
Screening Wild Pepper Germplasm for Resistance to Xanthomonas hortorum pv. gardneri

1
Institute of Genetics and Biotechnology, Applied Plant Genomics Group, Hungarian University of Agriculture and Life Sciences, Szent-Györgyi Albert Str. 4, 2100 Gödöllő, Hungary
2
GTIG, Török Ignác Secondary School of Gödöllő, Petőfi Sándor Str. 12-14, 2100 Gödöllő, Hungary
*
Author to whom correspondence should be addressed.
Sustainability 2023, 15(2), 908; https://doi.org/10.3390/su15020908
Submission received: 29 November 2022
/
Revised: 26 December 2022
/
Accepted: 30 December 2022
/
Published: 4 January 2023
(This article belongs to the Special Issue Plant Breeding Supporting the Sustainable Field Crop Production)
Abstract
:Bacterial spot disease on peppers is caused by four species of the genus Xanthomonas. This disease causes black spot lesions not only on the leaves but also on the fruit, leading to yield and quality loss. Xanthomonas species cause major disease outbreaks in tropical, subtropical and humid continental regions worldwide. Bacterial blight caused by xanthomonads occurs on both greenhouse- and field-grown peppers and is particularly important in areas characterized by hot and humid environmental conditions. As pesticides are currently not sufficiently effective in the control of bacterial spot, the development of pepper varieties resistant to Xanthomonas species, including X. hortorum pv. gardneri, is of primary importance for sustainable production. In our research, 119 lines of Capsicum baccatum from the USDA ARS gene bank (Griffin, GA) and MATE (Hungarian University of Agriculture and Life Sciences) were tested against strains of X. hortorum pv. gardneri under greenhouse conditions. Four accessions of the wild pepper species C. baccatum appeared to be resistant to seven strains of X. hortorum pv. gardneri in greenhouse trials. The resistant genotypes of X. hortorum pv. gardneri identified in this study can be used for the resistance gene pyramidation against different bacterial spotted Xanthomonas species in pepper.
1. Introduction
Pepper (Capsicum spp.) is one of the most important vegetables and spices worldwide because of its versatility, its high nutritional value, as well as its role in reducing human micronutrient deficiencies [1]. The chili pepper genus is currently known to consist of 43 species [2] that are native to temperate and tropical Central and South America, Mexico and the West Indies, five of which are domesticated: C. annuum, C. chinense, C. frutescens, C. baccatum, and C. pubescens [3]. Despite the crucial agronomic importance of this crop, diseases are still one of the most important damaging factors in pepper production, and only few can be controlled by chemical treatments. However, environmental and consumer concerns, as well as the risk of crop pathogen insensitivity, underline the importance of developing resistant pepper varieties as a more effective way of reducing the impact of disease [4,5]. Capsicum germplasm collections represent an immense genetic wealth resource this purpose, but screening for disease resistance is often a lengthy, labor-intensive, expensive and mostly complex process.
Bacterial spot disease of peppers occurs in both processing and fresh market pepper fields, particularly where growing conditions are characterized by high humidity. Known for its aggressiveness, the pathogen X. hortorum pv. gardneri causes large, star-shaped necrotic lesions on the foliage, stem and fruit. Diseased leaves drop prematurely, resulting in extensive defoliation, thus reducing plant productivity and exposing fruits to potential sunscalding. Stem lesions occur as narrow, light brown, longitudinally raised cankers. Fruit with such large lesions are non-marketable in both the fresh and processed markets.
Bacterial spot is caused by four bacterial species of the genus Xanthomonas: X. euvesicatoria, X. vesicatoria, X. perforans and X. hortorum pv. gardneri [6]. X. hortorum pv. gardneri was first described in 1957 [7]. The pathogen X. hortorum pv. gardneri was recognized as a causative agent of bacterial spot epidemics in the year 2000. Currently, there are few chemicals available that are effective against these pathogens, and even these are ineffective under high disease pressure and have a negative impact on the environment. Since the overuse of copper has led to the development of resistance in the bacterial population, there is a need to find new sustainable and environmentally friendly alternatives [8,9]. The growing number of bacterial spot diseases caused by emerging varieties of xanthomonads makes it increasingly urgent for growers to find practical and sustainable solutions. The use of varieties resistant to bacterial leaf spot offers a potential tool for disease control in field conditions. The spread of new Xanthomonas species over the last few years has made the discovery of new sources of resistance a major priority. Thus, host resistance is considered to be a critical element of disease management strategies.
Resistance breeding programs successfully developed commercial pepper lines with hypersensitive and quantitative resistance to various Xanthomonas species [10]. Genes regulating resistance have been found in X. euvesicatoria [11], X. perforans species T3 [12], X. perforans species T4 [13], and X. vesicatoria [14], and six dominant resistance genes have been identified in pepper to date, namely Bs1, Bs2, Bs3, Bs4C, Bs7 and BsT [15]. However, the persistence of resistance can change rapidly due to the changing geographical distribution of the pathogen and the rapid emergence of new pathogenic variants [16]. In the case of X. hortorum pv. gardneri, only one resistance gene source has been described so far, although in this case the infection was caused by a single Xantomonas strain (USVLXG1) and resistance was not tested with other Xanthomonas isolates [17]. The introduction of resistance genes with different mechanisms of action could provide a durable and sustainable solution to this ongoing host–pathogen arms race.
Commercial pepper varieties carrying recessive resistance genes, namely bs5 and bs6, have not proven effective against X. hortorum pv. gardneri. Based on current publications, the recessive resistance gene bs8 identified in C. annuum accession PI 163192 is the only one that was shown to confer resistance to X. hortorum pv. gardneri [18]. With the increasing frequency of Xanthomonas outbreaks around the world, there is a growing rationale to identify new sources of resistance in pepper to emerging bacterial species, including X. hortorum pv. gardneri and to incorporate these new resistance genes into breeding programs.
Preliminary results are presented below, which are aimed at screening wild pepper cultivars for heritable resistance to X. hortorum pv. gardneri. The aim of our current work was to identify new sources of resistance to X. hortorum pv. gardneri in pepper and to identify potential candidate genes involved in resistance to bacterial spot disease. A diverse USDA panel of pepper accessions were screened for resistance to X. hortorum pv. gardneri, and highly resistant accessions were identified in our work.
2. Materials and Methods
2.1. Plant Materials
The work was carried out on a collection of C. baccatum accessions originating from the USDA ARS genebank. We tested 119 accessions including 97 C. baccatum var. pendulum, 2 C. baccatum var. pratermissum, 7 C. baccatum var. baccatum, and 2 C. baccatum var. umbilicatum, and in 11 cases the variant was not specified (Table S1). In order to improve germination, the seeds were pre-soaked for four hours before they were sterilized with 10% calcium hypochlorite (Ca(ClO)2) solution for twenty minutes with a few drops of Tween20. The seeds were rinsed five times with sterile Milli-Q Water. After that, all of the seeds were placed on MS medium [19] with 20 g sucrose (MS20) in sterilized glass storage jars until the cotyledon stage. Afterwards, the plants were placed into pots filled with Klasmann Traysubstrate soil mixed with 50 g NPK and microelement fertilizer (14% N + 7% P + 21% K + 1% Mg + 1% microelements (B, Cu, Mn, Fe, Zn) to 70 L of soil.
The plants were then grown under controlled greenhouse conditions at 24 ± 2 °C, with 14/10 h photoperiods, and 50–70% relative humidity until they reached the six-mature-leaf stage. A cultivated variety C. annuum Fö “Fehérözön” was used as a susceptible control.
2.2. Bacterial Isolates and Plant Infiltration
To find a resistant phenotype, two Xg strains were used for plant infiltration—LMG962 from the Belgian Coordinated Collections of Microorganisms (BCCM) and SRB, which is a field isolate from Serbia [7]. Bacteria were cultivated in YDC [20] medium for two days in an incubator at a temperature of 30 °C. After two days of incubation, bacteria were collected and diluted with sterile Milli-Q Water to a final concentration of 106 cfu/mL. The infiltration was performed in planta with a needle-free syringe into the intercellular spaces on the abaxial side of the first true leaves. After infiltration, the plants were placed in plant growth chambers at 25 °C with a 16/8 h photoperiod, at 50–70% relative humidity.
The response to infection was monitored continuously, but the final evaluation was made after 7 days. The plants that showed complete resistance to both isolates were also tested with five further Xg strains: Xg51, Xg152, Xg153, Xg156, Xg177 (Table 1), which were donated by Brian Staskawicz (Innovative Genomics Institute University of California, Berkeley).
2.3. Bacterial Growth Test
In order to determine how the presence of resistance affected bacterial growth in resistant plants, leaf disc samples were collected from the infiltrated area at different times. For the test of the LMG962 isolate, four resistant and three susceptible plants were used in two replicates at 25 °C and 30 °C. The photoperiod and humidity were the same as for the accession testing. Sampling was performed at 0, 2, 4, 6, 8, 10, 12 days post inoculation (dpi) from the inoculated area. In the case of resistant plants, re-isolation was also performed 30 days after infection to monitor possible changes in bacterial counts. The isolated leaf discs were macerated in 100 µL sterilized water and prepared in dilution lines and were scattered on YDC medium. After two or three days, the colonies were counted. The cfu/cm2 of leaves was calculated by counting the number of adult colonies.
3. Results
3.1. Evaluation of C. baccatum Accessions for the Susceptibility and Resistance to X. hortorum pv. gardneri
One hundred and nineteen C. baccatum accessions were inoculated with two highly virulent strains (LMG962 and SRB) of X. hortorum pv. gardneri and evaluated for symptoms. Disease evaluation started one week after inoculation and the earliest typical disease symptoms were observed on the leaves 3–6 days after inoculation on the infected pepper plants. The average incubation period varied between 4 and 7 days until the initial abnormal discoloration appeared, while the length of time required to develop complete leaf necrosis in sensitive individuals varied between 6 and 10 days.
Susceptible symptoms typically started with chlorotic lesions, which developed into angular, water-soaked lesions and eventually into brown or black necrotic lesions. Phenotyping of the control line “Fehérözön” (highly susceptible) conformed to the expected disease reactions. In our studies, we found that C. baccatum accessions differed in terms of the earliness and intensity of symptoms. The vast majority of the accessions were susceptible to both strains, showing a similar phenotype to “Fehérözön” used as a control. A small part of the accessions showed partial resistance to one of the X. hortorum pv. gardneri strain. Based on the evaluation of their reactions, four of the 119 C. baccatum genotypes showed strong resistance to both Xg isolates used in the first selection round (Figure 1).
In the second phase, all four previously resistant phenotypes of C. baccatum accessions were challenged with five additional Xg isolates. Based on our results, all four accessions also showed strong resistance (Table 2) to all five additional Xg isolates (Xg51, Xg152, Xg153, Xg156, Xg 177).
In the resistant plants, the infected area turned light green, but at 30 dpi, it was no longer possible to distinguish between infiltrated and untreated areas. Small necrotic spots appeared on the leaves at 10–12 dpi typically along the leaf veins, but these lesions increased very little if at all after 30 days (Figure 2). No difference was observed between the emerging phenotype in resistant plants according to the bacterial strain used for inoculation.
3.2. Evaluation of the Bacterial Growth Test
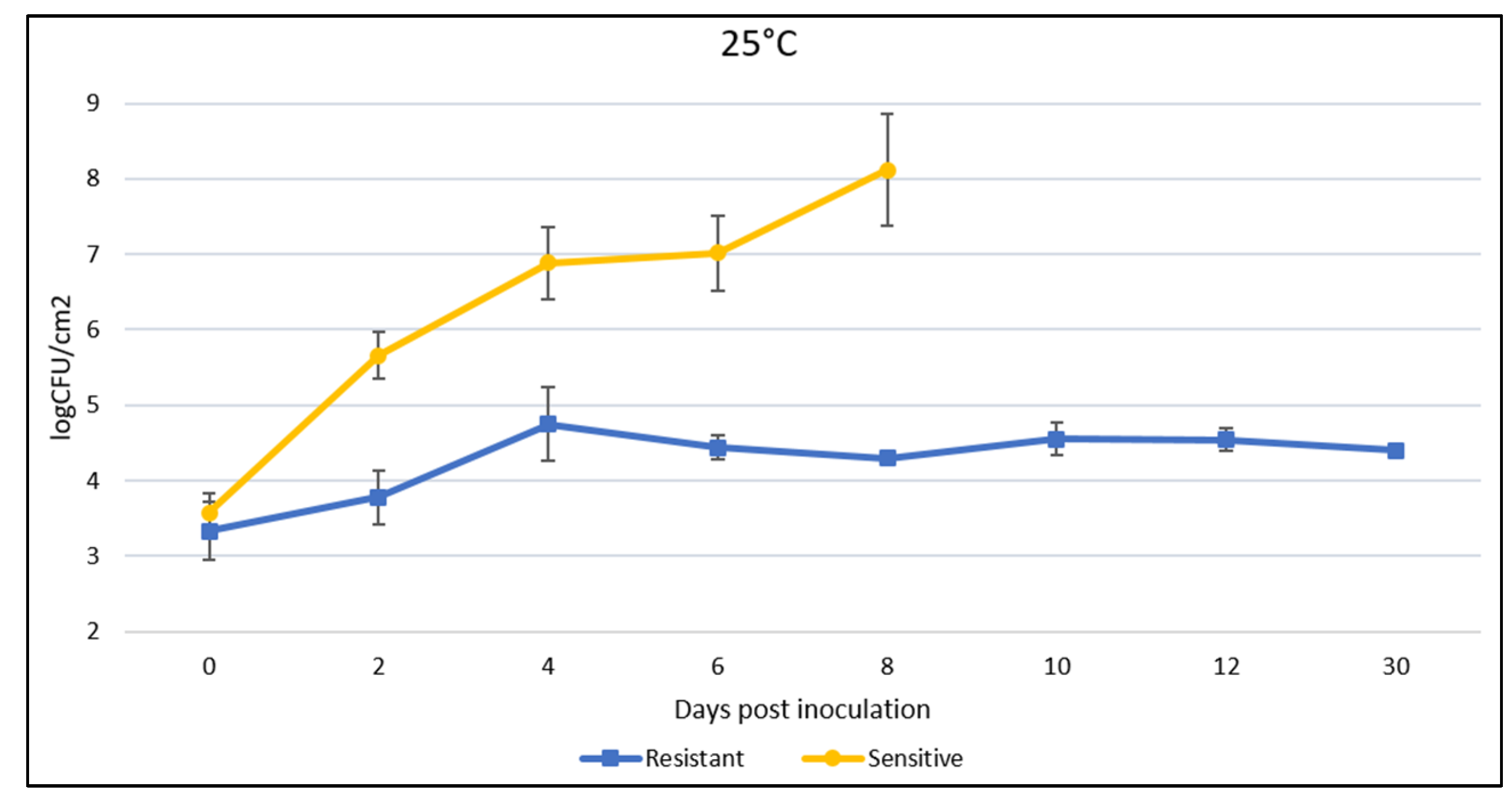
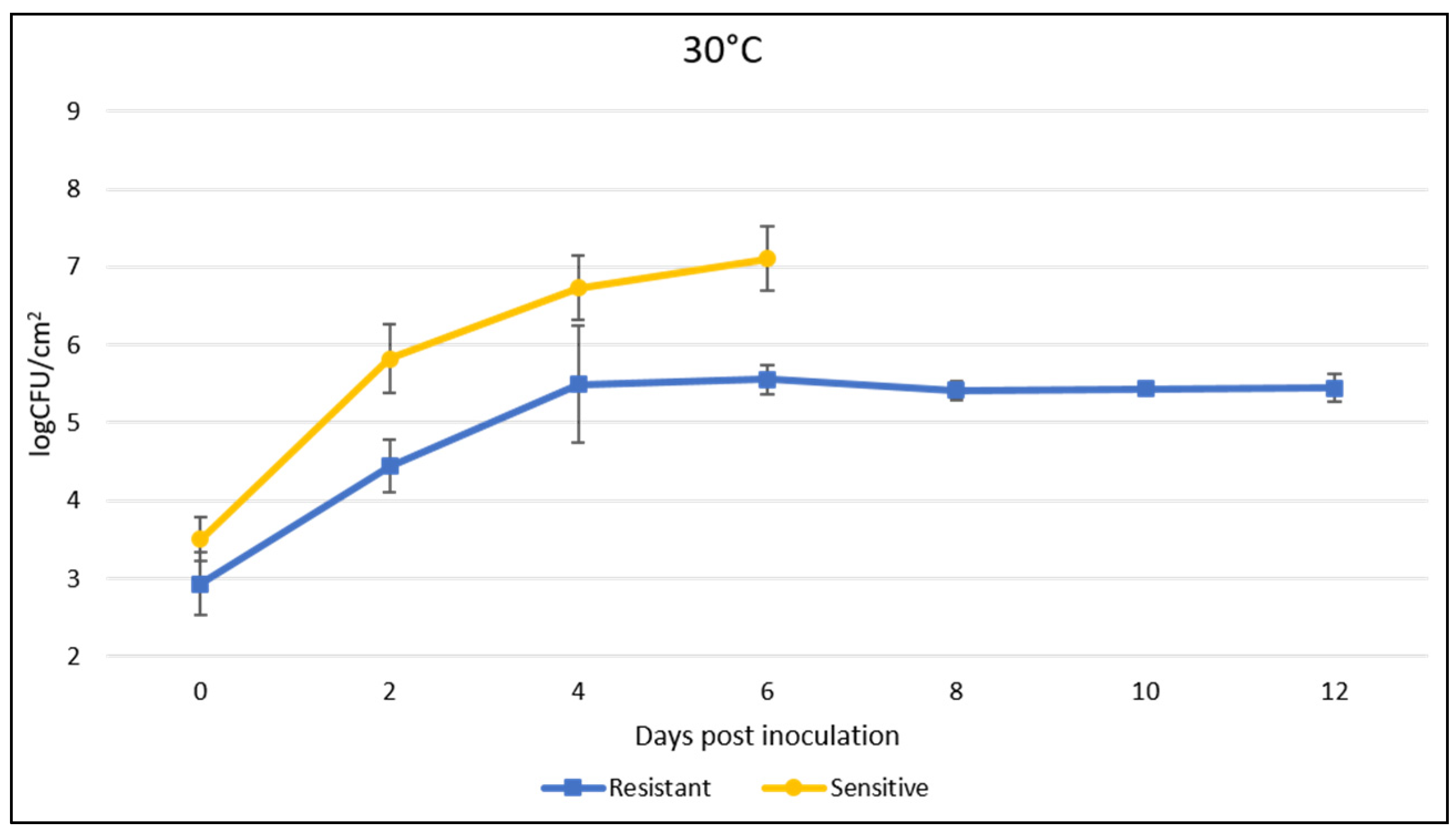
We determined the effect of the identified resistance gene on bacterial growth in the plant and the correlation of visual symptoms with bacterial population levels. The growth in the populations of different bacterial races was monitored for 12 days after inoculation and incubation at 25 °C and 30 °C. “Fehérözön” was used as a negative control. Compared to the almost equal initial values, the bacterial growth curve in planta at 25 °C indicated that resistant plants reached a maximum of 4.75 log at 4 dpi and then showed a minimal decrease with no significant change in the following 6–12 days. In contrast, in the case of the sensitive plants, a value of 6.7 log at 4 dpi was obtained and a steady increase was detected until leaf drop at 8 dpi (Figure 3). In the majority of cases, the infected leaves of the susceptible plants had fallen off by the eighth day and sampling could not be continued. At 30 °C, the resistant plants reached a maximum value of 5.55 log at 6 dpi and then a slight decrease was observed. Susceptible plants showed a value of 7.1 log at 6 dpi until leaf drop (Figure 4). In susceptible plants, the temperature had no significant effect on the growth curve of Xg bacteria, but significantly affected the tolerance of plants to infection, because inoculated leaves fell off earlier at higher temperatures. Plants kept at 25 °C were also sampled at 30 dpi and the results show that no significant change in the cfu value in the leaf occurred. In the case of plants kept at 30 °C, it was not possible to collect a sample after one month, due to early aging of the plant and leaf fall.
4. Discussion
C. baccatum is the most popular and widely cultivated chili pepper in South America. There is a wild form called C. baccatum var. baccatum and a domesticated form (C. baccatum var. pendulum) [21]. Brazil is considered to be the center of the genetic diversity of Capsicum spp., and at least 116 accessions already described are found in Brazil [22,23]. It is expected that new species [2] and new resistance genes for pepper breeding will be found in this important and untapped genetic resource. Pathogens are increasingly threatening the cultivation and yield of most Capsicum species worldwide. In this context, the search for new sources of resistance remains a major challenge today and Capsicum germplasm collections represent a promising target for this purpose.
Previous searches for resistance to X. hortorum pv. gardneri concentrated on the Xg isolates which are most prevalent in the US. In contrast to previous research, in our present work we also used the isolates most commonly found in Europe and the US. The results of this screening study in which pepper accessions were inoculated with seven Xg strains indicated that there were significant differences in resistance between PI lines from the USDA ARS pepper germplasm collection. Screening was carried out in two stages. As a first step, 119 accessions from the C. baccatum collection were challenged with two different strains of X. hortorum pv. gardneri to determine the extent and types of resistance in C. baccatum. Sensitive symptoms of bacterial spot were observed in 107 accessions and Xg was recovered from all plants, indicating that none of the accessions carried a source of resistance to this pathogen. Eight accessions showed partial resistance to one of the Xg isolates and strong resistance was identified in four PI lines (Cbp1, Cbp2, Cbp3 and Cbp4). In the second stage, screening was carried out on Cbp1, Cbp2, Cbp3 and Cbp4 accessions using the five potentially important isolates in Europe and the US. Complete screening was carried out under controlled conditions in growth chambers using a method (leaf screening of young plants with a concentrated bacterial suspension) that clearly indicated the presence of the pathogen and the rapid onset of symptoms.
The screening results showed two main types of resistance (Table 2) in the tested plant population. Species-specific resistance was the most common and was present in 6% of all accessions tested. Resistance to all species used (potential species-nonspecific resistance) was very rare and was present in 3% of accessions tested.
5. Conclusions
Species shifts in bacterial spot pathogen populations are very common [10,24] and pose a serious problem and challenge for pepper breeding programs due to the decline in resistance persistence [25]. Therefore, there is an increasing need to identify new sources of resistance to these pathogens. The resistance germplasm of X. hortorum pv. gardneri identified in this study can be of significant importance for both breeders and researchers and is highly relevant for introgression into pepper lines and in terms of gene pyramidation efforts against various Xanthomonas species. The resistance genes that we identified in C. baccatum might also have significant potential for resistance inhibition in other pepper species such as C. annuum. Interspecific crosses provide the opportunity to pass genes from one species to another, but in most cases, this can present several difficulties. A successful wide hybridization among species is highly dependent on the genetic distance between species. According to our current knowledge, two options are available for introgressing C. baccatum genes into C. annuum. The first option is using C. chinense and C. frutescens as bridge species [26], while the second option is to carry out direct crosses between C. annuum and C. baccatum combined with in vitro embryo rescue [27]. Our future goal is to use both methods to introduce our newly identified valuable resistance gene into C. annuum. Using the four resistant parental lines, crosses are underway with the aim of creating F1 and F2 progeny to determine the inheritance of disease resistance.
The new sources of resistance identified in this study provide a basis for further work on breeding disease-resistant varieties that are resilient to the changing population structure of pathogens. The current study showed that the different genotypes of C. baccatum differed in their response to wilt caused by the pathogen Xanthomonas, which also demonstrated that the pepper PI collection offered a valuable public source of resistance for pepper breeders to develop varieties resistant to bacterial spot. In conclusion, the information in this study can be summarized in that the use of a resistant genotype, together with cultural practices and sanitary control measures, is considered the most feasible and sustainable way to control X. hortorum pv. gardneri wilt.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/su15020908/s1, Table S1: Screening of C. baccatum germplasm to X. hortorum pv. gardneri.
Author Contributions
Conceptualization, Z.G.T. and Z.T.; methodology, Z.G.T. and Z.T.; software, Z.T.; validation, Z.G.T., Z.S. and Z.T.; formal analysis, Z.G.T., M.T., Z.S. and Z.T.; investigation, Z.G.T. and Z.T.; resources, Z.G.T. and Z.T.; data curation, Z.G.T. and Z.T.; writing—original draft preparation, Z.G.T. and Z.T.; writing—review and editing, Z.G.T., S.F., Z.S. and Z.T.; visualization, Z.T.; supervision, S.F. and Z.T.; project administration, Z.G.T. and Z.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- FAOSTAT, Food and Agriculture Organization (FAO). 2021. Available online: http://www.fao.org/faostat/en/#home (accessed on 15 April 2021).
- Barboza, G.E.; García, C.C.; Scaldaferro, M.; Bohs, L. An amazing new Capsicum (Solanaceae) species from the Andean-Amazonian Piedmont. PhytoKeys 2020, 167, 13–29. [Google Scholar] [CrossRef] [PubMed]
- USDA, ARS, National Genetic Resources Program. Germplasm Resources Information Network (GRIN). National Germplasm Resources Laboratory, Beltsville, Maryland. Available online: http://www.arsgrin.gov/cgi-bin/npgs/html/exsplist.pl (accessed on 23 May 2015).
- Mundt, C.C. Durable resistance: A key to sustainable management of pathogens and pests. Infect. Genet. Evol. 2014, 27, 446–455. [Google Scholar] [CrossRef] [PubMed]
- Collinge, D.B.; Sarrocco, S. Transgenic approaches for plant disease control: Status and prospects. Plant Pathol. 2022, 71, 207–225. [Google Scholar] [CrossRef]
- Jones, J.B.; Lacy, G.H.; Bouzar, H.; Stall, R.E.; Schaad, N.W. Reclassification of the xanthomonads associated with bacterial spot disease of tomato and pepper. Syst. Appl. Microbiol. 2004, 27, 755–762. [Google Scholar] [CrossRef] [Green Version]
- Šutic, D. Bakterioze crvenog patlidzana (Tomato bacteriosis). Rev. Appl. Mycol. 1957, 36, 734–735. [Google Scholar]
- Khanal, S.; Hind, S.R.; Babadoost, M. Occurrence of Copper-Resistant Xanthomonas perforans and X. gardneri in Illinois Tomato Fields. Plant Health Prog. 2020, 21, 338–344. [Google Scholar] [CrossRef]
- El-Fiki, I.A.I.; Youssef, M.; Eman, H.O. Controlling The Bacterial Leaf Spot Disease in Pepper Caused by Xanthomonas vesicatoria Using Natural Bacteritoxicants. Egypt. Acad. J. Biol. Sci. F. Toxicol. Pest Control. 2022, 14, 229–245. [Google Scholar] [CrossRef]
- Stall, R.E.; Jones, J.B.; Minsavage, G.V. Durability of resistance in tomato and pepper to xanthomonads causing bacterial spot. Annu. Rev. Phytopathol. 2009, 47, 265–284. [Google Scholar] [CrossRef]
- Yang, W.C.; Sacks, E.J.; Ivey, M.L.L.; Miller, S.A.; Francis, D.M. Resistance in Lycopersicon esculentum intraspecific crosses to race T1 strains of Xanthomonas campestris pv. vesicatoria causing bacterial spot of tomato. Phytopathology 2005, 95, 519–527. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Hutton, S.F.; Robbins, M.D.; Sim, S.C.; Scott, J.W.; Yang, W.; Jones, J.B.; Francis, D.M. Molecular mapping of hypersensitive resistance from tomato ‘Hawaii 7981’ to Xanthomonas perforans race T3. Phytopathology 2011, 101, 1217–1223. [Google Scholar] [CrossRef] [Green Version]
- Sharlach, M.; Dahlbeck, D.; Liu, L.; Chiu, J.; Jiménez-Gómez, J.M.; Kimura, S.; Koenig, D.; Maloof, J.N.; Sinha, N.; Minsavage, G.V.; et al. Fine genetic mapping of RXopJ4, a bacterial spot disease resistance locus from Solanum pennellii LA716. Theor. Appl. Genet. 2013, 126, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.W.; Francis, D.M.; Miller, S.A.; Somodi, G.C.; Jones, J.B. Tomato bacterial spot resistance derived from PI 114490; inheritance to race T2 and relationship across three pathogen races. J. Am. Soc. Hortic. Sci. 2003, 128, 698–703. [Google Scholar] [CrossRef] [Green Version]
- Potnis, N.; Krasileva, K.; Chow, V.; Almeida, N.F.; Patil, P.B.; Ryan, R.P.; Sharlach, M.; Behlau, F.; Dow, J.M.; Momol, M.T.; et al. Comparative genomics reveals diversity among xanthomonads infecting tomato and pepper. BMC Genom. 2011, 12, 146. [Google Scholar] [CrossRef] [Green Version]
- Potnis, N.; Timilsina, S.; Strayer, A.; Shantharaj, D.; Barak, J.D.; Paret, M.L.; Vallad, G.E.; Jones, J.B. Bacterial spot of tomato and pepper: Diverse Xanthomonas species with a wide variety of virulence factors posing a worldwide challenge. Mol. Plant Pathol. 2015, 16, 907–920. [Google Scholar] [CrossRef] [PubMed]
- Potnis, N.; Branham, S.E.; Jones, J.B.; Wechter, W.P. Genome-Wide Association Study of Resistance to Xanthomonas gardneri in the USDA Pepper (Capsicum) Collection. Phytopathology 2019, 109, 1217–1225. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Minsavage, G.V.; Gill, U.S.; Hutton, S.F.; Jones, J.B. Identification and Mapping of bs8, a Novel Locus Conferring Resistance to Bacterial Spot Caused by Xanthomonas gardneri. Phytopathology 2022, 112, 1640–1650. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Schaad, N.W. Laboratory Guide for the Identification of Plant Pathogenic Bacteria; The American Phytopathological Society: St. Paul, MN, USA, 1988; pp. 81–82. [Google Scholar]
- Dewitt, D.; Bosland, P.W. The Complete Chile Pepper Book—A Gardener ’s Guide to Choosing, Growing, Preserving and Cooking; Timber Press: Portland, OR, USA, 2009; p. 336. [Google Scholar]
- Moscone, E.A.; Scaldaferro, M.A.; Grabiele, M.; Cecchini, N.M.; García, Y.S.; Jarret, R.; Daviña, J.R.; Ducasse, D.A.; Barboza, G.E.; Ehrendorfer, F. The evolution of chili peppers (Capsicum-Solanaceae): A cytogenetic perspective. Acta Hortic. 2007, 745, 137–169. [Google Scholar] [CrossRef]
- Cardoso, R.; Ruas, C.F.; Giacomin, R.M.; Ruas, P.M.; Ruas, E.A.; Barbieri, R.L.; Rodrigues, R.; Gonçalves, L.S.A. Genetic variability in Brazilian Capsicum baccatum germplasm collection assessed by morphological fruit traits and AFLP markers. PLoS ONE 2018, 13, e0196468. [Google Scholar] [CrossRef] [Green Version]
- Horvath, D.M.; Stall, R.E.; Jones, J.B.; Pauly, M.H.; Vallad, G.E.; Dahlbeck, D.; Staskawicz, B.J.; Scott, J.W. Transgenic resistance confers effective field level control of bacterial spot disease in tomato. PLoS ONE 2012, 7, e42036. [Google Scholar] [CrossRef]
- Gassmann, W.; Dahlbeck, D.; Cjesnokova, O.; Minsavage, G.V.; Jones, J.B.; Staskawicz, B.J. Molecular evolution of virulence in natural field strains of Xanthomonas campestris pv. vesicatoria. J. Bacteriol. 2000, 182, 7053–7059. [Google Scholar] [CrossRef] [PubMed]
- Manzur, J.P.; Fita, A.; Prohens, J.; Rodríguez-Burruezo, A. Successful Wide Hybridization and Introgression Breeding in a Diverse Set of Common Peppers (Capsicum annuum) Using Different Cultivated Ají (C. baccatum) Accessions as Donor Parents. PLoS ONE 2015, 10, e0144142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potnis, N.; Minsavage, G.; Smith, J.K.; Hurlbert, J.C.; Norman, D.; Rodrigues, R.; Stall, R.E.; Jones, J.B. Avirulence proteins AvrBs7 from Xanthomonas gardneri and AvrBs1.1 from Xanthomonas euvesicatoria contribute to a novel gene-for-gene interaction in pepper. Mol. Plant-Microbe Interact. 2012, 25, 307–320. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Symptoms of bacterial spot 7 days after inoculation with strain LMG962 in susceptible Fö (a,b) and the resistant phenotype on Cbp2 plants infected with strain LMG962 (c,d) and SRB (e,f). Adaxial and abaxial view.
Figure 1.
Symptoms of bacterial spot 7 days after inoculation with strain LMG962 in susceptible Fö (a,b) and the resistant phenotype on Cbp2 plants infected with strain LMG962 (c,d) and SRB (e,f). Adaxial and abaxial view.

Figure 2.
Symptoms of bacterial spot 30 days after inoculation with strain LMG962, SRB and Xg51 in a resistant accession plant.
Figure 2.
Symptoms of bacterial spot 30 days after inoculation with strain LMG962, SRB and Xg51 in a resistant accession plant.

Figure 3.
Time course of bacterial population growth at 25 °C after infiltration of leaves of resistant and susceptible plants by LMG962 strains.
Figure 3.
Time course of bacterial population growth at 25 °C after infiltration of leaves of resistant and susceptible plants by LMG962 strains.

Figure 4.
Time course of bacterial population growth at 30 °C after infiltration of leaves of resistant and susceptible plants by LMG962 strains.
Figure 4.
Time course of bacterial population growth at 30 °C after infiltration of leaves of resistant and susceptible plants by LMG962 strains.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Xg strains used in this study and their origin.
| Species | Strain Name | Origin | Host Isolated | Year Isolated | Isolation ID | Collector |
|---|---|---|---|---|---|---|
| X. hortorum pv. gardneri | LMG962 | Republic of Yugoslavia | tomato | 1957 | - | Sutic D. |
| SRB | Republic of Yugoslavia | tomato | 1957 | - | Sutic D. | |
| Xg51 | - 1 | - | - | - | T. Minsavage | |
| Xg152 | - | - | - | - | - | |
| Xg153 | Gibsonburg, OH | tomato | 2010 | SM194-10 | S. Miller | |
| Xg156 | Blissfield, MI | tomato | 2010 | SM177-10 | S. Miller | |
| Xg177 | Sandusky Co., OH | tomato | 2012 | SM795-12 | S. Miller |
1 means that the exact origin, host, year, isolation ID or collector of the isolate has not been established.
Table 2.
C. baccatum var. pendulum accessions showing partial or complete resistance to the X. hortorum pv. gardneri strains used.
Table 2.
C. baccatum var. pendulum accessions showing partial or complete resistance to the X. hortorum pv. gardneri strains used.
| Species | Variety | Accession | SRB | LMG962 | XG51 | XG152 | XG153 | XG156 | XG177 |
|---|---|---|---|---|---|---|---|---|---|
| C.baccatum | pendulum | Cbp1 | + | + | + | + | + | + | + |
| Cbp2 | + | + | + | + | + | + | + | ||
| Cbp3 | + | + | + | + | + | + | + | ||
| Cbp4 | + | + | + | + | + | + | + | ||
| PI 441541 | + | − | |||||||
| PI 441542 | + | − | |||||||
| PI 370010 | − | + | |||||||
| PI 441543 | + | - | |||||||
| PI 441578 | + | − | |||||||
| PI 441552 | − | + | |||||||
| PI 441533 | − | + | |||||||
| PI 441520 | + | − |
+: resistant response to infection, −: sensitive response to infection.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tóth, Z.G.; Tóth, M.; Fekete, S.; Szabó, Z.; Tóth, Z. Screening Wild Pepper Germplasm for Resistance to Xanthomonas hortorum pv. gardneri. Sustainability 2023, 15, 908. https://doi.org/10.3390/su15020908
AMA Style
Tóth ZG, Tóth M, Fekete S, Szabó Z, Tóth Z. Screening Wild Pepper Germplasm for Resistance to Xanthomonas hortorum pv. gardneri. Sustainability. 2023; 15(2):908. https://doi.org/10.3390/su15020908
Chicago/Turabian StyleTóth, Zoltán Gábor, Máté Tóth, Sándor Fekete, Zoltán Szabó, and Zoltán Tóth. 2023. "Screening Wild Pepper Germplasm for Resistance to Xanthomonas hortorum pv. gardneri" Sustainability 15, no. 2: 908. https://doi.org/10.3390/su15020908
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.