
Response of Growth-Related Traits of Submerged Macrophytes to Light Reduction: A Meta-Analysis

1
Shanghai Investigation, Design & Research Institute Co., Ltd., Shanghai 200335, China
2
Yangtze Eco-Environment Engineering Research Center (Shanghai), China Three Gorges Corporation, Shanghai 200335, China
3
Northeast Asia Ecosystem Carbon Sink Research Center (NACC), Center for Ecological Research, Key Laboratory of Sustainable Forest Ecosystem Management-Ministry of Education, School of Forestry, Northeast Forestry University, Harbin 150040, China
*
Author to whom correspondence should be addressed.
Sustainability 2023, 15(7), 5918; https://doi.org/10.3390/su15075918
Submission received: 22 February 2023
/
Revised: 21 March 2023
/
Accepted: 27 March 2023
/
Published: 29 March 2023
(This article belongs to the Section Sustainability, Biodiversity and Conservation)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Underwater light reduction is presumably becoming more frequent and intensified due to eutrophication and algal blooms, which may significantly affect submerged macrophytes’ growth. However, a comprehensive understanding of how light reduction influences growth-related traits and responses is currently lacking. Here, we compiled data from 333 records of 62 published works that used controlled experiments to explore the responses of functional traits associated with growth to light reduction. Our results indicated that light reduction significantly decreased the relative growth rate (RGR), ramet number (RM), shoot biomass (SB), root biomass (RB), soluble carbohydrates (SC), and leaf number (LN) by 38.2%, 60.0%, 59.2%, 55.4%, 30.0%, and 56.1%, respectively, but elevated the chlorophyll content (Chl) by 25.8%. Meanwhile, the responses of RGR to light reduction increased significantly with the responses of RM, SB, RB, and root-to-shoot ratio (R/S). Considering the relationships among the growth-related traits, we further found that the responses of RGR to light reduction were mainly driven by the RM rather than leaf photosynthetic capability, indicating the importance of tiller ability under low light stress. We also identified a tipping point of the response of RGR to light reduction, which might be incorporated into hydrophyte dynamic models to improve precision. Our results highlight the importance of growth-related traits, andthese traits may need to be incorporated into models to improve the prediction of distribution and area for submerged species or to provide guidance for the restoration and sustainable development of aquatic ecosystems.
1. Introduction
Submerged macrophytes, as the main primary producers, play critical roles in structuring communities and stabilizing the functions of aquatic ecosystems [1,2,3], such as providing refuge and food for fish and zooplankton [4,5], reducing phytoplankton biomass through allelopathic effects [6,7], removing nutrients from water [8], and controlling sediment resuspension [9]. Additionally, submerged macrophytes also provide food and financial products for humans to advance the sustainability of aquatic ecosystems. However, the decline of submerged macrophytes is accelerating worldwide due to eutrophication and degradation of light conditions [10,11,12], expediting the collapse of aquatic ecosystems. Thus, successfully reviving submerged macrophytes is a crucial pathway for maintaining the functions and achieving sustainable development of aquatic ecosystems.
The growth and reproduction of submerged macrophytes are affected by abiotic and biotic factors such as light, nutrients, temperature, water level, and phytoplankton growth [13,14]. Among these factors, light availability determines the growth, reproduction, and distribution of submerged macrophytes [15,16], which could explain as much as 77% of the variation in macrophytes’ abundance [17]. In addition, light reception can facilitate inorganic carbon acquisition, photosynthesis, and growth of submerged macrophytes [18] as well as regulate oxygen evolution, thus enabling plants to adapt to anoxic sediments through root aeration [19,20]. Therefore, understanding the responses of submerged macrophytes to light is crucial for the ecological restoration of aquatic ecosystems.
Functional traits have been widely used to illuminate the interactions between plants and their environments in terrestrial ecosystems [21,22,23]. Hydrophyte adaptive strategies in aquatic ecosystems have been tested and can be directly compared with strategies for terrestrial plants using a functional trait-based approach [24,25]. Functional traits in hydrophytes are mainly assessed by referring to morphology (plant height and branch), production (biomass allocation and growth rate), reproduction (bud numbers), physiology (photosynthesis rate), and biochemistry (elemental contents of tissue) [26]. Under environmental stress, the plasticity of functional traits may enable macrophytes to be more adaptable. For instance, submerged macrophytes could ease low light stress via growth-related and physiological trait adjustments (e.g., shoot length, biomass allocation, and leaf photosynthesis parameters) to optimize light utilization and increase survival probability [27,28,29]. Thus, functional traits are considered vital drivers in regulating the growth strategies of submerged macrophytes [26,30].
In recent decades, substantial studies have been performed to examine the effects of light or shading on the functional traits of submerged macrophytes, which have considerably improved our understanding of the mechanisms underlying these light effects [20,25,31]. For example, after light reduction, Myriophyllum spicatum increased the plant height and branch length to maximize light interception and photosynthesis [27]. Moderate light reduction induced submerged macrophytes to increase chlorophyll content and glutamine synthetase to increase photosynthetic efficiency. However, extremely low light decreased photosynthesis and nutrient metabolic activity to avoid oxidative damage due to low antioxidant reactions [32]. Based on a field investigation, Arthaud et al. (2021) [31] identified tolerance and avoidance strategies for submerged macrophytes under reduced light. The tolerance strategy was characterized by a high investment in leaf area and photosynthetic efficiency to increase light energy utilization, and a greater investment in vertical growth characterized the avoidance strategy to obtain more light in shallow water [31]. Additionally, some studies have also shown that the responses of submerged macrophytes to reduced light occur mainly in terms of ramet and leaf numbers [33,34], but these responses might change with the percentage of light reduction [35]. Nonetheless, the current understanding of the global pattern of functional traits in response to light reduction remains highly uncertain due to the lack of complete datasets, which limits our ability to predict the distribution and dynamics of submerged macrophytes.
Moreover, the effects of light reduction on functional traits investigated in previous studies were highly diverse, which might be related to differences in species, leaf shapes, experimental durations, and light reduction intensities. For instance, after light reduction, Elodea canadensis showed decreases in ramet numbers and total biomass while biomass per ramet remained the same. In contrast, Myriophyllum spicatum and Zosterella dubia showed decreases in both ramet number and biomass per ramet [36]. To adapt to low-light environments, submerged macrophytes with flat leaves presented greater leaf areas, leaf dry weights, specific leaf areas, and Fv/Fm ratios, while those with needle leaves had greater Chl a/b ratios [3]. Shading had no significant effect on the Chl contents of waterweed over 8 weeks but significantly elevated Chl contents after 10 weeks [37]. Importantly, light reduction might affect submerged macrophytes’ relative growth rate by decreasing it, increasing it, or affecting it not at all depending on different species and percentage of light reduction [28,34,38,39,40]. In addition, whether the treatment method (artificial light source vs. shading), plant type (seedlings, mature, or shoots) and experiment end season influence the effects of light reduction has not been studied. Therefore, it is necessary to integrate the available data from different studies to reveal the patterns and mechanisms of functional traits in response to light reduction.
To support the use of results from individual studies to elucidate a general tendency, a meta-analysis, a quantitative and scientific synthesis tool to provide reliable conclusions [41], was conducted using 333 records of 62 published works describing the use of control experiments to evaluate the responses of 12 functional traits to light reduction. Specifically, we aimed to (1) examine global patterns of light-reduction-induced changes in the traits of submerged macrophytes and (2) investigate the influences of moderators (e.g., experimental conditions, plant type, and leaf shape) on traits and related responses to light reduction.
2. Materials and Methods
2.1. Traits Selection
Twelve functional traits of submerged macrophytes were included in our study: relative growth rate (RGR), shoot biomass (SB), root biomass (RB), root-to-shoot ratio (R/S), plant height (PH), ramet number (RM), leaf number (LN), soluble carbohydrates (SC), net photosynthesis rate (NPR), leaf maximal quantum yield of photosystem II complex (Fv/Fm), chlorophyll content (Chl), and chlorophyll a/b ratio (Chl a/b). These traits can reflect submerged plant growth performance [3,16,22]. Moreover, we also selected these traits based on data availability, as they were relatively easy to measure and thus were more frequently reported in the analyzed publications.
2.2. Data Source and Collection
A search for studies related to the responses of submerged plants to light availability was carried out using the Web of Science, Google Scholar, and China National Knowledge Infrastructure (CNKI) search engines. The search was performed between March and May 2022 using the following keyword string: ((“submer* plant*” or “submer* macrophyt*”) and (biomass or morpholog* or physiolog* or photosynthe* or growth) and (light or shad*)). Then, we screened the studies using the following criteria: (1) at least two light levels in the experimental design, (2) different light groups had similar sediment and water conditions, (3) the methods used to manipulate light were recorded, and (4) at least 1 of the 12 traits was reported at the species level. Furthermore, we excluded studies in which the different light levels resulted from the differences in water depth and turbidity due to the combined effects of other environmental factors such as hydraulic pressure and silt. We also excluded studies that investigated trait variation across a large water area without focusing on the effect of reduced light. A flowchart of the process and output of the literature search is provided in the supplementary materials (Figure S1).
According to these criteria, we compiled 62 published works between 1996 and 2022 that focused on intraspecific photosynthesis and growth trait variations of 37 species along light gradients. These submerged plants were sampled at 51 sites in East Asia, Oceania, Europe, and North America (Figure S2). Every study reported data for one or more traits considered in the analysis (Table S1).
We extracted the data directly from tables and text or indirectly from graphs using GetData Graph Digitizer (version 2.25). The mean value, standard deviation (or standard error), and sample size for each trait of a given species at each light level in each study were extracted. If the standard deviation was not reported, we calculated it by multiplying the standard error by the square root of the sample size. If only median values and ranges were reported, we estimated the mean value and standard deviation as proposed by Hozo et al. (2005, http://vassarstats.net/median_range.html, accessed on 23 May 2022) [42].
We regarded the highest and other light levels as control and treatment groups, respectively. Then, we calculated the percentage differences in light intensity between the control and treatment groups as the percentage of light reduction. In addition, we extracted the submerged plant type at the start of the experiment (mature, seedlings, shoots, tubers, or buds), end season, treatment method, and experimental duration for each study. We also extracted the scientific names of species and geographic coordinates of the locations at which species were sampled. To avoid species synonyms, the scientific names of the submerged plants in each study were confirmed using The Plant List (http://www.theplantlist.org/, accessed on 28 May 2022) and Tropicos (http://www.tropicos.org/, accessed on 28 May 2022).
2.3. Data Analysis
All data analysis and figure generation were performed in the R 4.1.3 statistical platform [43].
We calculated the natural log-transformed response ratio (lnRR) for each species’ traits along with light reduction gradients as the effect size in our meta-analysis [44]. The lnRR was calculated as follows:
where Xt and Xc were the traits’ mean values for the control and treatment pairs, respectively.
Because some selected studies reported traits for multiple light levels along a gradient, we calculated the lnRR for each light level compared with the highest light level. This did not meet the assumption of independence of effect sizes [45] (i.e., multiple treatments with a common control). Therefore, we computed the variance-covariance (VCV) matrix for each trait according to the method of Lajeunesse (2011) [46]. Before computing the VCV matrix, the sampling variance (v) of each study case’s effect size was determined as follows:
where SDt, nt and SDc, nc are the treatment and control groups’ standard deviations and sample sizes, respectively.
First, some missing standard deviations were imputed using the coefficient of variation from all complete study cases using the impute_SD function in the metagear package [47,48,49]. For each trait, the effect sizes and sampling variances were calculated using the escalc function in the metafor package [50], and the VCV matrix was computed using the covariance_commonControl function in the metagear package [48].
Then, we estimated the weighted average effect sizes (lnRR++) and 95% confidence intervals (CIs) of each trait with multilevel linear mixed-effect models using the rma.mv function in the metafor package [50]. The lnRR++ was significant if the estimated CIs did not overlap zero. The inverse of the variance of the VCV matrix was used to weight the precision of the lnRRs when averaged. The mixed-effect models were fitted with crossed, nested, and non-nested random effect terms: (1|reference/ID) + (1|species), which included the reference of each study and the species as non-nested random effects [51,52]. Considering the variation among different cases reported within a single publication (i.e., with the same reference), we nested each study case ID within the reference level in the random structure of the mixed-effect models.
To explore the influence of plant type, treatment method, end season, experimental periods, and the percentage of light reduction on the lnRRs of each trait, these factors were used as moderators in the multilevel linear mixed-effect models for meta-regression using the rma.mv function in the metafor package [50]. The heterogeneity of effect size was evaluated via a Q-test to determine whether these moderators significantly influenced the response of each trait to light reduction. The parameter and confidence interval estimate of moderators were based on the mixed-effect model that did not contain the intercept term. To improve the interpretation, the lnRR++ was transformed back to the percentage change (%) as follows:
To test the robustness of the meta-analysis for each trait, a file drawer analysis was performed using the fsn function in the metafor package [50]. The Rosenberg method was used to calculate the number of study cases averaging null results that would have to be added to the given outcomes to reduce the significance level of the weighted average effect size to 0.05 [50,53]. To test for potential publication bias, a funnel plot was made using the funnel function in the metafor package [50]. The modification of Egger’s test proposed by Nakagawa and Santos (2012) [54] was used to evaluate the funnel plot’s symmetry of the residuals.
Finally, to explore the pairwise relationships of the responses of traits to light reduction, asymmetric regressions were performed using the rma.mv function considering the random effect and the weight of the lnRRs. Specifically, the lnRRs of pairwise traits were used as response variables and moderators in the meta-regression analysis, respectively, and the variance of VCV matrix and “(1|reference/ID) + (1|species)” were used as the weight and random effect term, respectively. The pairwise Pearson correlation relationships of lnRRs of traits were also calculated to check the influence of random effect and the weight. The correlation coefficients were calculated using the corr.test function in the psych package [55], and a corrplot was made with the corrplot function in the corrplot package [56].
3. Results
3.1. Effects of Light Reduction on the Growth of Submerged Macrophytes
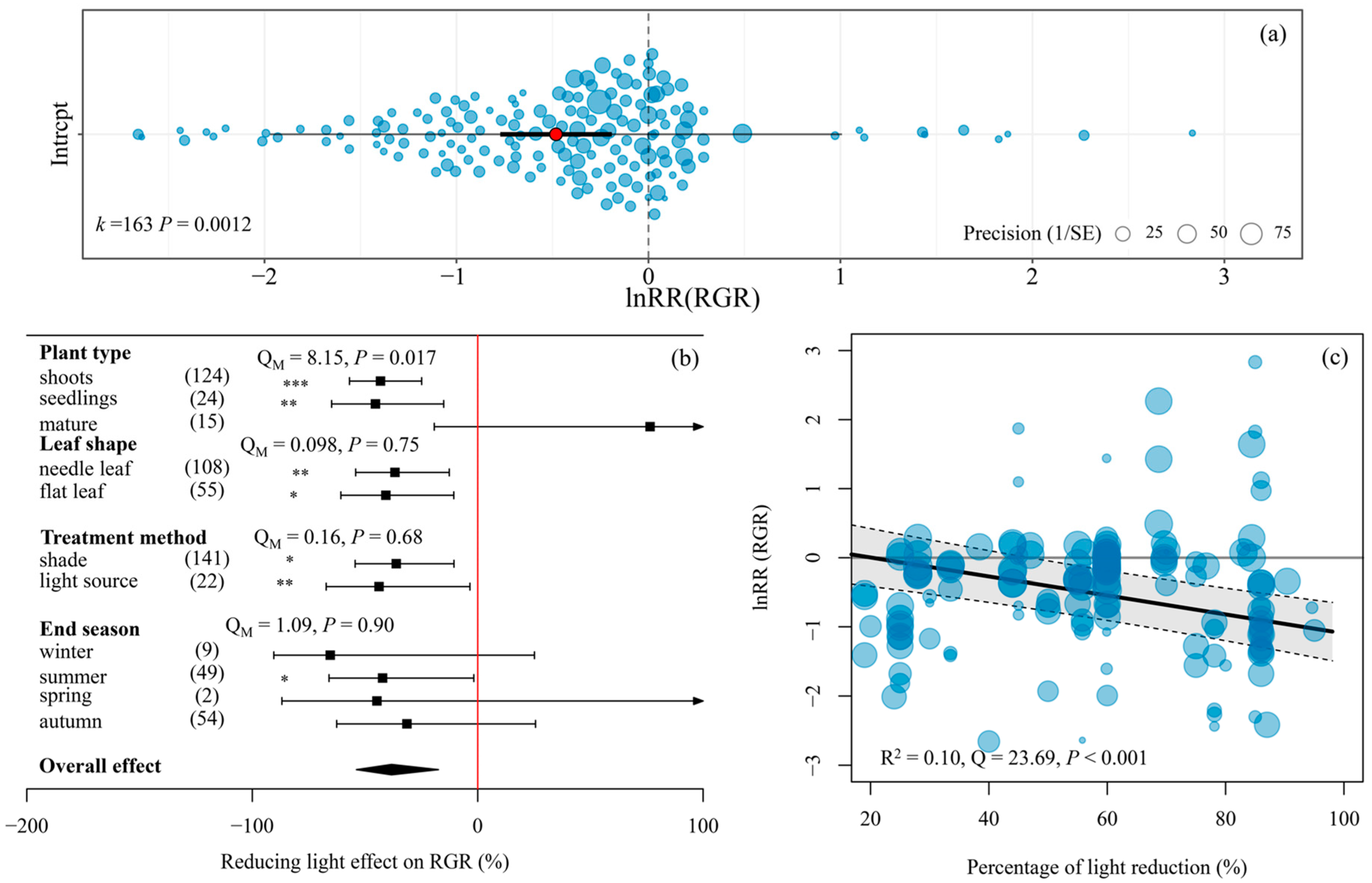
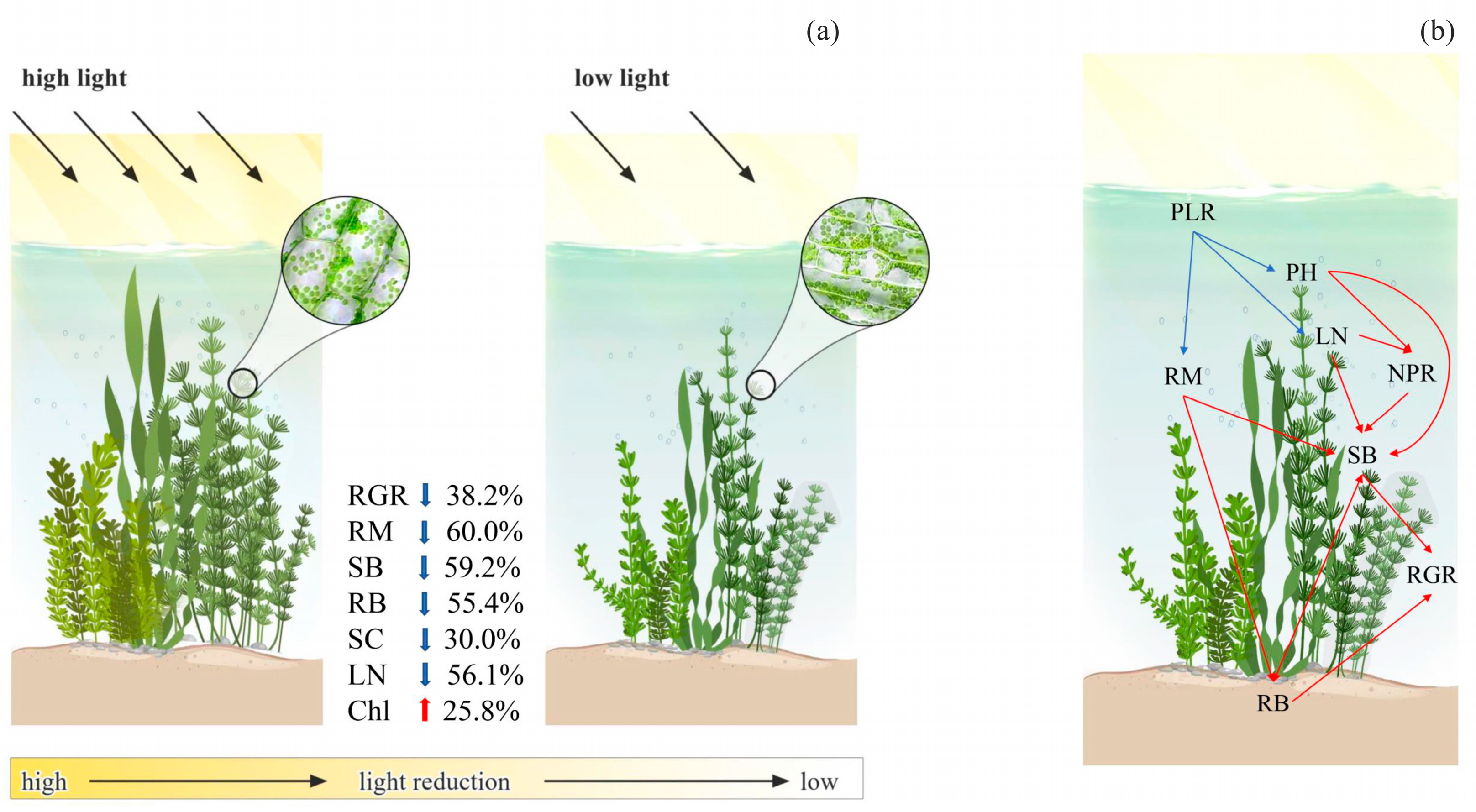
Our meta-analysis showed that light reduction resulted in a significant 38.2% decrease in the relative growth rate (RGR) of submerged macrophytes (Figure 1). The negative effects of light reduction on RGR resulted from the shoots and seedlings of submerged macrophytes (Figure 1b). Moderators (leaf shape, treatment method, experimental duration, and end season) had non-significant moderating effects on the response (lnRR) of RGR to light reduction (Figure 1b). Light reduction had significantly negative effects on RGR when experiments ended in the summer but a non-significant effect when experiments ended in other seasons (Figure 1b). With increases in the percentage of light reduction, the lnRR of RGR to light reduction decreased significantly (Figure 1c).
3.2. Effects of Light Reduction on the Whole Plant and Leaf Traits
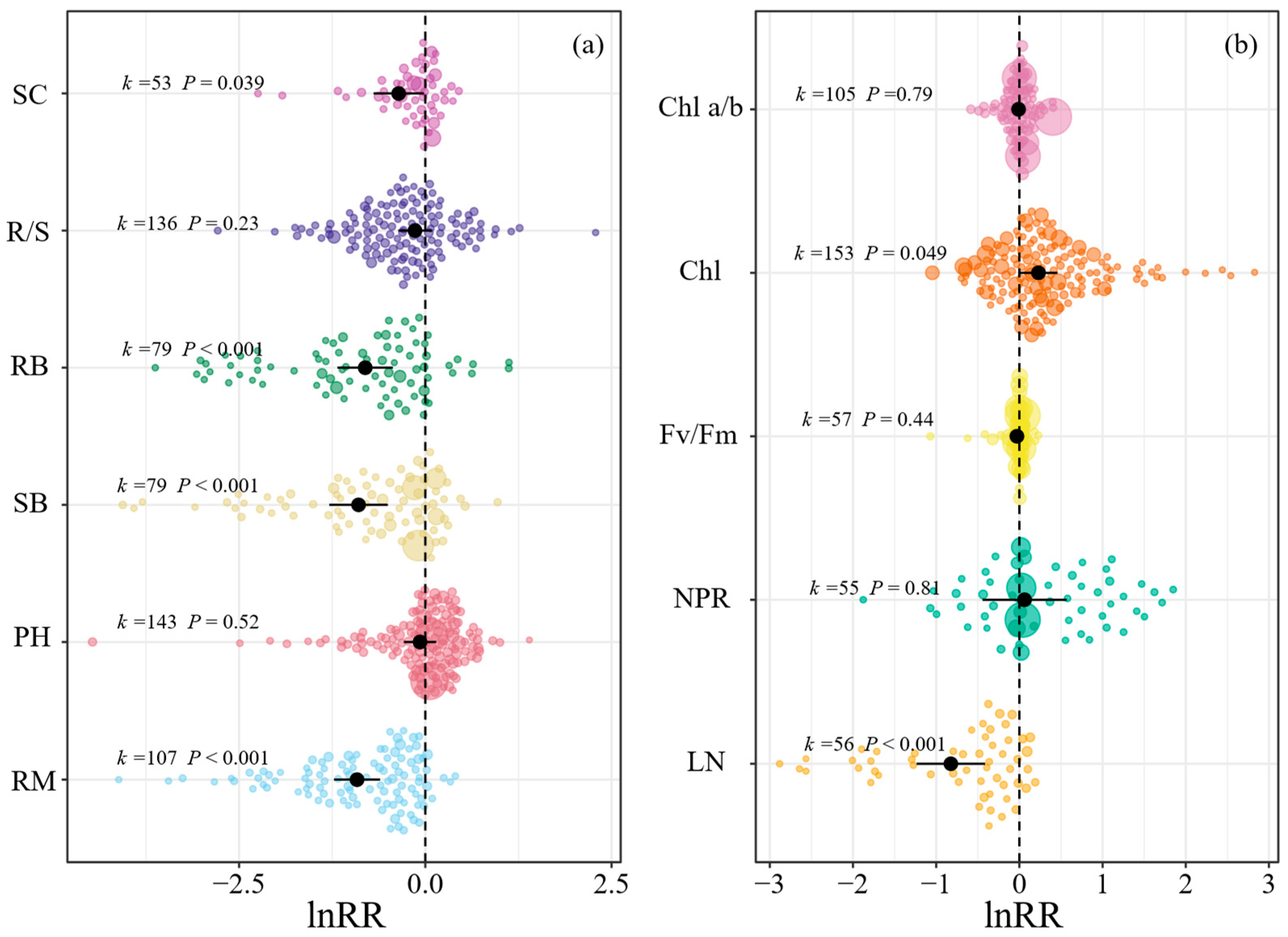
For the whole-plant traits, light reduction led to 60.0%, 59.2%, 55.4%, and 30.0% decreases in ramet number (RM), shoot biomass (SB), root biomass (RB), and soluble carbohydrates (SC), respectively (Figure 2a). For leaf traits, light reduction resulted in a 56.1% decrease in leaf number (LN) but a 25.8% increase in chlorophyll content (Chl, Figure 2b). Light reduction had non-significant influences on plant height (PH), root-to-shoot biomass ratio (R/S), net photosynthesis rate (NPR), maximal quantum yield of photosystem II complex (Fv/Fm), and chlorophyll a/b ratio (Chl a/b, Figure 2).
Most of the moderators did not significantly affect the response of whole-plant traits to light reduction, except for leaf shape and plant type (Figure S3). Specifically, light reduction had greater negative effects on the PH of plants grown from seedlings than those grown from other starting materials (Figure S3b). The lnRRs of R/S to light reduction were significantly negative for plants grown from shoots but non-significant for those grown from tubers, seedlings, and mature plants (Figure S3e). The responses of leaf traits to light reduction were altered by all moderators except the treatment method (Figure S4). The lnRRs of NPR and Chl a/b were influenced by the experiment end season, and the lnRRs of Chl were changed by plant type. The lnRRs of NPR for needle leaves were significantly greater than those for flat leaves. The lnRRs of R/S increased with experimental duration (Figure S5). Moreover, the lnRRs of RM, SB, RB, PH, LN, and Chl a/b decreased with the increasing percentage of light reduction (Figure S6).
3.3. Influence of Whole Plant and Leaf Traits on the Responses of RGR to Light Reduction
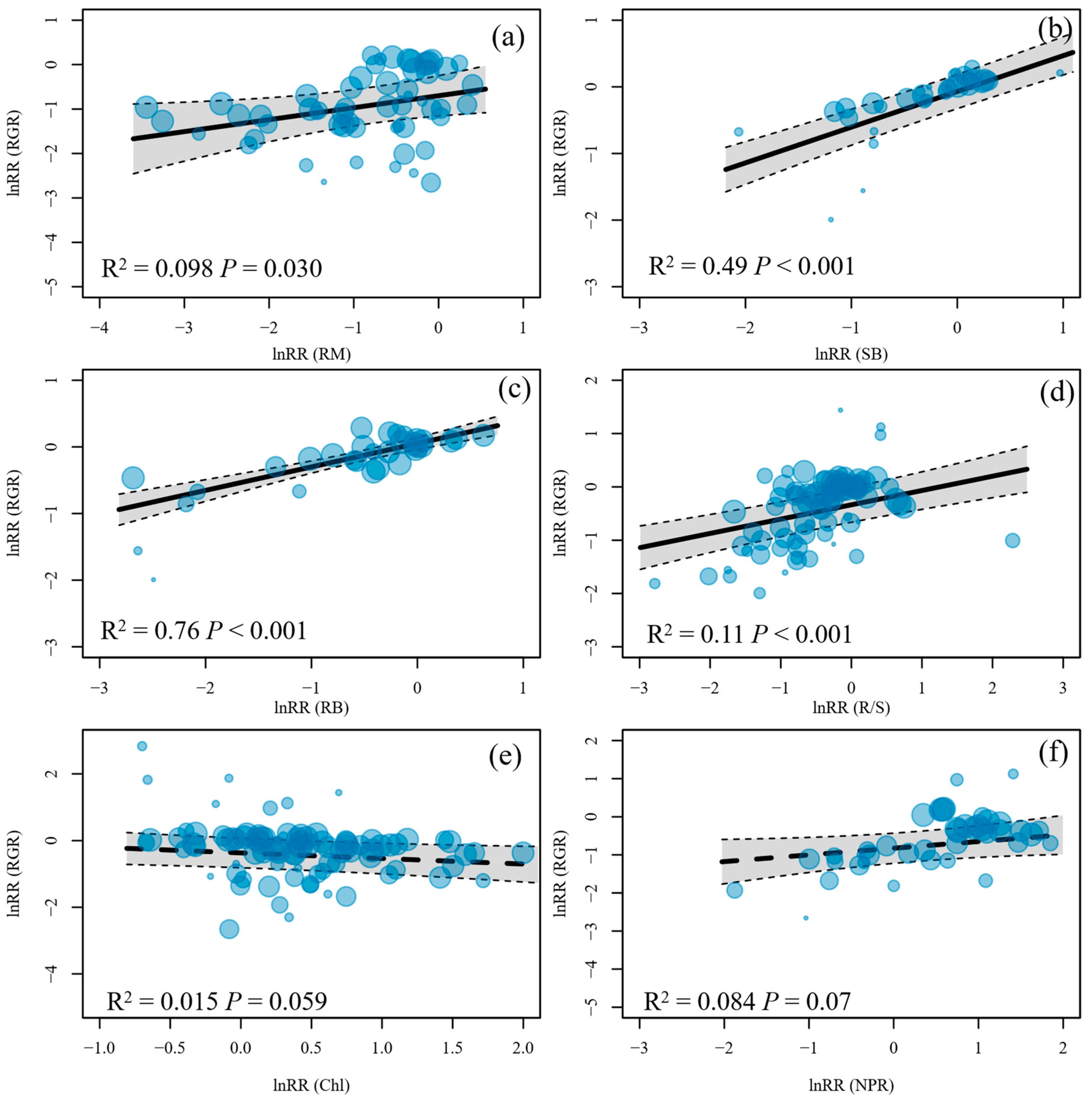
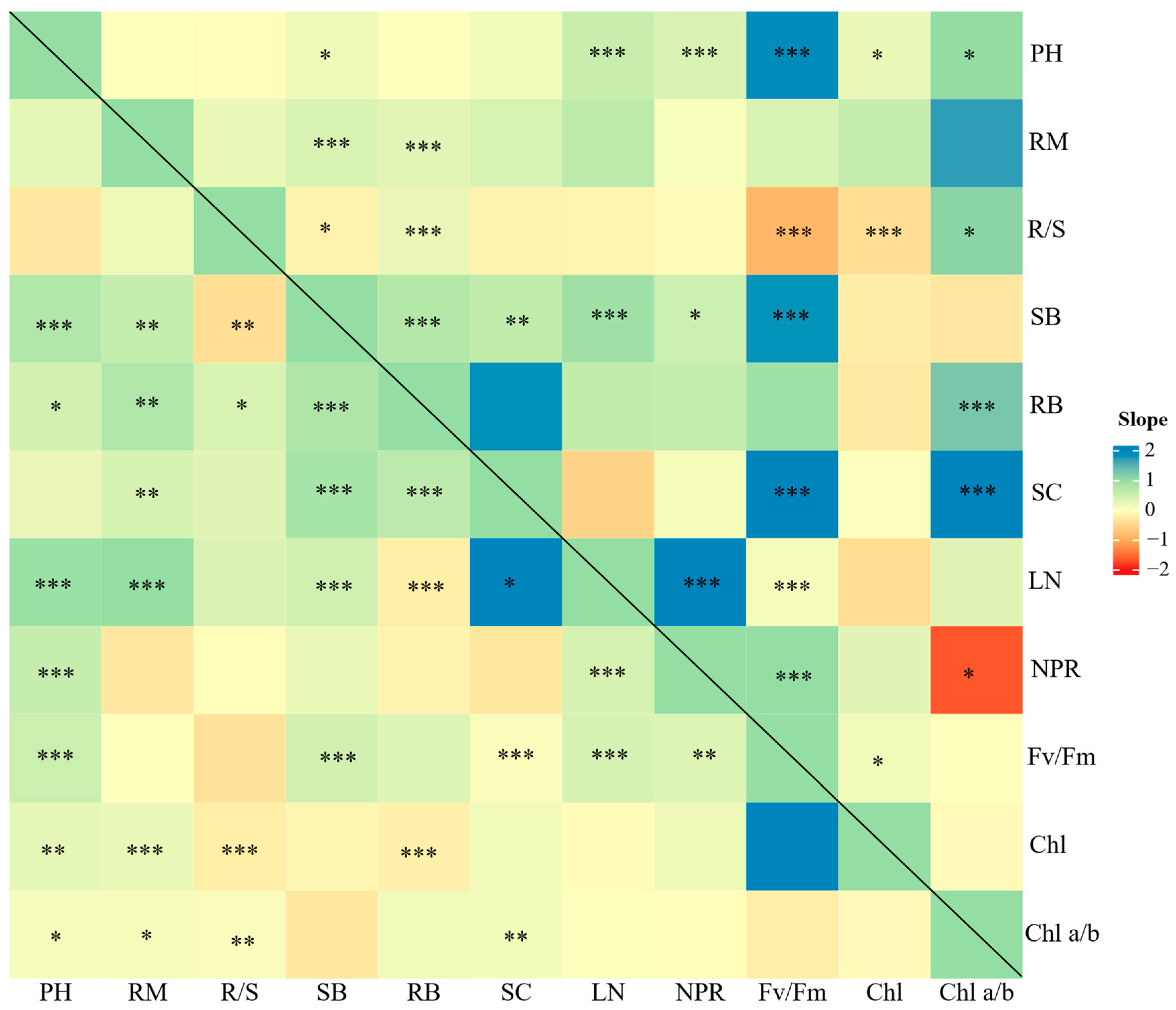
RGR was linked closely with whole-plant traits but non-significantly with leaf traits (Figure 3). Specifically, the lnRRs of RGR increased with the lnRRs of RM, SB, RB, and R/S. Some lnRRs of whole-plant traits correlated significantly with those of leaf traits (Figure 4 and Figure S7). For example, the lnRRs of SB were correlated positively and significantly with those of LN, NPR, and Fv/Fm. The lnRRs of PH were correlated significantly with those of LN, NPR, Fv/Fm, Chl, and Chl a/b. Furthermore, the lnRRs of RM and SC were also linked closely with those of SB and RB.
Based on these relationships, a potential regulatory pathway of the response of RGR to light reduction was proposed (Figure 5). With the increase in the percentage of light reduction (PLR), the lnRRs of RM, PH, and LN decreased. Subsequently, the lnRRs of NPR decreased due to the decreasing PH and LN. Finally, the increase in PLR resulted in SB, RB, and RGR decreasing significantly.
4. Discussion
4.1. Effects of Light Reduction on Growth-Related Whole Plant and Leaf Traits
Light is one of the major drivers (others include nutrients, temperature, and sediment type) that can affect plant growth and metabolism in aquatic ecosystems [15,57,58]. Our meta-analysis showed that light reduction significantly decreased the ramet number (RM), shoot biomass (SB), root biomass (RB), and soluble carbohydrate content (SC, Figure 2a). The decreased RM might be attributed to the adaptive strategy of submerged macrophytes. Submerged macrophytes tend to reduce the resistance of CO2 diffusion from water to chloroplasts and respiratory cost via decreasing RM [3,24,28,59], which might improve photosynthetic efficiency under low light stress. The decreased RM induced by light reduction further leads to decreased total carbon assimilation and significant reductions of SC, SB, and RB. Moreover, plant damage induced by light reduction is likely to affect the rhizosphere microbial community, resulting in the limited availability of nutrients and further decreases of SB and RB [60,61]. Specifically, light reduction could influence the rhizosphere microbes of submerged macrophytes via oxygen concentrations and root exudations [62,63,64]. Low light availability reduced the production of oxygen in the photosynthetic tissue of submerged macrophytes and decreased oxygen transportation from root tips into the rhizosphere [65,66], which might lead to a decreasing abundance of plant-growth-promoting bacteria (e.g., Azospirillum and Pseudomonas) and sulfide oxidizers (e.g., Sulfurimonas) [63,67,68]. Additionally, light deprivation could alter the compositions of root exudation resulting in a decrease in nitrogen-fixation bacteria and an increase in nirS denitrifying and anammox bacteria, which in turn intensifies the nitrogen limitation for the growth of submerged macrophytes [63,69]. However, how the type and amount of root exudation affect the rhizosphere nutrient cycles in submerged macrophytes should be further investigated.
Similarly, our results indicated that light reduction caused a significant decrease in leaf number (LN) but an increase in chlorophyll content (Chl) at the global scale (Figure 2b). When faced with limited light availability, the leaf increases Chl to adapt to low light stress and to increase low light utilization [32]. However, the increased Chl did not cause the increase in the net photosynthesis rate (NPR) in our meta-analysis, which might be the result of the consumption of more energy for nutrient metabolism activities (e.g., glutamine synthetase and alkaline phosphatase activity) [32]. Moreover, the light-reduction-induced decrease in LN (i.e., the photosynthetic organ) also contributed to the non-significant NPR.
4.2. Light-Reduction-Induced Changes of RGR and Its Relationships with Growth-Related Traits
Because suites of traits are linked to the growth of submerged macrophytes, the responses of growth-related traits to light reduction are closely connected with macrophytes’ eco-physiological adaptive strategies, resulting in changes in RGR [26,38,59]. Our meta-analysis indicated that light reduction significantly decreased the RGR (Figure 1). The growth rate decrease might be attributed to the following mechanisms. First, light reduction might decrease the photosynthetic capacity of whole plants. Our results showed that light reduction increased Chl by 25.8% but caused 56.1% and 59.2% decreases of LN and SB, respectively (Figure 2), resulting in decreased photosynthetic organ number and photosynthetic capacity of whole plants.
Second, submerged macrophytes might increase carbon investment in metabolism and thus decrease carbon fixation in response to light reduction [3,36]. For example, low light stress is likely to trigger the detoxification process, resulting in elevated antioxidant enzyme activity [70]. It has been shown that weak light stress could produce and accumulate superoxide dismutase and peroxidase activity in Hydrilla verticillate, Gracilaria lemaneiformis, and Vallisneria natans [71,72,73]. In our meta-analysis, NPR was affected non-significantly by light reduction, but the product of photosynthesis (i.e., SC) was reduced by light reduction (Figure 2), which directly suggested that more resources were invested in the metabolic and non-growth processes. Meanwhile, the responses (lnRRs) of RGR to light reduction were linked more closely with lnRRs of RB (R2 = 0.77) compared to photosynthetic organ biomasses (i.e., SB; R2 = 0.49) and positively linked with R/S (Figure 3b–d), indicating that the growth of submerged macrophytes relied more strongly on investment in non-photosynthetic organs in low-light environments.
Third, submerged macrophytes growing in poor light environments tended to decrease branch and lateral growth, leading to a decreased growth rate. Our meta-analysis also showed that the RM of submerged macrophytes was reduced by light reduction (Figure 2a), and the lnRRs of RGR increased significantly with those of RM (Figure 3). Among the three potential mechanisms, the decreased RGR was mainly driven by the decreased RM rather than leaf photosynthetic capability (Figure 5). On hand, the decreased RM reduced additional carbon investment in non-photosynthetic processes, contributing to the survival of submerged macrophytes under low light stress. On the other hand, light-reduction-induced changes in PH and LN showed opposite effects for NPR, resulting in a relatively weaker contribution of photosynthetic capability to lnRRs of RGR.
4.3. Factors Influencing the Responses of RGR and Other Traits to Light Reduction
Experimental factors (e.g., plant type, leaf shape, treatment method, experiment duration and percentage, and experimental end season) have been shown to affect the growth responses of plants to light reduction [3,30,74,75]. We found that the lnRRs of RGR were significantly negative for plants grown from shoots and seedlings but non-significantly positive for those grown from mature plants at the beginning of the experiment (Figure 1b), which might be related to more carbon storage in mature plants leading to greater adaptability. Meanwhile, some species dormant in low-temperature conditions might lead to non-significant effects of light reduction on RGR in spring, autumn, and winter.
The effect of light reduction on NPR for needle leaves was stronger than that for the flat leaves (Figure S4b), which might be due to flat leaves having a higher specific leaf area and being more sensitive to light changes compared to needle leaves [3]. Leaf originating from buds generally have high biological variability of photosynthesis pigments [76], leading to a stronger effect of light reduction on the chlorophyll (Chl) of plant materials originating from buds compared to those from shoots, seedlings, and mature plants (Figure S4d). Moreover, submerged macrophytes need more chlorophyll a content when they begin growing with more photosynthesis and a higher growth rate in spring, [31], leading to a stronger response of the Chl a/b ratio to light reduction in spring compared to other seasons (Figure S4e).
The lnRRs of the root-to-shoot ratio (R/S) increased with experimental duration (Figure S5), which might be attributed to the following reasons. When the duration was shorter than 70 days, to adapt to low light, submerged macrophytes elevated their investment in leaf construction to maximize photosynthetic capacity [77,78], leading to a negative response of R/S to light reduction. As the duration increased, weak light stress started to damage the leaves, and then the leaves withered, resulting in the R/S rising and the positive responses of R/S to light reduction.
4.4. Potential Publication Bias
Funnel plots, Egger’s test, and fail-safe analysis were used to test for potential publication bias in our database (Figures S8 and S9). There was no marked indication of publication bias for most trait datasets. Although the funnel plot for the LN was mildly asymmetrical, and the result of Egger’s test was significant (p < 0.001), the fail-safe analysis indicated that 3269 additional study cases with null results would need to be added to the dataset to reduce the significance level to p = 0.05. Therefore, the limited publication bias was not considered an issue for interpreting our main results.
4.5. Limitations and Implications
Due to the restriction of data coverage and the knowledge gap about functional traits of submerged macrophytes, our study may have a few limitations. First, we are aware that the responses of submerged macrophytes to light reduction were species-specific. Hence, our meta-analysis results might be inappropriate for predicting and modeling adaptive strategies for individual species. Second, most species in the database belonged to Vallisneria, Myriophyllum, Potamogeton, and Elodea (Table S1). The limited submerged species coverage of the field studies might hinder our understanding of growth strategies of the whole submerged macrophytes. Thus, more studies about growth strategies of submerged species, including other species (e.g., Stuckenia, Egeria, and Ceratophyllum), should be better explored in the future.
Despite these limitations, in this study, we found that light reduction significantly negatively influenced RGR, mainly resulting from the decreased RM. Our results may thus provide some suggestions for trait-based functional ecological research and the management and restoration of submerged macrophytes. We found that the growth responses of submerged macrophytes to light reduction were mainly driven by RM rather than leaf photosynthetic capability (Figure 3 and Figure 5). Thus, the tillering capacity of submerged macrophytes should be considered first when conducting ecological restoration and setting goals for the sustainable development of aquatic ecosystems.
Our results also showed that the responses of growth to light reduction had tipping points (at approximately 20% light reduction). Due to the restriction of data coverage, species-specific tipping points were still unclear. Nevertheless, based on the threshold effect of light reduction on RGR, the reported tipping point may help to improve hydrophyte dynamic models (HDMs). At present, the parameter used in most HDMs to reflect the effect of light on hydrophytes resulted from the light saturation point measured using photosynthetic light-response curves in the laboratory [79,80], which cannot represent plants’ actual growth and long-term light adaptation [81], thus limiting the predictions of the distribution and area of macrophytes. Recently, multiple environmental tipping points were successfully used in site selection for zebra mussel farming [82], suggesting that tipping points might also be applicable. The threshold effect of light reduction on RGR is essential for submerged species growth and distribution [16,17], which may help the management and restoration of submerged species [17]. Therefore, the more species-specific responses of RGR to light should be investigated, especially for common species in ecological restoration.
We also found a trend to covary with light reduction within the whole-plant (RM, SB, RB, and PH) and leaf traits (LN, NPR, Fv/Fm, and Chl) in our study (Figure 4 and Figure S7), which may imply a pattern to the plant economic spectrum existing in submerged plants. However, reports of functional traits related to resource acquisition (specific leaf area, nitrogen content, leaf longevity, enzyme activity, and root traits) are still scarce for submerged species compared to terrestrial plants [25,26], which limits the predictive capacity of these traits for community dynamics and ecosystem functions. Hence, there is an urgent need to examine the response of functional traits and their relationships to environmental changes to predict changes in submerged community structures and functions and to advance the sustainable development of aquatic ecosystems.
5. Conclusions
Growth-related traits play critical roles in the acclimation and adaptation of submerged macrophytes to light reduction. Our study examined the effect of light reduction on growth and related traits across 37 species from around the world. These results showed that light reduction markedly influenced growth traits and responses across the globe. Specifically, light reduction had significantly negative effects on RGR and whole-plant traits except for PH and R/S. At the leaf scale, light reduction induced a decrease in LN but an increase in Chl. The response of RGR to light reduction correlated significantly with whole-plant traits, including RM, SB, RB, and R/S, but non-significantly with leaf traits. The responses of RGR to light reduction were mainly driven by the RM rather than leaf photosynthetic capability. Moreover, most moderators had non-significant effects on the responses of traits, except for plant type and the percentage of light reduction. Our results highlighted the importance of these traits in response to light reduction, especially RGR and RM, and they might need to be incorporated into hydrophyte dynamic models for predicting the distribution and area of submerged macrophytes or to guide the management and ecological restoration of aquatic ecosystems.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/su15075918/s1: Figure S1: PRISMA flow chart showing the process of selecting publications; Figure S2: Geographical distribution of sampling sites of submerged plants in the study cases included in the meta-analysis; Figure S3: Meta-analysis of the effect of light reduction on the whole plant traits across different moderators; Figure S4: Meta-analysis of the effect of light reduction on the leaf traits across different moderators; Figure S5: Relationship of lnRRs of submerged macrophytes root shoot ratio with the experimental period (log scale) in the meta-analysis; Figure S6: Relationships of lnRRs of submerged macrophytes traits with the percentage of light reduction in the meta-analysis; Figure S7: Heatmap of the Pearson’s correlation coefficients between lnRRs of the paired photosynthesis and growth traits included in the meta-analysis; Figure S8: Contour-enhanced funnel plots for null-model residuals and p value of the Egger’s test for lnRRs of each variable; Figure S9: Bar plots of fail-safe numbers of each variable; and Table S1: List of primary studies, species and traits included in the meta-analysis. References [83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,129] are cited in the Supplementary Materials.
Author Contributions
J.G.: conceptualization, data collection and curation, formal analysis, visualization, writing—original draft, writing—review and editing; W.H.: funding acquisition, resources, writing—review and editing; J.W.: data collection and curation, writing—original; Y.C.: formal analysis, visualization; L.L.: formal analysis, visualization. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by funding from the China Three Gorges Corporation (Grand No.202103548) and Shanghai Investigation, Design & Research Institute Co., Ltd. (Grand No.2022QT(831)-002).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
The authors would like to acknowledge all scientists whose datasets and research results were included in the analysis.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Abbreviations
| Chl | Chlorophyll content |
| Chl a/b | Chlorophyll a/b ratio |
| Fv/Fm | Leaf maximal quantum yield of photosystem II complex |
| LN | Leaf number |
| lnRR | Response ratio |
| lnRR++ | Weighted average effect sizes |
| NPR | Net photosynthesis rate |
| PH | Plant height |
| PLR | Percentage of light reduction |
| RB | Root biomass |
| RGR | Relative growth rate |
| RM | Ramet number |
| R/S | Root-to-shoot ratio |
| SB | Shoot biomass |
| SC | Soluble carbohydrates |
References
- Jeppesen, E.; Søndergaard, M.; Søndergaard, M.; Christoffersen, K. The Structuring Role of Submerged Macrophytes in Lakes; Springer: New York, NY, USA, 1998; ISBN 978-0-387-98284-7. [Google Scholar]
- Su, H.; Chen, J.; Wu, Y.; Chen, J.; Guo, X.; Yan, Z.; Tian, D.; Fang, J.; Xie, P. Morphological traits of submerged macrophytes reveal specific positive feedbacks to water clarity in freshwater ecosystems. Sci. Total Environ. 2019, 684, 578–586. [Google Scholar] [CrossRef]
- Liu, H.; Liu, G.; Xing, W. Functional traits of submerged macrophytes in eutrophic shallow lakes affect their ecological functions. Sci. Total Environ. 2021, 760, 143332. [Google Scholar] [CrossRef]
- Henninger, T.O.; Froneman, P.W.; Richoux, N.B.; Hodgson, A.N. The role of macrophytes as a refuge and food source for the estuarine isopod Exosphaeroma hylocoetes (Barnard, 1940). Estuar. Coast. Shelf Sci. 2009, 82, 285–293. [Google Scholar] [CrossRef]
- Sato, M.; Nishijima, S.; Miyashita, T. Differences in refuge function for prey and tolerance to crayfish among macrophyte species. Limnology 2014, 15, 27–35. [Google Scholar] [CrossRef]
- Korner, S.; Nicklisch, A. Allelopathic growth inhibition of selected phytoplankton species by submerged macrophytes. J. Phycol. 2002, 38, 862–871. [Google Scholar] [CrossRef]
- Jiang, M.; Zhou, Y.; Wang, N.; Xu, L.; Zheng, Z.; Zhang, J. Allelopathic effects of harmful algal extracts and exudates on biofilms on leaves of Vallisneria natans. Sci. Total Environ. 2019, 655, 823–830. [Google Scholar] [CrossRef]
- Verhofstad, M.; Poelen, M.D.M.; van Kempen, M.M.L.; Bakker, E.S.; Smolders, A.J.P. Finding the harvesting frequency to maximize nutrient removal in a constructed wetland dominated by submerged aquatic plants. Ecol. Eng. 2017, 106, 423–430. [Google Scholar] [CrossRef]
- Carr, J.; D’Odorico, P.; McGlathery, K.; Wiberg, P. Stability and bistability of seagrass ecosystems in shallow coastal lagoons: Role of feedbacks with sediment resuspension and light attenuation. J. Geophys. Res-Biogeo. 2010, 115, 1–14. [Google Scholar] [CrossRef]
- Phillips, G.; Willby, N.; Moss, B. Submerged macrophyte decline in shallow lakes: What have we learnt in the last forty years? Aquat. Bot. 2016, 135, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Jeppesen, E.; Liu, X.; Qin, B.; Shi, K.; Zhou, Y.; Thomaz, S.M.; Deng, J. Global loss of aquatic vegetation in lakes. Earth-Sci. Rev. 2017, 173, 259–265. [Google Scholar] [CrossRef]
- Zhang, Q.; Dong, X.; Yang, X.; Liu, E.; Lin, Q.; Cheng, L.; Liu, L.; Jeppesen, E. Aquatic macrophyte fluctuations since the 1900s in the third largest Chinese freshwater lake (Lake Taihu): Evidences, drivers and management implications. Catena 2022, 213, 106153. [Google Scholar] [CrossRef]
- Lacoul, P.; Freedman, B. Environmental influences on aquatic plants in freshwater ecosystems. Environ. Rev. 2006, 14, 89–136. [Google Scholar] [CrossRef]
- Son, D.; Cho, H.; Lee, E.J. Determining factors for the occurrence and richness of submerged macrophytes in major Korean rivers. Aquat. Bot. 2018, 150, 82–88. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, X.; Qin, B.; Shi, K.; Deng, J.; Zhou, Y. Aquatic vegetation in response to increased eutrophication and degraded light climate in Eastern Lake Taihu: Implications for lake ecological restoration. Sci. Rep. 2016, 6, 23867. [Google Scholar] [CrossRef]
- Yang, C.; Shi, X.; Nan, J.; Huang, Q.; Shen, X.; Li, J. Morphological responses of the submerged macrophyte Vallisneria natans along an underwater light gradient: A mesocosm experiment reveals the importance of the Secchi depth to water depth ratio. Sci. Total Environ. 2022, 808, 152199. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Shi, K.; Lin, J.; Zhou, Y.; Qin, B. Determining critical light and hydrologic conditions for macrophyte presence in a large shallow lake: The ratio of euphotic depth to water depth. Ecol. Indic. 2016, 71, 317–326. [Google Scholar] [CrossRef]
- Ni, L. Growth of Potamageton maackianus under low-light stress in eutrophic water. J. Freshw. Ecol. 2001, 16, 249–256. [Google Scholar] [CrossRef] [Green Version]
- Sorrell, B.K. Regulation of root anaerobiosis and carbon translocation by light and root aeration in Isoetes alpinus. Plant Cell Environ. 2004, 27, 1102–1111. [Google Scholar] [CrossRef]
- Xie, Y.; Luo, W.; Ren, B.; Li, F. Morphological and physiological responses to sediment type and light availability in roots of the submerged plant Myriophyllum spicatum. Ann. Bot. 2007, 100, 1517–1523. [Google Scholar] [CrossRef] [Green Version]
- Wright, I.J.; Westoby, M.; Reich, P.B. Convergence towards higher leaf mass per area in dry and nutrient-poor habitats has different consequences for leaf life span. J. Ecol. 2002, 90, 534–543. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Wright, I.J.; Dong, N.; Maire, V.; Prentice, I.C.; Westoby, M.; Díaz, S.; Gallagher, R.V.; Jacobs, B.F.; Kooyman, R.; Law, E.A.; et al. Global climatic drivers of leaf size. Science 2017, 357, 917–921. [Google Scholar] [CrossRef] [Green Version]
- Poorter, H.; Niinemets, U.; Poorter, L.; Wright, I.J.; Villar, R. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef]
- Pierce, S.; Brusa, G.; Sartori, M.; Cerabolini, B.E.L. Combined use of leaf size and economics traits allows direct comparison of hydrophyte and terrestrial herbaceous adaptive strategies. Ann. Bot. 2012, 109, 1047–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalla Vecchia, A.; Villa, P.; Bolpagni, R. Functional traits in macrophyte studies: Current trends and future research agenda. Aquat. Bot. 2020, 167, 103290. [Google Scholar] [CrossRef]
- Strand, J.A.; Weisner, S.E.B. Morphological plastic responses to water depth and wave exposure in an aquatic plant (Myriophyllum spicatum). J. Ecol. 2001, 89, 166–175. [Google Scholar] [CrossRef]
- Riis, T.; Olesen, B.; Clayton, J.S.; Lambertini, C.; Brix, H.; Sorrell, B.K. Growth and morphology in relation to temperature and light availability during the establishment of three invasive aquatic plant species. Aquat. Bot. 2012, 102, 56–64. [Google Scholar] [CrossRef]
- Chen, J.; Cao, T.; Zhang, X.; Xi, Y.; Ni, L.; Jeppesen, E. Differential photosynthetic and morphological adaptations to low light affect depth distribution of two submersed macrophytes in lakes. Sci. Rep. 2016, 6, 34028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Ding, M.; Lan, Z.; Zhao, Y.; Chen, J. Light Availability and Patterns of Allocation to Reproductive and Vegetative Biomass in the Sexes of the Dioecious Macrophyte Vallisneria spinulosa. Front. Plant Sci. 2019, 10, 572. [Google Scholar] [CrossRef] [Green Version]
- Arthaud, F.; Toury, J.; Romestaing, C.; Bornette, G. Photosynthetic and morphological traits control aquatic plant distribution according to light stress. Evol. Ecol. 2021, 35, 739–760. [Google Scholar] [CrossRef]
- Li, H.; Li, Q.; Luo, X.; Fu, J.; Zhang, J. Responses of the submerged macrophyte Vallisneria natans to a water depth gradient. Sci. Total Environ. 2020, 701, 134944. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Li, H.; Luo, F. Effects of light heterogeneity on growth of a submerged clonal macrophyte. Plant Spec. Biol. 2013, 28, 156–164. [Google Scholar] [CrossRef]
- Yu, Y.; Li, Y.; Wang, H.; Wu, X.; Zhang, M.; Wang, H.; Hamilton, D.P.; Jeppesen, E. Submersed macrophyte restoration with artificial light-emitting diodes: A mesocosm experiment. Ecotoxicol. Environ. Saf. 2021, 228, 113044. [Google Scholar] [CrossRef]
- Chen, J.; Chou, Q.; Ren, W.; Su, H.; Zhang, M.; Cao, T.; Zhu, T.; Ni, L.; Liu, Z.; Xie, P. Growth, morphology and C/N metabolism responses of a model submersed macrophyte, Vallisneria natans, to various light regimes. Ecol. Indic. 2022, 136, 108652. [Google Scholar] [CrossRef]
- Zhu, B.; Mayer, C.M.; Rudstam, L.G.; Mills, E.L.; Ritchie, M.E. A comparison of irradiance and phosphorus effects on the growth of three submerged macrophytes. Aquat. Bot. 2008, 88, 358–362. [Google Scholar] [CrossRef]
- Chotikarn, P.; Kaewchana, P.; Prathep, A.; Roekngandee, P.; Sinutok, S. Effect of in situ experiment shading on the photosynthesis of Canadian waterweed (Elodea canadensis) from Songkhla Lagoon, Thailand. Appl. Ecol. Environ. Res. 2021, 19, 2593–2604. [Google Scholar] [CrossRef]
- Lu, J.; Wang, Z.; Xing, W.; Liu, G. Effects of substrate and shading on the growth of two submerged macrophytes. Hydrobiologia 2013, 700, 157–167. [Google Scholar] [CrossRef]
- Hussner, A.; Hofstra, D.; Jahns, P.; Clayton, J. Response capacity to CO2 depletion rather than temperature and light effects explain the growth success of three alien Hydrocharitaceae compared with native Myriophyllum triphyllum in New Zealand. Aquat. Bot. 2015, 120, 205–211. [Google Scholar] [CrossRef]
- Peng, H.; Ge, D.; Yuan, G.; Zou, D.; Fu, H.; Jeppesen, E. Effect of clonal fragmentation on the growth of Vallisneria natans (Lour.) Hara at contrasting nutrient and light conditions. Hydrobiologia 2021, 848, 903–912. [Google Scholar] [CrossRef]
- Gurevitch, J.; Koricheva, J.; Nakagawa, S.; Stewart, G. Meta-analysis and the science of research synthesis. Nature 2018, 555, 175–182. [Google Scholar] [CrossRef]
- Hozo, S.P.; Djulbegovic, B.; Hozo, I. Estimating the mean and variance from the median, range, and the size of a sample. BMC Med. Res. Methodol. 2005, 5, 13. [Google Scholar] [CrossRef] [Green Version]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: http://www.r-project.org/ (accessed on 13 April 2022).
- Hedges, L.V.; Gurevitch, J.; Curtis, P.S. The Meta-Analysis of Response Ratios in Experimental Ecology. Ecology 1999, 80, 1150–1156. [Google Scholar] [CrossRef]
- Gleser, L.J.; Olkin, I. Stochastically dependent effect sizes. In The Handbook of Research Synthesis and Meta-Analysis, 2nd ed.; Russell Sage Foundation: New York, NY, USA, 2009; pp. 357–376. ISBN 978-0-87154-163-5. [Google Scholar]
- Lajeunesse, M.J. On the meta-analysis of response ratios for studies with correlated and multi-group designs. Ecology 2011, 92, 2049–2055. [Google Scholar] [CrossRef] [PubMed]
- Bracken, M.B. Statistical methods for analysis of effects of treatment in overviews of randomized trials. In Effective Care of the Newborn Infant; Sinclair, J.C., Bracken, M.B., Eds.; Oxford University Press: Oxford, UK, 1992; pp. 13–20. ISBN 978-0192617378. [Google Scholar]
- Lajeunesse, M.J. Facilitating systematic reviews, data extraction and meta-analysis with the metagear package for r. Methods Ecol. Evol. 2016, 7, 323–330. [Google Scholar] [CrossRef]
- Kambach, S.; Bruelheide, H.; Gerstner, K.; Gurevitch, J.; Beckmann, M.; Seppelt, R. Consequences of multiple imputation of missing standard deviations and sample sizes in meta-analysis. Ecol. Evol. 2020, 10, 11699–11712. [Google Scholar] [CrossRef]
- Viechtbauer, W. Conducting Meta-Analyses in R with the metafor Package. J. Stat. Soft. 2010, 36, 1–48. [Google Scholar] [CrossRef] [Green Version]
- Benítez-López, A.; Alkemade, R.; Schipper, A.; Ingram, D.; Verweij, P.A.; Eikelboom, J.; Huijbregts, M. The impact of hunting on tropical mammal and bird populations. Science 2017, 356, 180–183. [Google Scholar] [CrossRef] [Green Version]
- Midolo, G.; De Frenne, P.; Hölzel, N.; Wellstein, C. Global patterns of intraspecific leaf trait responses to elevation. Glob. Chang. Biol. 2019, 25, 2485–2498. [Google Scholar] [CrossRef]
- Rosenberg, M.S. The file-drawer Problem Revisited: A General Weighted Method for Calculating fail-safe Numbers in meta-analysis. Evolution 2005, 59, 464–468. [Google Scholar] [CrossRef]
- Nakagawa, S.; Santos, E.S.A. Methodological issues and advances in biological meta-analysis. Evol. Ecol. 2012, 26, 1253–1274. [Google Scholar] [CrossRef]
- Revelle, W. Psych: Procedures for Personality and Psychological Research, 1.9.12; Northwestern University: Evanston, IL, USA, 2019; Available online: https://CRAN.R-project.org/package=psych (accessed on 13 April 2022).
- Wei, T.; Simko, V. R Package ‘Corrplot’: Visualization of a Correlation Matrix, 0.92. 2021. Available online: https://github.com/taiyun/corrplot (accessed on 13 April 2022).
- Chmara, R.; Szmeja, J.; Robionek, A. Leaf traits of macrophytes in lakes: Interspecific, plant group and community patterns. Limnologica 2019, 77, 125691. [Google Scholar] [CrossRef]
- Liu, H.; Zhou, W.; Li, X.; Chu, Q.; Tang, N.; Shu, B.; Liu, G.; Xing, W. How many submerged macrophyte species are needed to improve water clarity and quality in Yangtze floodplain lakes? Sci. Total Environ. 2020, 724, 138267. [Google Scholar] [CrossRef] [PubMed]
- Ellawala Kankanamge, C.; Matheson, F.E.; Riis, T. Shading constrains the growth of invasive submerged macrophytes in streams. Aquat. Bot. 2019, 158, 103125. [Google Scholar] [CrossRef]
- Kurtz, J.C.; Yates, D.F.; Macauley, J.M.; Quarles, R.L.; Genthner, F.J.; Chancy, C.A.; Devereux, R. Effects of light reduction on growth of the submerged macrophyte Vallisneria americana and the community of root-associated heterotrophic bacteria. J. Exp. Mar. Bio. Eco. 2003, 291, 199–218. [Google Scholar] [CrossRef]
- George, R.; Gullström, M.; Mangora, M.M.; Mtolera, M.S.P.; Björk, M. High midday temperature stress has stronger effects on biomass than on photosynthesis: A mesocosm experiment on four tropical seagrass species. Ecol. Evol. 2018, 8, 4508–4517. [Google Scholar] [CrossRef]
- Soda, S.; Ike, M.; Ogasawara, Y.; Yoshinaka, M.; Mishima, D.; Fujita, M. Effects of light intensity and water temperature on oxygen release from roots into water lettuce rhizosphere. Water Res. 2007, 41, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Martin, B.C.; Gleeson, D.; Statton, J.; Siebers, A.R.; Grierson, P.; Ryan, M.H.; Kendrick, G.A. Low Light Availability Alters Root Exudation and Reduces Putative Beneficial Microorganisms in Seagrass Roots. Front. Microbiol. 2018, 8, 2667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, H.; Jiang, M.; Zhou, N.; Jiang, C.; Liu, S. Submerged macrophytes recruit unique microbial communities and drive functional zonation in an aquatic system. Appl. Microbiol. Biotechnol. 2021, 105, 7517–7528. [Google Scholar] [CrossRef]
- Brodersen, K.E.; Nielsen, D.A.; Ralph, P.J.; Kühl, M. A split flow chamber with artificial sediment to examine the below-ground microenvironment of aquatic macrophytes. Mar. Biol. 2014, 161, 2921–2930. [Google Scholar] [CrossRef]
- Jovanovic, Z.; Pedersen, M.; Larsen, M.; Kristensen, E.; Glud, R.N. Rhizosphere O2 dynamics in young Zostera marina and Ruppia maritima. Mar. Ecol. Prog. Ser. 2015, 518, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Bagwell, C.E.; La Rocque, J.R.; Smith, G.W.; Polson, S.W.; Friez, M.J.; Longshore, J.W.; Lovell, C.R. Molecular diversity of diazotrophs in oligotrophic tropical seagrass bed communities. FEMS Microbiol. Ecol. 2002, 39, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Perner, M. The globally widespread genus Sulfurimonas: Versatile energy metabolisms and adaptations to redox clines. Front. Microbiol. 2015, 6, 989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Yin, X.; Chen, L.; Wang, J.; Wang, Y.; Liu, G.; Hua, Y.; Wan, X.; Xiao, N.; Zhao, J.; et al. Effects of illumination on nirS denitrifying and anammox bacteria in the rhizosphere of submerged macrophytes. Sci. Total Environ. 2021, 760, 143420. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Gu, X.; Song, R.; Wang, X.; Yang, L. Microcystin-LR induced oxidative stress and ultrastructural alterations in mesophyll cells of submerged macrophyte Vallisneria natans (Lour.) Hara. J. Hazard. Mater. 2011, 190, 188–196. [Google Scholar] [CrossRef]
- Xue, W.; Peng, Y.; Song, X.; Zou, G. Effects of light intensity on physiological and biochemical characteristics of Hydrilla verticillata (L.f.) Royle. J. Anhui Agr. Sci. 2012, 40, 4169–4172. [Google Scholar] [CrossRef]
- Wu, H.; Jiang, H.; Liu, C.; Deng, Y. Growth, pigment composition, chlorophyll fluorescence and antioxidant defenses in the red alga Gracilaria lemaneiformis (Gracilariales, Rhodophyta) under light stress. S. Afr. J. Bot. 2015, 100, 27–32. [Google Scholar] [CrossRef]
- Zhu, Z.; Song, S.; Yan, Y.; Li, P.; Jeelani, N.; Wang, P.; An, S.; Leng, X. Combined effects of light reduction and ammonia nitrogen enrichment on the submerged macrophyte Vallisneria natans. Mar. Freshw. Res. 2018, 69, 764–770. [Google Scholar] [CrossRef]
- Fritz, C.; Schneider, T.; Geist, J. Seasonal Variation in Spectral Response of Submerged Aquatic Macrophytes: A Case Study at Lake Starnberg (Germany). Water 2017, 9, 527. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.; Ibrahim, M.; Muhammad, S.; Khan, S.; Li, G. Light intensity effects on the growth and biomass production of submerged macrophytes in different water strata. Arab. J. Geosci. 2020, 13, 948. [Google Scholar] [CrossRef]
- Solymosi, K.; Morandi, D.; Boka, K.; Boddi, B.; Schoefs, B. High biological variability of plastids, photosynthetic pigments and pigment forms of leaf primordia in buds. Planta 2012, 235, 1035–1049. [Google Scholar] [CrossRef]
- Weiner, J. Allocation, plasticity and allometry in plants. Perspect. Plant Ecol. Evol. Syst. 2004, 6, 207–215. [Google Scholar] [CrossRef]
- Choudhury, M.I.; Urrutia-Cordero, P.; Zhang, H.; Ekvall, M.K.; Medeiros, L.R.; Hansson, L.A. Charophytes collapse beyond a critical warming and brownification threshold in shallow lake systems. Sci. Total Environ. 2019, 661, 148–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, H.; Shi, Q.; Qian, X. A multi-species modelling approach to select appropriate submerged macrophyte species for ecological restoration in Gonghu Bay, Lake Taihu, China. Ecol. Model. 2017, 360, 179–188. [Google Scholar] [CrossRef]
- Ji, Z. Hydrodynamics and Water Quality: Modeling Rivers, Lakes, and Estuaries; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2017; pp. 285–306. ISBN 9781118877159. [Google Scholar]
- Sviben, S.; Matoničkin Kepčija, R.; Vidaković-Cifrek, Ž.; Sertić Perić, M.; Kružić, P.; Popijač, A.; Primc, B. Chara spp. exhibit highly heterogeneous light adaptation, calcite encrustation and epiphyton patterns in a marl lake. Aquat. Bot. 2018, 147, 1–10. [Google Scholar] [CrossRef]
- Bagdanavičiūtė, I.; Umgiesser, G.; Vaičiūtė, D.; Bresciani, M.; Kozlov, I.; Zaiko, A. GIS-based multi-criteria site selection for zebra mussel cultivation: Addressing end-of-pipe remediation of a eutrophic coastal lagoon ecosystem. Sci. Total Environ. 2018, 634, 990–1003. [Google Scholar] [CrossRef]
- Carter, V.; Rybicki, N.B.; Turtora, M. Effect of increasing photon irradiance on the growth of Vallisneria americana in the tidal Potomac River. Aquat. Bot. 1996, 54, 337–345. [Google Scholar] [CrossRef]
- Asaeda, T.; Sultana, M.; Manatunge, J.; Fujino, T. The effect of epiphytic algae on the growth and production of Potamogeton perfoliatus L. in two light conditions. Environ. Exp. Bot. 2004, 52, 225–238. [Google Scholar] [CrossRef]
- Boedeltje, G.; Smolders, A.J.P.; Roelofs, J.G.M. Combined effects of water column nitrate enrichment, sediment type and irradiance on growth and foliar nutrient concentrations of Potamogeton alpinus. Freshw. Biol. 2005, 50, 1537–1547. [Google Scholar] [CrossRef]
- Li, Y.; Yu, D.; Xu, X.; Xie, Y. Light intensity increases the susceptibility of Vallisneria natans to snail herbivory. Aquat. Bot. 2005, 81, 265–275. [Google Scholar] [CrossRef]
- Chen, X.; Chen, K.; Xiao, Y.; Zhang, S.; Wang, Q. Effects of light and matrix on turion germination, seedling growth and leaf photosynthesis efficiency of Potamogeton crispus. Chin. J. Appl. Ecol. 2006, 17, 1413–1418, (Chinese article with English abstract). [Google Scholar]
- Li, H. The Influences of Light, Nutrition and Phytoplankton on Growth and Physiology of Submerged Macrophytes in Eutrophic Waters; Graduate University of the Chinese Academy of Sciences: Beijing, China, 2006. [Google Scholar]
- Xiao, Y. Effect of Main Environmental Factors on Photosynthesis of Three Submerged Macrophytes from Taihu Lake; Nanjing Agriculture University: Nanjing, China, 2006; (Chinese thesis with English abstract). [Google Scholar]
- Xiao, Y.; Chen, K.; Dai, X.; Chen, X.; Xu, X. Comparison of Adaptive Capacity to Low Light Intensity of Two Angiosperm Submerged Macrophytes from Taihu Lake. Plant Physiol. Commun. 2006, 42, 421–425, (Chinese article with English abstract). [Google Scholar]
- Imamoto, H.; Horiya, K.; Yamasaki, M.; Washitani, I. An experimental system to study ecophysiological responses of submerged macrophytes to temperature and light. Ecol. Res. 2007, 22, 172–176. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Q.; Chen, K. Impacts of Different Light Intensity on Morphology and Structure of Potamogeton crispus. J. Wuhan Bot. Res. 2008, 26, 163–169, (Chinese article with English abstract). [Google Scholar]
- Zhou, X.; Wang, G.; Feng, B. Influence of the growth and photosynthetic characteristics of Potamogeton cripus in light. Ecol. Environ. 2008, 17, 1342–1347, (Chinese article with English abstract). [Google Scholar]
- Zhou, X.; Wang, G.; Feng, B. Effects of Light on Growth and Some Characteristics of Light-Energy Conversion of Elodea nuttallii Seedling. J. Ecol. Rural Environ. 2008, 24, 46–52, (Chinese article with English abstract). [Google Scholar]
- Angelstein, S.; Schubert, H. Light acclimatisation of Elodea nuttallii grown under ambient DIC conditions. Plant Ecol. 2009, 202, 91–101. [Google Scholar] [CrossRef]
- Cao, T.; Xie, P.; Ni, L.; Zhang, M.; Xu, J. Carbon and nitrogen metabolism of an eutrophication tolerative macrophyte, Potamogeton crispus, under NH4+ stress and low light availability. Environ. Exp. Bot. 2009, 66, 74–78. [Google Scholar] [CrossRef]
- Zhu, G. Effects of Turbidity and Low Light Intensity on the Growth of Macrophytes; Nanjing Foresty University: Nanjing, China, 2009; (Chinese thesis with English abstract). [Google Scholar]
- Sultana, M.; Asaeda, T.; Azim, M.E.; Fujino, T. Morphological plasticity of submerged macrophyte Potamogeton wrightii Morong under different photoperiods and nutrient conditions. Chem. Ecol. 2010, 26, 223–232. [Google Scholar] [CrossRef]
- Wang, S. The Combined Effect of Light and N, P on Growth and Physiology of Vallisneria natans; South China University of Technology: Guangzhou, China, 2010; (Chinese thesis with English abstract). [Google Scholar]
- Zhang, M.; Cao, T.; Ni, L.; Xie, P.; Li, Z. Carbon, nitrogen and antioxidant enzyme responses of Potamogeton crispus to both low light and high nutrient stresses. Environ. Exp. Bot. 2010, 68, 44–50. [Google Scholar] [CrossRef]
- Cao, T.; Ni, L.; Xie, P.; Xu, J.; Zhang, M. Effects of moderate ammonium enrichment on three submersed macrophytes under contrasting light availability. Freshw. Biol. 2011, 56, 1620–1629. [Google Scholar] [CrossRef]
- Zhu, D.; Qiao, N.; Li, M.; Chen, P. Effect of light intensity, temperature, total nitrogen concentration and their interaction on Hydrilla Verticillata. Acta Hydrobiol. Sin. 2011, 35, 88–97, (Chinese article with English abstract). [Google Scholar] [CrossRef]
- Ge, F.; Liu, B.; Lu, Z.; Gao, Y.; Wu, Z. Effects of Light Intensity on Growth and Phenolic Contents of Myriophyllum spicatum. Environ. Sci. Technol. 2012, 35, 30–34, (Chinese article with English abstract). [Google Scholar]
- Yuan, L.; Li, W.; Liu, G.; Deng, G. Effects of different shaded conditions and water depths on the growth and reproductive strategy of Vallisneria spinulosa. Pak. J. Bot. 2012, 44, 911–918. [Google Scholar]
- Malheiro, A.C.E.; Jahns, P.; Hussner, A. CO2 availability rather than light and temperature determines growth and phenotypical responses in submerged Myriophyllum aquaticum. Aquat. Bot. 2013, 110, 31–37. [Google Scholar] [CrossRef]
- Cao, J.; Lu, J.; Ruan, H. Effects of underwater illumination compensation on growth and physiological indices of submerged macrophyte Vallisneria natans L. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2014, 38, 102–106, (Chinese article with English abstract). [Google Scholar]
- Lang, P.; Yuan, L.; Li, S. Effects of different light conditions on the photosynthesis of Potamogeton malaianus and Potamogeon pectinatus. Jiangsu Agric. Sci. 2014, 42, 323–325, (Chinese article with English abstract). [Google Scholar]
- Zhao, S.; Li, E.; Yang, J.; Wang, Z.; Wang, X.; Cai, X. Effects of light intensity on the seeds germination, seedlings growth and physiology of Ottelia acuminata. J. Lake Sci. 2014, 26, 107–112, (Chinese article with English abstract). [Google Scholar]
- Eller, F.; Alnoee, A.B.; Boderskov, T.; Guo, W.-Y.; Kamp, A.T.; Sorrell, B.K.; Brix, H. Invasive submerged freshwater macrophytes are more plastic in their response to light intensity than to the availability of free CO2 in air-equilibrated water. Freshw. Biol. 2015, 60, 929–943. [Google Scholar] [CrossRef]
- Hussner, A.; Jahns, P. European native Myriophyllum spicatum showed a higher use capacity than alien invasive Myriophyllum heterophyllum. Hydrobiologia 2015, 746, 171–182. [Google Scholar] [CrossRef]
- Song, Y.; Kong, F.; Wang, M.; Zhang, Y. Effects of Light Intensity and Epiphytic Algae on Physiological Parameters of Myriophyllum spicatum. J. Agro-Environ. Sci. 2015, 34, 233–239, (Chinese article with English abstract). [Google Scholar]
- Lin, C.; Han, C.; Pan, H.; You, W.; Yan, H.; Chen, H. Effects of different light intensity on growth of the eight submerged plants. Environ. Eng. 2016, 34, 16–19, (Chinese article with English abstract). [Google Scholar]
- Zhang, S. The influecne of Vallisneria natans Growth under Water Gradient/Light Intensity; Jiangxi Normal University: Nanchang, China, 2016; (Chinese thesis with English abstract). [Google Scholar]
- Cao, Y.; Luo, S.; Chen, B. Effects of light intensity on growth and antioxidant enzyme activity of Potamogeton crispus. Acta Hydrobiol. Sin. 2018, 42, 846–853, (Chinese article with English abstract). [Google Scholar]
- Jiang, H.; Zhang, Y.; Yin, L.; Li, W.; Jin, Q.; Fu, W.; Zhang, T.; Huang, W. Diurnal changes in photosynthesis by six submerged macrophytes measured using fluorescence. Aquat. Bot. 2018, 149, 33–39. [Google Scholar] [CrossRef]
- Li, P.; Zhu, Z.; Yan, Y.; An, S.; Leng, X. Effects of different light intensity and sediment nutrition on three submerged macrophytes. Ecol. Sci. 2018, 37, 101–107, (Chinese article with English abstract). [Google Scholar]
- Xue, R. Effect of Light Conditions on Growth Characteristic of Two Submersed Macrophyte; Chongqing University: Chongqing, China, 2018; (Chinese thesis with English abstract). [Google Scholar]
- He, L.; Bakker, E.S.; Nunez, M.M.A.; Hilt, S. Combined effects of shading and clipping on the invasive alien macrophyte Elodea nuttallii. Aquat. Bot. 2019, 154, 24–27. [Google Scholar] [CrossRef]
- Hillmann, E.R.; La Peyre, M.K. Effects of salinity and light on growth and interspecific interactions between Myriophyllum spicatum L. and Ruppia maritima L. Aquat. Bot. 2019, 155, 25–31. [Google Scholar] [CrossRef]
- Hu, Q.; Turnbull, M.; Hawes, I. Estimated light compensation depth explains growth of Stuckenia pectinata in Te Waihora. Aquat. Bot. 2019, 156, 57–64. [Google Scholar] [CrossRef]
- Tan, X.; Yuan, G.; Fu, H.; Peng, H.; Ge, D.; Lou, Q.; Zhong, J. Effects of ammonium pulse on the growth of three submerged macrophytes. PLoS ONE 2019, 14, e0219161. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X. Study on the Influence of Environmental on the Growth of Submerged Rotala Rotundifolia; Wenzhou University: Wenzhou, China, 2019; (Chinese thesis with English abstract). [Google Scholar]
- Chen, J.; Liu, Z.; Xiao, S.; Chen, R.; Luo, C.; Zhu, T.; Cao, T.; Ni, L.; Xie, P.; Su, H.; et al. Effects of benthivorous fish disturbance on chlorophyll a contents in water and the growth of two submersed macrophytes with different growth forms under two light regimes. Sci. Total Environ. 2020, 704, 135269. [Google Scholar] [CrossRef]
- Chen, J.; Ren, W.; Chou, Q.; Su, H.; Ni, L.; Zhang, M.; Liu, Z.; Xie, P. Alterations in biomass allocation indicate the adaptation of submersed macrophytes to low-light stress. Ecol. Indic. 2020, 113, 106235. [Google Scholar] [CrossRef]
- Kankanamge, C.E.; Matheson, F.E.; Riis, T. Shading may alter the colonization pattern and dominance between two invasive submerged aquatic plant species. Aquat. Ecol. 2020, 54, 721–728. [Google Scholar] [CrossRef]
- Yuan, G.; Fu, H.; Zhang, M.; Lou, Q.; Dai, T.; Jeppesen, E. Effects of plant size on the growth of the submersed macrophyte Vallisneria spinulosa S.Z.Yan at different light intensities: Implications for lake restoration. Hydrobiologia 2020, 847, 3609–3619. [Google Scholar] [CrossRef]
- Malea, L.; Nakou, K.; Papadimitriou, A.; Exadactylos, A.; Orfanidis, S. Physiological Responses of the Submerged Macrophyte Stuckenia pectinata to High Salinity and Irradiance Stress to Assess Eutrophication Management and Climatic Effects: An Integrative Approach. Water 2021, 13, 1706. [Google Scholar] [CrossRef]
- Yuan, G.; Yang, Z.; Sun, L.; Fu, H.; Peng, H.; Jeppesen, E. Asexual reproduction for overwintering of the submersed macrophyte Vallisneria spinulosa at different light intensities. Aquat. Sci. 2022, 84, 13. [Google Scholar] [CrossRef]
- Yuan, J.; Bai, Z.; Ye, S.; Liu, H.; Wang, Y.; Li, F.; Xie, Y.; Gao, A.; Wu, A. High-light inhibition of two submerged macrophytes in a shallow water experiment. AoB Plants 2022, 14, plac009. [Google Scholar] [CrossRef]
Figure 1.
Orchard plot (a) showing the mean estimate (red dot), confidence interval (bold black line), individual effect sizes (blue bubbles), and their precision (inverse variance) for the response ratios (lnRRs) of relative growth rate (RGR). A positive value of lnRR indicates that the mean value of a specific trait for a given species increases relative to that at the highest light level. Forest plot (b) showing the effect of light reduction on RGR across different moderators. Overall effect and 95% confidence intervals are given. Where the zero line (red solid line) is not crossed by the confidence intervals, the effect of light reduction was significant (shown by the asterisks, * p < 0.05, ** p < 0.01, *** p < 0.001). Arrows indicate 95% confidence intervals extending beyond the limits of the plot. Values in parentheses are sample sizes. The effects of moderators are tested by QM at the 0.05 level. Scatter plot (c) denoting the relationships of the lnRRs of RGR with the percentage of light reduction. The sizes of dots were weighted by the inverse variance.
Figure 1.
Orchard plot (a) showing the mean estimate (red dot), confidence interval (bold black line), individual effect sizes (blue bubbles), and their precision (inverse variance) for the response ratios (lnRRs) of relative growth rate (RGR). A positive value of lnRR indicates that the mean value of a specific trait for a given species increases relative to that at the highest light level. Forest plot (b) showing the effect of light reduction on RGR across different moderators. Overall effect and 95% confidence intervals are given. Where the zero line (red solid line) is not crossed by the confidence intervals, the effect of light reduction was significant (shown by the asterisks, * p < 0.05, ** p < 0.01, *** p < 0.001). Arrows indicate 95% confidence intervals extending beyond the limits of the plot. Values in parentheses are sample sizes. The effects of moderators are tested by QM at the 0.05 level. Scatter plot (c) denoting the relationships of the lnRRs of RGR with the percentage of light reduction. The sizes of dots were weighted by the inverse variance.

Figure 2.
Orchard plots showing mean estimate (black dot), confidence interval (bold black line), individual effect sizes (colored bubbles), and their precision (bubble sizes, inverse variance) for the response ratios (lnRRs) of (a) whole-plant traits (i.e., RM: ramet number, PH: plant height, SB: shoot biomass, RB: root biomass, R/S: root-to-shoot ratio, SC: soluble carbohydrates) and (b) leaf traits (i.e., LN: leaf number, NPR: net photosynthesis rate, Fv/Fm: leaf maximal quantum yield of photosystem II complex, Chl: chlorophyll content, Chl a/b: chlorophyll a/b ratio). The symbol k denotes the number of individual effects. When the confidence intervals do not cross the zero line (dash line), the effect of light reduction is significant.
Figure 2.
Orchard plots showing mean estimate (black dot), confidence interval (bold black line), individual effect sizes (colored bubbles), and their precision (bubble sizes, inverse variance) for the response ratios (lnRRs) of (a) whole-plant traits (i.e., RM: ramet number, PH: plant height, SB: shoot biomass, RB: root biomass, R/S: root-to-shoot ratio, SC: soluble carbohydrates) and (b) leaf traits (i.e., LN: leaf number, NPR: net photosynthesis rate, Fv/Fm: leaf maximal quantum yield of photosystem II complex, Chl: chlorophyll content, Chl a/b: chlorophyll a/b ratio). The symbol k denotes the number of individual effects. When the confidence intervals do not cross the zero line (dash line), the effect of light reduction is significant.

Figure 3.
Response ratio relationships between (a) ramet number (RM), (b) shoot biomass (SB), (c) root biomass (RB), (d) root-to-shoot ratio (R/S), (e) soluble carbohydrates (SC), and (f) net photosynthesis rate (NPR) with relative growth rate (RGR). The sizes of dots are weighted by the inverse variance.
Figure 3.
Response ratio relationships between (a) ramet number (RM), (b) shoot biomass (SB), (c) root biomass (RB), (d) root-to-shoot ratio (R/S), (e) soluble carbohydrates (SC), and (f) net photosynthesis rate (NPR) with relative growth rate (RGR). The sizes of dots are weighted by the inverse variance.

Figure 4.
Relationships between lnRRs of paired growth-related traits in the meta-analysis. Values in the matrix are slope estimates obtained through a mixed-effect regression model with the lnRR of a trait as a response (y-axis) and another trait as a predictor (x-axis). Significance levels of slope estimates are given for paired traits (* p < 0.05, ** p < 0.01, *** p < 0.001). RM: ramet number PH: plant height, R/S: root-to-shoot ratio, SB: shoot biomass, RB: root biomass, SC: soluble carbohydrates, LN: leaf number, NPR: net photosynthesis rate, Fv/Fm: leaf maximal quantum yield of photosystem II complex, Chl: chlorophyll content, Chl a/b: chlorophyll a/b ratio.
Figure 4.
Relationships between lnRRs of paired growth-related traits in the meta-analysis. Values in the matrix are slope estimates obtained through a mixed-effect regression model with the lnRR of a trait as a response (y-axis) and another trait as a predictor (x-axis). Significance levels of slope estimates are given for paired traits (* p < 0.05, ** p < 0.01, *** p < 0.001). RM: ramet number PH: plant height, R/S: root-to-shoot ratio, SB: shoot biomass, RB: root biomass, SC: soluble carbohydrates, LN: leaf number, NPR: net photosynthesis rate, Fv/Fm: leaf maximal quantum yield of photosystem II complex, Chl: chlorophyll content, Chl a/b: chlorophyll a/b ratio.

Figure 5.
Illustration of our main findings of the effect of light reduction on growth-related traits (a) and a simplified conceptual framework for potential mechanisms regulating the response of relative growth rate (RGR) to increased percentage of light reduction (PLR, (b)). Red arrows: positive effect, blue arrows: negative effect. RM: ramet number, PH: plant height, NPR: net photosynthesis rate, LN: leaf number, RB: root biomass, SB: shoot biomass.
Figure 5.
Illustration of our main findings of the effect of light reduction on growth-related traits (a) and a simplified conceptual framework for potential mechanisms regulating the response of relative growth rate (RGR) to increased percentage of light reduction (PLR, (b)). Red arrows: positive effect, blue arrows: negative effect. RM: ramet number, PH: plant height, NPR: net photosynthesis rate, LN: leaf number, RB: root biomass, SB: shoot biomass.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gao, J.; Hu, W.; Wang, J.; Cui, Y.; Li, L. Response of Growth-Related Traits of Submerged Macrophytes to Light Reduction: A Meta-Analysis. Sustainability 2023, 15, 5918. https://doi.org/10.3390/su15075918
AMA Style
Gao J, Hu W, Wang J, Cui Y, Li L. Response of Growth-Related Traits of Submerged Macrophytes to Light Reduction: A Meta-Analysis. Sustainability. 2023; 15(7):5918. https://doi.org/10.3390/su15075918
Chicago/Turabian StyleGao, Jing, Wei Hu, Jiawei Wang, Yichong Cui, and Liuxin Li. 2023. "Response of Growth-Related Traits of Submerged Macrophytes to Light Reduction: A Meta-Analysis" Sustainability 15, no. 7: 5918. https://doi.org/10.3390/su15075918
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.