Breaking the Dormancy of Snake’s Head Fritillary (Fritillaria meleagris L.) In Vitro Bulbs—Part 2: Effect of GA3 Soaking and Chilling on Sugar Status in Sprouted Bulbs


 , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
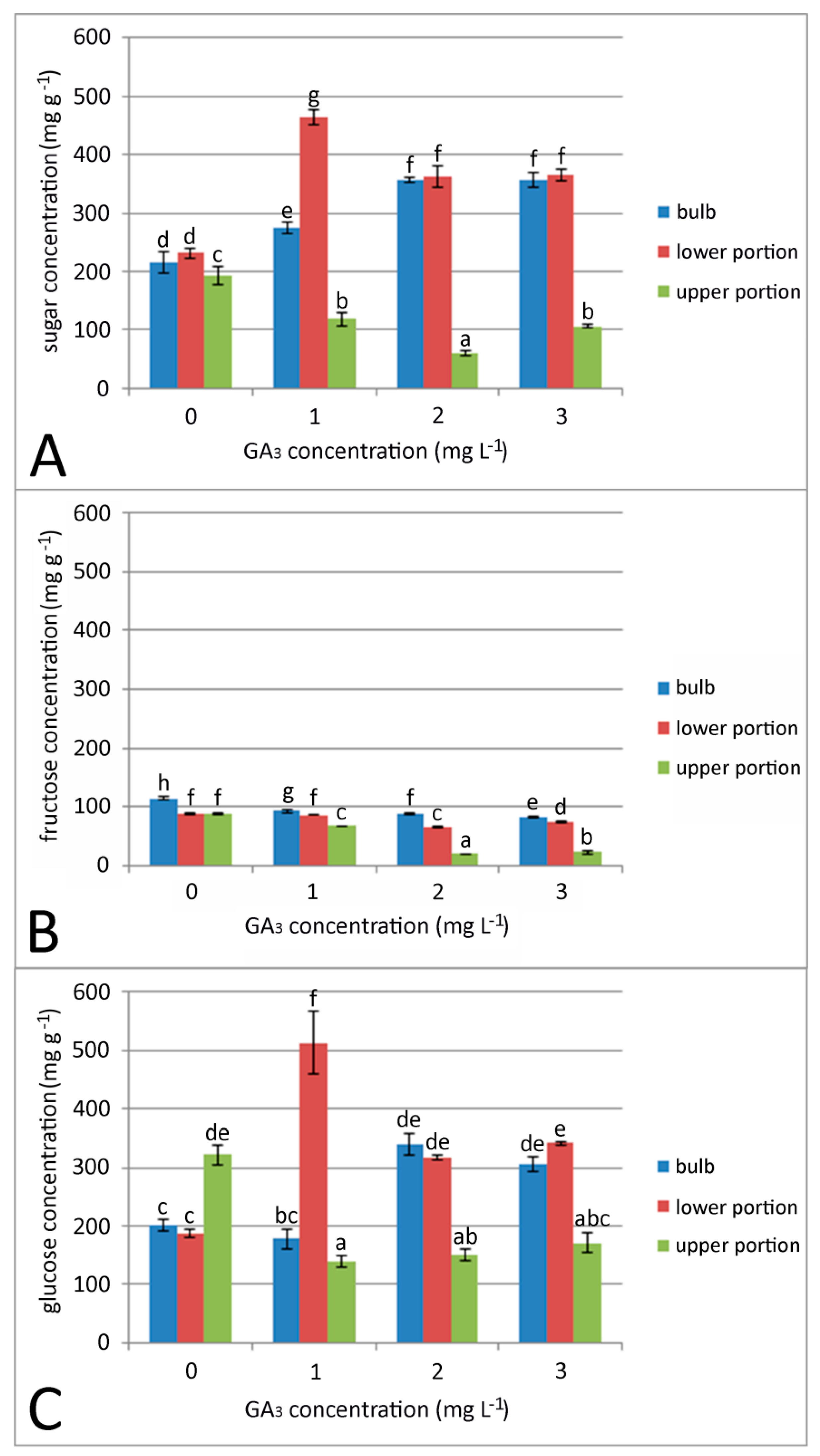
2.1. Effect of GA3 Soaking on Bulb Development and Sugar Content
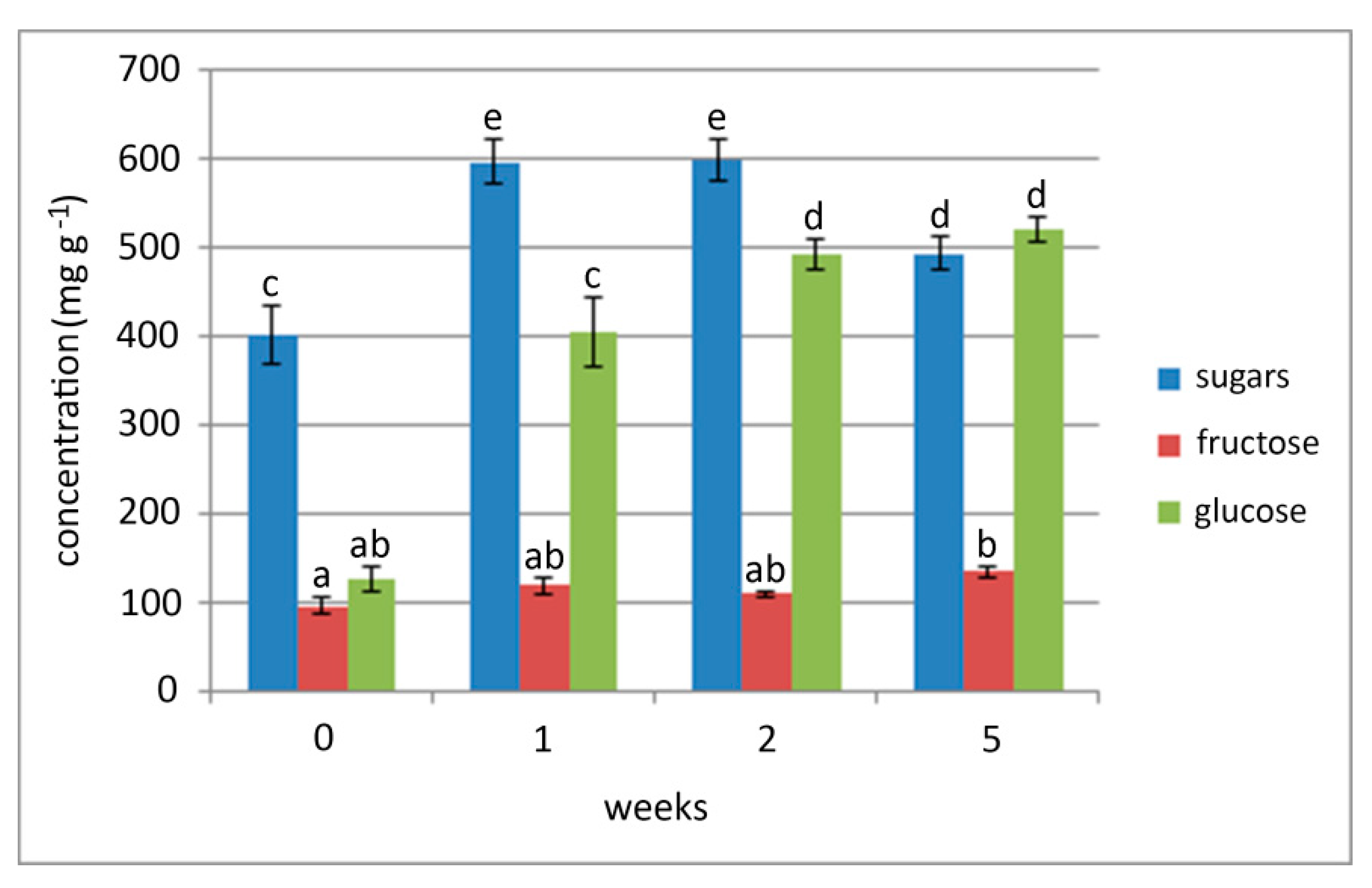
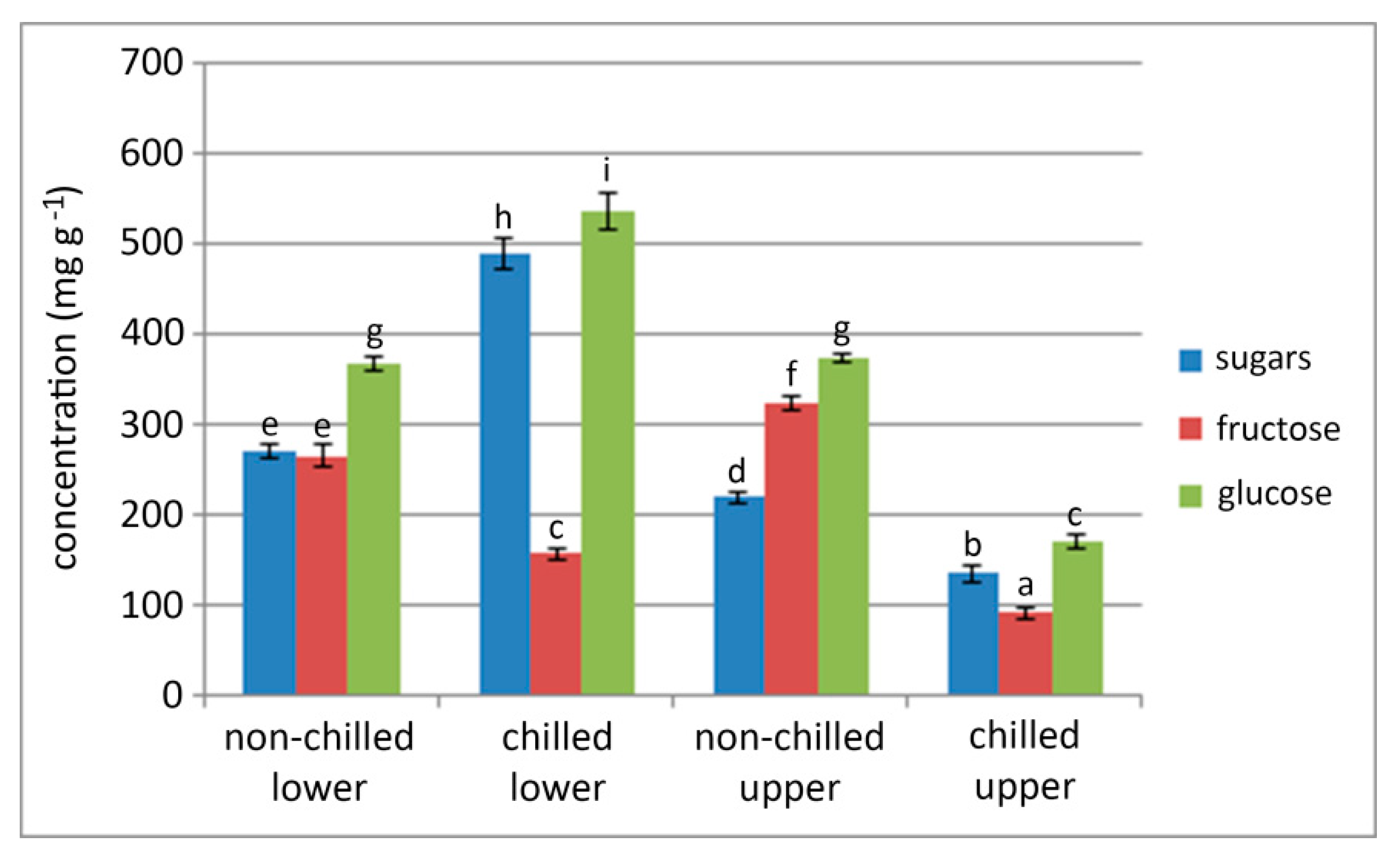
2.2. Effect of Chilling of Bulbs on Sugar Content in Different Parts of the Sprouted Bulbs
3. Discussion
4. Material and Methods
4.1. Plant Material and Culture Conditions
4.1.1. Bulb Soaking with GA3
4.1.2. Chilling of Bulbs
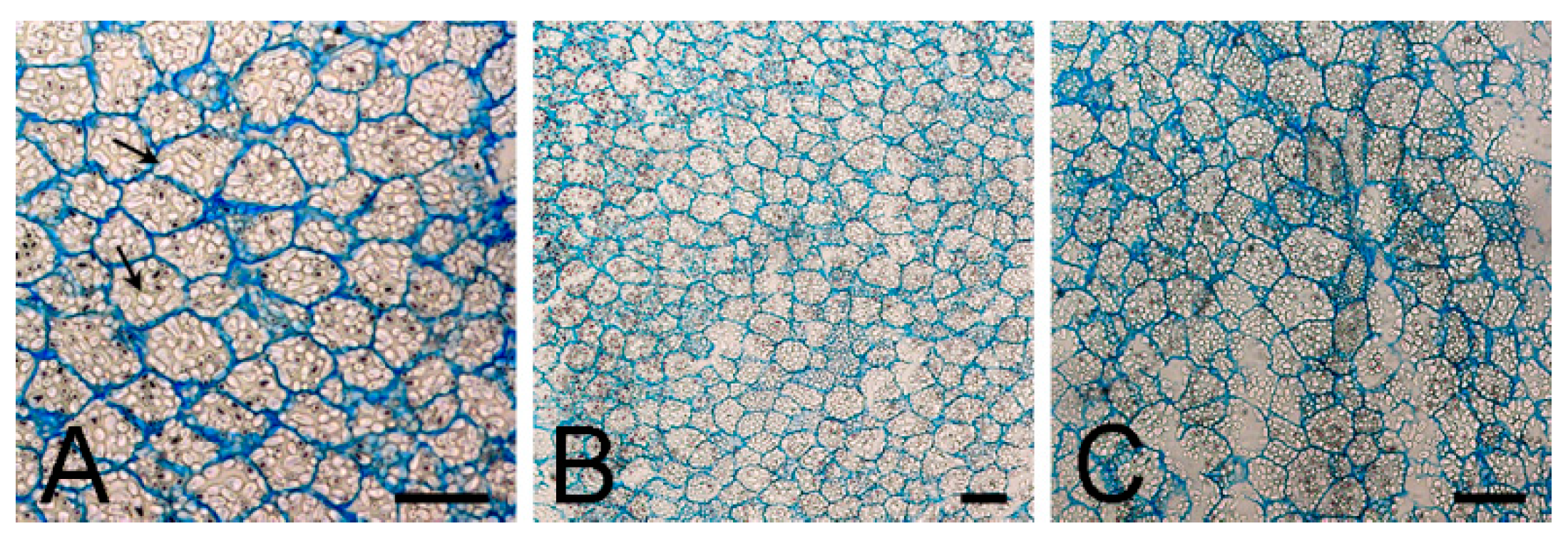
4.2. Morpho-Anatomical Analysis
4.3. Sugar Analysis
4.3.1. Determination of Total Sugar Content
4.3.2. Fructose Determination
4.3.3. Glucose Determination
4.4. Statistical Analysis of Data
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Petrić, M.; Jevremović, S.; Trifunović, M.; Tadić, V.; Milošević, S.; Dragićević, M.; Subotić, A. The effect of low temperature and GA3 treatments on dormancy breaking and activity of antioxidant enzymes in Fritillaria meleagris bulblets cultured in vitro. Acta Physiol. Plant. 2013, 35, 3223–3236. [Google Scholar] [CrossRef]
- Li, X.; Wang, C.; Cheng, J.; Zhang, J.; Da Silva, J.A.T.; Liu, X.; Duan, X.; Li, T.; Sun, H. Transcriptome analysis of carbohydrate metabolism during bulblet formation and development in Lilium davidii var. unicolor. BMC Plant Biol. 2014, 14, 358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viola, R.; Roberts, A.G.; Haupt, S.; Gazzani, S.; Hancock, R.D.; Marmiroli, N.; Machray, G.C.; Oparka, K.J. Tuberization in potato involves a switch from apoplastic to symplastic phloem unloading. Plant Cell 2001, 13, 385–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, K. Sucrose metabolism: Regulatory mechanisms and pivotal roles in sugar sensing and plant development. Curr. Opin. Plant Biol. 2004, 7, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Zhu, Z.; Guo, Q.; Yang, X.; Liu, L.; Sun, Y.; Wang, C. Dynamic changes in carbohydrate metabolism and endogenous hormones during Tulipa edulis stolon development into a new bulb. J. Plant Biol. 2016, 59, 121–132. [Google Scholar] [CrossRef]
- Atif, M.J.; Ahanger, M.A.; Amin, B.; Ghani, M.I.; Ali, M.; Cheng, Z. Mechanism of Allium Crops Bulb Enlargement in Response to Photoperiod: A Review. Int. J. Mol. Sci. 2020, 21, 1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.H.; Haque, M.S.; Karim, M.A.; Ahmed, M. Effects of gibberellic acid (GA3) on breaking dormancy in garlic (Allium sativum L.). Int. J. Agric. Biol. 2006, 8, 63–65. [Google Scholar]
- Podwyszyńska, M. The mechanisms of in vitro storage organ formation in ornamental geophytes. In Floriculture and Ornamental Biotechnology; Tuyl, J.M., Arens, P., Eds.; Global Science Books: London, UK, 2012; pp. 9–23. [Google Scholar]
- Langens-Gerrits, M.; Lilien-Kipnis, H.; Croes, T.; Miller, W.; Kollöffel, C.; De Klerk, G.-J.M. Bulb Growth in Lily Regenerated in Vitro. Acta Hortic. 1997, 430, 267–274. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, Y.; Yang, F.; Qi, X.; Ahmad, H.; Wu, C.; Cheng, Z. Effect of the mode and time of gibberellic acid treatment on plant architecture and bulb structure in garlic (Allium sativum L.). Sci. Hortic. 2019, 257, 108723. [Google Scholar] [CrossRef]
- Ranwala, A.P.; Miller, W.B. Analysis of nonstructural carbohydrates in storage organs of 30 ornamental geophytes by high-performance anion-exchange chromatography with pulsed amperometric detection. New Phytol. 2008, 180, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Rouhier, H.; Usuda, H. Spatial and Temporal Distribution of Sucrose Synthase in the Radish Hypocotyl in Relation to Thickening Growth. Plant Cell Physiol. 2001, 42, 583–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marković, M.; Trifunović Momčilov, M.; Uzelac, B.; Cingel, A.; Milošević, S.; Jevremović, S.; Subotić, A. Breaking the Dormancy of Snake’s Head Fritillary (Fritillaria meleagris L.) In Vitro Bulbs—Part 1: Effect of GA3, GA Inhibitors and Temperature on Fresh Weight, Sprouting and Sugar Content. Plants 2020, 9, 1449. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.-H.; Chung, Y.M.; Yang, S.J.; Ko, E.K.; Jeong, S.J.; Nam, J.S.; Kim, G.T.; Yi, Y.B. Effects of storage temperature and sucrose on bulblet growth, starch and protein contents in in vitro cultures of Hyacinthus orientalis. Biol. Plant. 2006, 50, 346–351. [Google Scholar] [CrossRef]
- Zaffryar, S.; Zimerman, B.; Abu-Abied, M.; Belausov, E.; Lurya, G.; Vainstein, A.; Kamenetsky, R.; Sadot, E. Development-specific association of amyloplasts with microtubules in scale cells of Narcissus tazetta. Protoplasma 2007, 230, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Fernie, A.R.; Willmitzer, L. Molecular and biochemical triggers of potato tuber development. Plant Physiol. 2001, 127, 1459–1465. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco culture. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Jensen, W.A. Botanical Histochemistry: Principles and Practices; WH Freeman and Company: San Francisco, CA, USA, 1962. [Google Scholar]
- Dreywood, R. Qualitative Test for Carbohydrate Material. Ind. Eng. Chem. Anal. Ed. 1946, 18, 499. [Google Scholar] [CrossRef]
- Messineo, L.; Musarra, E. Sensitive spectrophotometric determination of fructose, sucrose, and inulin without interference from aldohexoses, aldopentoses, and ketopentoses. Int. J. Biochem. 1972, 3, 691–699. [Google Scholar] [CrossRef]
- Amaral, L.I.V.D.; Gaspar, M.; Costa, P.M.F.; Aidar, M.P.M.; Buckeridge, M.S. Novo método enzimático rápido e sensível de extração e dosagem de amido em materiais vegetais. Hoehnea 2007, 34, 425–431. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marković, M.; Trifunović Momčilov, M.; Uzelac, B.; Radulović, O.; Milošević, S.; Jevremović, S.; Subotić, A. Breaking the Dormancy of Snake’s Head Fritillary (Fritillaria meleagris L.) In Vitro Bulbs—Part 2: Effect of GA3 Soaking and Chilling on Sugar Status in Sprouted Bulbs. Plants 2020, 9, 1573. https://doi.org/10.3390/plants9111573
Marković M, Trifunović Momčilov M, Uzelac B, Radulović O, Milošević S, Jevremović S, Subotić A. Breaking the Dormancy of Snake’s Head Fritillary (Fritillaria meleagris L.) In Vitro Bulbs—Part 2: Effect of GA3 Soaking and Chilling on Sugar Status in Sprouted Bulbs. Plants. 2020; 9(11):1573. https://doi.org/10.3390/plants9111573
Chicago/Turabian StyleMarković, Marija, Milana Trifunović Momčilov, Branka Uzelac, Olga Radulović, Snežana Milošević, Slađana Jevremović, and Angelina Subotić. 2020. "Breaking the Dormancy of Snake’s Head Fritillary (Fritillaria meleagris L.) In Vitro Bulbs—Part 2: Effect of GA3 Soaking and Chilling on Sugar Status in Sprouted Bulbs" Plants 9, no. 11: 1573. https://doi.org/10.3390/plants9111573