Printed Paper Waste as an Alternative Growing Medium Component to Produce Brassica Seedlings under Nursery Conditions

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Paper Waste
2.2. Growing Media Properties
2.3. Seed Emergence
2.4. Plant Growth and Tissue Analysis
2.5. Total Phenolics and Antioxidant Activity
2.6. Lipid Peroxidation, Hydrogen Peroxide, and Antioxidant Enzyme Activity
2.7. Statistical Analysis
3. Results and Discussion
3.1. Growing Media Characteristics
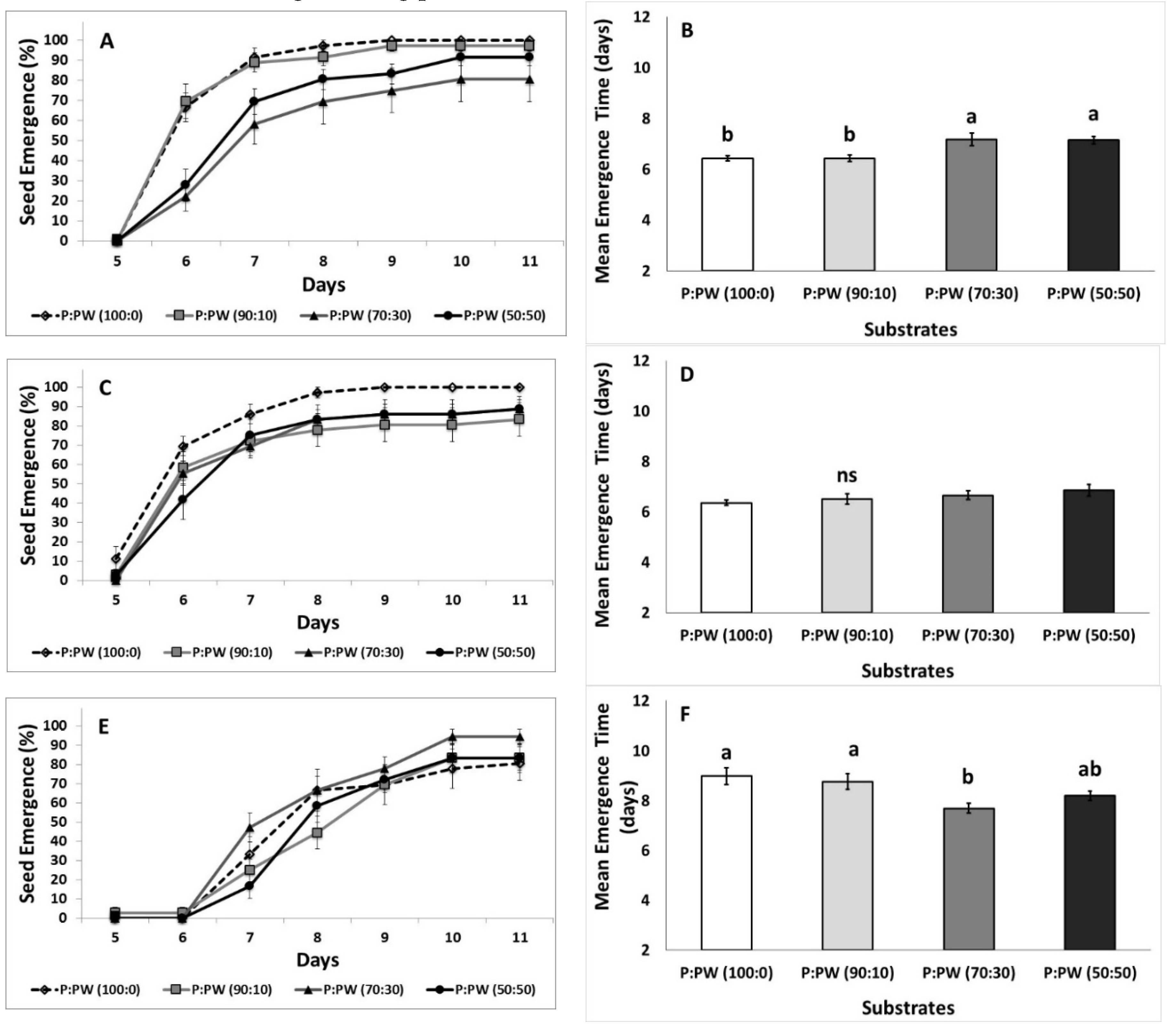
3.2. Seed Emergence
3.3. Plant Growth and Physiology
3.4. Plant Minerals
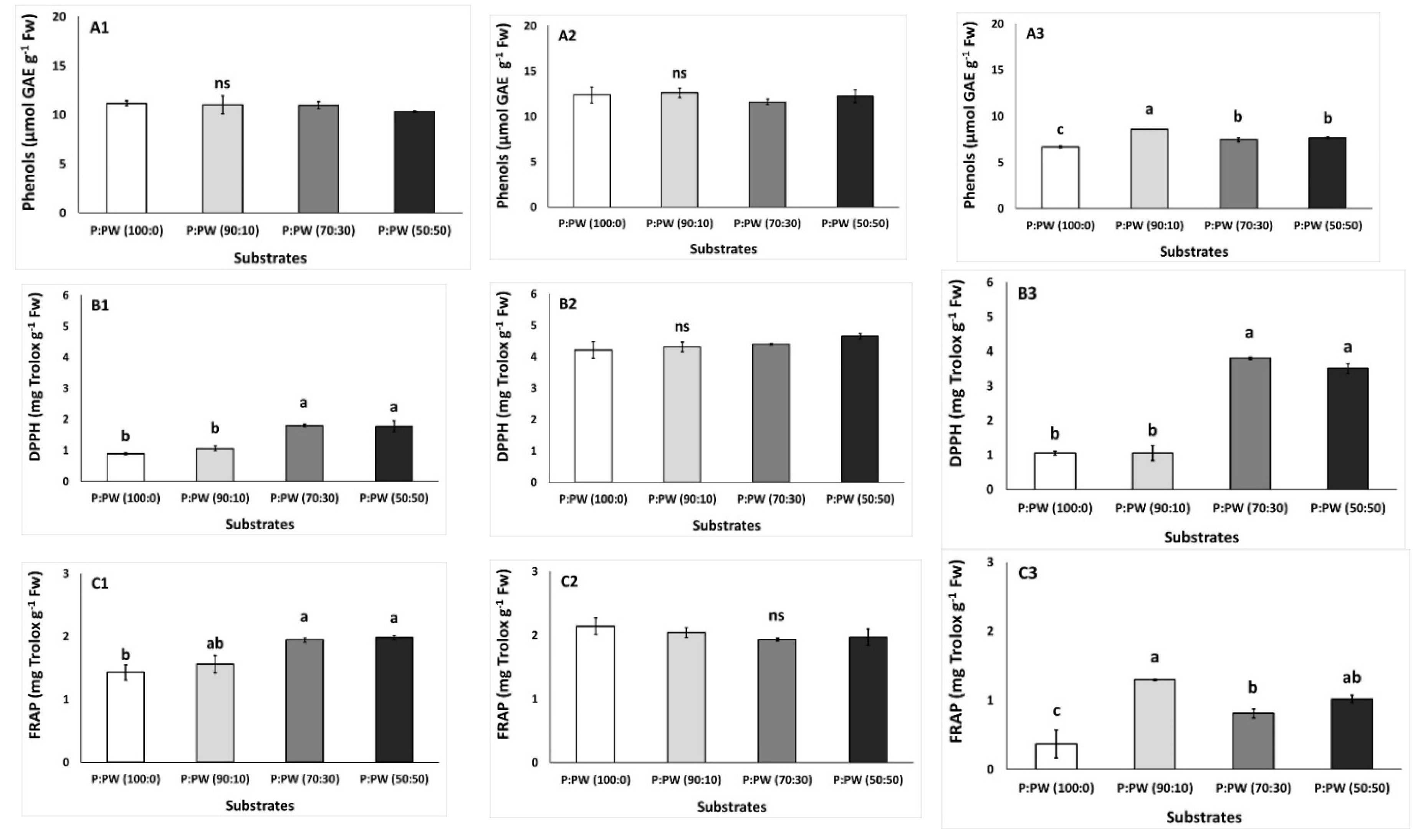
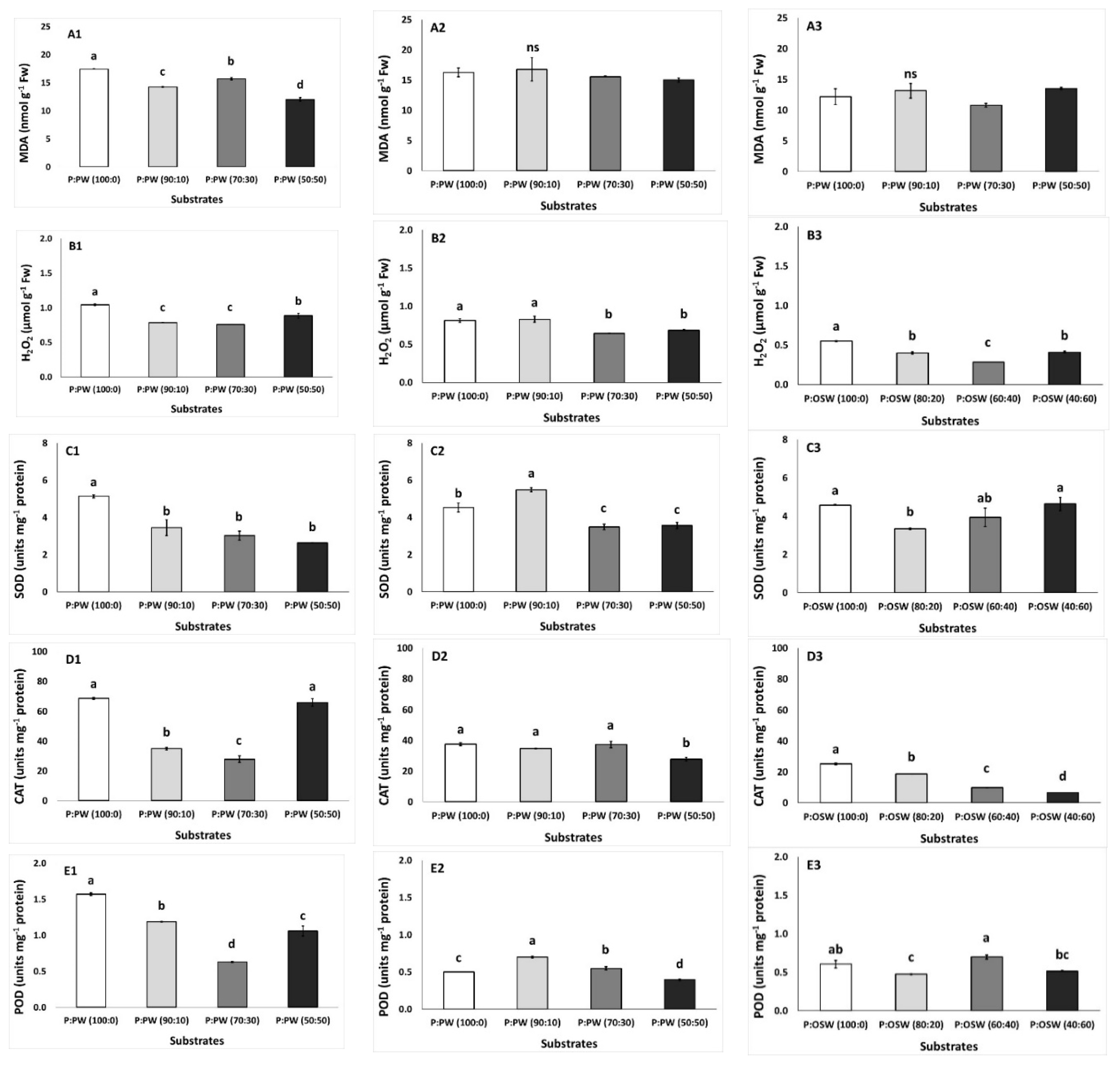
3.5. Plant Stress Factors and Antioxidants
3.6. Principal Component Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jaria, G.; Silva, C.P.; Ferreira, C.I.A.; Otero, M.; Calisto, V. Sludge from paper mill effluent treatment as raw material to produce carbon adsorbents: An alternative waste management strategy. J. Environ. Manag. 2017, 188, 203–211. [Google Scholar] [CrossRef] [Green Version]
- Rashid, M.; Barry, D.; Goss, M. Paper mill biosolids application to agricultural lands: Benefits and environmental concerns with special reference to situation in Canada. Soil Env. 2006, 25, 85–98. [Google Scholar]
- Oral, J.; Sikula, J.; Puchyr, R.; Hajny, Z.; Stehlik, P.; Bebar, L. Processing of waste from pulp and paper plant. J. Clean. Prod. 2005, 13, 509–515. [Google Scholar] [CrossRef]
- Tucker, P.; Douglas, P. Composted Paper Mill Waste as a Peat Substitute; Newspaper Industry Environmental Technology Initiative; University of Paisley: Glasgow, UK, 2006. [Google Scholar]
- Das, R.K.; Brar, S.K.; Verma, M. Potential use of pulp and paper solid waste for the bio-production of fumaric acid through submerged and solid state fermentation. J. Clean. Prod. 2016, 112, 4435–4444. [Google Scholar] [CrossRef]
- Méndez, A.; Barriga, S.; Fidalgo, J.M.; Gascó, G. Adsorbent materials from paper industry waste materials and their use in Cu(II) removal from water. J. Hazard. Mater. 2009, 165, 736–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Azevedo, A.R.G.; Alexandre, J.; Marvila, M.T.; Xavier, G.; de, C.; Monteiro, S.N.; Pedroti, L.G. Technological and environmental comparative of the processing of primary sludge waste from paper industry for mortar. J. Clean. Prod. 2020, 249, 119336. [Google Scholar] [CrossRef]
- Zhang, Q.; Khan, M.U.; Lin, X.; Yi, W.; Lei, H. Green-composites produced from waste residue in pulp and paper industry: A sustainable way to manage industrial wastes. J. Clean. Prod. 2020, 262, 121251. [Google Scholar] [CrossRef]
- Joshi, G.; Naithani, S.; Varshney, V.K.; Bisht, S.S.; Rana, V. Potential use of waste paper for the synthesis of cyanoethyl cellulose: A cleaner production approach towards sustainable environment management. J. Clean. Prod. 2017, 142, 3759–3768. [Google Scholar] [CrossRef]
- Kumar, P.R.; Jayaram, A.; Somashekar, R.K. Assessment of the performance of different compost models to manage urban household organic solid wastes. Clean Technol. Environ. Policy 2009, 11, 473–484. [Google Scholar] [CrossRef]
- Ravindran, B.; Mnkeni, P.N.S. Bio-optimization of the carbon-to-nitrogen ratio for efficient vermicomposting of chicken manure and waste paper using Eisenia fetida. Environ. Sci. Pollut. Res. 2016, 23, 16965–16976. [Google Scholar] [CrossRef]
- Yilmaz, M.; Gumuskaya, T. Recycling costs: A research in the waste paper industry. Eur. J. Acc. Aud. Fin. Res. 2015, 3, 58–68. [Google Scholar]
- Caputo, A.C.; Pelagagge, P.M. Waste-to-energy plant for paper industry sludges disposal: Technical-economic study. J. Hazard. Mater. 2001, 81, 265–283. [Google Scholar] [CrossRef]
- Abbasi, S.A.; Hussain, N.; Tauseef, S.M.; Abbasi, T. A novel FLippable Units Vermireactor Train System—FLUVTS—for rapidly vermicomposting paper waste to an organic fertilizer. J. Clean. Prod. 2018, 198, 917–930. [Google Scholar] [CrossRef]
- Vieira, C.; Pinheiro, R.; Rodriguez, R.; Candido, V.; Monteiro, S. Clay bricks added with effluent sludge from paper industry. Technical, economical and environmental benefits. Appl. Clay Sci. 2016, 132–133, 753–759. [Google Scholar] [CrossRef]
- Bugbee, G. Growth of ornamental plants in container media amended with biosolids composts. Compost. Sci. Util. 2002, 10, 92–98. [Google Scholar] [CrossRef]
- Landis, T.D.; Morgan, N. Growing Media Alternatives for Forest and Native Plant Nurseries. National Proceedings: Forest and Conservation Nursery Associations-2008; Rocky Mountain Research Station-P-58; US Department of Agricultur: Broomfield, CO, USA, 2009; pp. 26–31.
- Holmes, S. Growing media developments in the UK. Acta Hortic. 2009, 819, 23–26. [Google Scholar] [CrossRef]
- Kern, J.; Tammeorg, P.; Shanskiy, M.; Sakrabani, R.; Knicker, H.; Kammann, C.; Tuhkanen, E.M.; Smidt, G.; Prasad, M.; Tiilikkala, K.; et al. Synergistic use of peat and charred material in growing media–an option to reduce the pressure on peatlands? J. Environ. Eng. Landsc. Manag. 2017, 25, 160–174. [Google Scholar] [CrossRef]
- Ceglie, F.G.; Bustamante, M.A.; Amara, M.B.; Tittarelli, F. The challenge of peat substitution in organic seedling production: Optimization of growing media formulation through mixture design and response surface analysis. PLoS ONE 2015, 10, e0128600. [Google Scholar] [CrossRef]
- Abad, M.; Noguera, P.; Burés, S. National inventory of organic wastes for use as growing media for ornamental potted plant production: Case study in Spain. Bioresour. Technol. 2001, 77, 197–200. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Antoniou, O.; Athinodorou, F.; Vassiliou, R.; Papadaki, A.; Tzortzakis, N. Deployment of olive-stone waste as a substitute growing medium component for Brassica seedling production in nurseries. Environ. Sci. Pollut. Res. 2019. [Google Scholar] [CrossRef]
- Papamichalaki, M.; Papadaki, A.; Tzortzakis, N. Substitution of peat with municipal solid waste compost in watermelon seedling production combined with fertigation. Chil. J. Agric. Res. 2014, 74, 452–459. [Google Scholar] [CrossRef] [Green Version]
- Jayasinghe, G.Y.; Tokashiki, Y.; Kitou, M.; Kinjo, K. Oil palm waste and synthetic zeolite: An alternative soil-less growth substrate for lettuce production as a waste management practice. Waste Manag. Res. 2008, 26, 559–565. [Google Scholar] [CrossRef]
- Fascella, G. Growing substrates alternative to peat for ornamental plants. In Soilless Culture—Use of Substrates for the Production of Quality Horticultural Crops; Asaduzzaman, M., Ed.; InTech: Rijeka, Croatia, 2015; ISBN 978-953-51-1739-1. [Google Scholar]
- Kelepesi, S.; Tzortzakis, N.G. Olive mill wastesA growing medium component for seedling and crop production of lettuce and chicory. Int. J. Veg. Sci. 2009, 15, 325–339. [Google Scholar] [CrossRef]
- Prasad, M.; Tzortzakis, N.; McDaniel, N. Chemical characterization of biochar and assessment of the nutrient dynamics by means of preliminary plant growth tests. J. Environ. Manag. 2018, 216, 89–95. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Antoniou, O.; Tzionis, A.; Prasad, M.; Tzortzakis, N. Alternative soilless media using olive-mill and paper waste for growing ornamental plants. Environ. Sci. Pollut. Res. 2018, 25, 35915–35927. [Google Scholar] [CrossRef] [PubMed]
- Chrysargyris, A.; Stavrinides, M.; Moustakas, K.; Tzortzakis, N. Utilization of paper waste as growing media for potted ornamental plants. Clean Technol. Environ. Policy 2019, 21, 1937–1948. [Google Scholar] [CrossRef]
- WRAP Research Analysis of the Market Potential for Lower Grade Composted Materials in the UK; WRAP: Banbury, UK, 2002.
- Tzortzakis, N.G.; Economakis, C.D. Shredded maize stems as an alternative substrate medium: Effect on growth, flowering and yield of tomato in soilless culture. J. Veg. Sci. 2005, 11, 57–70. [Google Scholar] [CrossRef]
- Chong, C. Experiences with wastes and composts in nursery substrates. Horttechnology 2005, 15, 739–747. [Google Scholar] [CrossRef]
- Wuana, R.A.; Okieimen, F.E. Heavy Metals in Contaminated Soils: A Review of Sources, Chemistry, Risks and Best Available Strategies for Remediation. ISRN Ecol. 2011, 2011, 1–20. [Google Scholar] [CrossRef] [Green Version]
- European Committee for Standardization. European standard EN 13041. Soil Improvers and Growing Media—Determination of Physical Properties—Dry Bulk Density, Air Volume, Water Volume, Shrinkage Value and Total Pore Space; European Committee for Standardization: Brussels, Belgium, 1999. [Google Scholar]
- Chrysargyris, A.; Xylia, P.; Botsaris, G.; Tzortzakis, N. Antioxidant and antibacterial activities, mineral and essential oil composition of spearmint (Mentha spicata L.) affected by the potassium levels. Ind. Crops Prod. 2017, 103, 202–212. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Papakyriakou, E.; Petropoulos, S.A.; Tzortzakis, N. The combined and single effect of salinity and copper stress on growth and quality of Mentha spicata plants. J. Hazard. Mater. 2019, 368, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Tzortzakis, N.G.; Tzanakaki, K.; Economakis, C.D.C.D. Effect of origanum oil and vinegar on the maintenance of postharvest quality of tomato. Food Nutr. Sci. 2011, 2, 974–982. [Google Scholar] [CrossRef] [Green Version]
- Loreto, F.; Velikova, V. Isoprene produced by leaves protects the photosynthetic apparatus against ozone damage, quenches ozone products, and reduces lipid peroxidation of cellular membranes. Plant Physiol. 2001, 127, 1781–1787. [Google Scholar] [CrossRef] [PubMed]
- De Azevedo Neto, A.D.; Prisco, J.T.; Enéas-Filho, J.; De Abreu, C.E.B.; Gomes-Filho, E. Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant and salt-sensitive maize genotypes. Environ. Exp. Bot. 2006, 56, 87–94. [Google Scholar] [CrossRef]
- Ribeiro, H.M.; Vasconcelos, E.; Cabral, F.; Ribeiro, D. Fertilization of Pinus pinea L. seedlings with a sewage sludge-based compost. Waste Manag. Res. 2009, 27, 112–118. [Google Scholar] [CrossRef]
- Ouzounidou, G.; Asfi, M.; Sotirakis, N.; Papadopoulou, P.; Gaitis, F. Olive mill wastewater triggered changes in physiology and nutritional quality of tomato (Lycopersicon esculentum Mill.) depending on growth substrate. J. Hazard. Mater. 2008, 158, 523–530. [Google Scholar] [CrossRef]
- Mohammadi Torkashvand, A.; Haghighat, N.; Shadparvar, V. Effect of paper mill lime sludge as an acid soil amendment. Sci. Res. Essays 2010, 5, 1302–1306. [Google Scholar]
- He, J.; Lange, C.R.; Dougherty, M. Laboratory study using paper mill lime mud for agronomic benefit. Process Saf. Environ. Prot. 2009, 87, 401–405. [Google Scholar] [CrossRef]
- Havlin, J.; Beaton, J.; Tisdale, S.; Werner, L. Soil Fertility and Fertilizers. An Introduction to Nutrient Management; Prentice Hall: Upper saddle River, NJ, USA, 2005. [Google Scholar]
- Mertoglu-Elmas, G. The effect of colorants on the content of heavy metals in recycled corrugated board papers. BioResources 2017, 12, 2690–2698. [Google Scholar] [CrossRef] [Green Version]
- Perez-Murcia, M.D.; Moral, R.; Moreno-Caselles, J.; Perez-Espinosa, A.; Paredes, C. Use of composted sewage sludge in growth media for broccoli. Bioresour. Technol. 2006, 97, 123–130. [Google Scholar] [CrossRef]
- Kiarostami, K.; Mohseni, R.; Saboora, A. Biochemical changes of Rosmarinus officinalis under salt stress. J. Stress Physiol. Biochem. 2010, 6, 114–122. [Google Scholar]
- Hara, T.; Sonoda, Y. The role of macronutrients for cabbage-head formation: I. contribution to cabbage-head formation of nitrogen, phosphorus, or potassium supplied at different growth stages. Soil Sci. Plant Nutr. 1979, 25, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Tarchoune, I.; Sgherri, C.; Izzo, R.; Lachaal, M.; Ouerghi, Z.; Navari-Izzo, F. Antioxidative responses of Ocimum basilicum to sodium chloride or sodium sulphate salinization. Plant Physiol. Biochem. 2010, 48, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Morrish, R.; Hofstede, H. Alternatives to Peat: A Manual; Murdoch University: Perth, Australia, 2000. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| P:PW (100:0) | P:PW (90:10) | P:PW (70:30) | P:PW (50:50) | |
|---|---|---|---|---|
| EC (μS cm−1) | 277.16 ± 4.50c | 405.88 ± 23.99b | 412.01 ± 17.79b | 719.07 ± 8.98a |
| pH | 4.68 ± 0.08d | 5.09 ± 0.17c | 5.70 ± 0.02b | 6.16 ± 0.03a |
| Organic matter (%) | 99.02 ± 0.29a | 97.19 ± 0.43ab | 96.41 ± 0.59b | 92.44 ± 0.80c |
| Organic C (%) | 57.44 ± 0.17a | 56.37 ± 0.25ab | 55.92 ± 0.34b | 53.62 ± 0.46c |
| C/N | 132.21 ± 0.59c | 151.10 ± 2.93b | 156.59 ± 0.65b | 171.83 ± 9.15a |
| N (g kg−1) | 4.33 ± 0.02a | 3.72 ± 0.06b | 3.61 ± 0.01b | 3.13 ± 0.14c |
| K (g kg−1) | 0.068 ± 0.002a | 0.061 ± 0.005a | 0.071 ± 0.007a | 0.079 ± 0.004a |
| P (g kg−1) | 0.072 ± 0.007a | 0.041 ± 0.001a | 0.062 ± 0.016a | 0.057 ± 0.002a |
| Ca (g kg−1) | 1.21 ± 0.02c | 1.57 ± 0.08c | 5.98 ± 0.16b | 11.71 ± 1.38a |
| Mg (mg kg−1) | 0.273 ± 0.003c | 0.414 ± 0.028a | 0.303 ± 0.013bc | 0.342 ± 0.005b |
| Na (mg kg−1) | 0.138 ± 0.009c | 0.232 ± 0.239bc | 0.274 ± 0.031b | 0.501 ± 0.043a |
| Fe (mg kg−1) | 570.80 ± 10.36c | 583.13 ± 11.59c | 667.01 ± 6.21b | 1118.25 ± 6.89a |
| Cu (mg kg−1) | 53.93 ± 6.70b | 75.62 ± 5.51ab | 75.89 ± 8.14ab | 90.51 ± 11.10a |
| Zn (mg kg−1) | 32.13 ± 1.75a | 13.46 ± 0.93c | 16.55 ± 0.91bc | 20.23 ± 1.83b |
| Mn (mg kg−1) | 30.05 ± 17.40a | 22.14 ± 7.69a | 19.55 ± 11.32a | 7.66 ± 4.42a |
| TP (%) Z | 84.87 ± 0.27a | 82.94 ± 0.28b | 77.26 ± 0.26c | 75.93 ± 0.25d |
| AWHC (%) Z | 68.42 ± 0.23a | 66.95 ± 0.18b | 65.74 ± 0.21c | 65.94 ± 0.22c |
| AFP (%) Z | 16.49 ± 0.05a | 15.98 ± 0.05b | 11.49 ± 0.04c | 9.99 ± 0.03d |
| BD (g cm−3) Z | 0.236 ± 0.003c | 0.210 ± 0.000d | 0.256 ± 0.003b | 0.273 ± 0.002a |
| Species | Substrates | Plant Height | Leaf No | Fresh Weight | Dry Matter |
|---|---|---|---|---|---|
| Cauliflower | P:PW (100:0) | 7.98 ± 0.31aY | 6.33 ± 0.21a | 5.78 ± 1.11a | 11.89 ± 0.44ab |
| P:PW (90:10) | 8.63 ± 0.19a | 5.33 ± 021b | 4.83 ± 0.73ab | 16.31 ± 3.40a | |
| P:PW (70:30) | 5.88 ± 0.26b | 4.66 ± 0.33b | 2.92 ± 0.81bc | 9.36 ± 0.37b | |
| P:PW (50:50) | 5.28 ± 0.15b | 3.66 ± 0.51c | 1.85 ± 0.48c | 11.16 ± 0.48ab | |
| Broccoli | P:PW (100:0) | 12.11 ± 0.41a | 6.66 ± 0.33a | 6.85 ± 0.58a | 10.82 ± 1.13a |
| P:PW (90:10) | 11.72 ± 0.36b | 5.83 ± 0.16b | 5.06 ± 0.34b | 12.18 ± 0.69a | |
| P:PW (70:30) | 9.96 ± 0.29c | 5.16 ± 0.17c | 3.28 ± 0.33c | 11.81 ± 0.49a | |
| P:PW (50:50) | 8.85 ± 0.21d | 4.83 ± 0.16c | 2.62 ± 0.20c | 10.94 ± 0.58a | |
| Cabbage | P:PW (100:0) | 8.08 ± 0.35a | 7.66 ± 0.21a | 6.76 ± 0.93a | 11.78 ± 1.76a |
| P:PW (90:10) | 7.51 ± 0.28ab | 6.66 ± 0.21b | 4.65 ± 0.23b | 10.16 ± 0.29a | |
| P:PW (70:30) | 6.81 ± 0.23bc | 5.67 ± 0.20c | 2.23 ± 0.34c | 9.45 ± 1.50a | |
| P:PW (50:50) | 6.20 ± 0.26c | 5.02 ± 0.45c | 2.03 ± 0.46c | 8.58 ± 0.81a |
| Species | Substrates | Chl. Fluorescence | SPAD | Stomatal Conductance | Chl a | Chl b | Total Chl |
|---|---|---|---|---|---|---|---|
| Cauliflower | P:PW (100:0) | 0.846 ± 0.003aY | 50.13 ± 2.94a | 1.696 ± 0.308a | 0.793 ± 0.046ab | 0.373 ± 0.016ab | 1.166 ± 0.629ab |
| P:PW (90:10) | 0.835 ± 0.003ab | 51.30 ± 3.92a | 1.093 ± 0.211ab | 0.877 ± 0.038a | 0.413 ± 0.018a | 1.291 ± 0.057a | |
| P:PW (70:30) | 0.834 ± 0.004ab | 41.43 ± 3.43ab | 0.860 ± 0.085b | 0.714 ± 0.228b | 0.345 ± 0.011b | 1.059 ± 0.034b | |
| P:PW (50:50) | 0.824 ± 0.004b | 34.93 ± 1.70b | 0.850 ± 0.098b | 0.517 ± 0.002c | 0.267 ± 0.002c | 0.784 ± 0.001c | |
| Broccoli | P:PW (100:0) | 0.821 ± 0.005a | 49.76 ± 1.79a | 0.860 ± 0.072a | 0.821 ± 0.042bc | 0.397 ± 0.015c | 1.220 ± 0.057bc |
| P:PW (90:10) | 0.830 ± 0.011b | 47.66 ± 0.73a | 0.693 ± 0.046ab | 0.994 ± 0.010a | 0.490 ± 0.004a | 1.481 ± 0.017a | |
| P:PW (70:30) | 0.816 ± 0.001c | 41.46 ± 1.64b | 0.600 ± 0.052b | 0.870 ± 0.016b | 0.435 ± 0.012b | 0.301 ± 0.028b | |
| P:PW (50:50) | 0.813 ± 0.004c | 36.56 ± 1.61b | 0.620 ± 0.040b | 0.784 ± 0.011c | 0.392 ± 0.009c | 0.173 ± 0.020c | |
| Cabbage | P:PW (100:0) | 0.838 ± 0.002a | 39.03 ± 1.77a | 1.083 ± 0.082ab | 0.779 ± 0.093a | 0.374 ± 0.045a | 1.150 ± 0.138a |
| P:PW (90:10) | 0.824 ± 0.012ab | 39.26 ± 0.54a | 1.240 ± 0.135a | 0.637 ± 0.004ab | 0.323 ± 0.001ab | 0.956 ± 0.003ab | |
| P:PW (70:30) | 0.825 ± 0.002ab | 36.30 ± 0.55a | 0.803 ± 0.049b | 0.583 ± 0.001b | 0.270 ± 0.005bc | 0.806 ± 0.008b | |
| P:PW (50:50) | 0.807 ± 0.008b | 36.13 ± 1.87a | 0.910 ± 0.051b | 0.362 ± 0.010c | 0.215 ± 0.020c | 0.573 ± 0.020c |
| Minerals | P:PW (100:0) | P:PW (90:10) | P:PW (70:30) | P:PW (50:50) | |
|---|---|---|---|---|---|
| Cauliflower | N (g kg−1) | 22.14 ± 0.17d | 27.14 ± 0.16b | 35.67 ± 0.84a | 25.03 ± 0.17c |
| K (g kg−1) | 29.37 ± 0.03c | 37.37 ± 0.23b | 50.69 ± 0.17a | 36.65 ± 0.51b | |
| P (g kg−1) | 5.84 ± 0.08c | 6.91 ± 0.03b | 8.19 ± 0.12a | 7.26 ± 0.15b | |
| Mg (g kg−1) | 2.45 ± 0.09b | 1.42 ± 0.01c | 1.36 ± 0.03c | 3.17 ± 0.02a | |
| Ca (g kg−1) | 5.16 ± 0.08c | 5.88 ± 0.14c | 8.70 ± 0.38b | 12.21 ± 1.14a | |
| Na (g kg−1) | 15.99 ± 0.01a | 11.84 ± 0.18b | 11.74 ± 0.02b | 10.73 ± 0.19c | |
| Fe (mg kg−1) | 62.68 ± 1.96a | 46.77 ± 1.10b | 50.90 ± 1.62b | 36.21 ± 4.35c | |
| Cu (mg kg−1) | 21.42 ± 0.61c | 21.02 ± 0.49c | 36.23 ± 0.27b | 43.95 ± 0.58a | |
| Zn (mg kg−1) | 50.10 ± 3.05c | 115.39 ± 3.66a | 46.86 ± 0.40c | 78.75 ± 5.90b | |
| Broccoli | N (g kg−1) | 23.72 ± 0.00b | 22.21 ± 0.03c | 23.22 ± 0.11bc | 25.85 ± 0.62a |
| K (g kg−1) | 26.83 ± 0.11d | 30.04 ± 0.23c | 37.97 ± 0.80a | 35.40 ± 0.81b | |
| P (g kg−1) | 5.77 ± 0.00c | 6.47 ± 0.10b | 6.77 ± 0.03b | 7.03 ± 0.15a | |
| Mg (g kg−1) | 5.64 ± 0.01a | 5.38 ± 0.03b | 3.42 ± 0.03d | 4.62 ± 0.01c | |
| Ca (g kg−1) | 7.44 ± 0.14ab | 6.78 ± 0.42bc | 6.33 ± 0.17c | 7.74 ± 0.01a | |
| Na (g kg−1) | 9.49 ± 0.09b | 10.14 ± 0.02a | 7.85 ± 0.00c | 6.83 ± 0.13d | |
| Fe (mg kg−1) | 51.14 ± 0.42a | 46.02 ± 0.06b | 34.56 ± 0.04c | 29.26 ± 1.10d | |
| Cu (mg kg−1) | 27.78 ± 1.81b | 30.90 ± 1.81b | 30.14 ± 0.25b | 40.60 ± 1.09a | |
| Zn (mg kg−1) | 101.89 ± 4.74c | 187.20 ± 10.47a | 73.60 ± 0.37d | 143.73 ± 5.26b | |
| Cabbage | N (g kg−1) | 22.72 ± 0.45c | 23.45 ± 0.14bc | 32.39 ± 0.28a | 24.12 ± 0.16b |
| K (g kg−1) | 27.90 ± 0.22d | 32.70 ± 0.48c | 47.03 ± 0.83a | 36.80 ± 1.23b | |
| P (g kg−1) | 5.79 ± 0.11b | 5.82 ± 0.00b | 6.18 ± 0.09a | 5.78 ± 0.09b | |
| Mg (g kg−1) | 4.27 ± 0.09c | 3.43 ± 0.04d | 5.42 ± 0.16b | 6.68 ± 0.14a | |
| Ca (g kg−1) | 4.78 ± 0.30b | 5.43 ± 0.30b | 8.78 ± 0.14a | 8.06 ± 0.26a | |
| Na (g kg−1) | 7.83 ± 0.12ab | 7.01 ± 0.06c | 7.57 ± 0.02b | 8.13 ± 0.11a | |
| Fe (mg kg−1) | 57.24 ± 3.16a | 59.54 ± 0.97a | 35.71 ± 2.23b | 36.25 ± 2.54b | |
| Cu (mg kg−1) | 32.19 ± 1.20c | 37.01 ± 0.04c | 104.32 ± 4.24b | 120.41 ± 2.56a | |
| Zn (mg kg−1) | 23.05 ± 0.57d | 124.42 ± 3.58b | 87.48 ± 2.16c | 201.23 ± 0.99a |
| Data Set | Plant Growth and Physiology | Plant Stress Factors and Antioxidants | Leaf Minerals | |||
|---|---|---|---|---|---|---|
| Plant | Total Variance Explained by PCA,% | Variance of PW Containing GM,% | Total Variance Explained by PCA,% | Variance of PW Containing GM,% | Total Variance Explained by PCA,% | Variance of PW Containing GM,% |
| Cauliflower | 83.81 | 52.89 | 81.34 | 52.00 | 97.36 | 48.95 |
| Broccoli | 88.22 | 58.97 | 77.39 | 50.16 | 95.14 | 57.11 |
| Cabbage | 81.71 | 62.41 | 89.53 | 44.10 | 94.91 | 37.56 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chrysargyris, A.; Xylia, P.; Akinci, G.; Moustakas, K.; Tzortzakis, N. Printed Paper Waste as an Alternative Growing Medium Component to Produce Brassica Seedlings under Nursery Conditions. Sustainability 2020, 12, 5992. https://doi.org/10.3390/su12155992
Chrysargyris A, Xylia P, Akinci G, Moustakas K, Tzortzakis N. Printed Paper Waste as an Alternative Growing Medium Component to Produce Brassica Seedlings under Nursery Conditions. Sustainability. 2020; 12(15):5992. https://doi.org/10.3390/su12155992
Chicago/Turabian StyleChrysargyris, Antonios, Panayiota Xylia, Gorkem Akinci, Konstantinos Moustakas, and Nikolaos Tzortzakis. 2020. "Printed Paper Waste as an Alternative Growing Medium Component to Produce Brassica Seedlings under Nursery Conditions" Sustainability 12, no. 15: 5992. https://doi.org/10.3390/su12155992