Accumulation and Elimination of Tetrodotoxin in the Pufferfish Takifugu obscurus by Dietary Administration of the Wild Toxic Gastropod Nassarius semiplicata

Abstract
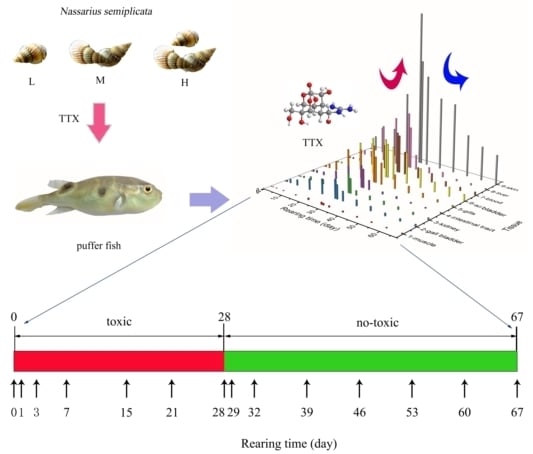
:
1. Introduction
2. Results
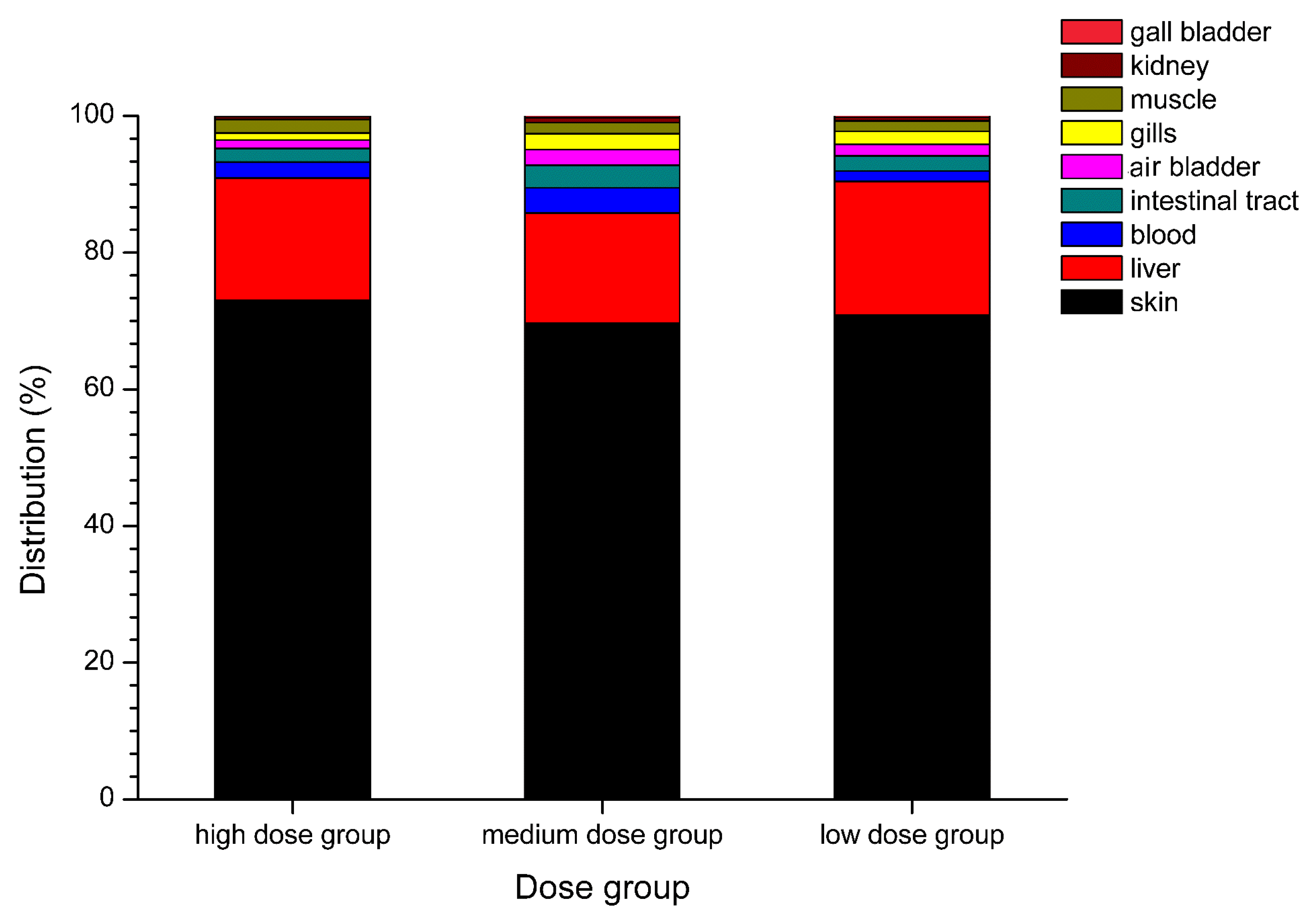
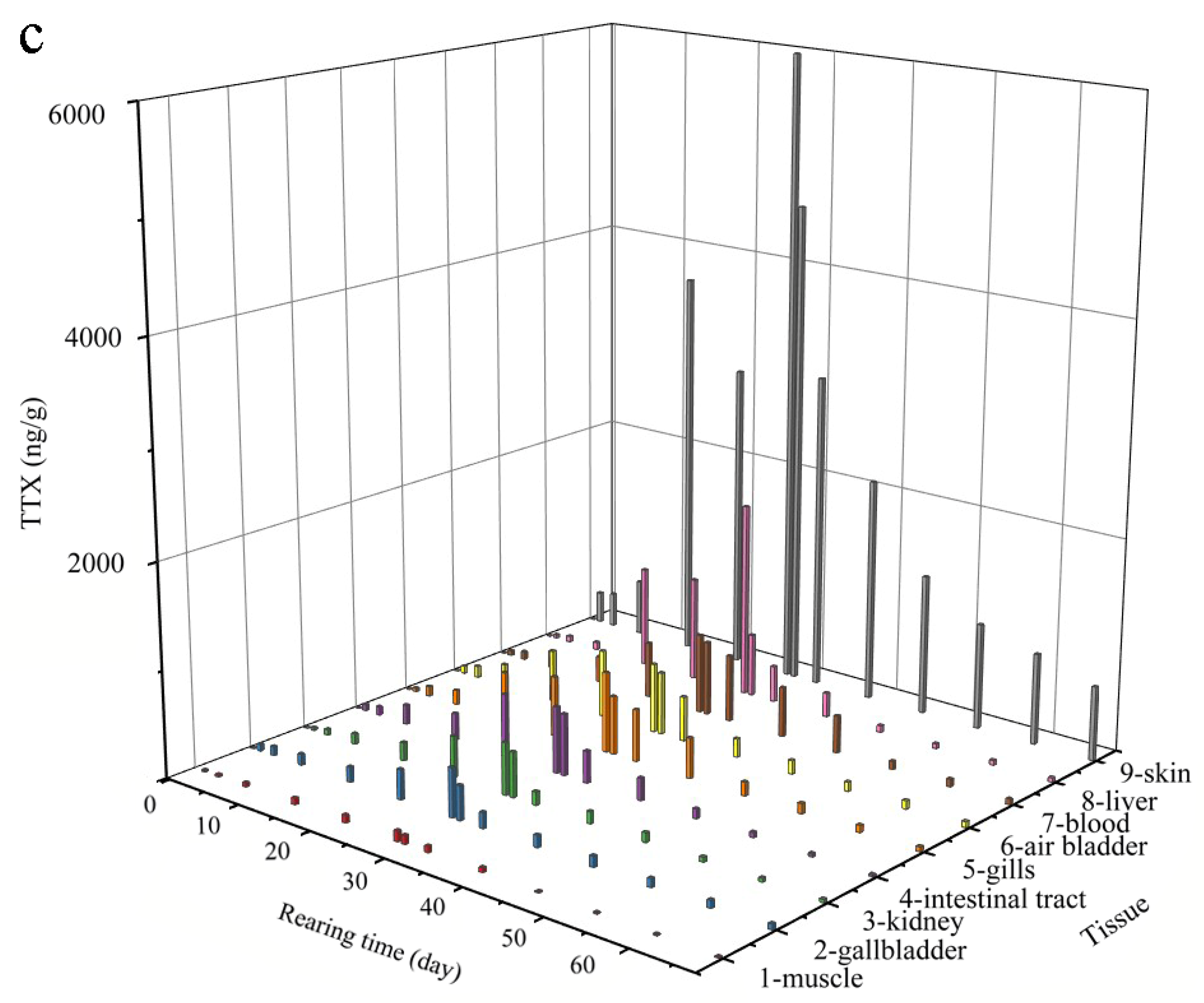
2.1. Anatomical Distribution of TTX
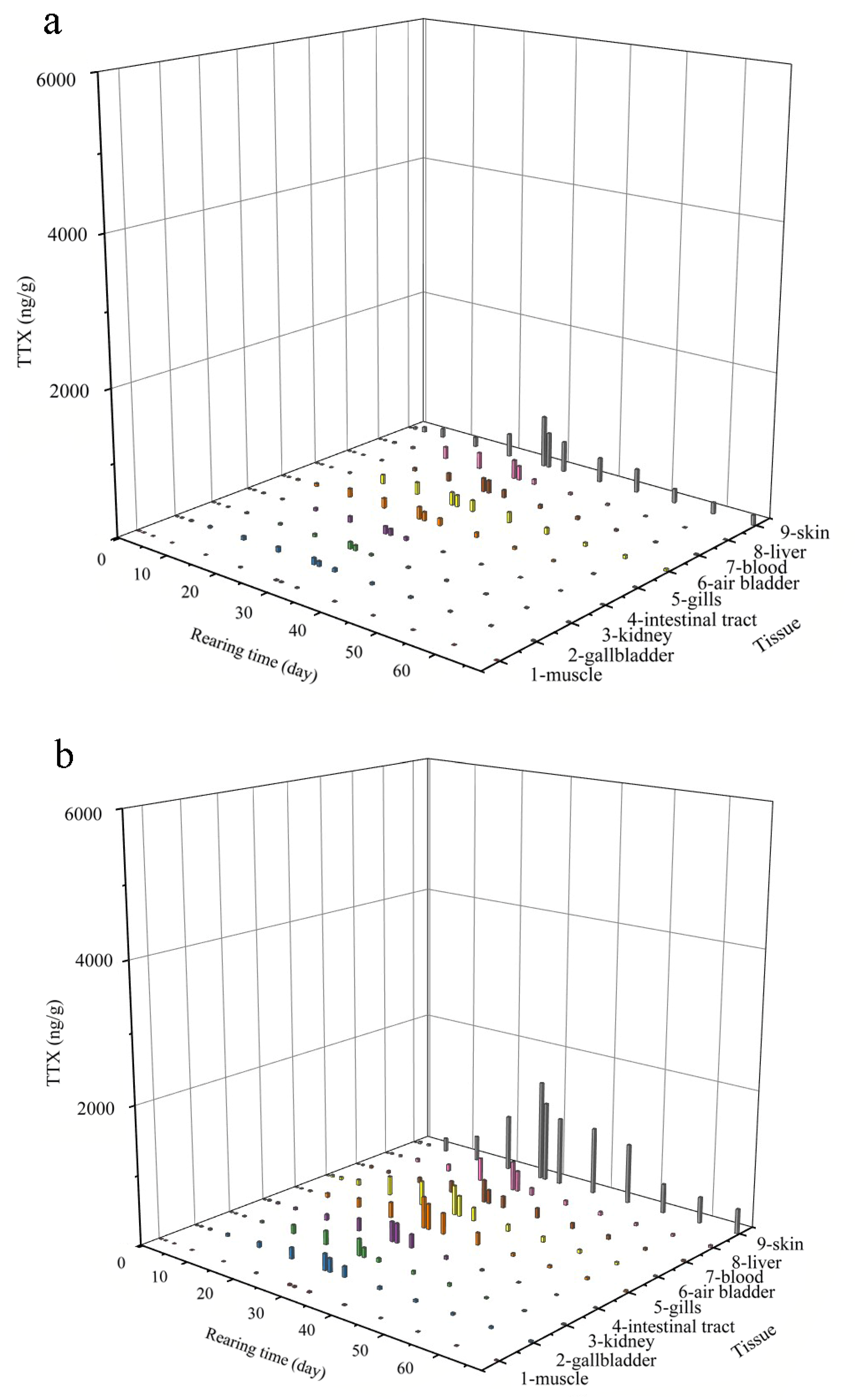
2.2. Patterns in TTX Accumulation and Elimination
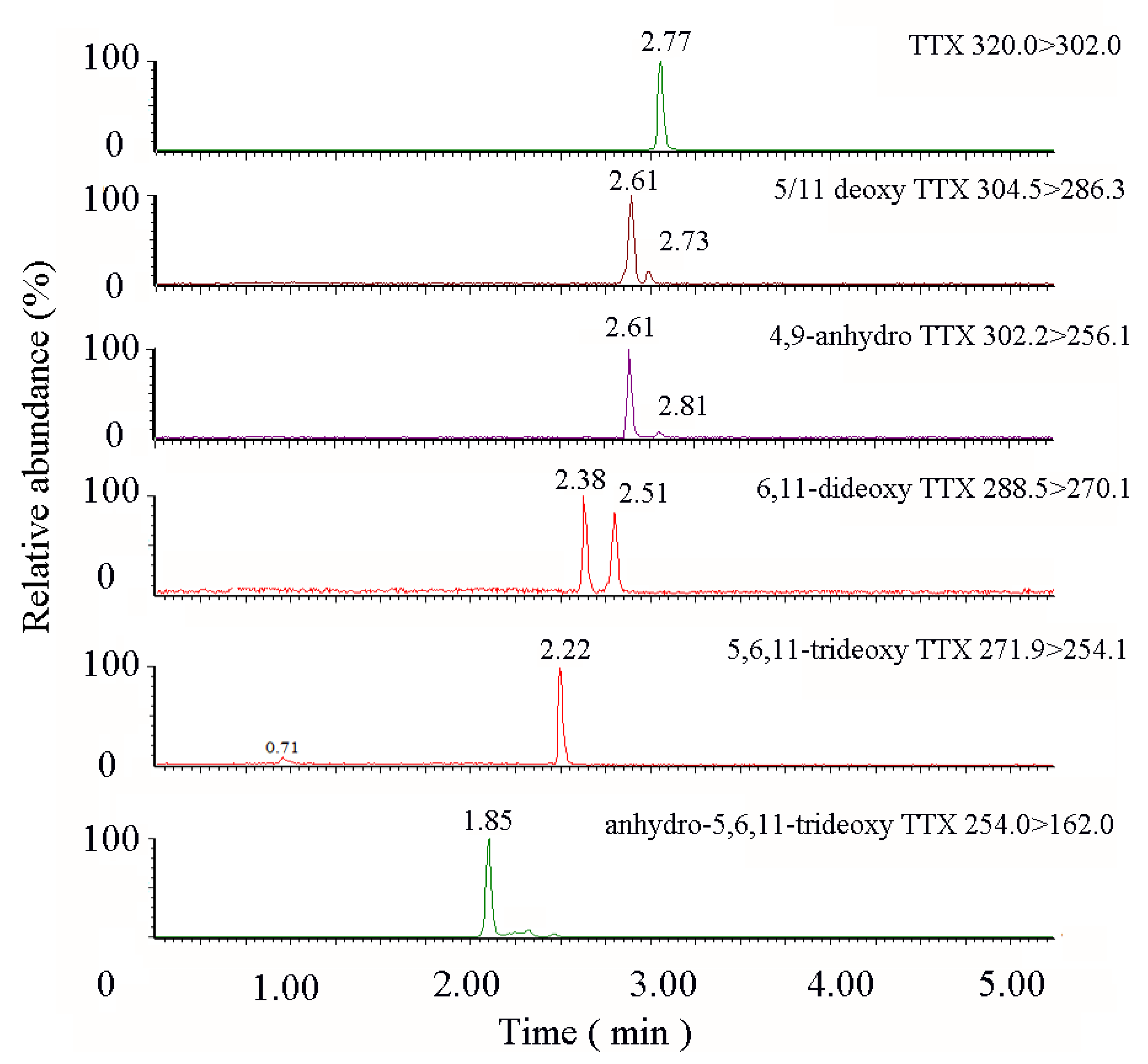
2.3. TTX Derivatives in Takifugu obscurus
3. Discussion
4. Conclusions
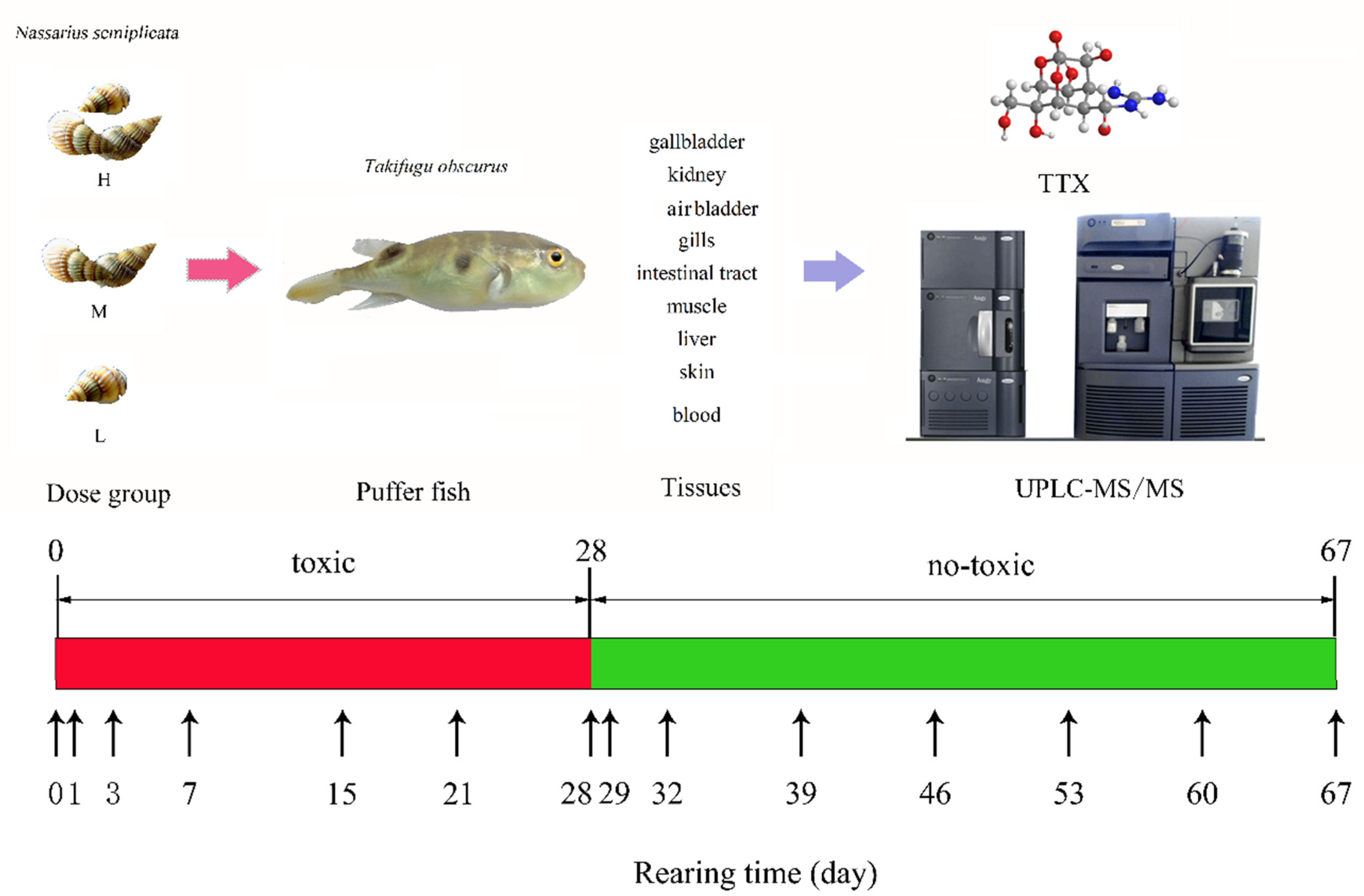
5. Materials and Methods
5.1. Materials
5.1.1. Instruments and Reagents
5.1.2. Biological Materials
5.2. Culture Environment and Diet
5.3. Analysis of TTX and Its Analogues
5.4. Kinetic TTX Elimination Model
5.5. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moczydlowski, E.G. The molecular mystique of tetrodotoxin. Toxicon 2013, 63, 165–183. [Google Scholar] [CrossRef]
- Noguchi, T.; Arakawa, O. Tetrodotoxin--distribution and accumulation in aquatic organisms, and cases of human intoxication. Mar. Drugs 2008, 6, 220–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bane, V.; Lehane, M.; Dikshit, M.; O’Riordan, A.; Furey, A. Tetrodotoxin: Chemistry, toxicity, source, distribution and detection. Toxins 2014, 6, 693–755. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Han, C.; Chen, S.; Li, L.; Zong, J.; Zeng, J.; Mei, G. Response Surface Methodology for the Optimization of Ultrasound-Assisted Extraction of Tetrodotoxin from the Liver of Takifugu pseudommus. Toxins 2018, 10, 529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorentz, M.N.; Stokes, A.N.; Rossler, D.C.; Lotters, S. Tetrodotoxin. Curr. Biol. 2016, 26, R870–R872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colquhoun, D.; Henderson, R.; Ritchie, J.M. The binding of labelled tetrodotoxin to non-myelinated nerve fibres. J. Physiol. 1972, 227, 95–126. [Google Scholar] [CrossRef] [PubMed]
- Narahashi, T. PHARMACOLOGY OF TETRODOTOXIN. J. Toxicol. Toxin Rev. 2001, 20, 67–84. [Google Scholar] [CrossRef]
- Noguchi, T.; Arakawa, O.; Takatani, T. TTX accumulation in pufferfish. Comp. Biochem. Physiol. Part D Genom. Proteom. 2006, 1, 145–152. [Google Scholar] [CrossRef]
- Noguchi, T.; Ebesu, J.S.M. PUFFER POISONING: EPIDEMIOLOGY AND TREATMENT. J. Toxicol. 2001, 20, 1–10. [Google Scholar] [CrossRef]
- Lago, J.; Rodriguez, L.P.; Blanco, L.; Vieites, J.M.; Cabado, A.G. Tetrodotoxin, an Extremely Potent Marine Neurotoxin: Distribution, Toxicity, Origin and Therapeutical Uses. Mar. Drugs 2015, 13, 6384–6406. [Google Scholar] [CrossRef]
- Chen, T.Y.; Hsieh, C.H.; Hwang, D.F. Development of standardized methodology for identifying toxins in clinical samples and fish species associated with tetrodotoxin-borne poisoning incidents. J. Food Drug Anal. 2016, 24, 9–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravaonindrina, N.; Andriamaso, T.H.; Rasolofonirina, N. Puffer fish poisoning in Madagascar: Four case reports. Arch. Inst. Pasteur Madag. 2001, 67, 61–64. [Google Scholar]
- Chowdhury, F.R.; Ahasan, H.A.; Al Mamun, A.; Rashid, A.K.; Al Mahboob, A. Puffer fish (Tetrodotoxin) poisoning: An analysis and outcome of six cases. Trop. Doct. 2007, 37, 263–264. [Google Scholar] [CrossRef] [PubMed]
- Islam, Q.T.; Razzak, M.A.; Islam, M.A.; Bari, M.I.; Basher, A.; Chowdhury, F.R.; Sayeduzzaman, A.B.; Ahasan, H.A.; Faiz, M.A.; Arakawa, O.; et al. Puffer fish poisoning in Bangladesh: Clinical and toxicological results from large outbreaks in 2008. Trans. R. Soc. Trop. Med. Hyg. 2011, 105, 74–80. [Google Scholar] [CrossRef]
- Fernandez-Ortega, J.F.; Morales-de los Santos, J.M.; Herrera-Gutierrez, M.E.; Fernandez-Sanchez, V.; Rodriguez Loureo, P.; Rancano, A.A.; Tellez-Andrade, A. Seafood intoxication by tetrodotoxin: First case in Europe. J. Emerg. Med. 2010, 39, 612–617. [Google Scholar] [CrossRef]
- Katikou, P.; Georgantelis, D.; Sinouris, N.; Petsi, A.; Fotaras, T. First report on toxicity assessment of the Lessepsian migrant pufferfish Lagocephalus sceleratus (Gmelin, 1789) from European waters (Aegean Sea, Greece). Toxicon 2009, 54, 50–55. [Google Scholar] [CrossRef]
- Yakes, B.J.; Deeds, J.; White, K.; Degrasse, S.L. Evaluation of surface plasmon resonance biosensors for detection of tetrodotoxin in food matrices and comparison to analytical methods. J. Agric. Food Chem. 2011, 59, 839–846. [Google Scholar] [CrossRef]
- Arakawa, O.; Takatani, T.; Taniyama, S.; Tatsuno, R. Toxins of Pufferfish—Distribution, Accumulation Mechanism, and Physiologic Functions. Aqua-Biosci. Monogr. 2017, 10, 41–80. [Google Scholar] [CrossRef] [Green Version]
- Tatsuno, R.; Gao, W.; Ibi, K.; Mine, T.; Okita, K.; Nishihara, G.N.; Takatani, T.; Arakawa, O. Profile differences in tetrodotoxin transfer to skin and liver in the pufferfish Takifugu rubripes. Toxicon 2017, 130, 73–78. [Google Scholar] [CrossRef]
- Kono, M.; Matsui, T.; Furukawa, K.; Yotsu-Yamashita, M.; Yamamori, K. Accumulation of tetrodotoxin and 4,9-anhydrotetrodotoxin in cultured juvenile kusafugu Fugu niphobles by dietary administration of natural toxic komonfugu Fugu poecilonotus liver. Toxicon 2008, 51, 1269–1273. [Google Scholar] [CrossRef]
- Noguchi, T.; Arakawa, O.; Takatani, T. Toxicity of pufferfish Takifugu rubripes cultured in netcages at sea or aquaria on land. Comp. Biochem. Physiol. Part D Genom. Proteom. 2006, 1, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Tatsuno, R.; Shikina, M.; Shirai, Y.; Wang, J.; Soyano, K.; Nishihara, G.N.; Takatani, T.; Arakawa, O. Change in the transfer profile of orally administered tetrodotoxin to non-toxic cultured pufferfish Takifugu rubripes depending of its development stage. Toxicon 2013, 65, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Araki, T.; Tatsuno, R.; Nina, S.; Ikeda, K.; Takatani, T.; Arakawa, O. Transfer profile of orally and intramuscularly administered tetrodotoxin to artificial hybrid specimens of the pufferfish Takifugu rubripes and Takifugu porphyreus. Shokuhin Eiseigaku Zasshi 2012, 53, 33–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, K.; Emoto, Y.; Tatsuno, R.; Wang, J.J.; Ngy, L.; Taniyama, S.; Takatani, T.; Arakawa, O. Maturation-associated changes in toxicity of the pufferfish Takifugu poecilonotus. Toxicon 2010, 55, 289–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, W.; Kanahara, Y.; Yamada, M.; Tatsuno, R.; Yoshikawa, H.; Doi, H.; Takatani, T.; Arakawa, O. Contrasting Toxin Selectivity between the Marine Pufferfish Takifugu pardalis and the Freshwater Pufferfish Pao suvattii. Toxins 2019, 11, 470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, K.; Murakami, Y.; Emoto, Y.; Ngy, L.; Taniyama, S.; Yagi, M.; Takatani, T.; Arakawa, O. Transfer profile of intramuscularly administered tetrodotoxin to non-toxic cultured specimens of the pufferfish Takifugu rubripes. Toxicon 2009, 53, 99–103. [Google Scholar] [CrossRef] [Green Version]
- Itoi, S.; Kozaki, A.; Komori, K.; Tsunashima, T.; Noguchi, S.; Kawane, M.; Sugita, H. Toxic Takifugu pardalis eggs found in Takifugu niphobles gut: Implications for TTX accumulation in the pufferfish. Toxicon 2015, 108, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Itoi, S.; Yoshikawa, S.; Asahina, K.; Suzuki, M.; Ishizuka, K.; Takimoto, N.; Mitsuoka, R.; Yokoyama, N.; Detake, A.; Takayanagi, C.; et al. Larval pufferfish protected by maternal tetrodotoxin. Toxicon 2014, 78, 35–40. [Google Scholar] [CrossRef] [Green Version]
- Itoi, S.; Suzuki, M.; Asahina, K.; Sawayama, E.; Nishikubo, J.; Oyama, H.; Takei, M.; Shiibashi, N.; Takatani, T.; Arakawa, O.; et al. Role of maternal tetrodotoxin in survival of larval pufferfish. Toxicon 2018, 148, 95–100. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Okoshi, N.; Watanabe, K.; Araki, N.; Yamaki, H.; Shoji, Y.; Terakawa, T. Localization of pufferfish saxitoxin and tetrodotoxin binding protein (PSTBP) in the tissues of the pufferfish, Takifugu pardalis, analyzed by immunohistochemical staining. Toxicon 2013, 72, 23–28. [Google Scholar] [CrossRef]
- Dyble, J.; Gossiaux, D.; Landrum, P.; Kashian, D.R.; Pothoven, S. A kinetic study of accumulation and elimination of microcystin-LR in yellow perch (Perca flavescens) tissue and implications for human fish consumption. Mar. Drugs 2011, 9, 2553–2571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, J.H.; Lee, J.S.; Yotsu-Yamashita, M. LC/MS analysis of tetrodotoxin and its deoxy analogs in the marine puffer fish Fugu niphobles from the southern coast of Korea, and in the brackishwater puffer fishes Tetraodon nigroviridis and Tetraodon biocellatus from Southeast Asia. Mar. Drugs 2010, 8, 1049–1058. [Google Scholar] [CrossRef] [PubMed]
- Tonon, L.A.C.; de Azevedo, G.P.R.; Monteiro, A.F.; Bernardi, D.I.; Gubiani, J.R.; Ioca, L.P.; Mattsson, H.K.; Moreira, A.P.B.; Gomes, A.F.; Pires Junior, O.R.; et al. New tetrodotoxin analogs in Brazilian pufferfishes tissues and microbiome. Chemosphere 2020, 242, 125211. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Xu, J.; Liang, S.; Ren, D.; Yan, X.; Bao, B. A novel TTX-producing Aeromonas isolated from the ovary of Takifugu obscurus. Toxicon 2010, 56, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Kudo, Y.; Yasumoto, T.; Konoki, K.; Cho, Y.; Yotsu-Yamashita, M. Isolation and structural determination of the first 8-epi-type tetrodotoxin analogs from the newt, Cynops ensicauda popei, and comparison of tetrodotoxin analogs profiles of this newt and the puffer fish, Fugu poecilonotus. Mar. Drugs 2012, 10, 655–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagashima, Y.; Tanaka, N.; Shimakura, K.; Shiomi, K.; Shida, Y. Occurrence of tetrodotoxin-related substances in the nontoxic puffer Takifugu xanthopterus. Toxicon 2001, 39, 415–418. [Google Scholar] [CrossRef]
- Zhang, X.; Yan, Z.; Wang, Y.; Jiang, T.; Wang, J.; Sun, X.; Guo, Y. Immunoaffinity chromatography purification and ultrahigh performance liquid chromatography tandem mass spectrometry determination of tetrodotoxin in marine organisms. J. Agric. Food Chem. 2015, 63, 3129–3134. [Google Scholar] [CrossRef]
- Gall, B.G.; Stokes, A.N.; French, S.S.; Brodie, E.D., 3rd; Brodie, E.D., Jr. Female newts (Taricha granulosa) produce tetrodotoxin laden eggs after long term captivity. Toxicon 2012, 60, 1057–1062. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exposure Dose Group | TTX in Feed (ng/g) | Feed Intake (g) | Total TTX (ng) | Actual Accumulation of TTX (ng) | Accumulation Ratio (%) |
|---|---|---|---|---|---|
| Low-dose group | 100 | 25.67 ± 2.1 | 2567 ± 210 | 1032 ± 104 | 40.20 ± 5.2 |
| Medium-dose group | 210 | 26.03 ± 3.2 | 5466 ± 320 | 1955 ± 110 | 35.76 ± 4.9 |
| High-dose group | 600 | 23.75 ± 2.4 | 14250 ± 240 | 5420 ± 173 | 38.03 ± 4.1 |
| Dose Group | Tissues | TTX-C0 Day 28 (ng/g) | TTX-Ct Day 67 (ng/g) | Elimination Rate K (d−1) | Half-Life Period B1/2 (d) |
|---|---|---|---|---|---|
| Low-dose group | gallbladder | 87.96 | 9.53 | 0.084 | 8.25 |
| kidney | 91.53 | 8.55 | 0.391 | 1.77 | |
| air bladder | 178.1 | 29.04 | 0.042 | 16.50 | |
| gills | 170.29 | 6.72 | 0.088 | 7.88 | |
| intestinal tract | 101.23 | 3.33 | 0.236 | 2.94 | |
| muscle | 10.68 | 2.32 | 0.040 | 9.16 | |
| liver | 262.18 | 4.42 | 0.387 | 1.79 | |
| skin | 717.34 | 135.66 | 0.042 | 16.50 | |
| blood | 189.35 | 14.42 | 0.105 | 6.60 | |
| Medium-dose group | gallbladder | 238.84 | 19.79 | 0.132 | 5.25 |
| kidney | 244.47 | 14.03 | 0.496 | 1.40 | |
| air bladder | 458.22 | 22.909 | 0.138 | 5.02 | |
| gills | 435.86 | 10.49 | 0.100 | 6.93 | |
| intestinal tract | 291.46 | 8.14 | 0.156 | 4.44 | |
| muscle | 20.51 | 2.79 | 0.056 | 12.38 | |
| liver | 427.62 | 27.88 | 0.399 | 1.74 | |
| skin | 1484.20 | 358.79 | 0.035 | 19.80 | |
| blood | 333.96 | 22.25 | 0.068 | 10.19 | |
| High-dose group | gallbladder | 460 | 50.25 | 0.100 | 6.93 |
| kidney | 486.47 | 29.07 | 0.320 | 2.17 | |
| air bladder | 757 | 43.86 | 0.098 | 7.07 | |
| gills | 650.12 | 54.82 | 0.075 | 9.24 | |
| intestinal tract | 622.93 | 18.1 | 0.12 | 5.78 | |
| muscle | 99.36 | 7.4 | 0.092 | 7.53 | |
| liver | 1840.3 | 41.18 | 0.952 | 0.73 | |
| skin | 6000 | 699.35 | 0.076 | 9.12 | |
| blood | 750.13 | 50.1 | 0.059 | 11.75 |
| TTX Analogues | Takifugu obscurus Tissues | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Gallbladder | Kidney | Air Bladder | Gills | Intestinal Tract | Muscle | Liver | Skin | Blood | |
| TTX | √ | √ | √ | √ | √ | √ | √ | √ | √ |
| 5-deoxy TTX | √ | √ | √ | √ | √ | √ | √ | √ | √ |
| 11-deoxy TTX | √ | √ | √ | √ | √ | √ | √ | √ | √ |
| 4,9-anhydro TTX | √ | √ | √ | √ | √ | / | √ | √ | √ |
| 6,11-dideoxy TTX | / | √ | √ | √ | √ | √ | √ | √ | √ |
| 5,6,11-trideoxy TTX | √ | / | √ | √ | √ | / | √ | √ | √ |
| anhydro-5,6,11-trideoxy TTX | / | / | / | / | / | / | / | √ | / |
| Analyte | Retention Time(min) | Precursor Ion(m/z) | Product Ion(m/z) | Cone Voltage (V) | Collision Energy(eV) |
|---|---|---|---|---|---|
| TTX | 2.77 | 320.0 | 302.0 * 161.8 | 45 | 25 35 |
| 5-deoxy TTX/ 11-deoxy TTX | 2.61 | 304.5 | 286.29 162.10 | 40 | 25 20 |
| 4,9-anhydro TTX | 2.61 | 302.20 | 256.13 161.83 | 40 | 35 25 |
| 6,11-dideoxy TTX | 2.51 | 288.54 | 270.11 224.69 | 40 | 25 25 |
| 5,6,11-trideoxy TTX | 2.22 | 271.90 | 254.11 162.07 | 40 | 25 20 |
| anhydro-5,6,11-trideoxy TTX | 1.85 | 254.02 | 161.98 | 40 | 25 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Zong, J.; Chen, S.; Li, M.; Lu, Y.; Wang, R.; Xu, H. Accumulation and Elimination of Tetrodotoxin in the Pufferfish Takifugu obscurus by Dietary Administration of the Wild Toxic Gastropod Nassarius semiplicata. Toxins 2020, 12, 278. https://doi.org/10.3390/toxins12050278
Zhang X, Zong J, Chen S, Li M, Lu Y, Wang R, Xu H. Accumulation and Elimination of Tetrodotoxin in the Pufferfish Takifugu obscurus by Dietary Administration of the Wild Toxic Gastropod Nassarius semiplicata. Toxins. 2020; 12(5):278. https://doi.org/10.3390/toxins12050278
Chicago/Turabian StyleZhang, Xiaojun, Jingjing Zong, Si Chen, Menglong Li, Yibo Lu, Ruirui Wang, and Hanxiang Xu. 2020. "Accumulation and Elimination of Tetrodotoxin in the Pufferfish Takifugu obscurus by Dietary Administration of the Wild Toxic Gastropod Nassarius semiplicata" Toxins 12, no. 5: 278. https://doi.org/10.3390/toxins12050278