A.; Li, G.; Findlay, D.L.; Watson, S.B. Iron-Mediated suppression of bloom-forming cyanobacteria by oxine in a eutrophic lake. Freshw. Biol. 2010, 55, 1102–1117. [Google Scholar] [CrossRef]
- Sevilla, E.; Martin-Luna, B.; Vela, L.; Bes, M.T.; Fillat, M.F.; Peleato, M.L. Iron availability affects mcyD expression and microcystin-LR synthesis in Microcystis aeruginosa PCC7806. Environ. Microbiol. 2008, 10, 2476–2483. [Google Scholar] [CrossRef] [PubMed]
- Zamyadi, A.; Dorner, S.; Sauvé, S.; Ellis, D.; Bolduc, A.; Bastien, C.; Prévost, M. Species-Dependence of cyanobacteria removal efficiency by different drinking water treatment processes. Water Res. 2013, 47, 2689–2700. [Google Scholar] [CrossRef] [PubMed]
- McQuaid, N.; Zamyadi, A.; Prevost, M.; Bird, D.F.; Dorner, S. Use of in vivophycocyanin fluorescence to monitor potential microcystin-producing cyanobacterial biovolume in a drinkingwater source. J. Environ. Monit. 2011, 13, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Jalili, F.; Trigui, H.; Maldonado, J.F.G.; Dorner, S.; Zamyadi, A.; Shapiro, B.J.; Terrat, Y.; Fortin, N.; Sauvé, S.; Prévost, M. Can Cyanobacterial Diversity in the Source Predict the Diversity in Sludge and the Risk of Toxin Release in a Drinking Water Treatment Plant? Toxins 2021, 13, 25. [Google Scholar] [CrossRef] [PubMed]
- Monchamp, M.-E.; Pick, F.R.; Beisner, B.E.; Maranger, R. Nitrogen Forms Influence Microcystin Concentration and Composition via Changes in Cyanobacterial Community Structure. PLoS ONE 2014, 9, e85573. [Google Scholar] [CrossRef] [PubMed]
- Bowling, L.C.; Blais, S.; Sinotte, M. Heterogeneous spatial and temporal cyanobacterial distributions in Missisquoi Bay, Lake Champlain: An analysis of a 9 year data set. J. Great Lakes Res. 2015, 41, 164–179. [Google Scholar] [CrossRef]
- International Joint Commission Report. Nutrient Loading and Impacts in Lake Champlain, Missisquoi Bay, and the Richelieu River. Available online: https://ijc.org/sites/default/files/2019-11/20191114_Missisquoi%20Bay%20Lit%20Review_LCBP_OBVBM_FR.pdf (accessed on 13 September 2021).
- Davis, T.; Harke, M.; Marcoval, M.; Goleski, J.; Orano-Dawson, C.; Berry, D.; Gobler, C. Effects of nitrogenous compounds and phosphorus on the growth of toxic and non-toxic strains of Microcystis during cyanobacterial blooms. Aquat. Microb. Ecol. 2010, 61, 149–162. [Google Scholar] [CrossRef] [Green Version]
- Schindler, D.W.; Hecky, R.E.; Findlay, D.L.; Stainton, M.P.; Parker, B.R.; Paterson, M.J.; Beaty, K.G.; Lyng, M.; Kasian, S.E.M. Eutrophication of lakes cannot be controlled by reducing nitrogen input: Results of a 37-year whole-ecosystem experiment. Proc. Natl. Acad. Sci. USA 2008, 105, 11254–11258. [Google Scholar] [CrossRef] [Green Version]
- Paerl, H. Nuisance Phytoplankton Blooms in Coastal, Estuarine, and Inland Waters. Limnol. Oceanogr. 1988, 33, 823–843. [Google Scholar] [CrossRef]
- Degerholm, J.; Gundersen, K.; Bergman, B.; Söderbäck, E. Phosphorus-Limited growth dynamics in two Baltic Sea cyanobacteria, Nodularia sp. and Aphanizomenon sp. FEMS Microbiol. Ecol. 2006, 58, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Dolinova, I.; Sevcu, A.; Jurczak, T.; Frankiewicz, P.; Wojtal-Frankiewicz, A.; Wan, L.; Deng, Q.; Song, C.; Zhou, Y.; et al. Strategies adopted by Aphanizomenon flos-aquae in response to phosphorus deficiency and their role on growth. Environ. Sci. Eur. 2020, 32, 1–13. [Google Scholar] [CrossRef]
- Saad, A.; Atia, A. Review on Freshwater Blue-Green Algae (Cyanobacteria): Occurrence, Classification and Toxicology. Biosci. Biotechnol. Res. Asia 2014, 11, 1319–1325. [Google Scholar] [CrossRef]
- Crayton, M. Toxic cyanobacteria blooms. In A Field/Laboratory Guide; Office of Environmenta1 Health Assessments Washington, Department of Health: Olympia, WA, USA, 2004. [Google Scholar]
- de Maagd, P.-J.; Hendriks, A.; Seinen, W.; Sijm, D.T. pH-Dependent hydrophobicity of the cyanobacteria toxin microcystin-LR. Water Res. 1999, 33, 677–680. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Nakahara, H. The formation and degradation of cyanobacterium Aphanizomenon flos-aquae blooms: The importance of pH, water temperature, and day length. Limnology 2005, 6, 1–6. [Google Scholar] [CrossRef]
- Li, H.; Alsanea, A.; Barber, M.; Goel, R. High-Throughput DNA sequencing reveals the dominance of pico- and other filamentous cyanobacteria in an urban freshwater Lake. Sci. Total Environ. 2019, 661, 465–480. [Google Scholar] [CrossRef] [PubMed]
- Henderson, R.; Parsons, S.A.; Jefferson, B. Successful Removal of Algae through the Control of Zeta Potential. Sep. Sci. Technol. 2008, 43, 1653–1666. [Google Scholar] [CrossRef]
- Henderson, R.; Parsons, S.A.; Jefferson, B. The impact of differing cell and algogenic organic matter (AOM) characteristics on the coagulation and flotation of algae. Water Res. 2010, 44, 3617–3624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takaara, T.; Sano, D.; Konno, H.; Omura, T. Affinity isolation of algal organic matters able to form complex with aluminium coagulant. Water Supply 2004, 4, 95–102. [Google Scholar] [CrossRef]
- Pivokonsky, M.; Kloucek, O.; Pivokonska, L. Evaluation of the production, composition and aluminum and iron complexation of algogenic organic matter. Water Res. 2006, 40, 3045–3052. [Google Scholar] [CrossRef] [PubMed]
- Henderson, R.; Parsons, S.A.; Jefferson, B. The impact of algal properties and pre-oxidation on solid–liquid separation of algae. Water Res. 2008, 42, 1827–1845. [Google Scholar] [CrossRef]
- Bernhardt, H.; Clasen, J. Flocculation of Micro-Organisms; Water Supply Res. Technol.-AQUA: London, UK, 1991; Volume 40, pp. 76–87. [Google Scholar]
- Algal cells and coagulation, flocculation and sedimentation processes. Water Sci. Technol. 1997, 36, 111–118. [CrossRef]
- Gonzalez-Torres, A.; Putnam, J.; Jefferson, B.; Stuetz, R.; Henderson, R. Examination of the physical properties of Microcystis aeruginosa flocs produced on coagulation with metal salts. Water Res. 2014, 60, 197–209. [Google Scholar] [CrossRef]
- Xu, H.; Pei, H.; Xiao, H.; Jin, Y.; Li, X.; Hu, W.; Ma, C.; Sun, J.; Li, H. Behaviors of Microcystis aeruginosa cells during floc storage in drinking water treatment process. Sci. Rep. 2016, 6, 34943. Available online: https://www.nature.com/articles/srep34943#supplementary-information (accessed on 9 August 2020). [CrossRef] [Green Version]
- Načeradská, J.; Pivokonska, L.; Pivokonský, M. On the importance of pH value in coagulation. J. Water Supply Res. Technol. 2019, 68, 222–230. [Google Scholar] [CrossRef]
- Zamyadi, A.; Dorner, S.; Ndong, M.; Ellis, D.; Bolduc, A.; Bastien, C.; Prévost, M. Low-Risk cyanobacterial bloom sources: Cell accumulation within full-scale treatment plants. J. Am. Water Work. Assoc. 2013, 105, E651–E663. [Google Scholar] [CrossRef] [Green Version]
- Fortin, N.; Aranda-Rodriguez, R.; Jing, H.; Pick, F.; Bird, D.; Greer, C.W. Detection of Microcystin-Producing Cyanobacteria in Missisquoi Bay, Quebec, Canada, Using Quantitative PCR. Appl. Environ. Microbiol. 2010, 76, 5105–5112. [Google Scholar] [CrossRef] [Green Version]
- Giani, A.; Bird, D.F.; Prairie, Y.; Lawrence, J.F. Empirical study of cyanobacterial toxicity along a trophic gradient of lakes. Can. J. Fish. Aquat. Sci. 2005, 62, 2100–2109. [Google Scholar] [CrossRef]
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management/Edited by Ingrid Chorus and Jamie Bertram; Taylor & Francis: Abingdon, UK, 1999. [Google Scholar]
- Pineda-Mendoza, R.; Zúñiga, G.; Martínez-Jerónimo, F. Microcystin production in Microcystis aeruginosa: Effect of type of strain, environmental factors, nutrient concentrations, and N:P ratio on mcyA gene expression. Aquat. Ecol. 2015, 50, 103–119. [Google Scholar] [CrossRef]
- Kaebernick, M.; Neilan, B.A.; Börner, T.; Dittmann, E. Light and the Transcriptional Response of the Microcystin Biosynthesis Gene Cluster. Appl. Environ. Microbiol. 2000, 66, 3387–3392. [Google Scholar] [CrossRef] [Green Version]
- Preußel, K.; Wessel, G.; Fastner, J.; Chorus, I. Response of cylindrospermopsin production and release in Aphanizomenon flos-aquae (Cyanobacteria) to varying light and temperature conditions. Harmful Algae 2009, 8, 645–650. [Google Scholar] [CrossRef]
- Rücker, J.; Stüken, A.; Nixdorf, B.; Fastner, J.; Chorus, I.; Wiedner, C. Concentrations of particulate and dissolved cylindrospermopsin in 21 Aphanizomenon-dominated temperate lakes. Toxicon 2007, 50, 800–809. [Google Scholar] [CrossRef]
- Büşra Şengül, A.; Ersan, G.; Tufekci, N. Removal of Intra- and Extracellular Microcystin by Submerged Ultrafiltration (UF) Membrane Combined with Coagulation/Flocculation and Powdered Activated Carbon (PAC) Adsorption. J. Hazard. Mater. 2017, 343, 29–35. [Google Scholar] [CrossRef]
- Dixon, M.B.; Richard, Y.; Ho, L.; Chow, C.; O’Neill, B.K.; Newcombe, G. A coagulation–powdered activated carbon–ultrafiltration—Multiple barrier approach for removing toxins from two Australian cyanobacterial blooms. J. Hazard. Mater. 2011, 186, 1553–1559. [Google Scholar] [CrossRef]
- Massey, I.Y.; Yang, F. A Mini Review on Microcystins and Bacterial Degradation. Toxins 2020, 12, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji, K.; Naito, S.; Kondo, F.; Ishikawa, N.; Watanabe, M.F.; Suzuki, M.; Harada, K.-I. Stability of microcystins from cyanobacteria: Effect of light on decomposition and isomerization. Environ. Sci. Technol. 1994, 28, 173–177. [Google Scholar] [CrossRef] [PubMed]
- de Figueiredo, D.; Azeiteiro, U.M.; Esteves, S.M.; Gonçalves, F.J.M.; Pereira, M.J. Microcystin-Producing blooms—A serious global public health issue. Ecotoxicol. Environ. Saf. 2004, 59, 151–163. [Google Scholar] [CrossRef]
- Harada, K.-I.; Tsuji, K.; Watanabe, M.F.; Kondo, F. Stability of microcystins from cyanobacteria—III. Effect of pH and temperature. Phycologia 1996, 35, 83–88. [Google Scholar] [CrossRef]
- Dinh, Q.T.; Munoz, G.; Simon, D.F.; Duy, S.V.; Husk, B.; Sauvé, S. Stability issues of microcystins, anabaenopeptins, anatoxins, and cylindrospermopsin during short-term and long-term storage of surface water and drinking water samples. Harmful Algae 2020, 101, 101955. [Google Scholar] [CrossRef]
- Maghsoudi, E.; Fortin, N.; Greer, C.; Duy, S.V.; Fayad, P.; Sauvé, S.; Prévost, M.; Dorner, S. Biodegradation of multiple microcystins and cylindrospermopsin in clarifier sludge and a drinking water source: Effects of particulate attached bacteria and phycocyanin. Ecotoxicol. Environ. Saf. 2015, 120, 409–417. [Google Scholar] [CrossRef]
- Han, J.; Jeon, B.-S.; Futatsugi, N.; Park, H.-D. The effect of alum coagulation for in-lake treatment of toxic Microcystis and other cyanobacteria related organisms in microcosm experiments. Ecotoxicol. Environ. Saf. 2013, 96, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Pei, H.; Xu, H.; Jin, Y.; Sun, J. Behavior of Cylindrospermopsis raciborskii during coagulation and sludge storage—Higher potential risk of toxin release than Microcystis aeruginosa? J. Hazard. Mater. 2018, 347, 307–316. [Google Scholar] [CrossRef]
- Ho, L.; Dreyfus, J.; Boyer, J.; Lowe, T.; Bustamante, H.; Duker, P.; Meli, T.; Newcombe, G. Fate of cyanobacteria and their metabolites during water treatment sludge management processes. Sci. Total Environ. 2012, 424, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Dreyfus, J.; Monrolin, Y.; Pestana, C.; Reeve, P.J.; Sawade, E.; Newton, K.; Ho, L.; Chow, C.W.K.; Newcombe, G. Identification and assessment of water quality risks associated with sludge supernatant recycling in the presence of cyanobacteria. J. Water Supply Res. Technol. AQUA 2016, 65, 441–452. [Google Scholar] [CrossRef]
- Maghsoudi, E.; Prévost, M.; Duy, S.V.; Sauvé, S.; Dorner, S. Adsorption characteristics of multiple microcystins and cylindrospermopsin on sediment: Implications for toxin monitoring and drinking water treatment. Toxicon 2015, 103, 48–54. [Google Scholar] [CrossRef]
- Jones, G.J.; Negri, A.P. Persistence and degradation of cyanobacterial paralytic shellfish poisons (PSPs) in freshwaters. Water Res. 1997, 31, 525–533. [Google Scholar] [CrossRef]
- Chen, W.; Song, L.; Peng, L.; Wan, N.; Zhang, X.; Gan, N. Reduction in microcystin concentrations in large and shallow lakes: Water and sediment-interface contributions. Water Res. 2008, 42, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Wood, S. Increasing Microcystis cell density enhances microcystin synthesis: A mesocosm study. Inland Waters 2012, 2, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Edler, L.; Elbrächter, M. The Utermöhl method for quantitative phytoplankton analysis. Microsc. Mol. Methods Quant. Phytoplankton Anal. 2010, 110, 13–20. [Google Scholar]
- Lund, J.W.G. A Simple Counting Chamber for Nannoplankton1. Limnol. Oceanogr. 1959, 4, 57–65. [Google Scholar] [CrossRef]
- Munoz, G.; Duy, S.V.; Roy-Lachapelle, A.; Husk, B.; Sauvé, S. Analysis of individual and total microcystins in surface water by on-line preconcentration and desalting coupled to liquid chromatography tandem mass spectrometry. J. Chromatogr. A 2017, 1516, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Roy-Lachapelle, A.; Duy, S.V.; Munoz, G.; Dinh, Q.T.; Bahl, E.; Simon, D.F.; Sauvé, S. Analysis of multiclass cyanotoxins (microcystins, anabaenopeptins, cylindrospermopsin and anatoxins) in lake waters using on-line SPE liquid chromatography high-resolution Orbitrap mass spectrometry. Anal. Methods 2019, 11, 5289–5300. [Google Scholar] [CrossRef] [Green Version]
- United States Environmental Protection Agency (USEPA). Method 415.1. Organic Carbon, Total (Combustion or Oxidation); United States Environmental Protection Agency: Washington, DC, USA, 1974; pp. 1–3.
- United States Environmental Protection Agency (USEPA). Method 353.2, Revision 2.0: Determination of Nitrate-Nitrite Nitrogen by Automated Colorimetry; United States Environmental Protection Agency: Washington, DC, USA, 1993; pp. 1–15.
- United States Environmental Protection Agency (USEPA). Method 365.3: Phosphorous, All Forms (Colorimetric, Ascorbic Acid, Two Reagent); United States Environmental Protection Agency: Washington, DC, USA, 1978; pp. 1–5.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Missisquoi Bay | Petit Lac St. François | ||||
|---|---|---|---|---|---|---|
| (A) 10 September 2018 | (B) 24 September 2018 | (C) 13 August 2019 | (a) 26 June 2019 | (b) 24 July 2019 | (c) 05 August 2019 | |
| Total cell counts (cells/mL) | 998,183 35,034 | 547,325 16,578 | 17,408,158 138,898 | 6,033,197 316,425 | 109,193 3578 | 235,723 5986 |
| Chlorophyll-a (RFU) | - | - | 62.22 1.03 | 3.97 0.04 | 5.14 0.06 | 6.27 0.08 |
| Phycocyanin (RFU) | - | - | 93.43 0.52 | 16.51 1.54 | 1.77 0.04 | 4.83 0.13 |
| pH | 6.5 0.08 | 6.4 0.07 | 8.05 0.24 | 8.08 0.07 | 7.84 0.03 | 8.01 0.01 |
| TDS (mg/L) | 101 0.00 | 100 0.00 | 98.00 0.00 | 122.50 0.71 | 116.00 0.00 | 115.00 0.00 |
| Temp (°C) | 21.8 0.01 | 18.7 0.12 | 26.89 0.21 | 25.96 0.49 | 25.14 0.12 | 24.43 0.24 |
| TOC (mg C/L) | 15.22 0.25 | 5.55 0.07 | 885.00 33.34 | 175.00 9.50 | 11.10 0.38 | 10.76 0.13 |
| DOC (mg C/L) | 7.50 0.05 | 5.00 0.00 | 19.34 1.67 | 9.80 0.11 | 9.83 0.13 | 11.34 1.58 |
| TN (mg N/L) | 5.55 0.65 | 2.75 0.08 | 7.84 2.95 | 12.85 0.57 | 1.11 0.05 | 1.29 0.008 |
| TP (μg P/L) | 360.51 0.01 | 292.11 13.28 | 4336.92 74.65 | 723.55 24.97 | 110.38 1.89 | 131.78 7.71 |
| DN (mg N/L) | 0.45 0.004 | 0.52 0.009 | 2.66 0.12 | 0.73 0.08 | 0.60 0.02 | 0.50 0.02 |
| DP (μg P/L) | 17.05 0.07 | 17.99 0.45 | 302.17 12.98 | 139.06 3.35 | 23.92 4.91 | 42.86 1.13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le, K.T.N.; Goitom, E.; Trigui, H.; Sauvé, S.; Prévost, M.; Dorner, S. The Effects of Ferric Sulfate (Fe2(SO4)3) on the Removal of Cyanobacteria and Cyanotoxins: A Mesocosm Experiment. Toxins 2021, 13, 753. https://doi.org/10.3390/toxins13110753
Le KTN, Goitom E, Trigui H, Sauvé S, Prévost M, Dorner S. The Effects of Ferric Sulfate (Fe2(SO4)3) on the Removal of Cyanobacteria and Cyanotoxins: A Mesocosm Experiment. Toxins. 2021; 13(11):753. https://doi.org/10.3390/toxins13110753
Chicago/Turabian StyleLe, Kim Thien Nguyen, Eyerusalem Goitom, Hana Trigui, Sébastien Sauvé, Michèle Prévost, and Sarah Dorner. 2021. "The Effects of Ferric Sulfate (Fe2(SO4)3) on the Removal of Cyanobacteria and Cyanotoxins: A Mesocosm Experiment" Toxins 13, no. 11: 753. https://doi.org/10.3390/toxins13110753