
Microalgae Cultivated under Magnetic Field Action: Insights of an Environmentally Sustainable Approach
 , ,
, ,  ,
,  , ,
, ,  , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Microalga Cultures and Magnetic Fields
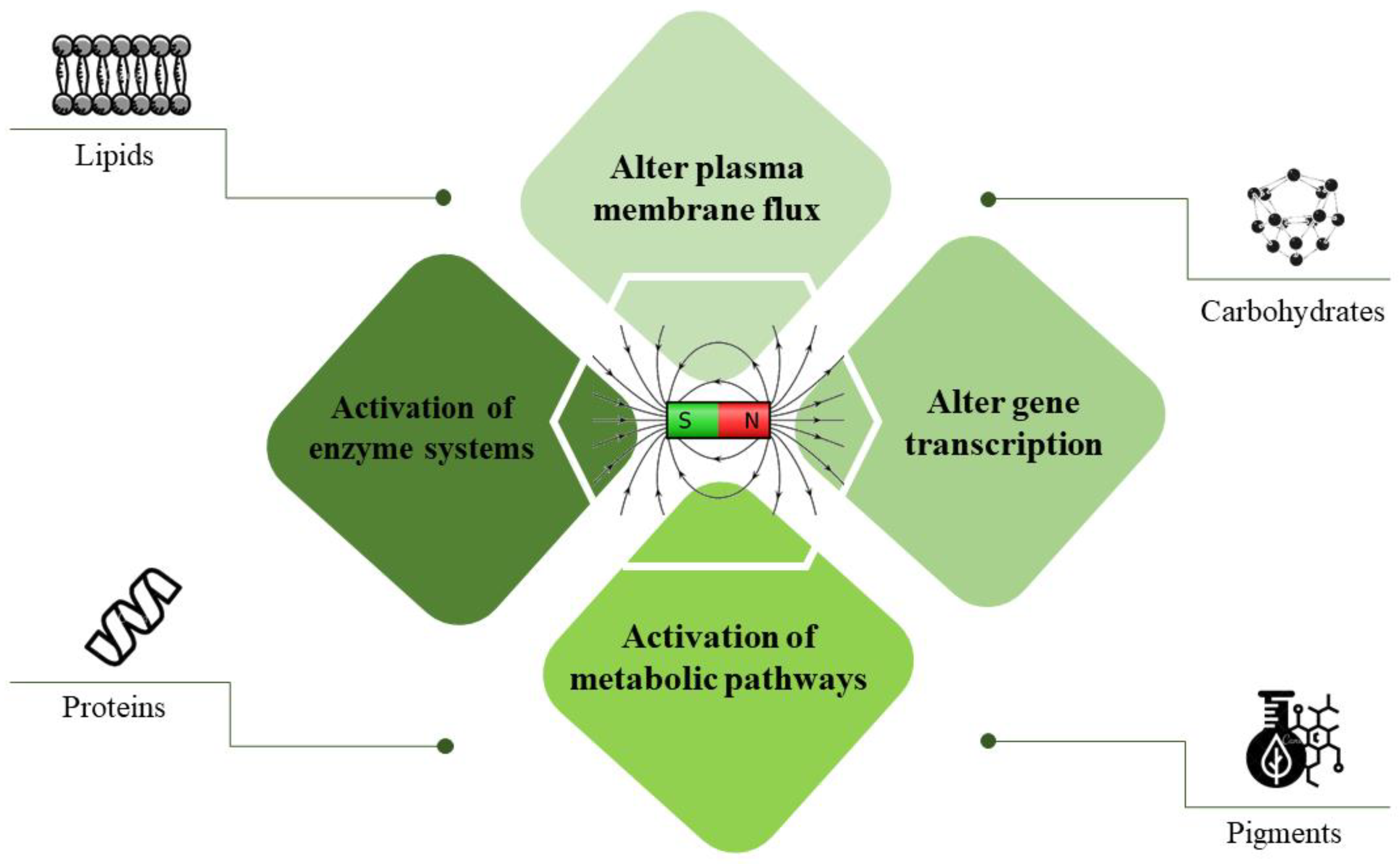
2.1. Bioeffects of Magnetic Field on Microalga Cultivation
2.2. Magnetic Field Effect on Microalga Cell Growth and Cell Size
2.3. Magnetic Field Effect on the Chemical Composition of Microalgae
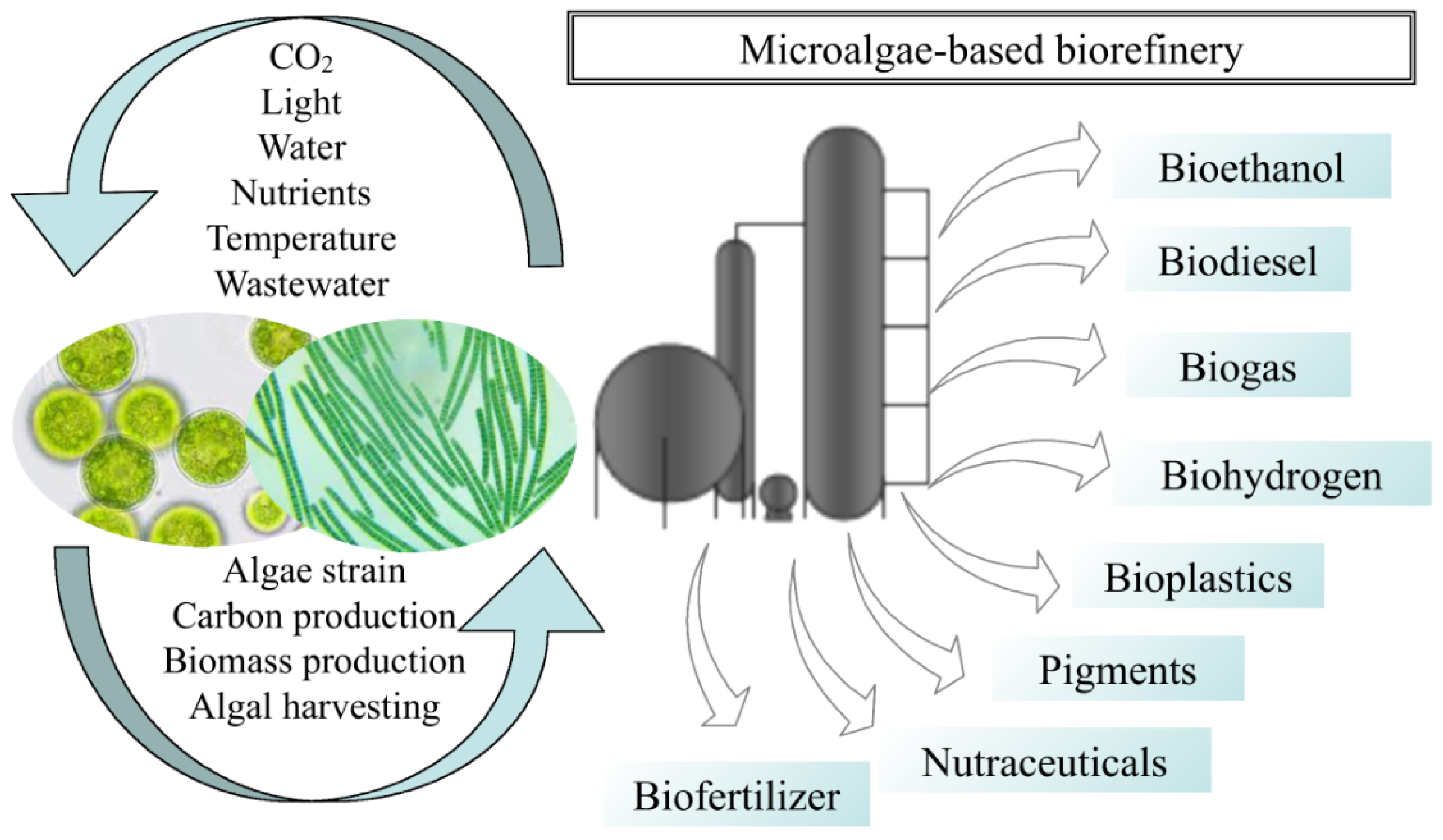
3. Microalga-Based Biorefinery
3.1. CO2 Mitigation
3.2. Wastewater Treatment
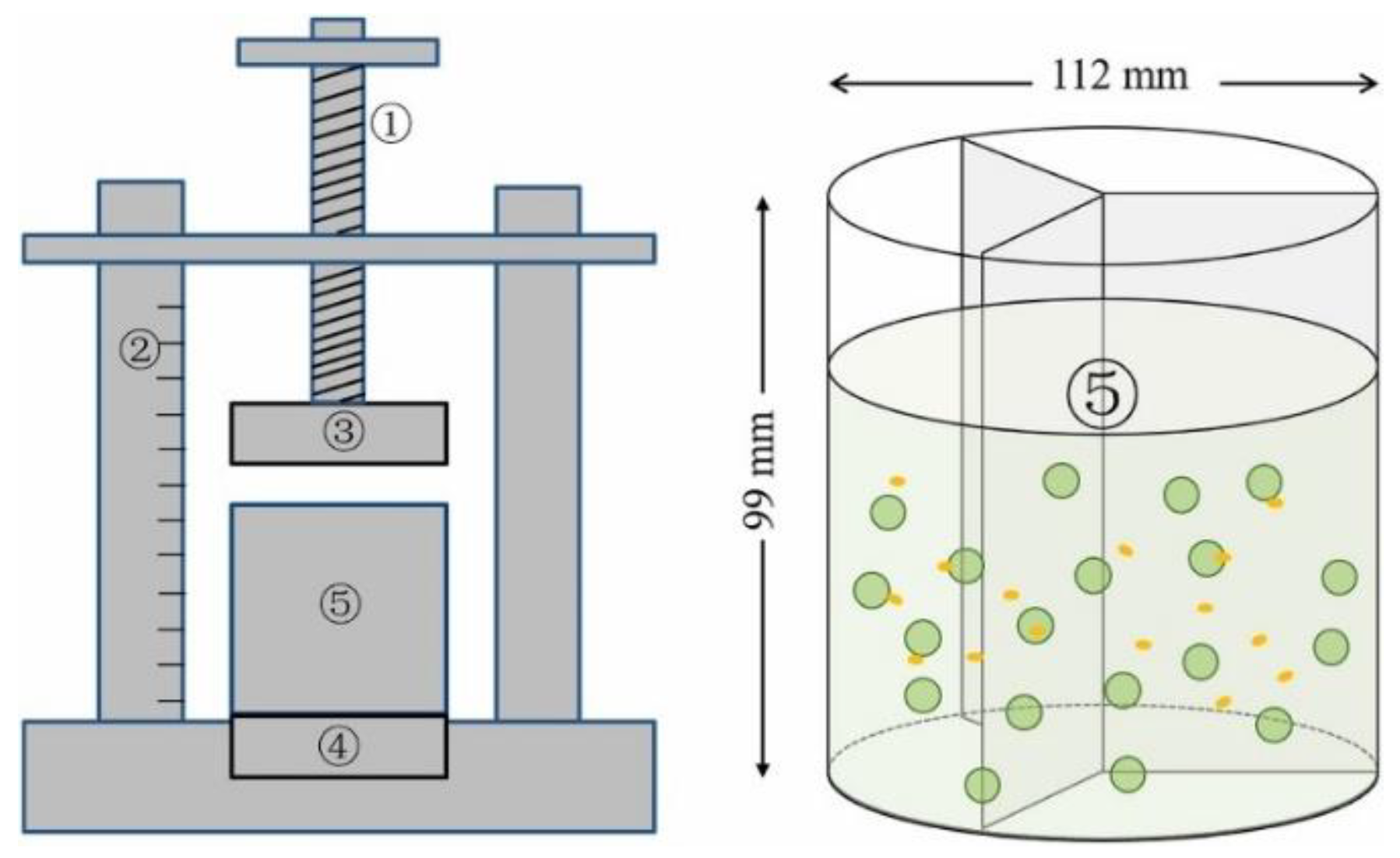
3.3. Different Ways of MF Application to Microalga Cultivation
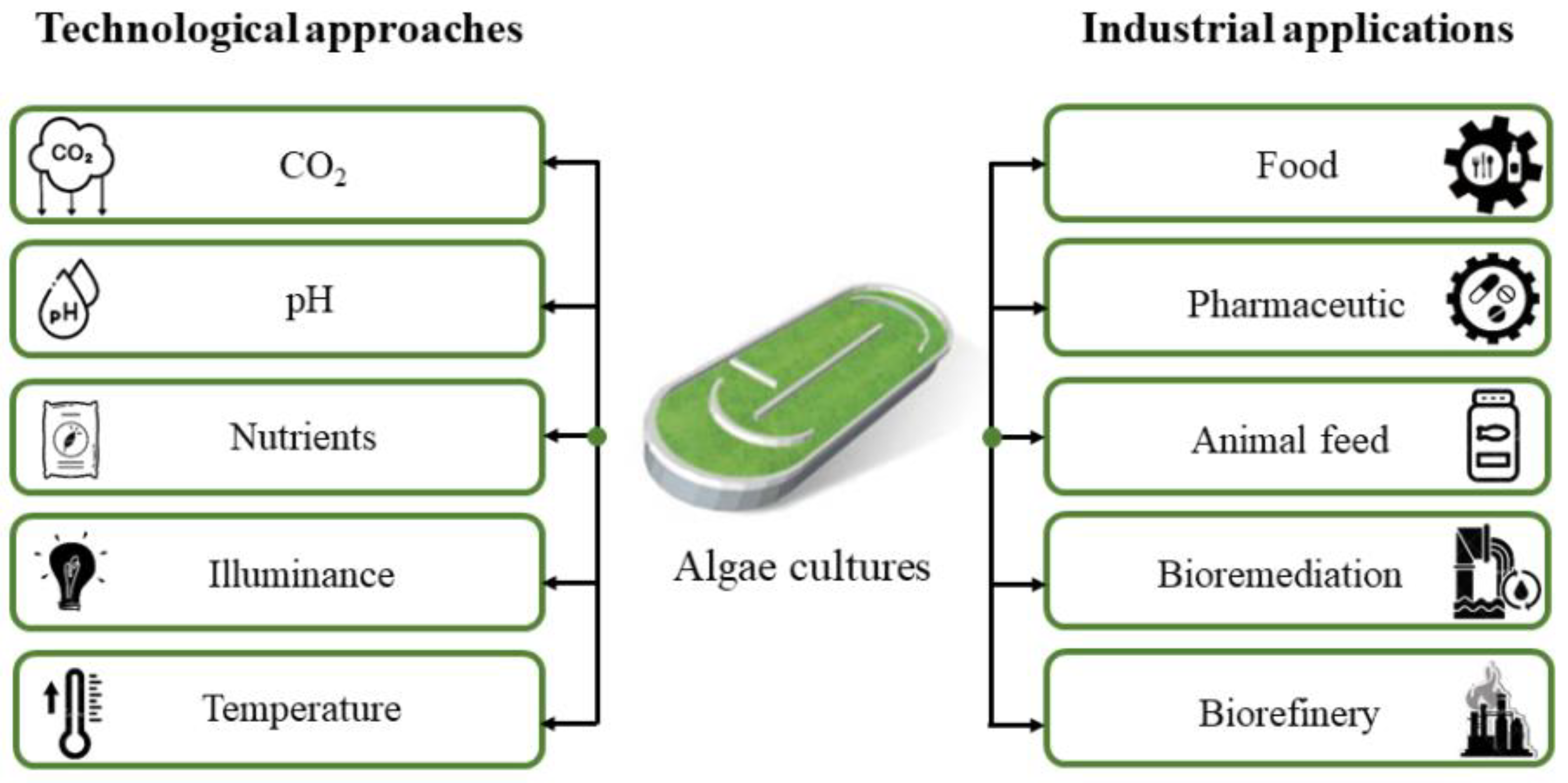
4. Sustainable and Environmentally Friendly Application of Microalgae
5. Final Considerations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chew, K.W.; Yap, J.Y.; Show, P.L.; Suan, N.H.; Juan, J.C.; Ling, T.C.; Lee, D.-J.; Chang, J.-S. Microalgae biorefinery: High value products perspectives. Bioresour. Technol. 2017, 229, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Hunt, R.W.; Zavalin, A.; Bhatnagar, A.; Chinnasamy, S.; Das, K.C. Electromagnetic biostimulation of living cultures for biotechnology, biofuel and bioenergy applications. Int. J. Mol. Sci. 2009, 10, 4515–4558. [Google Scholar] [CrossRef]
- Spanò, N.; Di Paola, D.; Albano, M.; Manganaro, A.; Sanfilippo, M.; D’Iglio, C.; Capillo, G.; Savoca, S. Growth performance and bioremediation potential of Gracilaria gracilis (Steentoft, L.M. Irvine & Farnham, 1995). Int. J. Environ. Stud. 2022, 79, 748–760. [Google Scholar]
- Tu, R.; Jin, W.; Xi, T.; Yang, Q.; Han, S.-F.; Abomohra, A.E.-F. Effect of static magnetic field on the oxygen production of Scenedesmus obliquus cultivated in municipal wastewater. Water Res. 2015, 86, 132–138. [Google Scholar] [CrossRef]
- Han, S.; Jin, W.; Chen, Y.; Tu, R.; Abomohra, A.E.-F. Enhancement of lipid production of Chlorella pyrenoidosa cultivated in municipal wastewater by magnetic treatment. Appl. Biochem. Biotechnol. 2016, 180, 1043–1055. [Google Scholar] [CrossRef]
- Santos, L.O.; Deamici, K.M.; Menestrino, B.C.; Garda-Buffon, J.; Costa, J.A.V. Magnetic treatment of microalgae for enhanced product formation. World J. Microbiol. Biotechnol. 2017, 33, 169. [Google Scholar] [CrossRef] [PubMed]
- Zieliński, M.; Rusanowska, P.; Dębowski, M.; Hajduk, A. Influence of static magnetic field on sludge properties. Sci. Total Environ. 2018, 625, 738–742. [Google Scholar] [CrossRef] [PubMed]
- Bauer, L.M.; Costa, J.A.V.; da Rosa, A.P.C.; Santos, L.O. Growth stimulation and synthesis of lipids, pigments and antioxidants with magnetic fields in Chlorella kessleri cultivations. Bioresour. Technol. 2017, 244, 1425–1432. [Google Scholar] [CrossRef] [PubMed]
- Serrano, G.; Miranda-Ostojic, C.; Ferrada, P.; Wulff-Zotelle, C.; Maureira, A.; Fuentealba, E.; Gallardo, K.; Zapata, M.; Rivas, M. Response to Static Magnetic Field-Induced Stress in Scenedesmus obliquus and Nannochloropsis gaditana. Mar. Drugs 2021, 19, 527. [Google Scholar] [CrossRef]
- Costa, S.S.; Peres, B.P.; Machado, B.R.; Costa, J.A.V.; Santos, L.O. Increased lipid synthesis in the culture of Chlorella homosphaera with magnetic fields application. Bioresour. Technol. 2020, 315, 123880. [Google Scholar] [CrossRef]
- Deamici, K.M.; Santos, L.O.; Costa, J.A.V. Magnetic field action on outdoor and indoor cultures of Spirulina: Evaluation of growth, medium consumption and protein profile. Bioresour. Technol. 2018, 249, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Small, D.P.; Hüner, N.P.; Wan, W. Effect of static magnetic fields on the growth, photosynthesis and ultrastructure of Chlorella kessleri microalgae. Bioelectromagnetics 2012, 33, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Deamici, K.M.; Costa, J.A.V.; Santos, L.O. Magnetic fields as triggers of microalga growth: Evaluation of its effect on Spirulina sp. Bioresour. Technol. 2016, 220, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Veiga, M.C.; Fontoura, M.M.; de Oliveira, M.G.; Costa, J.A.V.; Santos, L.O. Magnetic fields: Biomass potential of Spirulina sp. for food supplement. Bioprocess Biosyst. Eng. 2020, 43, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- Deamici, K.M.; Santos, L.O.; Costa, J.A.V. Magnetic field as promoter of growth in outdoor and indoor assays of Chlorella fusca. Bioprocess Biosyst. Eng. 2021, 44, 1453–1460. [Google Scholar] [CrossRef]
- Deamici, K.M.; de Morais, M.G.; Santos, L.O.; Muylaert, K.; Gardarin, C.; Costa, J.A.V.; Laroche, C. Static magnetic fields effects on polysaccharides production by different microalgae strains. Appl. Sci. 2021, 11, 5299. [Google Scholar] [CrossRef]
- Deamici, K.M.; Santos, L.O.; Costa, J.A.V. Use of static magnetic fields to increase CO2 biofixation by the microalga Chlorella fusca. Bioresour. Technol. 2019, 276, 103–109. [Google Scholar] [CrossRef]
- Chu, F.-J.; Wan, T.-J.; Pai, T.-Y.; Lin, H.-W.; Liu, S.-H.; Huang, C.-F. Use of magnetic fields and nitrate concentration to optimize the growth and lipid yield of Nannochloropsis oculata. J. Environ. Manag. 2020, 253, 109680. [Google Scholar] [CrossRef]
- Dębowski, M.; Zieliński, M.; Kisielewska, M.; Hajduk, A. Effect of constant magnetic field on anaerobic digestion of algal biomass. Environ. Technol. 2016, 37, 1656–1663. [Google Scholar] [CrossRef]
- Huo, S.; Chen, X.; Zhu, F.; Zhang, W.; Chen, D.; Jin, N.; Cobb, K.; Cheng, Y.; Wang, L.; Ruan, R. Magnetic field intervention on growth of the filamentous microalgae Tribonema sp. in starch wastewater for algal biomass production and nutrients removal: Influence of ambient temperature and operational strategy. Bioresour. Technol. 2020, 303, 122884. [Google Scholar] [CrossRef]
- Sirisha, K.; Suganya, B.; Sivasubramanian, V.; Bs, V.; Swaminathan, D.; Babu, A.; Meyyappan, N. Studies on the effect of pulsed magnetic field on the productivity of algae grown in dye industry effluent. J. Appl. Biotechnol. Bioeng. 2017, 3, 409–413. [Google Scholar]
- Sambusiti, C.; Bellucci, M.; Zabaniotou, A.; Beneduce, L.; Monlau, F. Algae as promising feedstocks for fermentative biohydrogen production according to a biorefinery approach: A comprehensive review. Renew. Sustain. Energy Rev. 2015, 44, 20–36. [Google Scholar] [CrossRef]
- Goswami, R.K.; Mehariya, S.; Verma, P.; Lavecchia, R.; Zuorro, A. Microalgae-based biorefineries for sustainable resource recovery from wastewater. J. Water Process Eng. 2021, 40, 101747. [Google Scholar] [CrossRef]
- Jalilian, N.; Najafpour, G.D.; Khajouei, M. Macro and micro algae in pollution control and biofuel production—A review. ChemBioEng Rev. 2020, 7, 18–33. [Google Scholar] [CrossRef]
- Vieira, M.V.; Pastrana, L.M.; Fuciños, P. Microalgae encapsulation systems for food, pharmaceutical and cosmetics applications. Mar. Drugs 2020, 18, 644. [Google Scholar] [CrossRef]
- Santos, L.O.; Silva, P.G.P.; Costa, S.S.; Machado, T.B. Magnetic Field Application to Increase Yield of Microalgal Biomass in Biofuel Production. In Biotechnological Applications of Biomass; IntechOpen: London, UK, 2020. [Google Scholar]
- Alam, M.A.; Xu, J.-L.; Wang, Z. Microalgae Biotechnology for Food, Health and High Value Products; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Esteves, A.F.; Soares, O.S.; Vilar, V.J.; Pires, J.C.; Gonçalves, A.L. The effect of light wavelength on CO2 capture, biomass production and nutrient uptake by green microalgae: A step forward on process integration and optimisation. Energies 2020, 13, 333. [Google Scholar] [CrossRef] [Green Version]
- Menegazzo, M.L.; Fonseca, G.G. Biomass recovery and lipid extraction processes for microalgae biofuels production: A review. Renew. Sustain. Energy Rev. 2019, 107, 87–107. [Google Scholar] [CrossRef]
- Debnath, C.; Bandyopadhyay, T.K.; Bhunia, B.; Mishra, U.; Narayanasamy, S.; Muthuraj, M. Microalgae: Sustainable resource of carbohydrates in third-generation biofuel production. Renew. Sustain. Energy Rev. 2021, 150, 111464. [Google Scholar] [CrossRef]
- de Carvalho Silvello, M.A.; Gonçalves, I.S.; Azambuja, S.P.H.; Costa, S.S.; Silva, P.G.P.; Santos, L.O.; Goldbeck, R. Microalgae-based carbohydrates: A green innovative source of bioenergy. Bioresour. Technol. 2022, 344, 126304. [Google Scholar] [CrossRef]
- Daneshvar, E.; Ok, Y.S.; Tavakoli, S.; Sarkar, B.; Shaheen, S.M.; Hong, H.; Luo, Y.; Rinklebe, J.; Song, H.; Bhatnagar, A. Insights into upstream processing of microalgae: A review. Bioresour. Technol. 2021, 329, 124870. [Google Scholar] [CrossRef]
- Berlot, M.; Rehar, T.; Fefer, D.; Berovic, M. The influence of treatment of Saccharomyces cerevisiae inoculum with a magnetic field on subsequent grape must fermentation. Chem. Biochem. Eng. Q. 2013, 27, 423–429. [Google Scholar]
- Deutmeyer, A.; Raman, R.; Murphy, P.; Pandey, S. Effect of magnetic field on the fermentation kinetics of Saccharomyces cerevisiae. Adv. Biosci. Biotechnol. 2011, 2011, 6857. [Google Scholar]
- Santos, L.O.; Alegre, R.M.; Garcia-Diego, C.; Cuellar, J. Effects of magnetic fields on biomass and glutathione production by the yeast Saccharomyces cerevisiae. Process Biochem. 2010, 45, 1362–1367. [Google Scholar] [CrossRef]
- da Silva, P.G.P.; Júnior, D.P.; Sala, L.; de Medeiros Burkert, J.F.; Santos, L.O. Magnetic field as a trigger of carotenoid production by Phaffia rhodozyma. Process Biochem. 2020, 98, 131–138. [Google Scholar] [CrossRef]
- Tan, L.; Shao, Y.; Mu, G.; Ning, S.; Shi, S. Enhanced azo dye biodegradation performance and halotolerance of Candida tropicalis SYF-1 by static magnetic field (SMF). Bioresour. Technol. 2020, 295, 122283. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, K.; Wang, L.; Gao, M. Extremely low-frequency magnetic fields affect pigment production of Monascus purpureus in liquid-state fermentation. Eur. Food Res. Technol. 2014, 238, 157–162. [Google Scholar] [CrossRef]
- Gao, M.; Zhang, J.; Feng, H. Extremely low frequency magnetic field effects on metabolite of Aspergillus niger. Bioelectromagnetics 2011, 32, 73–78. [Google Scholar] [CrossRef]
- Canli, O.; Kurbanoglu, E.B. Application of low magnetic field on inulinase production by Geotrichum candidum under solid state fermentation using leek as substrate. Toxicol. Ind. Health 2012, 28, 894–900. [Google Scholar] [CrossRef]
- Menestrino, B.d.C.; Pintos, T.H.C.; Sala, L.; Costa, J.A.V.; Santos, L.O. Application of static magnetic fields on the mixotrophic culture of Chlorella minutissima for carbohydrate production. Appl. Biochem. Biotechnol. 2020, 192, 822–830. [Google Scholar] [CrossRef]
- Zhang, X.; Yarema, K.; Xu, A. Impact of static magnetic field (SMF) on microorganisms, plants and animals. Biol. Eff. Static Magn. Fields 2017, 133–172. [Google Scholar]
- Gómez Luna, L.; Álvarez, I.; Rivero, R. Culture of Chlorella vulgaris using soy waste and applying a magnetic field Cultivo de C. vulgaris con soja y CM. Rev. Colomb. Biotecnol. 2011, 13, 27–38. [Google Scholar]
- da Costa Menestrino, B.; Sala, L.; Costa, J.A.V.; Buffon, J.G.; Santos, L.O. Magnetic fields exhibit a positive impact on lipid and biomass yield during phototrophic cultivation of Spirulina sp. Bioprocess Biosyst. Eng. 2021, 44, 2087–2097. [Google Scholar] [CrossRef] [PubMed]
- Deamici, K.M.; Cardias, B.B.; Costa, J.A.V.; Santos, L.O. Static magnetic fields in culture of Chlorella fusca: Bioeffects on growth and biomass composition. Process Biochem. 2016, 51, 912–916. [Google Scholar] [CrossRef]
- Sarraf, M.; Kataria, S.; Taimourya, H.; Santos, L.O.; Menegatti, R.D.; Jain, M.; Ihtisham, M.; Liu, S. Magnetic field (MF) applications in plants: An overview. Plants 2020, 9, 1139. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, W.W.C.; Costa, R.M.P.B.; e Fernandes, T.d.S.; Porto, A.L.F. Evidences of the static magnetic field influence on cellular systems. Prog. Biophys. Mol. Biol. 2016, 121, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Hirano, M.; Ohta, A.; Abe, K. Magnetic field effects on photosynthesis and growth of the cyanobacterium Spirulina platensis. J. Ferment. Bioeng. 1998, 86, 313–316. [Google Scholar] [CrossRef]
- Wang, H.Y.; Zeng, X.B.; Guo, S.Y.; Li, Z.T. Effects of magnetic field on the antioxidant defense system of recirculation-cultured Chlorella vulgaris. Bioelectromagn. J. Bioelectromagn. Soc. Soc. Phys. Regul. Biol. Med. Eur. Bioelectromagn. Assoc. 2008, 29, 39–46. [Google Scholar] [CrossRef]
- Baldev, E.; MubarakAli, D.; Sivasubramanian, V.; Pugazhendhi, A.; Thajuddin, N. Unveiling the induced lipid production in Chlorella vulgaris under pulsed magnetic field treatment. Chemosphere 2021, 279, 130673. [Google Scholar] [CrossRef]
- Yamaoka, Y.; Takimura, O.; Fuse, H.; Kaminura, K. Effect of magnetism on growth of Dunaliella salina. Res. Photosynth. 1992, 3, 87–90. [Google Scholar]
- Oliveira, M. Magnetic stimulation on the growth of the microalga Nannochloropsis oculata. In Electronic Thesis and Dissertation Repository; The University of Western Ontario: London, ON, Canada, 2017. [Google Scholar]
- Li, Z.-Y.; Guo, S.-Y.; Li, L.; Cai, M.-Y. Effects of electromagnetic field on the batch cultivation and nutritional composition of Spirulina platensis in an air-lift photobioreactor. Bioresour. Technol. 2007, 98, 700–705. [Google Scholar] [CrossRef]
- Shao, W.; Ebaid, R.; Abomohra, A.E.-F.; Shahen, M. Enhancement of Spirulina biomass production and cadmium biosorption using combined static magnetic field. Bioresour. Technol. 2018, 265, 163–169. [Google Scholar] [CrossRef]
- Deamici, K.M.; Cuellar-Bermudez, S.P.; Muylaert, K.; Santos, L.O.; Costa, J.A.V. Quantum yield alterations due to the static magnetic fields action on Arthrospira platensis SAG 21.99: Evaluation of photosystem activity. Bioresour. Technol. 2019, 292, 121945. [Google Scholar] [CrossRef] [PubMed]
- Schlesinger, D.; Molot, L.; Shuter, B. Specific growth rates of freshwater algae in relation to cell size and light intensity. Can. J. Fish. Aquat. Sci. 1981, 38, 1052–1058. [Google Scholar] [CrossRef]
- Kagami, M.; Urabe, J. Phytoplankton growth rate as a function of cell size: An experimental test in Lake Biwa. Limnology 2001, 2, 111–117. [Google Scholar]
- Young, A.; Frank, H. Energy transfer reactions involving carotenoids: Quenching of chlorophyll fluorescence. J. Photochem. Photobiol. B Biol. 1996, 36, 3–15. [Google Scholar] [CrossRef]
- Coballase-Urrutia, E.; Navarro, L.; Ortiz, J.; Verdugo-Díaz, L.; Gallardo, J.; Hernández, M.E.; Estrada-Rojo, F. Static magnetic fields modulate the response of different oxidative stress markers in a restraint stress model animal. BioMed Res. Int. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Kthiri, A.; Hidouri, S.; Wiem, T.; Jeridi, R.; Sheehan, D.; Landouls, A. Biochemical and biomolecular effects induced by a static magnetic field in Saccharomyces cerevisiae: Evidence for oxidative stress. PLoS ONE 2019, 14, e0209843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, L.O.; Silva, P.G.P.; Machado, B.R.; Sala, L.; Deamici, K.M. Update on the application of magnetic fields to microalgal cultures. World J. Microbiol. Biotechnol. 2022, 38, 211. [Google Scholar] [CrossRef]
- Pérez, A.T.E.; Camargo, M.; Rincón, P.C.N.; Marchant, M.A. Key challenges and requirements for sustainable and industrialized biorefinery supply chain design and management: A bibliographic analysis. Renew. Sustain. Energy Rev. 2017, 69, 350–359. [Google Scholar] [CrossRef]
- Pessôa, L.C.; Deamici, K.M.; Pontes, L.A.M.; Druzian, J.I.; de Jesus Assis, D. Technological prospection of microalgae-based biorefinery approach for effluent treatment. Algal Res. 2021, 60, 102504. [Google Scholar] [CrossRef]
- Wang, Y.; Ho, S.-H.; Cheng, C.-L.; Guo, W.-Q.; Nagarajan, D.; Ren, N.-Q.; Lee, D.-J.; Chang, J.-S. Perspectives on the feasibility of using microalgae for industrial wastewater treatment. Bioresour. Technol. 2016, 222, 485–497. [Google Scholar] [CrossRef]
- Ferreira, G.F.; Rios Pinto, L.F.; Carvalho, P.O.; Coelho, M.B.; Eberlin, M.N.; Maciel Filho, R.; Fregolente, L.V. Biomass and lipid characterization of microalgae genera Botryococcus, Chlorella, and Desmodesmus aiming high-value fatty acid production. Biomass Convers. Biorefinery 2021, 11, 1675–1689. [Google Scholar] [CrossRef]
- Suganya, T.; Varman, M.; Masjuki, H.; Renganathan, S. Macroalgae and microalgae as a potential source for commercial applications along with biofuels production: A biorefinery approach. Renew. Sustain. Energy Rev. 2016, 55, 909–941. [Google Scholar] [CrossRef]
- UNEP. Emissions Gap Report 2021: The Heat Is on—A World of Climate Promises Not Yet Delivered; United Nations Environment Programme: Nairobi, Kenya, 2021. [Google Scholar]
- Lu, W.; Alam, M.A.; Luo, W.; Asmatulu, E. Integrating Spirulina platensis cultivation and aerobic composting exhaust for carbon mitigation and biomass production. Bioresour. Technol. 2019, 271, 59–65. [Google Scholar] [CrossRef]
- Song, C.; Liu, Q.; Qi, Y.; Chen, G.; Song, Y.; Kansha, Y.; Kitamura, Y. Absorption-microalgae hybrid CO2 capture and biotransformation strategy—A review. Int. J. Greenh. Gas Control 2019, 88, 109–117. [Google Scholar] [CrossRef]
- Daneshvar, E.; Wicker, R.J.; Show, P.-L.; Bhatnagar, A. Biologically-mediated carbon capture and utilization by microalgae towards sustainable CO2 biofixation and biomass valorization—A review. Chem. Eng. J. 2022, 427, 130884. [Google Scholar] [CrossRef]
- de Morais, M.G.; de Morais, E.G.; Duarte, J.H.; Deamici, K.M.; Mitchell, B.G.; Costa, J.A.V. Biological CO2 mitigation by microalgae: Technological trends, future prospects and challenges. World J. Microbiol. Biotechnol. 2019, 35, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Razzak, S.A.; Ali, S.A.M.; Hossain, M.M.; deLasa, H. Biological CO2 fixation with production of microalgae in wastewater—A review. Renew. Sustain. Energy Rev. 2017, 76, 379–390. [Google Scholar] [CrossRef]
- Choudhary, M.; Peter, C.; Shukla, S.K.; Govender, P.P.; Joshi, G.M.; Wang, R. Environmental issues: A challenge for wastewater treatment. In Green Materials for Wastewater Treatment; Springer: Berlin/Heidelberg, Germany, 2020; pp. 1–12. [Google Scholar]
- Rashid, N.; Rehman, M.S.U.; Han, J.-I. Recycling and reuse of spent microalgal biomass for sustainable biofuels. Biochem. Eng. J. 2013, 75, 101–107. [Google Scholar] [CrossRef]
- Liu, X.; Saydah, B.; Eranki, P.; Colosi, L.M.; Mitchell, B.G.; Rhodes, J.; Clarens, A.F. Pilot-scale data provide enhanced estimates of the life cycle energy and emissions profile of algae biofuels produced via hydrothermal liquefaction. Bioresour. Technol. 2013, 148, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Renuka, N.; Sood, A.; Prasanna, R.; Ahluwalia, A. Phycoremediation of wastewaters: A synergistic approach using microalgae for bioremediation and biomass generation. Int. J. Environ. Sci. Technol. 2015, 12, 1443–1460. [Google Scholar] [CrossRef] [Green Version]
- Nagarajan, D.; Lee, D.-J.; Chen, C.-Y.; Chang, J.-S. Resource recovery from wastewaters using microalgae-based approaches: A circular bioeconomy perspective. Bioresour. Technol. 2020, 302, 122817. [Google Scholar] [CrossRef]
- Das, P.; AbdulQuadir, M.; Thaher, M.; Khan, S.; Chaudhary, A.K.; Alghasal, G.; Al-Jabri, H.M.S. Microalgal bioremediation of petroleum-derived low salinity and low pH produced water. J. Appl. Phycol. 2019, 31, 435–444. [Google Scholar] [CrossRef]
- Chavan, R.; Mutnuri, S. Tertiary treatment of domestic wastewater by Spirulina platensis integrated with microalgal biorefinery. Biofuels 2019, 10, 33–44. [Google Scholar] [CrossRef]
- Ammar, S.H.; Khadim, H.J.; Mohamed, A.I. Cultivation of Nannochloropsis oculata and Isochrysis galbana microalgae in produced water for bioremediation and biomass production. Environ. Technol. Innov. 2018, 10, 132–142. [Google Scholar] [CrossRef]
- Shabana, E.; Senousy, H.; Khourshid, E. Pharmaceutical wastewater treatment using free and immobilized Cyanobacteria. Egypt. J. Phycol. 2019, 20, 123–154. [Google Scholar] [CrossRef]
- Acién, F.G.; Gómez-Serrano, C.; Morales-Amaral, M.d.M.; Fernández-Sevilla, J.M.; Molina-Grima, E. Wastewater treatment using microalgae: How realistic a contribution might it be to significant urban wastewater treatment? Appl. Microbiol. Biotechnol. 2016, 100, 9013–9022. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, J.; Kwan, T.A.; Zimmerman, J.B.; Peccia, J. Ammonia inhibition in oleaginous microalgae. Algal Res. 2016, 19, 123–127. [Google Scholar] [CrossRef]
- Brennan, L.; Owende, P. Biofuels from microalgae—A review of technologies for production, processing, and extractions of biofuels and co-products. Renew. Sustain. Energy Rev. 2010, 14, 557–577. [Google Scholar] [CrossRef]
- Craggs, R.; Heubeck, S.; Lundquist, T.; Benemann, J. Algal biofuels from wastewater treatment high rate algal ponds. Water Sci. Technol. 2011, 63, 660–665. [Google Scholar] [CrossRef] [PubMed]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Zhang, H.; Zhang, J. The influence of a static magnetic field on a Chlorella vulgaris-Bacillus licheniformis consortium and its sewage treatment effect. J. Environ. Manag. 2021, 295, 112969. [Google Scholar] [CrossRef]
- Ryu, B.-G.; Kim, E.J.; Kim, H.-S.; Kim, J.; Choi, Y.-E.; Yang, J.-W. Simultaneous treatment of municipal wastewater and biodiesel production by cultivation of Chlorella vulgaris with indigenous wastewater bacteria. Biotechnol. Bioprocess Eng. 2014, 19, 201–210. [Google Scholar] [CrossRef]
- Woertz, I.; Feffer, A.; Lundquist, T.; Nelson, Y. Algae grown on dairy and municipal wastewater for simultaneous nutrient removal and lipid production for biofuel feedstock. J. Environ. Eng. 2009, 135, 1115. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Luo, S.; Fan, X.; Yang, Z.; Guo, R. Biomass and lipid production of marine microalgae using municipal wastewater and high concentration of CO2. Appl. Energy 2011, 88, 3336–3341. [Google Scholar] [CrossRef]
- Feng, X.; Chen, Y.; Lv, J.; Han, S.; Tu, R.; Zhou, X.; Jin, W.; Ren, N. Enhanced lipid production by Chlorella pyrenoidosa through magnetic field pretreatment of wastewater and treatment of microalgae-wastewater culture solution: Magnetic field treatment modes and conditions. Bioresour. Technol. 2020, 306, 123102. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.A.V.; Freitas, B.C.B.; Cruz, C.G.; Silveira, J.; Morais, M.G. Potential of microalgae as biopesticides to contribute to sustainable agriculture and environmental development. J. Environ. Sci. Health Part B 2019, 54, 366–375. [Google Scholar] [CrossRef]
- Bhattacharjee, R.; Dey, U. Biofertilizer, a way towards organic agriculture: A review. Afr. J. Microbiol. Res. 2014, 8, 2332–2343. [Google Scholar]
- Thirumurthy, P.; Mol, I. Microalgae as bio-pesticides for the development of sustainable agriculture. Wide Spectr. 2020, 6, 5–22. [Google Scholar]
- Costa, J.A.V.; Cassuriaga, A.P.A.; Moraes, L.; de Morais, M.G. Biosynthesis and potential applications of terpenes produced from microalgae. Bioresour. Technol. Rep. 2022, 101166. [Google Scholar] [CrossRef]
- Stirk, W.; Bálint, P.; Tarkowská, D.; Novák, O.; Maróti, G.; Ljung, K.; Turečková, V.; Strnad, M.; Ördög, V.; Van Staden, J. Effect of light on growth and endogenous hormones in Chlorella minutissima (Trebouxiophyceae). Plant Physiol. Biochem. 2014, 79, 66–76. [Google Scholar] [CrossRef] [PubMed]
- GMI (Global Market Insights). Algae Oil Market Size, Regional Outlook, Application Growth Potential, COVID-19 Impact Analysis, Competitive Market Growth & Forecast, 2022–2028; Global Market Insights Inc.: Selbyville, DE, USA, 2022. [Google Scholar]
- Qari, H.; Rehan, M.; Nizami, A.-S. Key issues in microalgae biofuels: A short review. Energy Procedia 2017, 142, 898–903. [Google Scholar] [CrossRef]
- Dineshbabu, G.; Goswami, G.; Kumar, R.; Sinha, A.; Das, D. Microalgae—Nutritious, sustainable aqua-and animal feed source. J. Funct. Foods 2019, 62, 103545. [Google Scholar] [CrossRef]
- The State of World Fisheries and Aquaculture 2018—Meeting the Sustainable Development Goals; The State of The World series of the Food and Agriculture Organization of the United Nations. Aquaculture; FAO: Rome, Italy, 2018; Volume 35.
- Hunter, M.C.; Smith, R.G.; Schipanski, M.E.; Atwood, L.W.; Mortensen, D.A. Agriculture in 2050: Recalibrating targets for sustainable intensification. Bioscience 2017, 67, 386–391. [Google Scholar] [CrossRef]
- Hasan, M.; Soto, D. Improving Feed Conversion Ratio and Its Impact on Reducing Greenhouse Gas Emissions in Aquaculture; FAO: Rome, Italy, 2017. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microalga Specie | Type of Microalgae | MF Intensity and Exposure Time | Cultivation Time (Days) | Positive Effect | Reference |
|---|---|---|---|---|---|
| Scenedesmus obliquus | Green algae | 50 mT for 1 h/d | 6 | 11.5% chlorophyll a content | [4] |
| Spirulina sp. | Blue-green algae | 60 mT for 24 h/d | 15 | 57.2% phycocyanin content | [13] |
| Chlorella kessleri | Green algae | 30 mT for 24 h/d | 10 | 25% carotenoid content | [8] |
| Chlorella vulgaris | Green algae | 30 mT for 1 h/d | 7 | 84% protein content | [43] |
| Spirulina sp. | Blue-green algae | 60 mT for 24 h/d | 15 | 16.7% protein content | [13] |
| Tribonema sp. | Yellow-green algae | 30 mT for 24 h/d | 9 | 85.4% protein content | [20] |
| Chlorella kessleri | Green algae | 10 mT for 24 h/d | 12 | 47% lipid content | [12] |
| Chlorella kessleri | Green algae | 30 mT for 24 h/d | 10 | 13.7% lipid content | [8] |
| Haematococcus pluvialis | Red algae | 16.9 mT for 24 h/d | 7 | 25% lipid content | [18] |
| Chlorella homosphaera | Green algae | 15 mT for 1 h/d | 15 | 22.4% lipid content | [10] |
| Spirulina sp. | Blue-green algae | 30 mT for 24 h/d | 15 | 45.5% lipid content | [44] |
| Chlorella kessleri | Green algae | 10 mT for 24 h/d | 12 | 8.5% carbohydrate content | [12] |
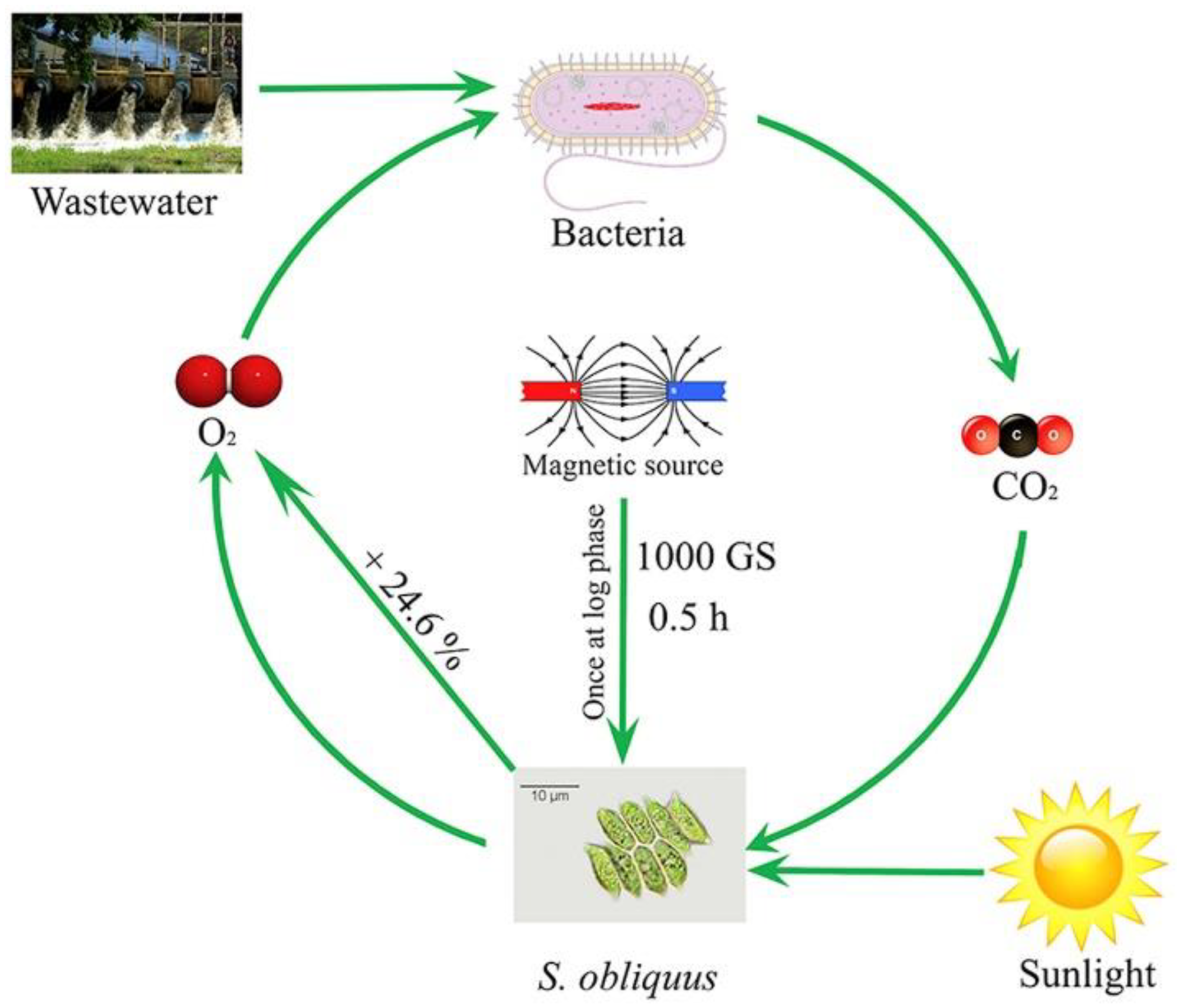
| Chlorella fusca | Green algae | 60 mT for 24 h/d | 15 | 24.6% carbohydrate content | [45] |
| Chlorella minutissima | Green algae | 30 mT for 24 h/d | 12 | 162.9% carbohydrate content | [41] |
| Chlorella fusca LEB 111 | Green algae | 25 mT for 1 h/d | 15 | 10.6% carbohydrate content | [15] |
| Microalga Species | Intensity/Exposure Time | Growth Rate Increase (↑)/Decrease (↓) | Biomass Increase (↑)/Decrease (↓) | Reference |
|---|---|---|---|---|
| Chlorella fusca | 60 mT/24 h/d | ↓ 12% | ↑ 21% | [45] |
| Chlorella fusca | 60 mT/1 h/d | ↓ 14% | ↑ 13% | [17] |
| Chlorella fusca | 25 mT/24 h/d (outdoors) 1 h/d (indoors) | ↓ 18%—outdoors ↑ 100%—indoors | ↑ 31%—outdoors ↑ 19%—indoors | [16] |
| Chlorella homosphaera | 30 mT/1 h/d | 106% | ↑ 21% | [10] |
| Chlorella kessleri | 10 mT/ every 18 min | ↑ 126%—in flask ↑ 88%—in raceway pond | ↑ 77% | [12] |
| Chlorella pyrenoidosa | 500 mT/3 h/d | n.a. | ↑ 12% | [5] |
| Chlorella vulgaris | 10 mT/12 h | ↑ 72% | n.a. | [47] |
| Chlorella vulgaris | PEMF * 0.07 mT, 1 Hz/4 h/d | n.a. | ↑ 315% compared to 0.09 mT | [50] |
| Dunaliella salina | 10 mT, with 1 mg/L Fe-EDTA | n.a. | ↑ 84% | [51] |
| Nannochloropsis oculata | 10 mT/n.a. | n.a. | ↑ 43% | [52] |
| Spirulina platensis | 10 mT/n.a. | n.a. | ↑ 10% | [48] |
| Spirulina platensis | 250 mT/n.a. | n.a. | ↑ 47% | [53] |
| Spirulina platensis | 30 mT/ 3, 6 and 12 h/d | n.a. | ↑ 16% (the highest for 6 h/d) | [54] |
| Spirulina platensis | 30 mT/1 h/d | n.a. | ↑ 49% | [55] |
| Spirulina sp. | 60 mT/1 h/d | ↑ 48% | ↑ 95% | [13] |
| Spirulina sp. | 25 mT/24 h/d | the same for outdoors and indoors as in control | ↑ 6%—outdoors ↓ 6%—indoors | [11] |
| Spirulina sp. | 60 mT, 1.9 g/L NaNO3 24 h/d | ↑ 17% | ↑ 37% | [14] |
| Microalga Species | Intensity/Exposure Time | Effect of MF vs. Control | Reference |
|---|---|---|---|
| Carbohydrates | |||
| Chlorella fusca | 30, 60 mT/1, 24 h/d | 31 vs. 25% w/w | [45] |
| Chlorella fusca | 30, 60 mT/1, 24 h/d | 25 vs. 17% w/w | [17] |
| Chlorella fusca | 25 mT/1, 24 h/d | 34 vs. 31% w/w—outdoors 28 vs. 25% w/w—indoors | [15] |
| Chlorella homosphaera | 15 mT/1 h/d 30, 60 mT/1, 24 h/d | 45 vs. 47% w/w | [10] |
| Chlorella kessleri | 10 mT/every 18 min | 42 vs 39% w/w | [12] |
| Chlorella kessleri | 30, 60 mT/1, 24 h/d | 21 vs. 19% w/w | [8] |
| Spirulina platensis | 30 mT/3, 6, 12 h/d | 13 vs. 10% c.d.w. | [54] |
| Spirulina platensis | 30 mT/1, 24 h/d | 16 vs. 13% w/w | [55] |
| Spirulina sp. | 30, 60 mT/1, 24 h/d | 30 vs. 13% w/w | [13] |
| Spirulina sp. | 30, 60 mT/24 h/d | 7–15 vs. 12–22% w/w | [14] |
| Pigments: Chlorophyll a | |||
| Chlorella kessleri | 10 mT/every 18 min | 2.9 vs. 2.5% w/w | [12] |
| Spirulina platensis | 250 mT/n.a. | 13.5 vs. 10.3 g/L | [53] |
| Spirulina platensis | 30 mT/1, 24 h/d | 2.6 vs. 2.2% w/w | [55] |
| Nannochloropsis oculata | 5, 10, 15 mT/n.a. | 1.9 to 2.0-fold increase at 5 and 10 mT, respectively; no change at 15 mT | [52] |
| Pigments: Chlorophyll b | |||
| Chlorella kessleri | 10 mT/every 18 min | 0.87 vs. 0.53% w/w | [12] |
| Pigments: Carotenoids | |||
| Chlorella kessleri | 10 mT /every 18 min | 0.08 vs. 0.07% w/w | [12] |
| Pigments: Phycocyanin | |||
| Spirulina platensis | 30 mT/1, 24 h/d | 11.5 vs. 12.6% w/w | [45] |
| Spirulina sp. LEB 18 | 30, 60 mT/1, 24 h/d | 13.2 vs. 8.4 g/L | [13] |
| Spirulina platensis | 10 mT/n.a. | increase by 54% vs. control | [48] |
| Protein | |||
| Chlorella fusca | 30, 60 mT/1, 24 h/d | 64 vs. 60% w/w | [45] |
| Chlorella fusca | 30, 60 mT/1, 24 h/d | 56 vs. 52% w/w | [17] |
| Chlorella fusca | 25 mT/1, 24 h/d | 33 vs. 29% w/w—outdoors 36 vs. 28% w/w—indoors | [15] |
| Chlorella homosphaera | 15 mT/1 h/d 30, 60 mT/1, 24 h/d | 23 vs. 18% w/w | [10] |
| Chlorella kessleri | 10 mT/every 18 min | 32 vs. 30% w/w | [12] |
| Chlorella kessleri | 30, 60 mT/1, 24 h/d | 59 vs. 54% w/w | [8] |
| Spirulina platensis | 30 mT/3, 6, 12 h/d | 65 vs. 59% c.d.w. | [54] |
| Spirulina platensis | 30 mT/1, 24 h/d | 67 vs. 67% w/w | [55] |
| Spirulina sp. | 30, 60 mT/1, 24 h/d | 73 vs. 63% w/w | [13] |
| Spirulina sp. | 30, 60 mT/24 h/d | 66–73 vs. 58–71% w/w | [14] |
| Lipids | |||
| Chlorella fusca | 30, 60 mT/1, 24 h/d | 13 vs. 13% w/w | [45] |
| Chlorella fusca | 30, 60 mT/1, 24 h/d | 19 vs. 18% w/w | [17] |
| Chlorella fusca | 25 mT/1, 24 h/d | 32 vs. 30% w/w—outdoors 36 vs. 34% w/w—indoors | [15] |
| Chlorella homosphaera | 15 mT/1 h/d 30, 60 mT/1, 24 h/d | 44 vs. 29% w/w | [10] |
| Chlorella kessleri | 10 mT/every 18 min | 20 vs. 25% w/w | [12] |
| Chlorella kessleri | 30, 60 mT/1, 24 h/d | 24 vs. 21% w/w | [8] |
| Chlorella pyrenoidosa | 500 mT/3 h/d | 216 vs. 212 mg/g c.d.w. | [5] |
| Chlorella vulgaris | PEMF * 0.06, 0.07, 0.08, 0.09 mT at 1 Hz/4 h/d | 55 d.w. vs. 49% d.w. | [50] |
| Spirulina platensis | 30 mT/3, 6, 12 h/d | 18 vs. 16% c.d.w. | [54] |
| Spirulina sp. | 30, 60 mT/1, 24 h/d | 10 vs. 8% w/w | [13] |
| Spirulina sp. | 30, 60 mT/24 h/d | 9–15 vs. 11–17% w/w | [14] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deamici, K.M.; Dziergowska, K.; Silva, P.G.P.; Michalak, I.; Santos, L.O.; Detyna, J.; Kataria, S.; Brestic, M.; Sarraf, M.; Islam, M. Microalgae Cultivated under Magnetic Field Action: Insights of an Environmentally Sustainable Approach. Sustainability 2022, 14, 13291. https://doi.org/10.3390/su142013291
Deamici KM, Dziergowska K, Silva PGP, Michalak I, Santos LO, Detyna J, Kataria S, Brestic M, Sarraf M, Islam M. Microalgae Cultivated under Magnetic Field Action: Insights of an Environmentally Sustainable Approach. Sustainability. 2022; 14(20):13291. https://doi.org/10.3390/su142013291
Chicago/Turabian StyleDeamici, Kricelle Mosquera, Katarzyna Dziergowska, Pedro Garcia Pereira Silva, Izabela Michalak, Lucielen Oliveira Santos, Jerzy Detyna, Sunita Kataria, Marian Brestic, Mohammad Sarraf, and Monirul Islam. 2022. "Microalgae Cultivated under Magnetic Field Action: Insights of an Environmentally Sustainable Approach" Sustainability 14, no. 20: 13291. https://doi.org/10.3390/su142013291