
Dynamics and Drivers of Vegetation Phenology in Three-River Headwaters Region Based on the Google Earth Engine

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
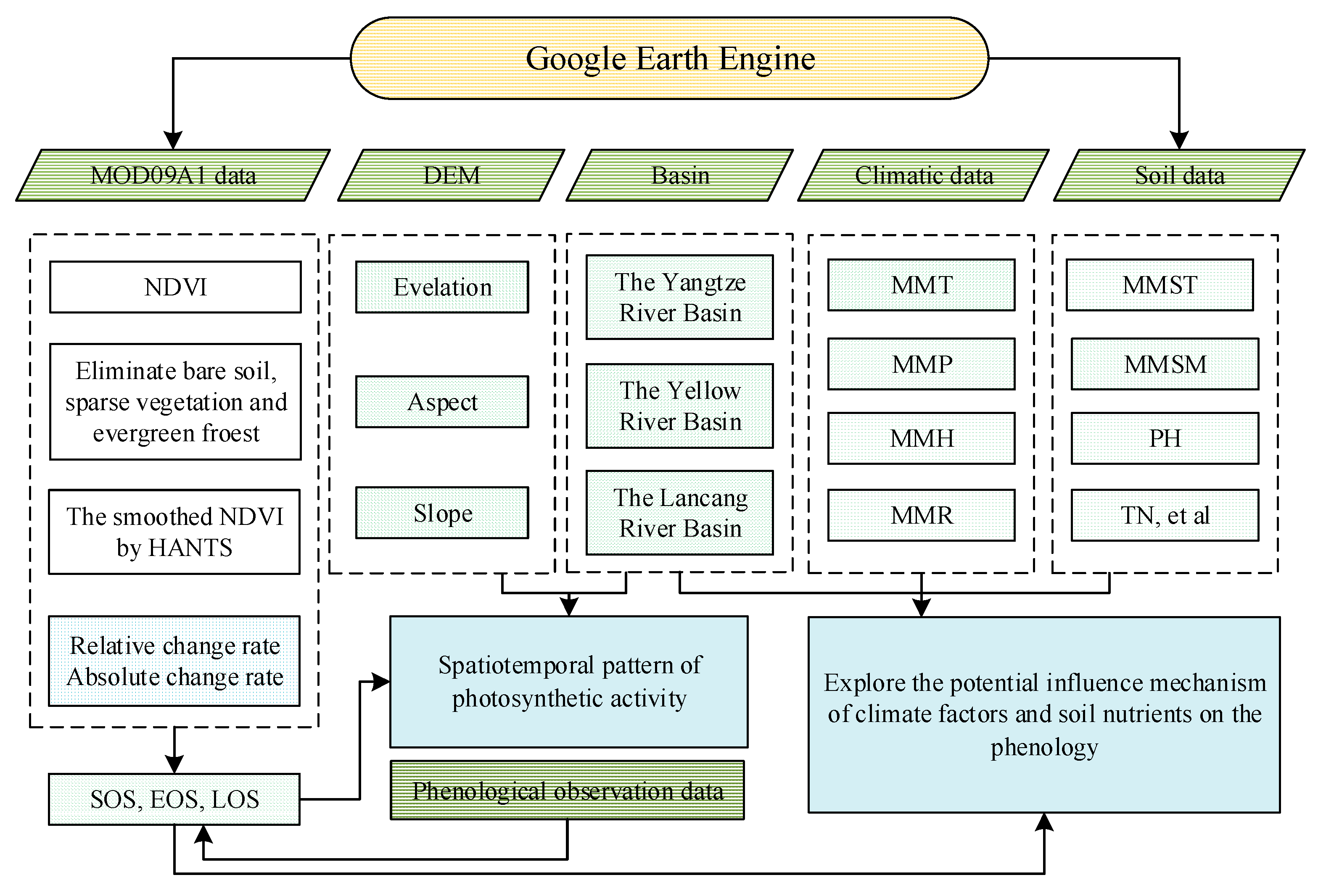
2.1. Processing
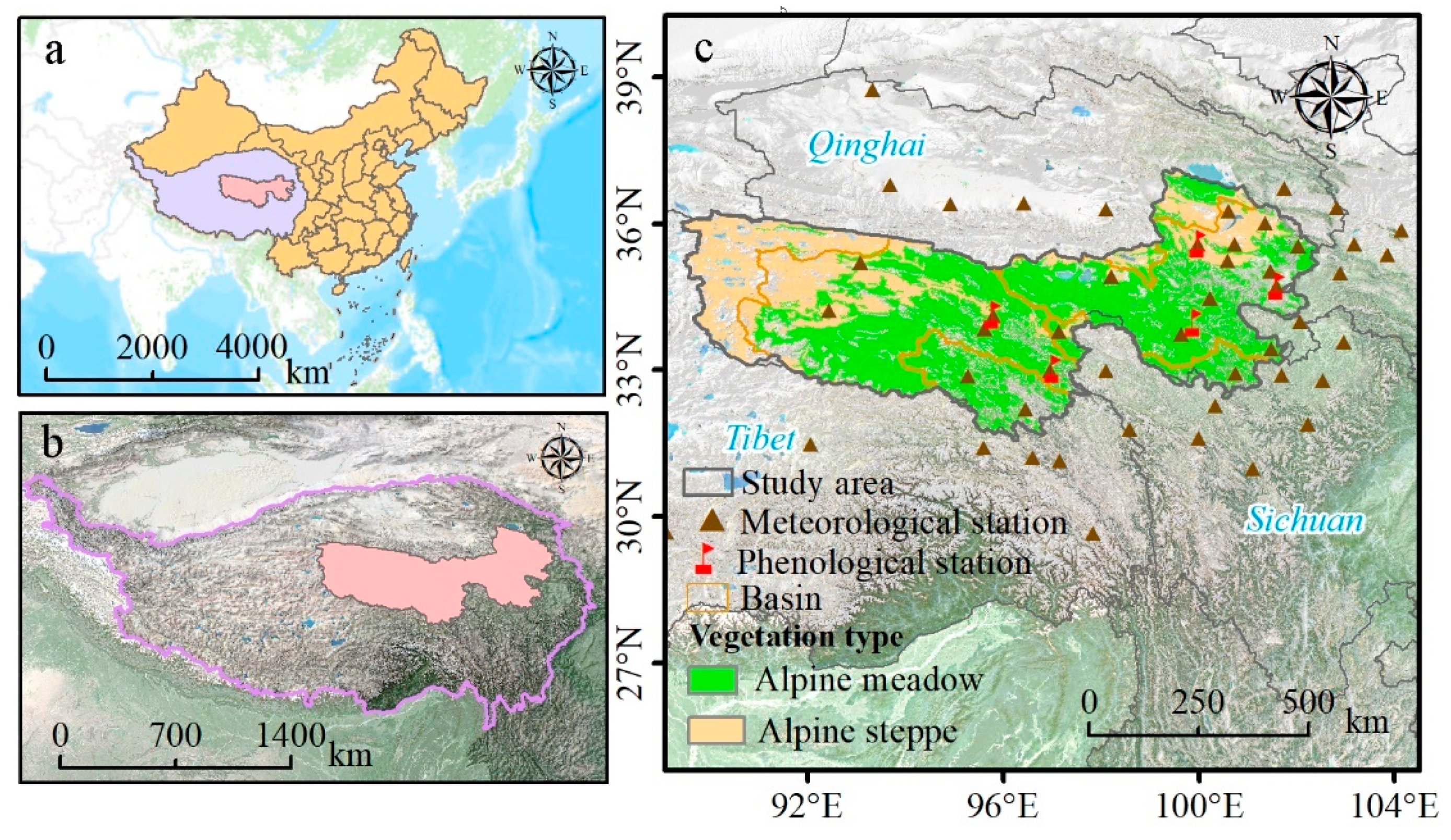
2.2. Study Area
2.3. Data Sources
2.3.1. MOD09A1 Data
2.3.2. Phenological Observation Data
2.3.3. Climate Datasets
2.3.4. Soil Characteristics Database
2.3.5. Digital Elevation Model
2.4. Methods
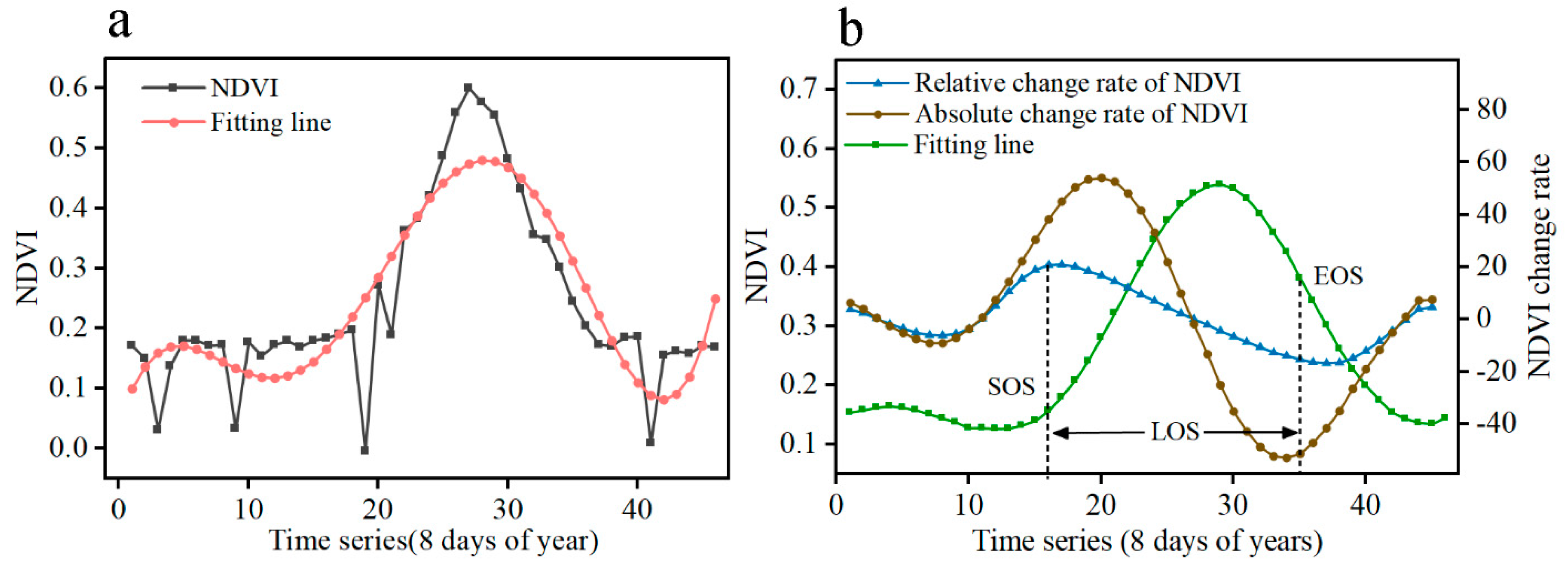
2.4.1. Extraction of Plant Phenological Information
- (1)
- NDVI
- (2)
- Determination of the SOS and EOS
2.4.2. The Spatiotemporal Pattern of Plant Phenology
- (1)
- Linear Regression Analysis
- (2)
- Standard Deviation Analysis
2.4.3. Driving Force Analysis
- (1)
- Pearson Correlation Coefficient
- (2)
- Structural Equation Model
3. Results
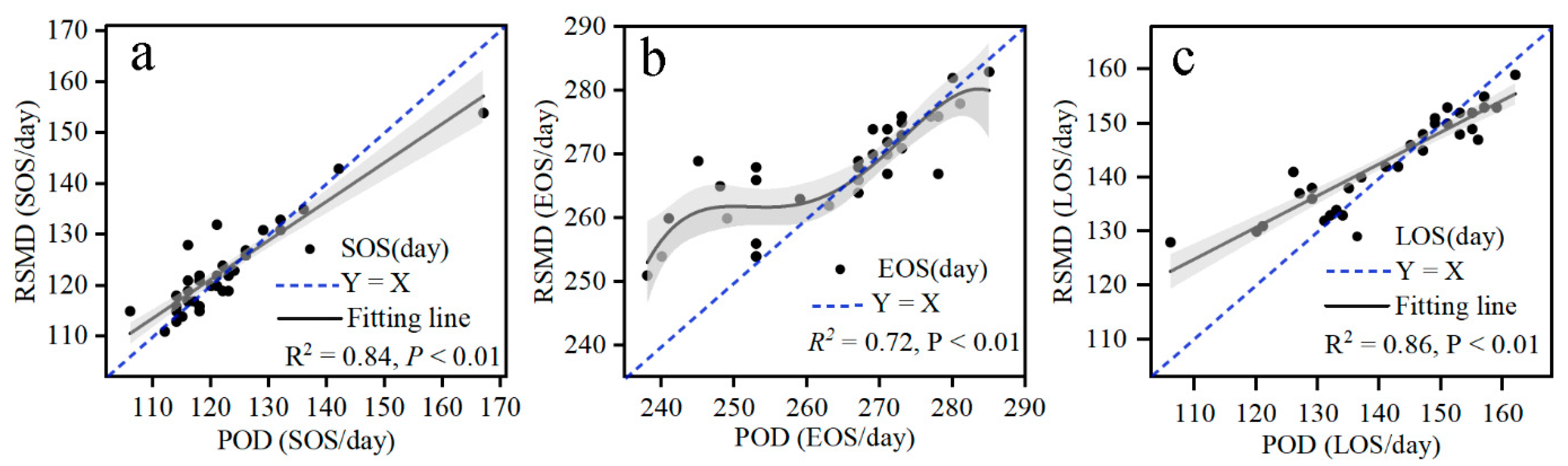
3.1. The Verification of Vegetation Phenological Results
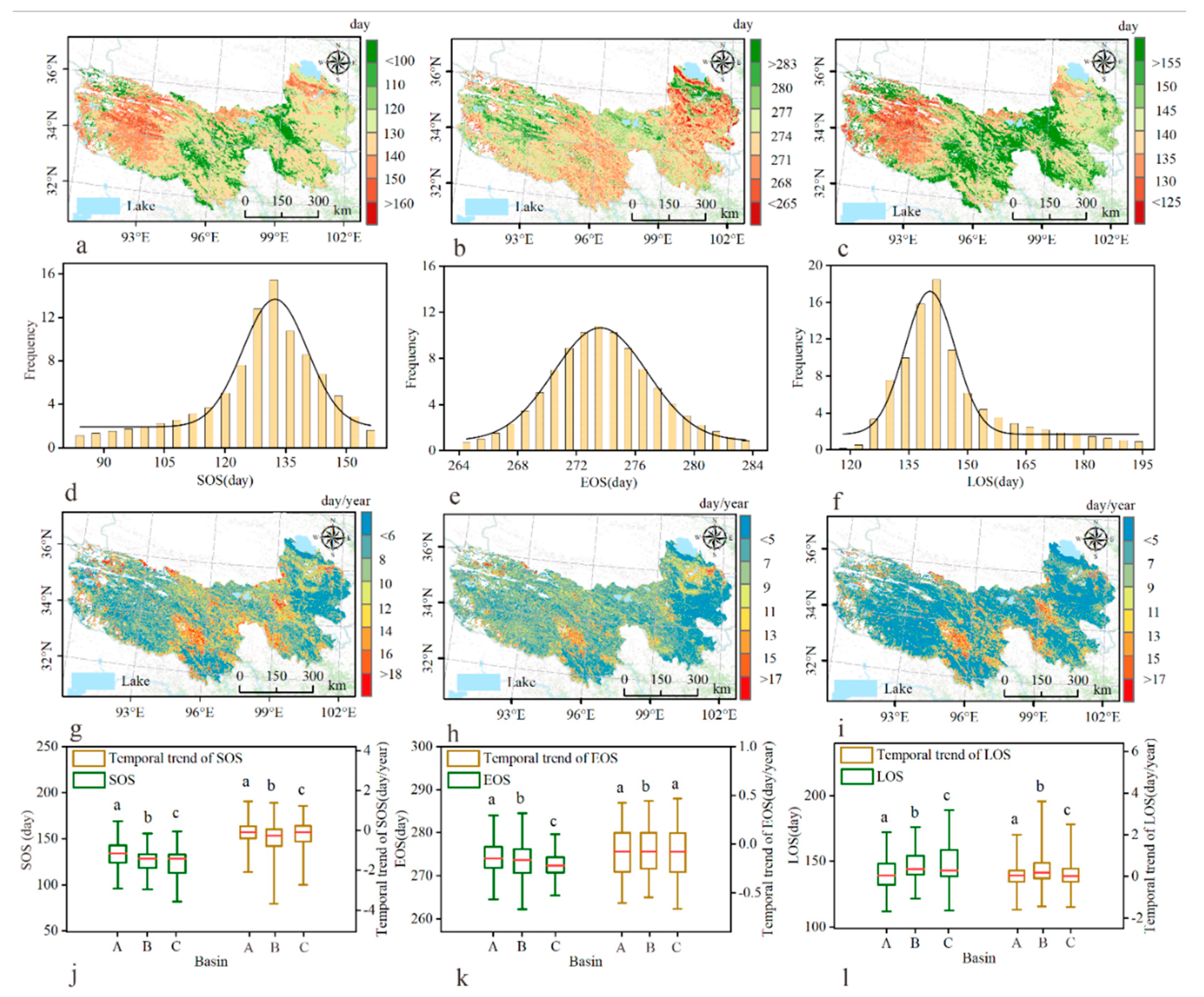
3.2. Spatiotemporal Pattern of Plant Phenology
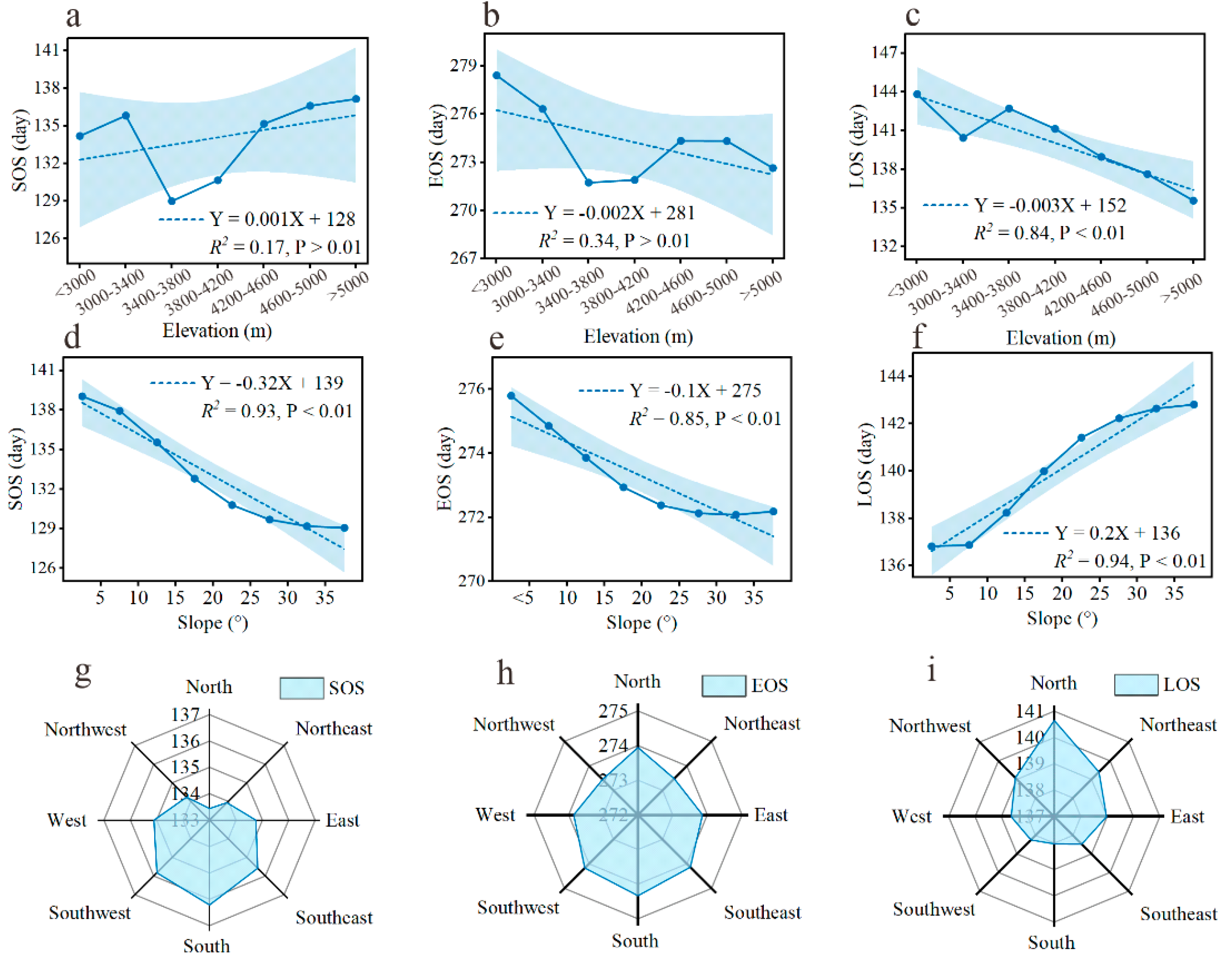
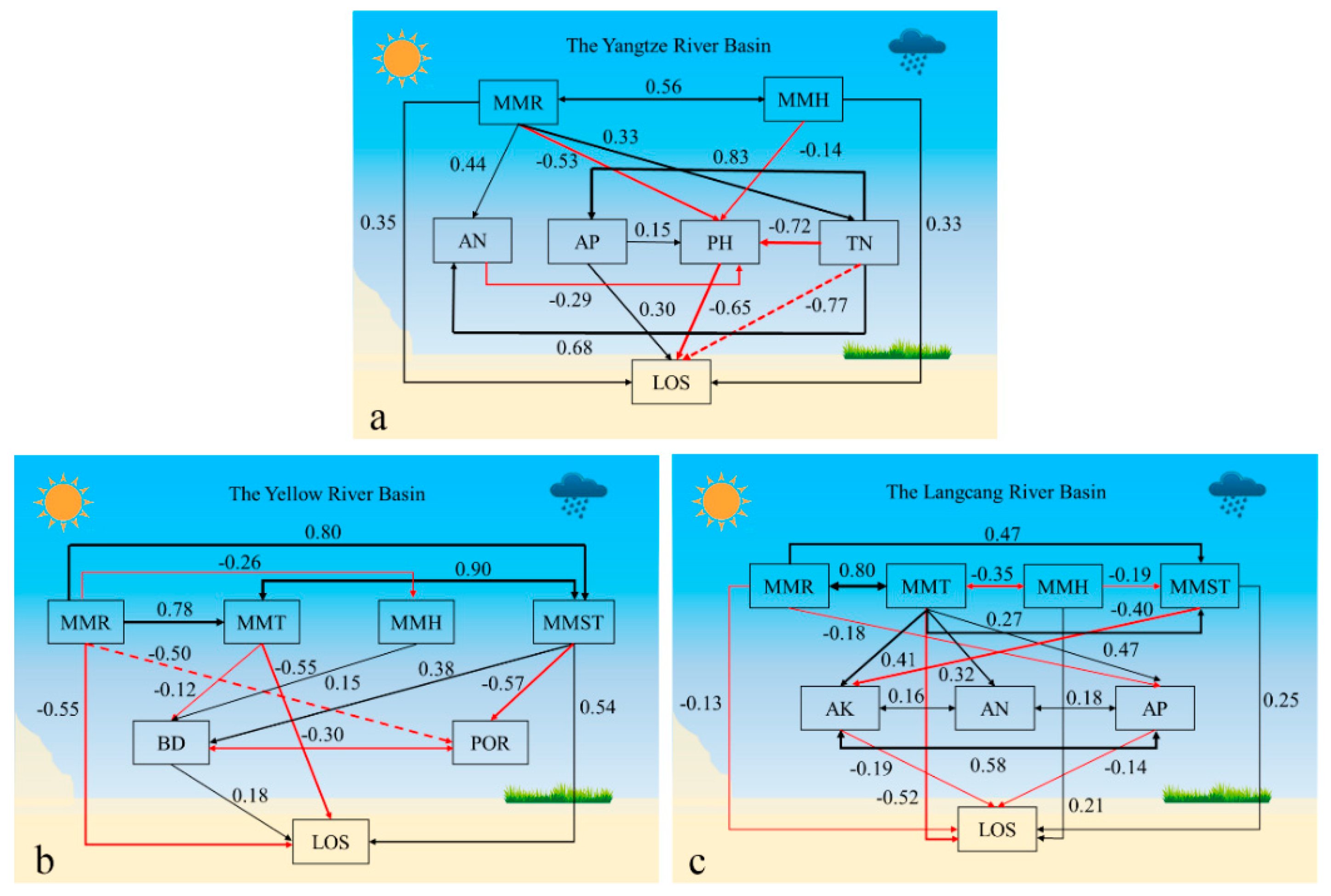
3.3. Linking Climatic and Soil Factors to Plant Phenology
4. Discussion
4.1. Spatial–Temporal Patterns of Plant Phenology
4.2. The Response of the Plant Phenology to Climate Change
4.3. Limitations of the Current Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- O’Neill, B.C.; Oppenheimer, M.; Warren, R.; Hallegatte, S.; Kopp, R.E.; Pörtner, H.O.; Scholes, R.; Birkmann, J.; Foden, W.; Licker, R.; et al. IPCC reasons for concern regarding climate change risks. Nat. Clim. Chang. 2017, 7, 28–37. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Zhang, Y.; Dong, J.; Xiao, X. Green-up dates in the Tibetan Plateau have continuously advanced from 1982 to 2011. Proc. Natl. Acad. Sci. USA 2013, 110, 4309–4314. [Google Scholar] [CrossRef] [Green Version]
- Brandt, L.A.; Butler, P.R.; Handler, S.D.; Janowiak, M.K.; Shannon, P.D.; Swanston, C.W. Integrating science and management to assess forest ecosystem vulnerability to climate change. J. For. 2017, 115, 212–221. [Google Scholar] [CrossRef]
- Piao, S.; Friedlingstein, P.; Ciais, P.; Viovy, N.; Demarty, J. Growing season extension and its impact on terrestrial carbon cycle in the Northern Hemisphere over the past 2 decades. Glob. Biogeochem. Cycles 2007, 21. [Google Scholar] [CrossRef]
- Gusewell, S.; Furrer, R.; Gehrig, R.; Pietragalla, B. Changes in temperature sensitivity of spring phenology with recent climate warming in Switzerland are related to shifts of the preseason. Glob. Chang. Biol. 2017, 23, 5189–5202. [Google Scholar] [CrossRef]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Dragoni, D.; Schmid, H.P.; Wayson, C.A.; Potter, H.; Grimmond, C.S.B.; Randolph, J.C. Evidence of increased net ecosystem productivity associated with a longer vegetated season in a deciduous forest in south-central Indiana, USA. Glob. Chang. Biol. 2011, 17, 886–897. [Google Scholar] [CrossRef]
- Geng, X.; Fu, Y.H.; Hao, F.; Zhou, X.; Zhang, X.; Yin, G.; Vitasse, Y.; Piao, S.; Niu, K.; de Boeck, H.J.; et al. Climate warming increases spring phenological differences among temperate trees. Glob. Chang. Biol. 2020, 26, 5979–5987. [Google Scholar] [CrossRef]
- Gao, M.; Wang, X.; Meng, F.; Liu, Q.; Li, X.; Zhang, Y.; Piao, S. Three-dimensional change in temperature sensitivity of northern vegetation phenology. Glob. Chang. Biol. 2020, 26, 5189–5201. [Google Scholar] [CrossRef]
- Shen, X.; An, R.; Feng, L.; Ye, N.; Zhu, L.; Li, M. Vegetation changes in the Three-River Headwaters Region of the Tibetan Plateau of China. Ecol. Indic. 2018, 93, 804–812. [Google Scholar] [CrossRef]
- Zhang, W.; Jin, H.; Shao, H.; Li, A.; Li, S.; Fan, W. Temporal and spatial variations in the leaf area index and its response to topography in the Three-River Source Region, China from 2000 to 2017. ISPRS Int. J. Geo Inf. 2021, 10, 33. [Google Scholar] [CrossRef]
- Binghong, H.; Bingrong, Z.; Henghe, Z.; Mingming, S.; Ying, S.; Decao, N.; Hua, F. Relationships between grassland vegetation turngreen and climate factors in the Three-river Resource region. Acta Ecol. Sin. 2019, 39, 5635–5641. [Google Scholar]
- Qiang, L. Phenology response of vegetation to hydrothermal condition in Three-river Source Region for the last 12 years. Arid Zone Res. 2016, 33, 150–158. [Google Scholar]
- Chen, T.; Yi, G.; Zhang, T.; Wang, Q.; Bie, X. A method for determining vegetation growth process using remote sensing data: A case study in the Three-River Headwaters Region, China. J. Mt. Sci. 2019, 16, 2001–2014. [Google Scholar] [CrossRef]
- Han, H.; Bai, J.; Ma, G.; Yan, J. Vegetation phenological changes in multiple landforms and responses to climate change. ISPRS Int. J. Geo-Inf. 2020, 9, 111. [Google Scholar] [CrossRef] [Green Version]
- Niu, B.; Zhang, X.; Piao, S.; Janssens, I.A.; Fu, G.; He, Y.; Zhang, Y.; Shi, P.; Dai, E.; Yu, C.; et al. Warming homogenizes apparent temperature sensitivity of ecosystem respiration. Sci. Adv. 2021, 7, eabc7358. [Google Scholar] [CrossRef]
- Chen, A.; Huang, L.; Liu, Q.; Piao, S. Optimal temperature of vegetation productivity and its linkage with climate and elevation on the Tibetan Plateau. Glob. Chang. Biol. 2021, 27, 1942–1951. [Google Scholar] [CrossRef]
- Cong, N.; Shen, M.; Yang, W.; Yang, Z.; Zhang, G.; Piao, S. Varying responses of vegetation activity to climate changes on the Tibetan Plateau grassland. Int. J. Biometeorol. 2017, 61, 1433–1444. [Google Scholar] [CrossRef]
- Menzel, A.; Sparks, T.H.; Estrella, N.; Koch, E.; Aasa, A.; Ahas, R.; Alm-Kübler, K.; Bissolli, P.; Braslavská, O.G.; Briede, A.; et al. European phenological response to climate change matches the warming pattern. Glob. Chang. Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef]
- Yun, J.; Jeong, S.-J.; Ho, C.-H.; Park, C.-E.; Park, H.; Kim, J. Influence of winter precipitation on spring phenology in boreal forests. Glob. Chang. Biol. 2018, 24, 5176–5187. [Google Scholar] [CrossRef]
- Nemani, R.R.; Keeling, C.D.; Hashimoto, H.; Jolly, W.M.; Piper, S.C.; Tucker, C.J.; Myneni, R.B.; Running, S.W. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 2003, 300, 1560–1563. [Google Scholar] [CrossRef] [Green Version]
- Chapin, F.S., III; Matson, P.A.; Vitousek, P. Principles of Terrestrial Ecosystem Ecology; Springer Science & Business Media: Berlin, Germany, 2002; pp. 369–397. [Google Scholar]
- Yue, K.; Peng, Y.; Peng, C.; Yang, W.; Peng, X.; Wu, F. Stimulation of terrestrial ecosystem carbon storage by nitrogen addition: A meta-analysis. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Zhou, T.C.; Liu, M.; Chen, Y.C.; Liu, G.H.; Xu, M.; Shi, P.L.; Peng, F.; Tsunekawa, A.; Liu, Y.; et al. Water and heat availability are drivers of the aboveground plant carbon accumulation rate in alpine grasslands on the Tibetan Plateau. Glob. Ecol. Biogeogr. 2019, 29, 50–64. [Google Scholar] [CrossRef] [Green Version]
- Gower, S.T.; Kucharik, C.J.; Norman, J.M. Direct and indirect estimation of leaf area index, f APAR, and net primary production of terrestrial ecosystems. Remote Sens. Environ. 1999, 70, 29–51. [Google Scholar] [CrossRef]
- Jiang, C.; Zhang, L. Ecosystem change assessment in the Three-river Headwater Region, China: Patterns, causes, and implications. Ecol. Eng. 2016, 93, 24–36. [Google Scholar] [CrossRef]
- Xianfeng, L.; Jinshui, Z.; Xiufang, Z.; Yaozhong, P.; Yanxu, L.; Donghai, Z.; Zhihui, L. Spatiotemporal changes in vegetation coverage and its driving factors in the Three-River Headwaters Region during 2000–2011. J. Geogr. Sci. 2014, 24, 288–302. [Google Scholar]
- Zheng, D.; Wang, Y.; Hao, S.; Xu, W.; Lv, L.; Yu, S. Spatial-temporal variation and tradeoffs/synergies analysis on multiple ecosystem services: A case study in the Three-River Headwaters region of China. Ecol. Indic. 2020, 116, 106494. [Google Scholar] [CrossRef]
- Zheng, Y.; Han, J.; Huang, Y.; Fassnacht, S.R.; Xie, S.; Lv, E.; Chen, M. Vegetation response to climate conditions based on NDVI simulations using stepwise cluster analysis for the Three-River Headwaters region of China. Ecol. Indic. 2018, 92, 18–29. [Google Scholar] [CrossRef]
- Zeng, N.; He, H.; Ren, X.; Zhang, L.; Zeng, Y.; Fan, J.; Li, Y.; Niu, Z.; Zhu, X.; Chang, Q. The utility of fusing multi-sensor data spatio-temporally in estimating grassland aboveground biomass in the three-river headwaters region of China. Int. J. Remote Sens. 2020, 41, 7068–7089. [Google Scholar] [CrossRef]
- Piao, S.; Cui, M.; Chen, A.; Wang, X.; Ciais, P.; Liu, J.; Tang, Y. Altitude and temperature dependence of change in the spring vegetation green-up date from 1982 to 2006 in the Qinghai-Xizang Plateau. Agric. For. Meteorol. 2011, 151, 1599–1608. [Google Scholar] [CrossRef]
- Li, L.; Zhang, Y.; Liu, L.; Wu, J.; Wang, Z.; Li, S.; Zhang, H.; Zu, J.; Ding, M.; Paudel, B. Spatiotemporal patterns of vegetation greenness change and associated climatic and anthropogenic drivers on the Tibetan Plateau during 2000–2015. Remote Sens. 2018, 10, 1525. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Liu, H.; Lin, Z.; Jiao, F.; Gong, H. Relationship of abrupt vegetation change to climate change and ecological engineering with multi-timescale analysis in the Karst Region, Southwest China. Remote Sens. 2019, 11, 1564. [Google Scholar] [CrossRef] [Green Version]
- He, D.; Huang, X.; Tian, Q.; Zhang, Z. Changes in vegetation growth dynamics and relations with climate in inner Mongolia under more strict multiple pre-processing (2000–2018). Sustainability 2020, 12, 2534. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Wang, J.; Xiong, J.; Bian, J.; Jin, H.; Cheng, W.; Li, A. Vegetation change and its response to climate change in Yunnan Province, China. Adv. Meteorol. 2021, 2021, 8857589. [Google Scholar] [CrossRef]
- Shipley, B. Cause and correlation in biology. Ann. Bot. 2002, 90, 777–778. [Google Scholar]
- Zhang, Y.; Gao, J.; Liu, L.; Wang, Z.; Ding, M.; Yang, X. NDVI-based vegetation changes and their responses to climate change from 1982 to 2011: A case study in the Koshi River Basin in the middle Himalayas. Glob. Planet. Chang. 2013, 108, 139–148. [Google Scholar] [CrossRef]
- Kong, D.; Zhang, Q.; Singh, V.P.; Shi, P. Seasonal vegetation response to climate change in the Northern Hemisphere (1982–2013). Glob. Planet. Chang. 2017, 148, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Ye, C.; Sun, J.; Liu, M.; Xiong, J.; Zong, N.; Hu, J.; Huang, Y.; Duan, X.; Tsunekawa, A. Concurrent and lagged effects of extreme drought induce net reduction in vegetation carbon uptake on Tibetan Plateau. Remote Sens. 2020, 12, 2347. [Google Scholar] [CrossRef]
- Yao, Y.; Wang, X.; Li, Y.; Wang, T.; Shen, M.; Du, M.; He, H.; Li, Y.; Luo, W.; Ma, M.; et al. Spatiotemporal pattern of gross primary productivity and its covariation with climate in China over the last thirty years. Glob. Chang. Biol. 2018, 24, 184–196. [Google Scholar] [CrossRef]
- Davidson, E.A.; Janssens, I.A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 2006, 440, 165–173. [Google Scholar] [CrossRef]
- Kim, D.G.; Vargas, R.; Bond-Lamberty, B.; Turetsky, M.R. Effects of soil rewetting and thawing on soil gas fluxes: A review of current literature and suggestions for future research. Biogeosciences 2012, 9, 2459–2483. [Google Scholar] [CrossRef] [Green Version]
- Bao, G.; Qin, Z.; Bao, Y.; Zhou, Y.; Li, W.; Sanjjav, A. NDVI-based long-term vegetation dynamics and its response to climatic change in the Mongolian Plateau. Remote Sens. 2014, 6, 8337–8358. [Google Scholar] [CrossRef] [Green Version]
- Duan, A.; Wu, G.; Liu, Y.; Ma, Y.; Zhao, P. Weather and climate effects of the Tibetan Plateau. Adv. Atmos. Sci. 2012, 29, 978–992. [Google Scholar] [CrossRef]
- Immerzeel, W.W.; Bierkens, M.F.P. Asian water towers: More on monsoons—Response. Science 2010, 330, 584–585. [Google Scholar]
- Huang, K.; Zhang, Y.; Zhu, J.; Liu, Y.; Zu, J.; Zhang, J. The influences of climate change and human activities on vegetation dynamics in the Qinghai-Tibet Plateau. Remote Sens. 2016, 8, 876. [Google Scholar] [CrossRef] [Green Version]
- Guojin, P.; Xuejia, W.; Meixue, Y. Using the NDVI to identify variations in, and responses of, vegetation to climate change on the Tibetan Plateau from 1982 to 2012. Quat. Int. 2016, 444, 87–96. [Google Scholar] [CrossRef]
- Pu, Y.; Long, G.; Liu, S.; Lu, C.; Kang, Q. Research progress in slope-directive variation of mountain soils. Chin. J. Soil Sci. 2007, 38, 753–757. [Google Scholar] [CrossRef]
- Gao, T.; Li, J.; Lu, J.; Zheng, W.; Chen, J.; Wang, J.; Duan, F. Soil nutrient and fertility of different slope directions in the Abies georgei var. smithii forest in Sejila Mountain. Acta Ecol. Sin. 2020, 40, 1331–1341. [Google Scholar]
- Liu, M. Response of plant element content and soil factors to the slope gradient of alpine meadows in Gannan. Acta Ecol. Sin. 2017, 37, 8275–8284. [Google Scholar]
- Makiranta, P.; Laiho, R.; Mehtatalo, L.; Strakova, P.; Sormunen, J.; Minkkinen, K.; Penttila, T.; Fritze, H.; Tuittila, E.S. Responses of phenology and biomass production of boreal fens to climate warming under different water-table level regimes. Glob. Chang. Biol. 2018, 24, 944–956. [Google Scholar] [CrossRef]
- Reichstein, M.; Bahn, M.; Ciais, P.; Frank, D.; Mahecha, M.D.; Seneviratne, S.I.; Zscheischler, J.; Beer, C.; Buchmann, N.; Frank, D.C.; et al. Climate extremes and the carbon cycle. Nature 2013, 500, 287–295. [Google Scholar] [CrossRef]
- Gang, C.; Zhou, W.; Chen, Y.; Wang, Z.; Sun, Z.; Li, J.; Qi, J.; Odeh, I. Quantitative assessment of the contributions of climate change and human activities on global grassland degradation. Environ. Earth Sci. 2014, 72, 4273–4282. [Google Scholar] [CrossRef]
- Keenan, T.; Sabate, S.; Gracia, C. The importance of mesophyll conductance in regulating forest ecosystem productivity during drought periods. Glob. Chang. Biol. 2010, 16, 1019–1034. [Google Scholar] [CrossRef]
- Zeng, L.; Wardlow, B.D.; Xiang, D.; Hu, S.; Li, D. A review of vegetation phenological metrics extraction using time-series, multispectral satellite data. Remote Sens. Environ. 2020, 237, 111511. [Google Scholar] [CrossRef]
- Li, N.; Zhan, P.; Pan, Y.; Zhu, X.; Li, M.; Zhang, D. Comparison of remote sensing time-series smoothing methods for grassland spring phenology extraction on the Qinghai-Tibetan Plateau. Remote Sens. 2020, 12, 3383. [Google Scholar] [CrossRef]
- Zhu, W.; Pan, Y.; He, H.; Wang, L.; Mou, M.; Liu, J. A changing-weight filter method for reconstructing a high-quality NDVI time series to preserve the integrity of vegetation phenology. IEEE Trans. Geosci. Remote Sens. 2012, 50, 1085–1094. [Google Scholar] [CrossRef]
- Spiess, A.N.; Deutschmann, C.; Burdukiewicz, M.; Himmelreich, R.; Klat, K.; Schierack, P.; Rodiger, S. Impact of smoothing on parameter estimation in quantitative DNA amplification experiments. Clin. Chem 2015, 61, 379–388. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MMST | MMH | MMT | MMP | MMSM | AN | AP | TN | SOM | CEC | TP | AK | POR | MMR | PH | TK | BD | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SOS a | −0.06 ** | −0.38 ** | −0.13 ** | −0.68 ** | −0.37 ** | −0.39 ** | −0.31 ** | −0.24 ** | −0.22 ** | −0.23 ** | 0.04 ** | 0.09 ** | −0.03 * | 0.73 ** | 0.50 ** | 0.44 ** | 0.06 ** |
| SOS b | 0.48 ** | −0.16 ** | 0.50 ** | −0.04 ** | 0.31 ** | −0.04 ** | 0.10 ** | 0.09 ** | 0.09 ** | 0.12 ** | 0.14 ** | 0.15 ** | −0.19 ** | 0.50 ** | 0.09 ** | 0.10 ** | −0.05 ** |
| SOS c | 0.55 ** | −0.36 ** | 0.65 ** | 0 | 0.43 ** | 0.31 ** | 0.35 ** | 0.29 ** | 0.23 ** | 0.26 ** | 0.05 | 0.50 ** | −0.26 ** | 0.53 ** | −0.09 ** | 0.12 ** | −0.08 ** |
| EOS a | 0.22 ** | −0.16 ** | 0.10 ** | −0.45 ** | −0.27 ** | −0.27 ** | −0.17 ** | −0.17 ** | −0.16 ** | −0.19 ** | 0.08 ** | 0.05 ** | 0.02 | 0.44 ** | 0.40 ** | 0.41 ** | 0.02 |
| EOS b | 0.24 ** | −0.23 ** | 0.20 ** | −0.23 ** | −0.39 ** | −0.32 ** | 0 | −0.12 ** | −0.07 ** | −0.07 ** | 0.05 ** | 0.04 ** | −0.21 ** | 0.03 | 0.37 ** | 0.01 | −0.06 ** |
| EOS c | −0.41 ** | 0.08 ** | −0.43 ** | −0.33 ** | −0.22 ** | −0.12 ** | 0.13 ** | 0.01 | 0.04 | 0.04 | 0.09 ** | −0.12 ** | 0.24 ** | −0.36 ** | 0.07 ** | −0.26 ** | −0.18 ** |
| LOS a | 0.17 ** | 0.52 ** | 0.21 ** | 0.21 ** | −0.02 | 0.38 ** | 0.33 ** | 0.24 ** | 0.21 ** | 0.21 ** | −0.01 | −0.09 ** | 0.06 ** | 0.53 ** | −0.46 ** | −0.37 ** | −0.07 ** |
| LOS b | −0.43 ** | 0.10 ** | −0.46 ** | 0.02 | −0.02 | −0.05 ** | −0.10 ** | −0.13 ** | −0.11 ** | −0.14 ** | −0.13 ** | −0.15 ** | 0.14 ** | −0.55 ** | 0.01 | −0.10 ** | 0.04 ** |
| LOS c | −0.65 ** | 0.41 ** | −0.69 ** | −0.08 ** | −0.16 ** | −0.32 ** | −0.33 ** | −0.28 ** | −0.22 ** | −0.25 ** | −0.03 | −0.50 ** | 0.28 ** | −0.58 ** | 0.10 ** | −0.15 ** | 0.06 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Sun, H.; Xiong, J.; He, D.; Cheng, W.; Ye, C.; Yong, Z.; Huang, X. Dynamics and Drivers of Vegetation Phenology in Three-River Headwaters Region Based on the Google Earth Engine. Remote Sens. 2021, 13, 2528. https://doi.org/10.3390/rs13132528
Wang J, Sun H, Xiong J, He D, Cheng W, Ye C, Yong Z, Huang X. Dynamics and Drivers of Vegetation Phenology in Three-River Headwaters Region Based on the Google Earth Engine. Remote Sensing. 2021; 13(13):2528. https://doi.org/10.3390/rs13132528
Chicago/Turabian StyleWang, Jiyan, Huaizhang Sun, Junnan Xiong, Dong He, Weiming Cheng, Chongchong Ye, Zhiwei Yong, and Xianglin Huang. 2021. "Dynamics and Drivers of Vegetation Phenology in Three-River Headwaters Region Based on the Google Earth Engine" Remote Sensing 13, no. 13: 2528. https://doi.org/10.3390/rs13132528