Removal of Aflatoxin B1 by Edible Mushroom-Forming Fungi and Its Mechanism

1
Department of Integrated Biomedical and Life Science, Korea University, Seoul 02841, Korea
2
Department of Food and Nutrition, Hanyang University, Seoul 04763, Korea
*
Author to whom correspondence should be addressed.
†
Equal contribution.
Toxins 2021, 13(9), 668; https://doi.org/10.3390/toxins13090668
Submission received: 30 August 2021
/
Revised: 15 September 2021
/
Accepted: 16 September 2021
/
Published: 18 September 2021
(This article belongs to the Special Issue Determination and Detoxification Strategies of Mycotoxins)
Abstract
:Aflatoxins (AFs) are biologically active toxic metabolites, which are produced by certain toxigenic Aspergillus sp. on agricultural crops. In this study, five edible mushroom-forming fungi were analyzed using high-performance liquid chromatography fluorescence detector (HPLC-FLD) for their ability to remove aflatoxin B1 (AFB1), one of the most potent naturally occurring carcinogens known. Bjerkandera adusta and Auricularia auricular-judae showed the most significant AFB1 removal activities (96.3% and 100%, respectively) among five strains after 14-day incubation. The cell lysate from B. adusta exhibited higher AFB1 removal activity (35%) than the cell-free supernatant (13%) after 1-day incubation and the highest removal activity (80%) after 5-day incubation at 40 °C. In addition, AFB1 analyses using whole cells, cell lysates, and cell debris from B. adusta showed that cell debris had the highest AFB1 removal activity at 5th day (95%). Moreover, exopolysaccharides from B. adusta showed an increasing trend (24–48%) similar to whole cells and cell lysates after 5- day incubation. Our results strongly suggest that AFB1 removal activity by whole cells was mainly due to AFB1 binding onto cell debris during early incubation and partly due to binding onto cell lysates along with exopolysaccharides after saturation of AFB1 binding process onto cell wall components.
Key Contribution: AFB1 removal mechanism by an edible mushroom-forming fungus Bjerkandera adusta was revealed in this study. Our data suggest that after AFB1 binds onto cell debris of B. adusta during early incubation, it binds onto cell lysates along with exopolysaccharides when the AFB1 binding process onto cell wall components is saturated.
1. Introduction
Aflatoxins (AFs) are a group of highly toxic secondary metabolites, which are produced by certain toxigenic Aspergillus species (A. flavus, A. parasiticus, and A. nominus) commonly found in crops such as cotton seed, tree nuts, corn, and peanuts [1]. There are four major types of AFs: AFB1, AFB2, AFG1, and AFG2. Of these AFs, AFB1 is the most potent carcinogen [2]. After AFB1 is bioactivated to AFB1-8,9-epoxide by cytochrome P450 (CYP450) in liver, it forms adducts at N7 guanine residues on DNA. This can cause hepatotoxicity, teratogenicity, immunotoxicity, and carcinogenicity in human and animals [3]. The International Agency for Research on Cancer (IARC) classified AFB1 as a group 1 human carcinogen [4]. Due to high AFB1 contamination of food and feed, many efforts have been sought to reduce or eliminate AFB1 in them. Physical methods such as use of microwave, UV irradiation, and absorbent materials as well as chemical methods such as use of ozone, bisulfite, and ammonia have been proposed. However, these two methods limit their application to food manufacturing systems because they could reduce the nutritional value of food and alter the food quality, causing undesirable health effects [5,6,7,8,9,10,11,12,13]. On the other hand, biological methods using microorganisms or their enzymes offer the most promising alternatives for detoxification of AFs in food and feed. The microbial degradation of AFs takes some advantages such as utilization of specific reactions and mild reaction conditions to detoxify AFs to less or non-toxic metabolites [14,15]. Many studies have reported degradation of AFs by bacteria and fungi [8,16,17,18,19,20]. Shantha (1999) showed that some fungi (Rhizopus sp., Trichoderma sp., Phoma sp., Sporotrichum sp., and Alternaria sp.) were able to degrade AFB1 [18]. Interestingly, several researchers have documented that white rot fungi have the potential to degrade lignin or polycyclic aromatic hydrocarbons including AFs by their enzymes such as laccases and peroxidases [21,22,23]. Alberts and colleagues reported biodegradation of AFB1 through oxidation of phenolic compounds by laccase, a low specific enzyme, from white rot fungi including Trametes versicolor [24]. Wang and collaborators showed AFB1 detoxification by Mn-peroxidase from Phanerochaete sodida strain [20]. Although biodegradation mechanisms were not identified, Motomura and co-workers isolated and purified an AFs-degrading enzyme from an edible mushroom Pleurotus ostreatus [25]. Yehia showed that Mn-peroxidase from P. ostreatus was able to detoxify AFB1 [26]. However, some problems still remain regarding practical applications of AFs biodegradation in the food industry. The microorganisms that have AFs degradation activity must be safe and should not produce undesirable byproducts or adverse effects on the quality of the foods. Another biological method for reduction of AFs in food and feed is elimination of the toxin by adhesion or adsorption using several microorganisms such as lactic acid bacteria or yeasts [27,28]. A number of studies have shown that several different strains of lactic acid bacteria such as Lactobacillus sp., Lactococcus sp., and Bifidobacterium sp. can reduce levels of AFs in food and feed by a binding process onto their cell wall components [29,30,31,32,33]. Other researchers have also reported binding of AFs by yeast such as Saccharomyces cerevisiae [34,35,36,37,38].
In this study, we evaluated possibilities of AFB1 biodegradation by edible mushroom-forming fungi collected from South Korea and investigated the mechanisms of the AFB1 elimination in the process. In particular, the ability of Bjerkandera adusta to remove AFB1 was analyzed after enzymatic, physical, and chemical treatments to degrade or change the fungal cellular components in order to better understand the role of fungal components in AFB1 removal activity. Our data suggest that AFB1 was removed by its binding onto cell wall components of B. adusta. To the best of our knowledge, this is the first report on AFB1 binding activity and mechanism by edible mushroom-forming fungi.
2. Results
2.1. Time Course of Fungal Growth, pH, and Removal of AFB1 Using Five Edible Mushroom-Forming Fungi
Five edible mushroom-forming fungi (B. adusta, Auricularia auricular-judae, Lentinula edodes, Hericium erinaceus, and Poria cocos) in Basidiomycota, which had been obtained from mushroom farms in Gyunggi province in South Korea, were tested for possibilities of AFB1 biodegradation.
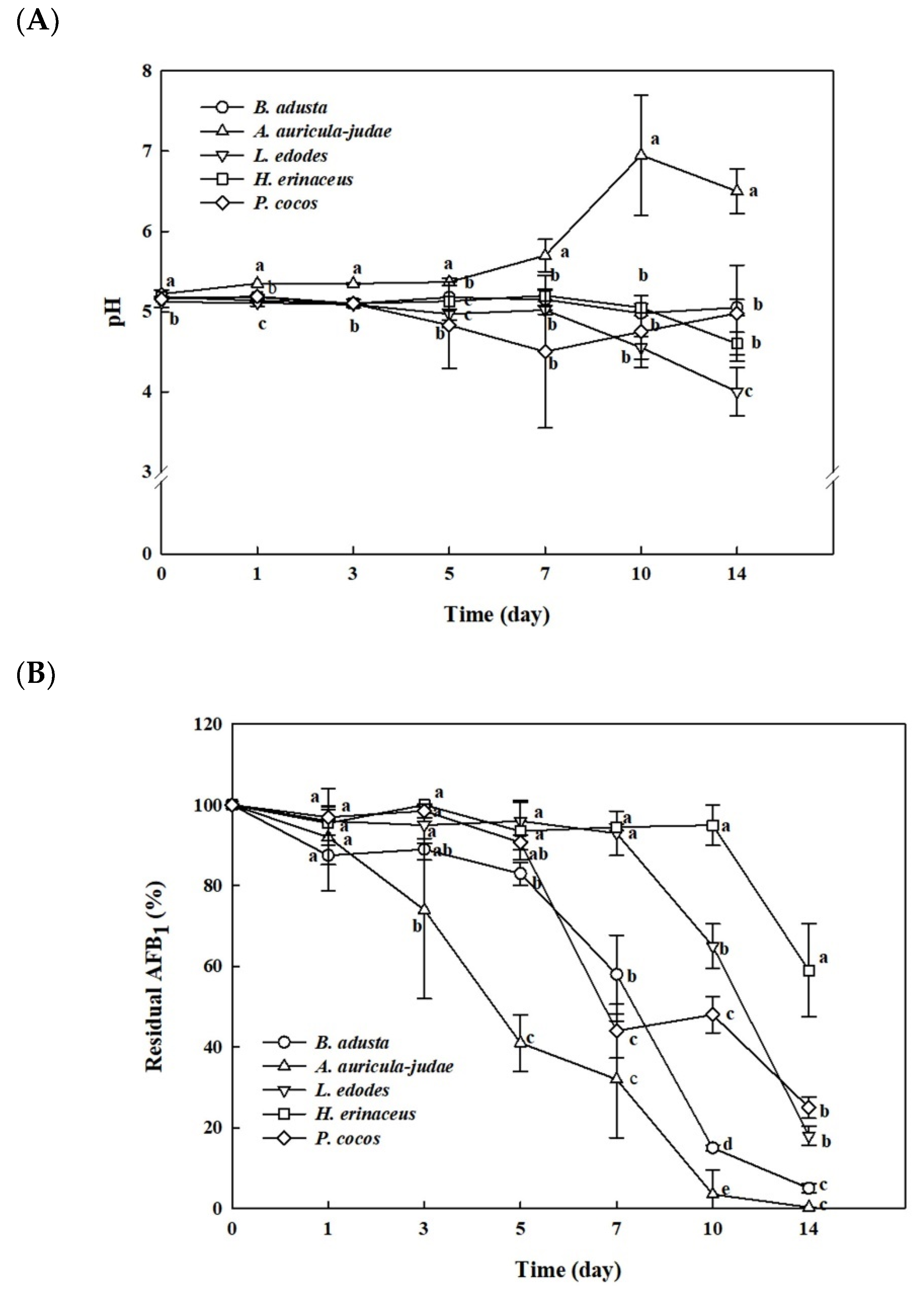
The fungal stains were grown in 30 mL of potato dextrose broth (PDB) containing 50 ng/mL of AFB1 for 14 days. All of the cultures were maintained at pH 5 until the 5th day (Figure 1A). After 5-day incubation, the pH of H. erinaceus and L. edodes cultures were decreased to 4 and 4.5, respectively, while the pH of A. auricular-judae culture showed a rapid increase from 5 to 6–7. The pH of both B. adusta and P. cocos was continuously maintained at pH 5 for 14 days. The high-performance liquid chromatography fluorescence detector (HPLC-FLD) results indicated that all five strains decreased the levels of AFB1 in the media throughout the incubation. B. adusta and A. auricular-judae showed the most significant AFB1 removal activities (96.3% and 100% AFB1 reduction, respectively) among five strains after 14 days (Figure 1B). A. auricular-judae removed AFB1 in the medium more rapidly after 1st day than other mushroom-forming fungi, whereas B. adusta showed a sharp decrease in levels of AFB1 between 5 and 10 days. On the other hand, L. edodes and P. cocos reduced the levels of AFB1 by 81.0% and 75.5%, respectively, after 14 days, whereas H. erinaceus removed AFB1 by only 42.2%.
The colony diameters of five mushroom-forming fungi were measured for their growth rates on potato dextrose agar (PDA) plates. B. adusta showed fast growth until 4th day and maximum growth on 5th day, while others grew slowly until the 7th day (data not shown).
2.2. Test for Degradation of Remazol Brilliant Blue R (RBBR) and Coumarin by Mushroom-Forming Fungi
It has been reported that white rot fungi such as L. edodes, Bjerkandera sp., and T. versicolor produced Mn peroxidases or laccases as extracellular ligninolytic enzymes, which mediate degradation of recalcitrant phenolic compounds [24,39,40,41,42]. In order to investigate the mechanisms of AFB1 detoxification by five edible mushroom-forming fungi, RBBR dye or coumarin was used as the sole carbon source in fungal cultures, since the RBBR decolorization method has been used as a fast screening assay to identify potential ligninolytic fungi, and coumarin is the basic molecular structure of all AFs (bisfuranocoumarin derivatives) including AFB1 [22,43,44,45]. After 7- to 10-day incubation, only B. adusta showed decolorization of dark blue RBBR agar plates to brown color plates, suggesting that B. adusta has a capability to degrade lignin (data not shown). In case of coumarin agar plates, none of the five fungal strains was grown on the agar plate after 7- to 10-day incubation (data not shown). It suggests that all five mushroom-forming fungi were not able to use AFs as the sole carbon source.
2.3. AFB1 Removal by Cell-Free Supernatants and Cell Lysates
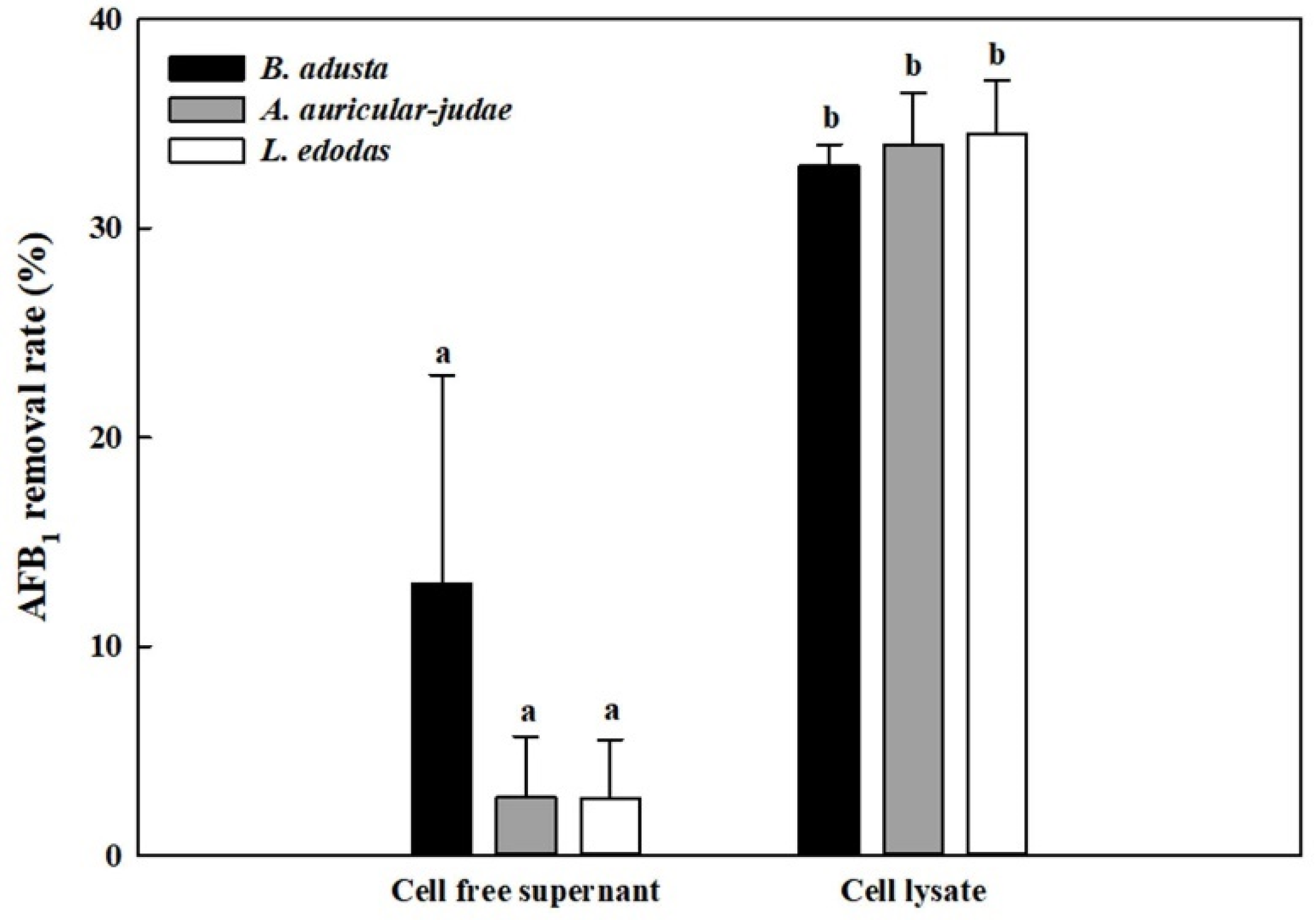
Since the results from time course and RBBR decolorization experiments suggest that mushroom-forming fungi may remove AFB1 by either extracellular enzymes or intracellular enzymes, we prepared cell-free supernatants and cell lysates from three mushroom-forming fungal cultures (B. adusta, A. auricular-judae, and L. edodes), which showed high AFB1 removal activities. Cell-free supernatants from B. adusta showed about 13% AFB1 removal activity after 1 day incubation, while those from A. auricular-judae and L. edodes showed about 3% of AFB1 removal activities (Figure 2). In contrast, cell lysates from all three mushroom-forming fungi exhibited higher AFB1 removal activities (about 35%) than those of cell-free supernatants at 1st day (p < 0.01). These data suggest that cell lysates from all three mushroom-forming fungi play a major role in AFB1 removal.
2.4. Effects of Different Reaction Temperatures on AFB1 Removal by Cell Lysates
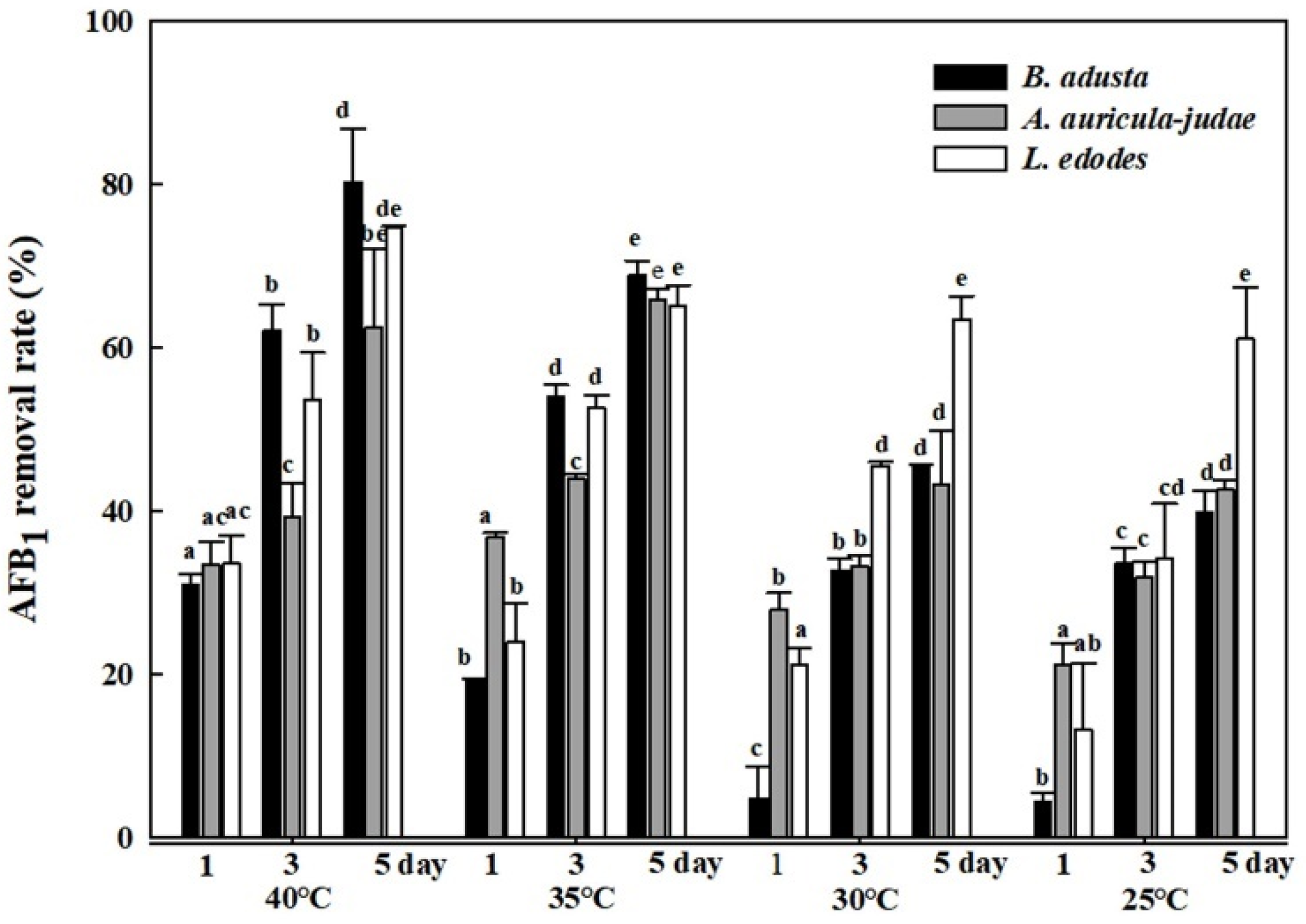
In order to investigate effects of reaction temperatures on AFB1 removal activity, cell lysates from the three mushroom-forming fungal cultures (B. adusta, A. auricular-judae, and L. edodes) were incubated with AFB1 for 1, 3, and 5 days at four different temperatures (25, 30, 35, and 40 °C). As incubation time becomes longer at the same temperature, cell lysates from all three mushroom-forming fungal cultures showed higher AFB1 removal activity. In addition, the cell lysates showed significant AFB1 removal activities (63% to 80%) at 40 °C after 5-day incubation, whereas they showed only 40% to 61% AFB1 removal activities at 25 °C after 5-day incubation (Figure 3). In particular, among cell lysates from the three mushroom-forming fungal cultures, the cell lysate from B. adusta culture exhibited higher removal activity (80%) than that from L. edodes culture (75%) or A. auricular-judae (60%) at 40 °C. These results suggest that B. adusta removes much more levels of AFB1 at 40 °C compared to the other two mushroom-forming fungi.
2.5. Effects of NADPH and NaIO4 on AFB1 Removal by Cell Lysates
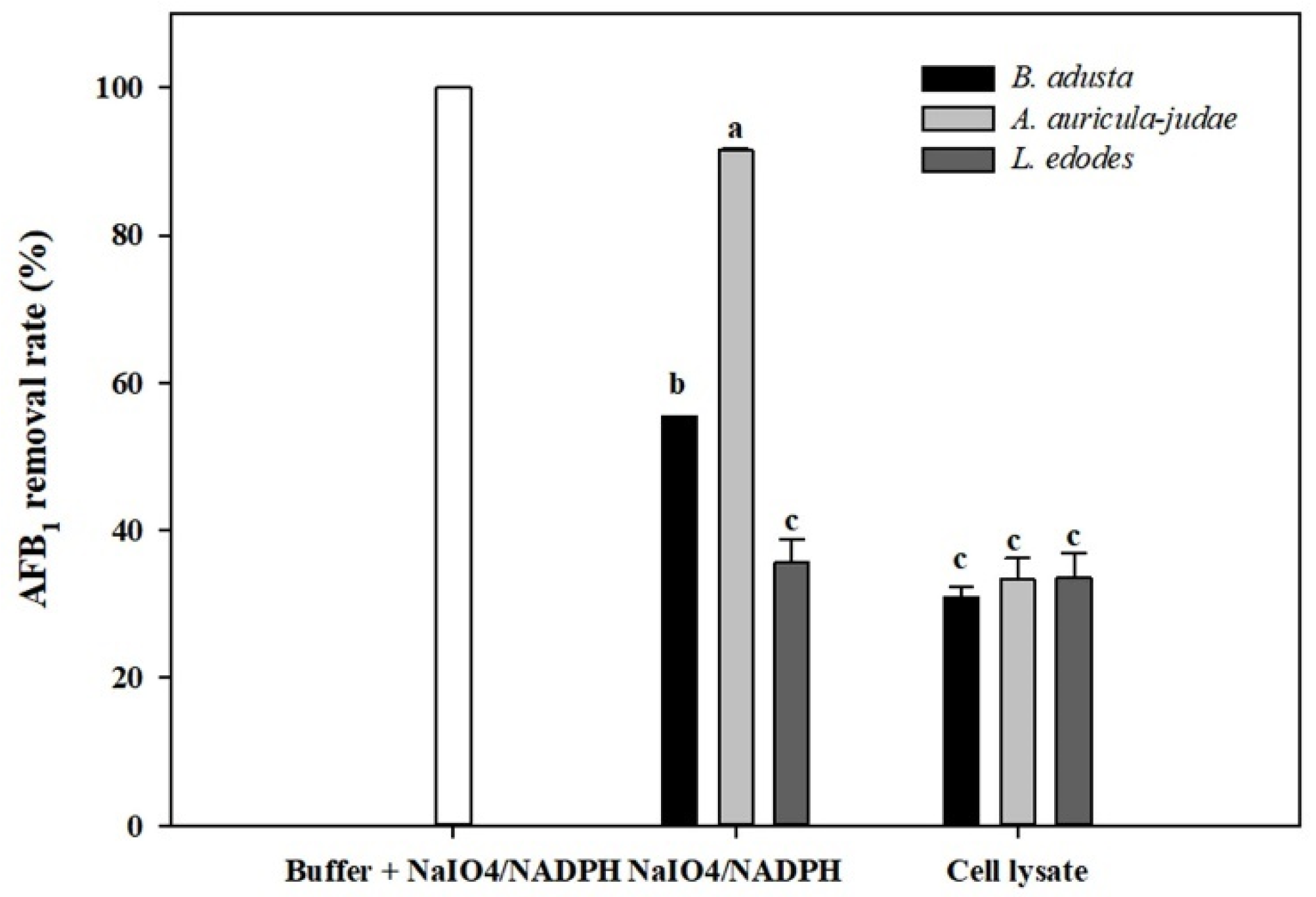
Previously, Hamid and co-worker reported that AFs degradation was enhanced by addition of NADPH and NaIO4 to cell-free extracts of A. flavus and that the AFs degradative activity may be involved in cytochrome P-450 monooxygenases [46]. Therefore, AFB1 removal activity was measured using cell lysates from three mushroom-forming fungal cultures (B. adusta, A. auricular-judae, and L. edodes) after addition of NaIO4 and NADPH to them. The NADPH- and NaIO4-treated cell lysates from all three mushroom-forming fungal cultures showed higher AFB1 removal activities than those without treatment after 2-day incubation (Figure 4). However, approximately 100% of AFB1 was degraded in a buffer solution to which only NaIO4 and NADPH were added without cell lysates. Thus, we concluded that the AFB1 removal activities in cell lysates including NaIO4 and NADPH were not due to enzyme activities in the lysate, but due to oxidation of AFB1 by NaIO4 and NADPH.
2.6. Effects of Heat or Proteinase Treatment on AFB1 Removal by Whole Cells and Cell Lysates from B. adusta Cultures
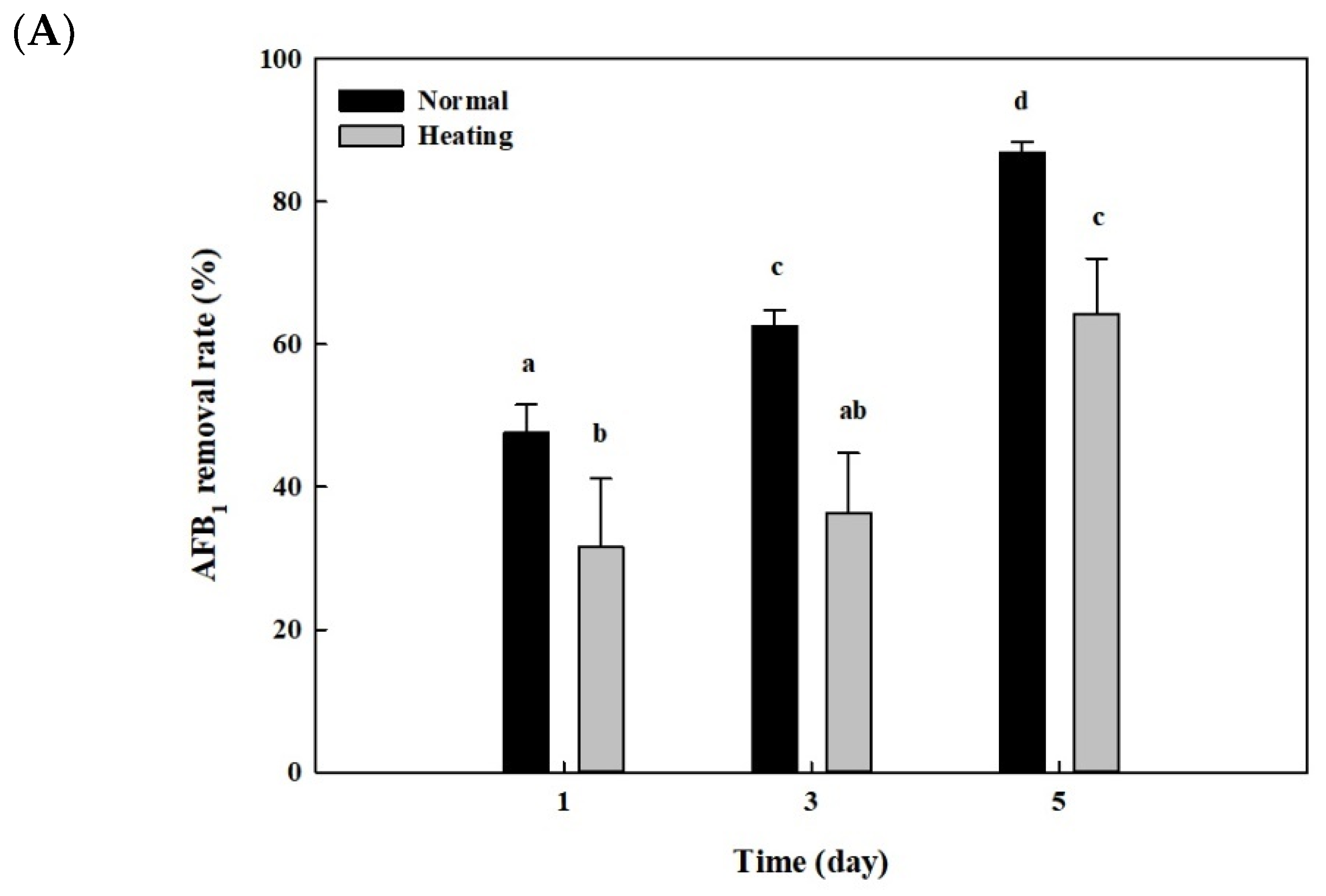
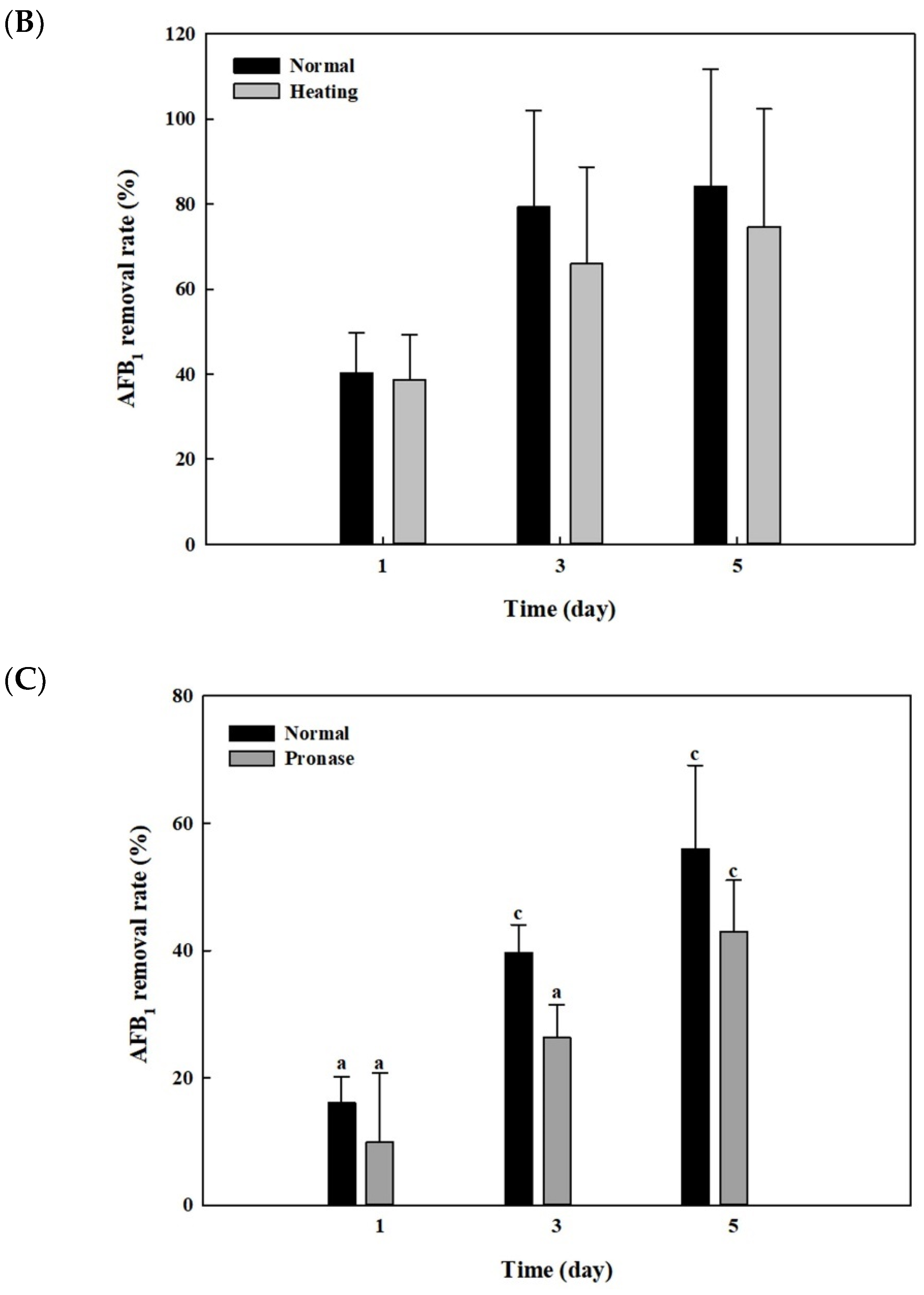
After whole cells and cell lysates from B. adusta culture were heat-treated at 121 °C for 15 min and 95 °C for 10 min, respectively, AFB1 quantification assays were performed to see whether the AFB1 removal activities shown in Figure 2 and Figure 3 were due to enzymes. The whole cells from B. adusta without heat treatment showed 87% of AFB1 removal activity at 5th day, while those after heat treatment showed 64% of AFB1 removal activity (Figure 5A). The AFB1 removal activity using cell lysates from B. adusta culture also had similar trends to that using whole cells. The cell lysates with or without heat treatment were 75% and 84% of AFB1 removal activities on the 5th day, respectively, which did not show statistically significant differences. (Figure 5B). These data suggest that heat treatment, which may cause enzyme inactivation by protein denaturation, did not affect much of the AFB1 removal activities in cell lysates.
In order to investigate effects of pronase E on AFB1 removal by cell lysates from B. adusta culture, AFB1 analyses were performed after treatment with pronase E, one of proteases. The cell lysates after pronase E treatment showed a slightly decreased AFB1 removal activity (42%) on the 5th day compared to those without treatment (56%) (Figure 5C). These results suggest that heat-stable proteins in cell lysates play a minor role in AFB1 removal.
2.7. AFB1 Removal by Whole Cells, Cell Lysates, Cell Debris, and Exopolysaccharides from B. adusta Cultures
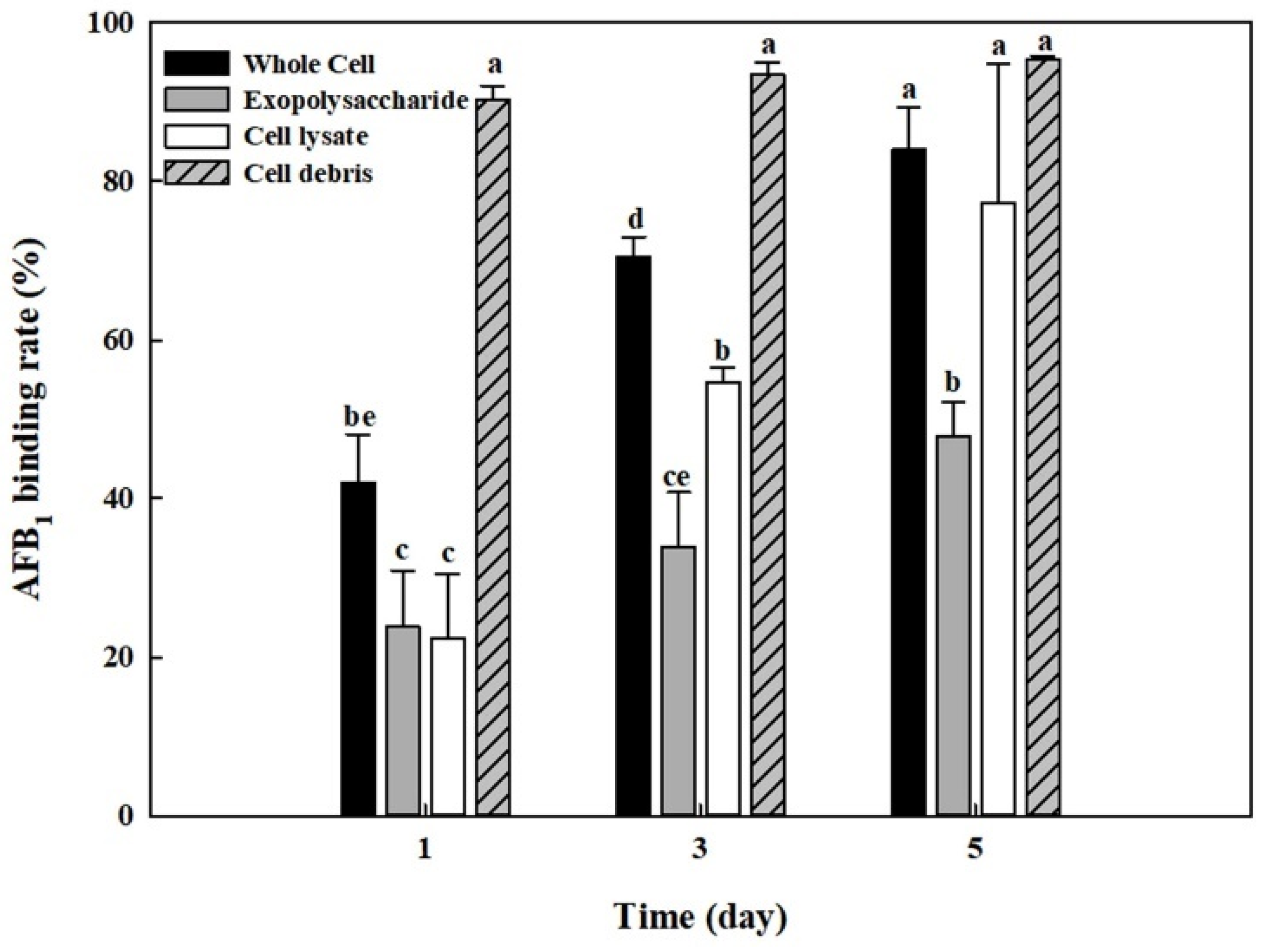
The AFB1 removal results using cell-free supernatants shown in Figure 2 indicated that cell-free supernatants also had the removal activities. In addition, it was reported that exopolysaccharides produced by microorganisms are possibly involved in mycotoxin removal [47]. Thus, we analyzed the amounts of total carbohydrates, protein, and glucosamine in cell-free supernatants, cell lysates, and cell debris. Table 1 shows that the cell-free supernatant from B. adusta culture has a high level of total carbohydrates (p < 0.01). Therefore, exopolysaccharides were extracted from cell-free supernatants of B. adusta culture to test if exopolysaccharides are responsible for AFB1 removal in cell-free supernatants. In addition, it has been documented that not only lactic acid bacteria such as Lactobacillus sp. and Streptococcus sp. but also yeast such as S. cerevisiae can reduce levels of AFs by binding AFB1 onto their cell wall components [28,30,38,48]. Thus, AFB1 quantification assays were conducted on whole cells, cell lysates, exopolysaccharides, and cell debris from B. adusta culture. The cell debris showed the highest AFB1 binding activity (95%) after 5-day incubation, while exopolysaccharides showed a lower AFB1 removal activity (48%) compared to the cell debris and cell lysates (77%) (Figure 6). Moreover, AFB1 removal activities by cell debris had negligible changes throughout the 5-day incubation (91–95%), whereas those using whole cells, cell lysates, or exopolysaccharides showed a gradually increasing trend for 5 days (42–84%, 22–77%, and 24–48%, respectively). These results strongly suggest that AFB1 removal activity by whole cells was mainly due to AFB1 binding onto cell debris during early incubation and that it was due to proteins in cell lysates and partly exopolysaccharides after saturation of AFB1 binding process onto cell wall components.
3. Discussion
This study aimed to investigate AFB1 removal activities of edible mushroom-forming fungi and the mechanisms of AFB1 removal by the fungi. It has been reported that biological control methods by microorganisms have more practical application in elimination of mycotoxins in food and feed than chemical or physical methods because microorganisms can degrade mycotoxins to less toxic or nontoxic products [27]. In particular, some researchers described that laccase or peroxidase from some mushroom-forming fungi including P. ostreatus was able to degrade AFB1 [24,26]. In this study, B. adusta showed decolorization of RBBR dye, indicating that it may produce potential lignocellulolytic enzymes. This result is consistent with previous studies. Alberts and colleagues described that B. adusta SCC0169 strain degraded Poly R-478 dye effectively, which is used to screen for potential polycyclic aromatic hydrocarbon degrading fungi [24]. In addition, it was documented that one B. adusta strain produced versatile peroxidase, a hybrid enzyme between Mn-peroxidase and lignin-peroxidase, and decolorized industrial dyes [39,41,42]. However, B. adusta, in our study, was not able to degrade coumarin (a basic structure of AFs), while the SCC0169 strain showed a relatively low AFB1 degradation activity (28.19%) [24]. These results suggest that our B. adusta strain could degrade other phenolic or aromatic compounds than coumarin derivatives including AFs. In addition, in the present study, cell-free supernatants from B. adusta showed very low AFB1 removal activity (13%) compared to cell lysates from the strain (35%) (Figure 2). Moreover, heat or protease treatment of cell lysates from B. adusta did not exhibit much decrease in AFB1 removal activity (Figure 5B,C). Previous studies reported that heat treatment at 120 °C for 20 min did not affect AFB1 removing ability in Lactobacillus acidophilus and S. cerevisiae [49,50]. Additionally, heat treatment of cell-free extracts from a filamentous fungus Phoma sp. in boiling water for 10 min did not show a significant difference compared to unheated cell-free extracts [18]. These are in agreement with our results. In addition, exopolysaccharides isolated from cell-free supernatants also had much lower AFB1 removal activity (48%) compared to cell lysates (77%), cell debris (95%), or whole cells (84%) in our study (Figure 6). This result is not consistent with Taheur and co-workers’ study, in which they described that an exopolysaccharide (kefiran) from Lactobacillus kefiri strain on Kefir grains may be involved in AFB1 removal by adsorption [50]. The discrepancy may have come from different components of exopolysaccharides between L. kefiri (a lactic acid bacterium) and B. adusta (a mushroom-forming fungus). Taken together, our data suggest that ligninolytic enzymes such as laccases or peroxidases were not involved in AFB1 removal by B. adusta, and that cell lysates from the strain play a major role in AFB1 removal with a minor role of exopolysaccharides in the present study.
Previously, a number of reports have shown that lactic acid bacteria and yeasts can eliminate AFs by adhesion to their cell wall components [27,29,33,36,38,48,51,52]. Peltonen and collaborators showed that two Lactobacillus amylovorus strains and one Lactobacillus rhamnosus strain, which are commonly used in the food industry as a starter culture, removed more than 50% AFB1 by binding onto cell wall during 72 h incubation [33]. Another study reported that five probiotic strains such as L. acidophilus, L. rhamnosus, Lactobacillus reuteri, Lactobacillus johnsonii, and Bifidobacterium bifidum were able to bind approximately 20% of AFM1 [48]. L. rhamnosus strain GG also reduced AFB1 uptake into and toxicity in Caco-2 cells by binding to bacterial cell wall [29]. In addition, some researchers showed that yeast removed AFB1 by the similar mechanism to lactic acid bacteria [36,38]. Kusumaningtyas and colleagues reported that S. cerevisiae reduced 60% of AFs in chicken feed by binding at 5th day when it was co-cultured with toxigenic A. flavus [36]. Another researcher showed that S. cerevisiae strains were able to bind approximately 50% of AFB1 [38]. In the current study, our results indicate that AFB1 removal activity during an early incubation period was due to cell debris, while the activity during the late incubation period was due to cell lysates (Figure 6). Therefore, we concluded that AFB1 binds to cell wall components in B. adusta during the early incubation period and it then binds to cell lysates after saturation of the AFB1 binding process onto the cell wall. Furthermore, in this study, AFB1 removal activity was more than 80% in the whole cell and cell debris after 5-day incubation (84% and 95%, respectively), which showed higher binding efficiencies compared to those in yeast strains (50–60%) as described previously [36,38]. It has been documented that there is a variation in AF binding ability between different strains and that it was due to different number of binding sites for AFs in cell walls components of different strains [33,47,49]. It is known that polysaccharides and proteins in cell walls of yeast and lactic acid bacteria play a major role in mycotoxin binding [38,49]. Available literature also indicated that in S. cerevisiae β-D-glucans and mannoproteins (glucomannan) in its cell wall are involved in adsorption of mycotoxins such as AFB1 and zearalenone, while in lactic acid, bacteria peptidoglycan and teichoic acids in their cell wall are responsible for AFB1 binding activity [38,49,53,54,55,56]. Ruiz-Herrera reported that the cell wall of Basidiomycota is mainly composed of glucans, chitins, and mannoproteins, the percentages of which are significantly different from those of yeasts (1% chitin in Ascomycota to which S. cerevisiae belong, while 35% chitin in Basidiomycota to which Coprinus belong) [57]. It is possible that in addition to glucans and mannoproteins, other polysaccharide chitins in the cell wall of B. adusta contributed to the AFB1 binding. Moreover, Figure 5C showed that pronase E treatment of cell lysates from B. adusta slightly decreased AFB1 removal activity compared to the control. It suggests that proteins in cell lysates play a minor role in AFB1 removal by B. adusta and that pronase may have released other components (involved in AFB1 binding process) associated with proteins by protein degradation, which is in agreement with explanation by other researchers [38]. In summary, our data indicate that after AFB1 binds to mainly cell wall components in B. adusta, it binds to cell lysates (possibly components associated with proteins in cell lysates) along with exopolysaccharides because of saturation of AFB1 binding process onto cell wall.
The edible mushroom-forming fungal strain such as B. adusta may be of interest as a novel microorganism for reduction in the contamination of AFs in the food and feed industries. It could form complexes with AFs and prevent absorption of AFs in the gastrointestinal tract when these mushroom-forming fungi are ingested by human or given to animals as feeds. Thus, our findings in this study will contribute to the development of preventive strategies to eliminate contamination of AFB1 in food and feed.
4. Conclusions
The AFB1 removal test using whole cells, cell lysates, exopolysaccharides, and cell debris from B. adusta culture exhibited that cell debris had the highest AFB1 removal activity (95%) and that the removal activities of cell lysates (77%) and exopolysaccharides (48%) were high in that order after 5-day incubation. Furthermore, AFB1 removal activities by cell debris had negligible changes throughout the 5-day incubation (91–95%), whereas those using whole cells (42–84%), cell lysates (22–77%), or exopolysaccharides (24–48%) showed a gradually increasing tendency for 5 days. Thus, based on these results, we concluded that in B. adusta after AFB1 binds onto cell debris during early incubation, it binds onto cell lysates along with exopolysaccharides when the AFB1 binding process onto cell wall components is saturated.
5. Materials and Methods
5.1. Chemicals and Reagents
AFB1 standard, RBBR, coumarin, sodium periodate, and trifluoroacetic acid (TFA) were obtained from Sigma-Aldrich Co. (St. Louis, MO, USA). Pronase E from Streptomyces griseus was also purchased from Sigma-Aldrich. HPLC grade methanol and acetonitrile were obtained from J.T. Baker (Avantor Performance Materials, Inc., Center Valley, PA, USA). Ethyl acetate was purchased from Daejung Chemicals and Metals Co. (Gyeonggi-do, Korea).
5.2. Fungal Strains and Culture Conditions
Five edible mushroom-forming fungi (B. adusta, A. auricular-judae, L. edodes, H. erinaceus, and P. cocos) in Basidiomycota were collected from mushroom farms in Gyeonggi province in South Korea. Each fungal culture was prepared by incubation at 25 °C for 10–14 days after center-inoculation of a block (1 cm × 1 cm × 0.5 cm) of the fruiting body onto PDA (MB Cell, Seoul, South Korea). The fungal culture was used as a source of a mycelial inoculum for subsequent cultures. All other mushroom-forming fungal cultures were performed at 25 °C for 7–10 days.
For large scale culture, B. adusta was cultured in 150 mL of PDB (MB Cell, Seoul, South Korea) at 25 °C for 7 days with shaking at 100 rpm. Then, fungal cells were transferred into and cultured in 5 L jar fermentor (Fermentech Co, Cheongju, ChungCheongBuk-do, Korea) at 25 °C for 7 days with shaking at 150 rpm.
5.3. Test for Degradation of RBBR and Coumarin by Mushrooms
In order to detect mushroom-forming fungi that can produce ligninolytic enzymes, RBBR was added to agar media (20 g glucose, 5 g peptone, 2 g yeast extract, 1 g KH2PO4, 0.5 g MgSO4 7H2O, 1 g RBBR, 15 g agar, and 1000 mL distilled water) [58]. In addition, for detection of mushroom-forming fungi that can degrade AFB1 (a coumarin derivative), coumarin media including coumarin as the sole carbon source were used: 10 g coumarin, 0.05 g KH2PO4, 1 g NH4NO3, 1 g CaCl2, 0.25 g MgSO4.7H2O, 1 mg FeSO4, 15 g agar, and 1000 mL DW [59].
5.4. Time Course of Fungal Growth, Change in pH, and Removal of AFB1
In order to measure fungal growth on solid culture media, each mushroom-forming fungus was inoculated onto PDA plates using agar plugs (5 mm diameter) cut from the periphery of the actively growing mycelial colony, which had been cultured on the same medium (PDA). It was incubated at 25 °C for 7–10 days. Colony diameter was measured every day for radial growth of mushrooms.
For time course experiments of AFB1 removal by mushroom-forming fungi, each fun gal strain (10 agar plugs; 5 mm diameter) was cultured in 30 mL of PDB containing 50 ng/mL of AFB1 at 25 °C for 14 days with shaking at 100 rpm. The fungal culture media were taken at the 0, 1, 3, 5, 7, 10, and 14th day in triplicate. The pH of culture was measured using a pH meter (Hanna, Smithfield, RI, USA) after filtration with Whatman No.4 filter paper (Whatman Inc., Clifton, NJ, USA).
5.5. AFB1 Extraction from Fungal Culture Media
For extraction of AFB1 from fungal culture media, the fungal culture was filtrated using Whatman No. 4 filter paper. Then, 3 mL of ethyl acetate and 1 mL of filtrate were mixed by a vortex mixer (Fisher Scientific, Springfield, NJ, USA) for 30 s. After the mixture was placed at 25 °C for 30 min, 2 mL of ethyl acetate upper layer was transferred to a new glass test tube. Two microliters of ethyl acetate was added to the lower layer, and they were mixed by vortexing for 30 s. Again, after the mixture was placed at 25 °C, its upper layer was combined with the first extract. The 4 mL of ethyl acetate extracts were evaporated to dryness under a gentle stream of nitrogen at 60 °C.
5.6. AFB1 Assays Using Whole Cells, Cell Lysates after Cell Disruption, Cell-Free Supernatants, and Cell Debris
After large-scale fermentation, B. adusta cells were harvested by centrifugation (8000 rpm, 20 min, 4 °C) and freeze-dried using a freeze-dryer (IlshinBioBase, Dongducheon, Korea) for 4 days. Freeze-dried B. adusta cells (75 mg) were transferred to a vial, and 4.5 mL of citrate–phosphate buffer (pH 7) was added to it. Samples were spiked with 0.5 mL of AFB1 standard solution to give 1 μg/mL of AFB1 as the final concentration and incubated for 5 days at 40 °C with shaking at 100 rpm. Fifty microliters of samples were taken at the 1, 3, and 5th day.
A portion of freeze-dried mycelia (1.2 g) were ground in liquid nitrogen in a mortar with a pestle. The powdered mycelia were resuspended in 1 mL of ice-cold citrate–phosphate buffer (pH 7). After cell debris were pelleted by centrifugation at 13,000 rpm for 10 min at 4 °C, supernatants were filtered through a syringe filter (47 mm × 0.45 µm, GHP; Pall Corporation, Port Washington, NY, USA) and used as cell lysates. The precipitated materials were used as cell debris. AFB1 assay using cell debris was performed by the same procedure with that using whole cells as described above. For measurement of AFB1 removal activity at different temperatures, cell lysates (900 μL) were transferred to a vial and spiked with 100 μL of AFB1 to give 1 μg/mL of AFB1 as the final concentration. Samples were incubated for 5 days at 25, 30, 35, and 40 °C with shaking at 100 rpm, and 50 μL of samples were taken at the 1, 3, and 5th day.
The cell-free supernatant from B. adusta fermentation broth was used for either AFB1 quantification assay or extraction of exopolysaccharides. For AFB1 assay using supernatants, they were filtered through a syringe filter (0.45 μm). The supernatant (900 μL) was then transferred to a vial and spiked with 100 μL of AFB1 to give 1 μg/mL of AFB1 as the final concentration. Samples were incubated for 1 day at 40 °C with shaking at 100 rpm. For extraction of exopolysaccharides, ice-cold ethanol was gradually added to the supernatant up to 80% (v/v) saturation, and it was stirred at 4 °C overnight. The exopolysaccharide was obtained by centrifugation (8000 rpm, 20 min, 4 °C) and freeze-dried using a freeze-dryer for 4 days. AFB1 assay using exopolysaccharides was performed by the same procedure with that using whole cells as described above. All experiments were carried out in triplicate.
5.7. Effects of 3 mM NaIO4 and 0.2 mM NADHP on AFB1 Removal by Cell Lysates
Two-hundred microliters of sodium periodate (10 mg/mL), which was prepared in acetate buffer (pH 4.5), and 51 μL of NADPH (10 mg/mL) were added to 3 mL of cell lysate, and they were incubated at 25 °C for 1 day. Then, samples were transferred into 100 mL of citrate–phosphate buffer (pH 7), and they were dialyzed for 5 h at room temperature. Samples were spiked with AFB1 standard solution to give 1 μg/mL of AFB1 as the final concentration and incubated for 2 days at 40 °C with shaking at 100 rpm. All experiments were carried out in triplicate.
5.8. AFB1 Assays Using Heat- or Pronase-Treated Whole Cells and Cell Lysates
Freeze-dried cells (75 mg) were transferred to a vial, and 4.5 mL of citrate–phosphate buffer (pH 7) was added to it. After samples were autoclaved at 121 °C for 15 min and cooled down, they were spiked with 0.5 mL of AFB1 (1 μg/mL). Then, samples were incubated for 5 days at 40 °C with shaking at 100 rpm, and 50 μL of samples were taken at the 1, 3, and 5th day.
Cell lysates (900 μL) were transferred to a vial and heated in a 95 °C water bath (Vision Scientific Co., Daejeon, Korea) for 10 min. Samples were spiked with 100 μL of AFB1 (final concentration: 1 μg/mL) and incubated for 5 days at 40 °C with shaking at 100 rpm. Fifty microliters of samples were taken at the 1, 3, and 5th day.
One milliliter of pronase E solution (0.5 mg/mL), which was prepared in phosphate buffer (pH 7.6), was added to 3 mL of cell lysate, and they were incubated at 25 °C for 1 day. Then, samples were transferred into 100 mL of citrate–phosphate buffer (pH 7), and they were dialyzed for 5 h at room temperature. Samples were spiked with AFB1 standard solution to give 1 μg/mL of AFB1 as the final concentration and incubated for 5 days at 40 °C with shaking at 100 rpm. Fifty microliters of samples were taken at the 1, 3, and 5th day. All experiments were carried out in triplicate.
5.9. AFB1 Analysis by HPLC-FLD
After liquid–liquid extraction of AFB1 in culture broth as described above, the dried culture extracts were dissolved with 1 mL of TFA-10% acetonitrile (10:90, v/v) and mixed by a vortex mixer for 30 s. The mixture was placed in darkness for 3 h and, then, filtered through a syringe filter (13 mm × 0.2 μm, GH polypro membrane [GHP], Pall corporation, Port Washington, NY, USA). The AFB1 standard solution was also derivatized with TFA using the same procedure as described above.
Dionex Ultimate 3000 HPLC (Thermo Fisher Scientific, Sunnyvale, CA, USA) was programmed to inject 10 μL of samples and AFB1 standard solutions and run for 20 min through a Ultrasphere® C18 column (4.6 mm i.d. × 250 mm, 5 μm; Beckman Coulter, Miami, FL, USA). The mobile phase was acetonitrile–methanol–water (15:15:70, v/v/v) pumped at a constant flow rate of 1 mL/min. The determination of AFB1 was carried out using a fluorescence detector with 360 nm and 440 nm for excitation and emission, respectively [60].
For AFB1 assays using whole cells, cell lysates, and cell-free supernatants, 50 μL of samples were dissolved with 950 μL of TFA-10% acetonitrile (10:90, v/v) and mixed by a vortex mixer for 30 s.
The sensitivity of the analytical method using HPLC-FLD was determined by a limit of detection (LOD) and limit of quantification (LOQ). They were calculated as signal-to-noise (S/N) ratios of 3 and 10, respectively, which were measured by using Chromeleon 6.8 Chromatography Data System (Thermo Fisher Scientific). The LOD and LOQ for AFB1 were 0.03 and 0.1 μg/L, respectively.
The linearity of a series of AFB1 concentrations in the analytical method was assessed by a standard curve using seven levels of AFB1 standard solutions (0.5, 5, 20, 50, 100, 200, and 500 ng/mL). The linearity was determined by linear regression analysis and expressed as a coefficient of determination (r2). The curve showed an r2 value of 0.9986.
The repeatability (within-day precision) was determined by three consecutive inject-ions of AFB1 solutions within a day. Relative standard deviation (RSD) was in the range of 0.0–3.7%.
5.10. Determination of Total Carbohydrates, Protein, and Glucosamine in Cell-Free Supernatants, Cell Lysates, and Cell Debris
The amounts of total carbohydrates were determined by the phenol-sulfuric acid method with minor modifications according to Dubois et al. [61]. Briefly, 5% phenol (200 μL) was added to 200 μL of each sample in a test tube, and it was mixed by vortexing for 30 s. Then, 1 mL of sulfuric acid was added to it, and it was mixed by vortexing for 30 s. After it was cooled down for 20 min at room temperature, the absorbance of each sample was measured at 490 nm using a spectrophotometer (Genesys, 10S UV-VIS, Thermo Fisher Scientific, Waltham, MA, USA). Glucose solutions (0–200 μg/mL) were used to construct a standard curve.
The amounts of protein were determined using a bicinchoninic acid (BCA) assay (Pierce BCA protein assay kit; Thermo Fischer Scientific, Waltham, MA, USA) according to the manufacturer’s instructions. The absorbance of each sample was measured at 562 nm using a spectrophotometer (Genesys, 10S UV-VIS). Bovine serum albumin (BSA) (0–2000 μg/mL) was used to create a standard curve.
The amounts of glucosamine were determined using the method by Rondle et al. with minor modifications [62]. Briefly, each sample was hydrolyzed with 6 N HCl at 105 °C for 1 h under N2 atmosphere. The hydrolyzed sample (0.5 mL) was added to 0.5 mL of distilled water in a Pyrex tube (18 mm × 150 mm; Corning, NY, USA) with a ground-glass stopper, and 1 mL of acetylacetone reagent (4% acetylacetone in 1.5 N NaCO3) was added to it. It was incubated for 30 min in a boiling water bath. After cooled down in water, 5 mL of ethanol was added, and it was mixed with vortexing for 30 s. Then, 1 mL of Ehrlich reagent (2.67% [w/v] of p-dimethylaminnobenzaldehyde in a solution of 95% ethanol and conc. HCl [1:1]) was added to it. It was mixed with vortexing for 30 s and incubated at room temperature for 25 min. The absorbance of each sample was measured at 530 nm using a spectrophotometer (Genesys, 10S UV-VIS).
A serial dilution of 0.2% glucosamine-HCl standard solution (0–100 μg/mL) was used to construct a standard curve.
5.11. Statistical Analysis
Data were statistically analyzed by t-test or a one-way analysis of variance (ANOVA) and expressed as the mean ± standard deviation using SigmaStat software (Jandel Corporation, San Rafael, CA, USA). Tukey’s test was performed for post hoc comparisons. A p value < 0.05 was considered statistically different.
Author Contributions
Conceptualized and designed experiments, S.-Y.H., S.-H.C. and A.-S.O.; performed experiments, M.-J.C.; analyzed data, M.-J.C., S.-Y.H., S.-H.C. and A.-S.O.; wrote the manuscript, M.-J.C. and S.-Y.H. All authors have read and agreed to the published version of the manuscript.
Funding
No funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cotty, P.J.; Bayman, P.; Egel, D.S.; Elias, D.S. Agriculture, aflatoxins, and Asperigiilus. In The Genus Aspergillus; Powell, K.A., Fenwick, A., Peberdy, J.F., Eds.; Plenum Press: New York, NY, USA, 1994; pp. 1–27. [Google Scholar]
- Gourama, H.; Bullerman, L.B. Aspergillus flavus and Aspergillus parasiticus: Aflatoxigenic fungi of concern in foods and feeds: A review. J. Food Prot. 1995, 58, 1395–1404. [Google Scholar] [CrossRef] [PubMed]
- Eaton, D.L.; Gallagher, E.P. Mechanism of aflatoxin carcinogenesis. Annu. Rev. Pharm. 1994, 34, 135–172. [Google Scholar] [CrossRef]
- IARC. Summaries and Evaluations: Aflatoxins; IARC: Lyon, France, 2003; p. 171. [Google Scholar]
- Herzallah, S.; Alshawabkeh, K.; Al Fataftah, A. Aflatoxin decontamination of artificially contaminated feeds by sunlight, gamma-radiation, and microwave heating. J. Appl. Poult. Res. 2008, 17, 515–521. [Google Scholar] [CrossRef]
- Diaz, D.E.; Hagler, W.M.; Blackwelder, J.T.; Eve, J.A.; Hopkins, B.A.; Anderson, K.L.; Jones, F.T.; Whitlow, L.W. Aflatoxin Binders II: Reduction of aflatoxin M1 in milk by sequestering agents of cows consuming aflatoxin in feed. Mycopathologia 2004, 157, 233–241. [Google Scholar] [CrossRef]
- Grant, P.G.; Phillips, T.D. Isothermal adsorption of aflatoxin B1 on HSCAS clay. J. Agr. Food Chem. 1998, 46, 599–605. [Google Scholar] [CrossRef]
- Mishra, H.N.; Das, C. A review on biological control and metabolism of aflatoxin. Crit. Rev. Food Sci. 2003, 43, 245–264. [Google Scholar] [CrossRef]
- Dwarakanath, C.T.; Rayner, E.T.; Mann, G.E.; Dollear, F.G. Reduction of aflatoxin levels in cottonseed and peanut meals by ozonization. J. Am. Oil Chem. Soc. 1968, 45, 93–95. [Google Scholar] [CrossRef] [PubMed]
- Doyle, M.P.; Marth, E.H. Bisulfite degrades aflatoxin—Effect of temperature and concentration of bisulfite. J. Food Prot. 1978, 41, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Park, D.L.; Lee, L.; Koltun, S.A. Distribution of ammonia-related aflatoxin reaction-products in cottonseed meal. J. Am. Oil Chem. Soc. 1984, 61, 1071–1074. [Google Scholar] [CrossRef]
- Kabak, B.; Dobson, A.D.W.; Var, I. Strategies to prevent mycotoxin contamination of food and animal feed: A review. Crit. Rev. Food Sci. 2006, 46, 593–619. [Google Scholar] [CrossRef]
- Huwig, A.; Freimund, S.; Kappeli, O.; Dutler, H. Mycotoxin detoxication of animal feed by different adsorbents. Toxicol. Lett. 2001, 122, 179–188. [Google Scholar] [CrossRef]
- Samuel, S.M.; Aiko, V.; Panda, P.; Mehta, A. Aflatoxin B1 occurrence, biosynthesis and its degradation. J. Pure Appl. Microbio. 2013, 7, 965–971. [Google Scholar]
- Kolosova, A.; Stroka, J. Substances for reduction of the contamination of feed by mycotoxins: A review. World Mycotoxin J. 2011, 4, 225–256. [Google Scholar] [CrossRef]
- Hormisch, D.; Brost, I.; Kohring, G.W.; Giffhorn, E.; Kroppenstedt, R.M.; Stackebrandt, E.; Farber, P.; Holzapfel, W.H. Mycobacterium fluoranthenivorans sp nov., a fluoranthene and aflatoxin B1 degrading bacterium from contaminated soil of a former coal gas plant. Syst. Appl. Microbiol. 2004, 27, 653–660. [Google Scholar] [CrossRef]
- Teniola, O.D.; Addo, P.A.; Brost, I.M.; Farber, P.; Jany, K.D.; Alberts, J.F.; van Zyl, W.H.; Steyn, P.S.; Holzapfel, W.H. Degradation of aflatoxin B1 by cell-free extracts of Rhodococcus erythropolis and Mycobacterium fluoranthenivorans sp nov DSM44556(T). Int. J. Food Microbiol. 2005, 105, 111–117. [Google Scholar] [CrossRef]
- Shantha, T. Fungal degradation of aflatoxin B1. Nat. Toxins 1999, 7, 175–178. [Google Scholar] [CrossRef]
- Doyle, M.P.; Applebaum, R.S.; Brackett, R.E.; Marth, E.H. Physical, chemical and biological degradation of mycotoxins in foods and agricultural commodities. J. Food Prot. 1982, 45, 964–971. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Q.; Ogata, M.; Hirai, H.; Kawagishi, H. Detoxification of aflatoxin B1 by manganese peroxidase from the white-rot fungus Phanerochaete sordida YK-624. FEMS Microbiol. Lett. 2011, 314, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Arora, D.S.; Sharma, R.K. Ligninolytic fungal laccases and their biotechnological applications. Appl. Biochem. Biotech. 2010, 160, 1760–1788. [Google Scholar] [CrossRef]
- Vyas, B.R.M.; Molitoris, H.P. Involvement of an extracellular H2O2-dependent ligninolytic activity of the white-rot fungus Pleurotus ostreatus in the decolorization of remazol-brilliant-blue-R. Appl. Environ. Microbiol. 1995, 61, 3919–3927. [Google Scholar] [CrossRef] [Green Version]
- Pickard, M.A.; Roman, R.; Tinoco, R.; Vazquez-Duhalt, R. Polycyclic aromatic hydrocarbon metabolism by white rot fungi and oxidation by Coriolopsis gallica UAMH 8260 laccase. Appl. Environ. Microbiol. 1999, 65, 3805–3809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberts, J.F.; Gelderblom, W.C.A.; Botha, A.; van Zyl, W.H. Degradation of aflatoxin B1 by fungal laccase enzymes. Int. J. Food Microbiol. 2009, 135, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Motomura, M.; Toyomasu, T.; Mizuno, K.; Shinozawa, T. Purification and characterization of an aflatoxin degradation enzyme from Pleurotus ostreatus. Microbiol. Res. 2003, 158, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Yehia, R.S. Aflatoxin detoxification by manganese peroxidase purified from Pleurotus ostreatus. Braz. J. Microbiol. 2014, 45, 127–133. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Jezkova, A.; Yuan, Z.; Pavlikova, L.; Dohnal, V.; Kuca, K. Biological degradation of aflatoxins. Drug Metab. Rev. 2009, 41, 1–7. [Google Scholar] [CrossRef]
- Adebo, O.A.; Njobeh, P.B.; Gbashi, S.; Nwinyi, O.C.; Mavumengwana, V. Review on microbial degradation of aflatoxins. Crit. Rev. Food Sci. 2017, 57, 3208–3217. [Google Scholar] [CrossRef]
- Gratz, S.; Wu, Q.K.; El-Nezami, H.; Juvonen, R.O.; Mykkanen, H.; Turner, P.C. Lactobacillus rhamnosus strain GG reduces aflatoxin B1 transport, metabolism, and toxicity in caco-2 cells. Appl. Environ. Microbiol. 2007, 73, 3958–3964. [Google Scholar] [CrossRef] [Green Version]
- El Khoury, A.; Atoui, A.; Yaghi, J. Analysis of aflatoxin M1 in milk and yogurt and AFM1 reduction by lactic acid bacteria used in Lebanese industry. Food Control 2011, 22, 1695–1699. [Google Scholar] [CrossRef]
- El-Nezami, H.; Mykkanen, H.; Kankaanpaa, P.; Salminen, S.; Ahokas, J. Ability of Lactobacillus and Propionibacterium strains to remove aflatoxin B1 from the chicken duodenum. J. Food Prot. 2000, 63, 549–552. [Google Scholar] [CrossRef]
- Oluwafemi, F.; Kumar, M.; Bandyopadhyay, R.; Ogunbanwo, T.; Ayanwande, K.B. Bio-detoxification of aflatoxin B1 in artificially contaminated maize grains using lactic acid bacteria. Toxin Rev. 2010, 29, 115–122. [Google Scholar] [CrossRef]
- Peltonen, K.; El-Nezami, H.; Haskard, C.; Ahokas, J.; Salminen, S. Aflatoxin B1 binding by dairy strains of lactic acid bacteria and bifidobacteria. J. Dairy Sci. 2001, 84, 2152–2156. [Google Scholar] [CrossRef]
- El-Shiekh, H.H.; Mahdy, H.M.; El-Aaser, M.M. Bioremediation of aflatoxins by some reference fungal strains. Pol. J. Microbiol. 2007, 56, 215–223. [Google Scholar] [PubMed]
- Goncalves, B.L.; Rosim, R.E.; de Oliveira, C.A.F.; Corassin, C.H. The in vitro ability of different Saccharomyces cerevisiae—Based products to bind aflatoxin B1. Food Control 2015, 47, 298–300. [Google Scholar] [CrossRef]
- Kusumaningtyas, E.; Widiastuti, R.; Maryam, R. Reduction of aflatoxin B1 in chicken feed by using Saccharomyces cerevisiae, Rhizopus oligosporus and their combination. Mycopathologia 2006, 162, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Shetty, P.H.; Hald, B.; Jespersen, L. Surface binding of aflatoxin B1 by Saccharomyces cerevisiae strains with potential decontaminating abilities in indigenous fermented foods. Int. J. Food Microbiol. 2007, 113, 41–46. [Google Scholar] [CrossRef]
- Shetty, P.H.; Jespersen, L. Saccharomyces cerevisiae and lactic acid bacteria as potential mycotoxin decontaminating agents. Trends Food Sci. Technol. 2006, 17, 48–55. [Google Scholar] [CrossRef]
- Asgher, M.; Bhatti, H.N.; Ashraf, M.; Legge, R.L. Recent developments in biodegradation of industrial pollutants by white rot fungi and their enzyme system. Biodegradation 2008, 19, 771–783. [Google Scholar] [CrossRef]
- Mester, T.; Field, J.A. Characterization of a novel manganese peroxidase-lignin peroxidase hybrid isozyme produced by Bjerkandera species strain BOS55 in the absence of manganese. J. Biol. Chem. 1998, 273, 15412–15417. [Google Scholar] [CrossRef] [Green Version]
- Pogni, R.; Baratto, M.C.; Giansanti, S.; Teutloff, C.; Verdin, J.; Valderrama, B.; Lendzian, F.; Lubitz, W.; Vazquez-Duhalt, R.; Basosi, R. Tryptophan-based radical in the catalytic mechanism of versatile peroxidase from Bjerkandera adusta. Biochemistry 2005, 44, 4267–4274. [Google Scholar] [CrossRef]
- Tinoco, R.; Verdin, J.; Vazquez-Duhalt, R. Role of oxidizing mediators and tryptophan 172 in the decoloration of industrial dyes by the versatile peroxidase from Bjerkandera adusta. J. Mol. Catal. B-Enzym. 2007, 46, 1–7. [Google Scholar] [CrossRef]
- Boer, C.G.; Obici, L.; de Souza, C.G.M.; Peralta, R.M. Decolorization of synthetic dyes by solid state cultures of Lentinula (Lentinus) edodes producing manganese peroxidase as the main ligninolytic enzyme. Bioresour. Technol. 2004, 94, 107–112. [Google Scholar] [CrossRef]
- Bergot, B.J.; Stanley, W.L.; Masri, M.S. Reaction of coumarin with aqua ammonia—implications in detoxification of aflatoxin. J. Agr. Food Chem. 1977, 25, 965–966. [Google Scholar] [CrossRef]
- Grove, M.D.; Plattner, R.D.; Weisleder, D. Ammoniation products of an aflatoxin model coumarin. J. Agr. Food Chem. 1981, 29, 1161–1164. [Google Scholar] [CrossRef]
- Hamid, A.B.; Smith, J.E. Degradation of aflatoxin by Aspergillus flavus. J. Gen. Microbiol. 1987, 133, 2023–2029. [Google Scholar] [CrossRef] [Green Version]
- Taheur, F.B.; Fedhila, K.; Chaieb, K.; Kouidhi, B.; Bakhrouf, A.; Abrunhosa, L. Adsorption of aflatoxin B1, zearalenone and ochratoxin A by microorganisms isolated from Kefir grains. Int. J. Food Microbiol. 2017, 251, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrano-Nino, J.C.; Cavazos-Garduno, A.; Hernandez-Mendoza, A.; Applegate, B.; Ferruzzi, M.G.; San Martin-Gonzalez, M.F.; Garcia, H.S. Assessment of probiotic strains ability to reduce the bioaccessibility of aflatoxin M1 in artificially contaminated milk using an in vitro digestive model. Food Control 2013, 31, 202–207. [Google Scholar] [CrossRef]
- Bueno, D.J.; Casale, C.H.; Pizzolitto, R.P.; Salvano, M.A.; Oliver, G. Physical adsorption of aflatoxin B1 by lactic acid bacteria and Saccharomyces cerevisiae: A theoretical model. J. Food Prot. 2007, 70, 2148–2154. [Google Scholar] [CrossRef]
- Pizzolitto, R.P.; Bueno, D.J.; Armando, M.R.; Cavaglieri, L.; Dalcero, A.M.; Salvano, M.A. Binding of aflatoxin B1 to lactic acid bacteria and Saccharomyces cerevisiae in vitro: A useful model to determine the most efficient microorganism. In Aflatoxins-Biochemistry and Molecular Biology, Guevara-Gonzalez; Guevara-Gonzalez, R.G., Ed.; InTech: Rijeka, Croatia, 2011; pp. 323–346. [Google Scholar] [CrossRef] [Green Version]
- Haskard, C.A.; El-Nezami, H.S.; Kankaanpaa, P.E.; Salminen, S.; Ahokas, J.T. Surface binding of aflatoxin B1 by lactic acid bacteria. Appl. Environ. Microbiol. 2001, 67, 3086–3091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahaie, S.; Emam-Djomeh, Z.; Razavi, S.H.; Mazaheri, M. Evaluation of aflatoxin decontaminating by two strains of Saccharomyces cerevisiae and Lactobacillus rhamnosus strain GG in pistachio nuts. Int. J. Food Sci. Technol. 2012, 47, 1647–1653. [Google Scholar] [CrossRef]
- Yiannikouris, A.; Francois, J.; Poughon, L.; Dussap, C.G.; Bertin, G.; Jeminet, G.; Jouany, J.P. Adsorption of zearalenone by beta-D-glucans in the Saccharomyces cerevisiae cell wall. J. Food Prot. 2004, 67, 1195–1200. [Google Scholar] [CrossRef] [PubMed]
- Yiannikouris, A.; Andre, G.; Poughon, L.; Francois, J.; Dussap, C.G.; Jeminet, G.; Bertin, G.; Jouany, J.P. Chemical and conformational study of the interactions involved in mycotoxin complexation with beta-D-glucans. Biomacromolecules 2006, 7, 1147–1155. [Google Scholar] [CrossRef] [PubMed]
- Karaman, M.; Basmacioglu, H.; Ortatatli, M.; Oguz, H. Evaluation of the detoxifying effect of yeast glucomannan on aflatoxicosis in broilers as assessed by gross examination and histopathology. Brit. Poult. Sci. 2005, 46, 394–400. [Google Scholar] [CrossRef]
- Lahtinen, S.J.; Haskard, C.A.; Ouwehand, A.C.; Salminen, S.J.; Ahokas, J.T. Binding of aflatoxin B1 to cell wall components of Lactobacillus rhamnosus strain GG. Food Addit. Contam. A 2004, 21, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Herrera, J. Fungal Cell Wall: Structure, Synthesis and Assembly; CRC Press: Boca Raton, FL, USA, 1992. [Google Scholar]
- Ha, H.-C. Screening and production of lignocellulolytic enzymes secreted by the edible basidiomycete Pleurotus ostreatus. J. Mushrooms 2012, 10, 74–82. [Google Scholar] [CrossRef]
- Guan, S.; Ji, C.; Zhou, T.; Li, J.X.; Ma, Q.G.; Niu, T.G. Aflatoxin B1 degradation by Stenotrophomonas maltophilia and other microbes selected using coumarin medium. Int. J. Mol. Sci. 2008, 9, 1489–1503. [Google Scholar] [CrossRef] [Green Version]
- Abbas, H.K.; Zablotowicz, R.M.; Bruns, H.A.; Abel, C.A. Biocontrol of aflatoxin in corn by inoculation with non-aflatoxigenic Aspergillus flavus isolates. Biocontrol Sci. Technol. 2006, 16, 437–449. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Rondle, C.J.; Morgan, W.T. The determination of glucosamine and galactosamine. Biochem. J. 1955, 61, 586–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Time course of pH and AFB1 removal activity during 5 edible mushroom-forming fungal cultures. Each fungal strain was grown in PDB at 25 °C for 14 days with shaking at 100 rpm. (A) The pH and (B) the levels of AFB1 were measured in triplicate. The values are expressed as the mean ± standard deviation. Different letters at the same culture time point indicate statistically significant differences (p < 0.05).
Figure 1.
Time course of pH and AFB1 removal activity during 5 edible mushroom-forming fungal cultures. Each fungal strain was grown in PDB at 25 °C for 14 days with shaking at 100 rpm. (A) The pH and (B) the levels of AFB1 were measured in triplicate. The values are expressed as the mean ± standard deviation. Different letters at the same culture time point indicate statistically significant differences (p < 0.05).

Figure 2.
AFB1 removal activity by cell-free supernatants and cell lysates from 3 mushroom-forming fungal cultures (B. adusta, A. auricular-judae, and L. edodes). Cell-free supernatants or cell lysates from 3 mushroom-forming fungal cultures, which were spiked with AFB1 (final concentration: 1 μg/mL), were incubated for 1 day at 40 °C with shaking at 100 rpm. The levels of AFB1 were measured in triplicate. The data are expressed as the mean ± standard deviation. Different letters indicate statistically significant differences (p < 0.05).
Figure 2.
AFB1 removal activity by cell-free supernatants and cell lysates from 3 mushroom-forming fungal cultures (B. adusta, A. auricular-judae, and L. edodes). Cell-free supernatants or cell lysates from 3 mushroom-forming fungal cultures, which were spiked with AFB1 (final concentration: 1 μg/mL), were incubated for 1 day at 40 °C with shaking at 100 rpm. The levels of AFB1 were measured in triplicate. The data are expressed as the mean ± standard deviation. Different letters indicate statistically significant differences (p < 0.05).

Figure 3.
Effects of different reaction temperatures on AFB1 removal by cell lysates from the 3 mushroom-forming fungal cultures (B. adusta, A. auricular-judae, and L. edodes). Cell lysates from 3 mushroom-forming fungal cultures, which were spiked with AFB1 (final concentration: 1 μg/mL), were incubated for 5 days at 25, 30, 35, and 40 °C with shaking at 100 rpm. The levels of AFB1 were measured in triplicate. The data are expressed as the mean ± standard deviation. Different letters at the same temperature indicate statistically significant differences (p < 0.05).
Figure 3.
Effects of different reaction temperatures on AFB1 removal by cell lysates from the 3 mushroom-forming fungal cultures (B. adusta, A. auricular-judae, and L. edodes). Cell lysates from 3 mushroom-forming fungal cultures, which were spiked with AFB1 (final concentration: 1 μg/mL), were incubated for 5 days at 25, 30, 35, and 40 °C with shaking at 100 rpm. The levels of AFB1 were measured in triplicate. The data are expressed as the mean ± standard deviation. Different letters at the same temperature indicate statistically significant differences (p < 0.05).

Figure 4.
Effects of NaIO4 and NADPH on AFB1 removal by cell lysates from the 3 mushroom-forming fungal cultures (B. adusta, A. auricular-judae, and L. edodes). NaIO4- and NADPH-treated cell lysates from 3 mushroom-forming fungal cultures (final concentration: 3 mM and 0.2 mM, respectively) were incubated for 2 days at 40 °C with shaking at 100 rpm after spiked with AFB1 (final concentration: 1 μg/mL). The levels of AFB1 were measured in triplicate. The data are expressed as the mean ± standard deviation. Different letters indicate statistically significant differences (p < 0.05).
Figure 4.
Effects of NaIO4 and NADPH on AFB1 removal by cell lysates from the 3 mushroom-forming fungal cultures (B. adusta, A. auricular-judae, and L. edodes). NaIO4- and NADPH-treated cell lysates from 3 mushroom-forming fungal cultures (final concentration: 3 mM and 0.2 mM, respectively) were incubated for 2 days at 40 °C with shaking at 100 rpm after spiked with AFB1 (final concentration: 1 μg/mL). The levels of AFB1 were measured in triplicate. The data are expressed as the mean ± standard deviation. Different letters indicate statistically significant differences (p < 0.05).

Figure 5.
Effects of heat or proteinase treatment on AFB1 removal by whole cells and cell lysates from B. adusta cultures. (A) Heat-treated whole cells, (B) heat-treated cell lysates, and (C) pronase E-treated cell lysates from B. adusta cultures, which were spiked with AFB1 (final concentration: 1 μg/mL), were incubated for 5 days at 40 °C with shaking at 100 rpm. The levels of AFB1 were measured in triplicate. The data are expressed as the mean ± standard deviation. Different letters indicate statistically significant differences (p < 0.05).
Figure 5.
Effects of heat or proteinase treatment on AFB1 removal by whole cells and cell lysates from B. adusta cultures. (A) Heat-treated whole cells, (B) heat-treated cell lysates, and (C) pronase E-treated cell lysates from B. adusta cultures, which were spiked with AFB1 (final concentration: 1 μg/mL), were incubated for 5 days at 40 °C with shaking at 100 rpm. The levels of AFB1 were measured in triplicate. The data are expressed as the mean ± standard deviation. Different letters indicate statistically significant differences (p < 0.05).


Figure 6.
AFB1 binding activity by whole cells, cell lysates, cell debris, and exopolysaccharides from B. adusta cultures. Whole cells, cell lysates, cell debris, and exopolysaccharides from B. adusta cultures, which were spiked with AFB1 (final concentration: 1 μg/mL), were incubated for 5 days at 40 °C with shaking at 100 rpm. The levels of AFB1 were measured in triplicate. The data are expressed as the mean ± standard deviation. Different letters indicate statistically significant differences (p < 0.05).
Figure 6.
AFB1 binding activity by whole cells, cell lysates, cell debris, and exopolysaccharides from B. adusta cultures. Whole cells, cell lysates, cell debris, and exopolysaccharides from B. adusta cultures, which were spiked with AFB1 (final concentration: 1 μg/mL), were incubated for 5 days at 40 °C with shaking at 100 rpm. The levels of AFB1 were measured in triplicate. The data are expressed as the mean ± standard deviation. Different letters indicate statistically significant differences (p < 0.05).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The amounts of total carbohydrates, protein, and glucosamine in cell-free supernatants, cell lysates, and cell debris from B. adusta cultures.
Table 1.
The amounts of total carbohydrates, protein, and glucosamine in cell-free supernatants, cell lysates, and cell debris from B. adusta cultures.
| Cell Fraction | Total Carbohydrate (mg/mL) | Protein (mg/mL) | Glucosamine (mg/mL) |
|---|---|---|---|
| Cell-free supernatant | 42.75 ± 2.64 a | 10.52 ± 1.16 a | N.D. 1 |
| Cell lysate | 11.15 ± 3.63 b | 11.78 ± 0.03 a | N.D. |
| Cell debris | 2.41 ± 0.03 c | 5.12 ± 0.20 b | 0.30 ± 0.01 |
1 N.D. indicates not detected. Total carbohydrates, protein, and glucosamine were measured in triplicate. The data are expressed as the mean ± standard deviation. Different letters in the same column indicate statistically significant differences (p < 0.05).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Choo, M.-J.; Hong, S.-Y.; Chung, S.-H.; Om, A.-S. Removal of Aflatoxin B1 by Edible Mushroom-Forming Fungi and Its Mechanism. Toxins 2021, 13, 668. https://doi.org/10.3390/toxins13090668
AMA Style
Choo M-J, Hong S-Y, Chung S-H, Om A-S. Removal of Aflatoxin B1 by Edible Mushroom-Forming Fungi and Its Mechanism. Toxins. 2021; 13(9):668. https://doi.org/10.3390/toxins13090668
Chicago/Turabian StyleChoo, Min-Jung, Sung-Yong Hong, Soo-Hyun Chung, and Ae-Son Om. 2021. "Removal of Aflatoxin B1 by Edible Mushroom-Forming Fungi and Its Mechanism" Toxins 13, no. 9: 668. https://doi.org/10.3390/toxins13090668
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.