Integrating Multiple Lines of Evidence to Explore Intraspecific Variability in a Rare Endemic Alpine Plant and Implications for Its Conservation


Abstract
:1. Introduction
2. Results
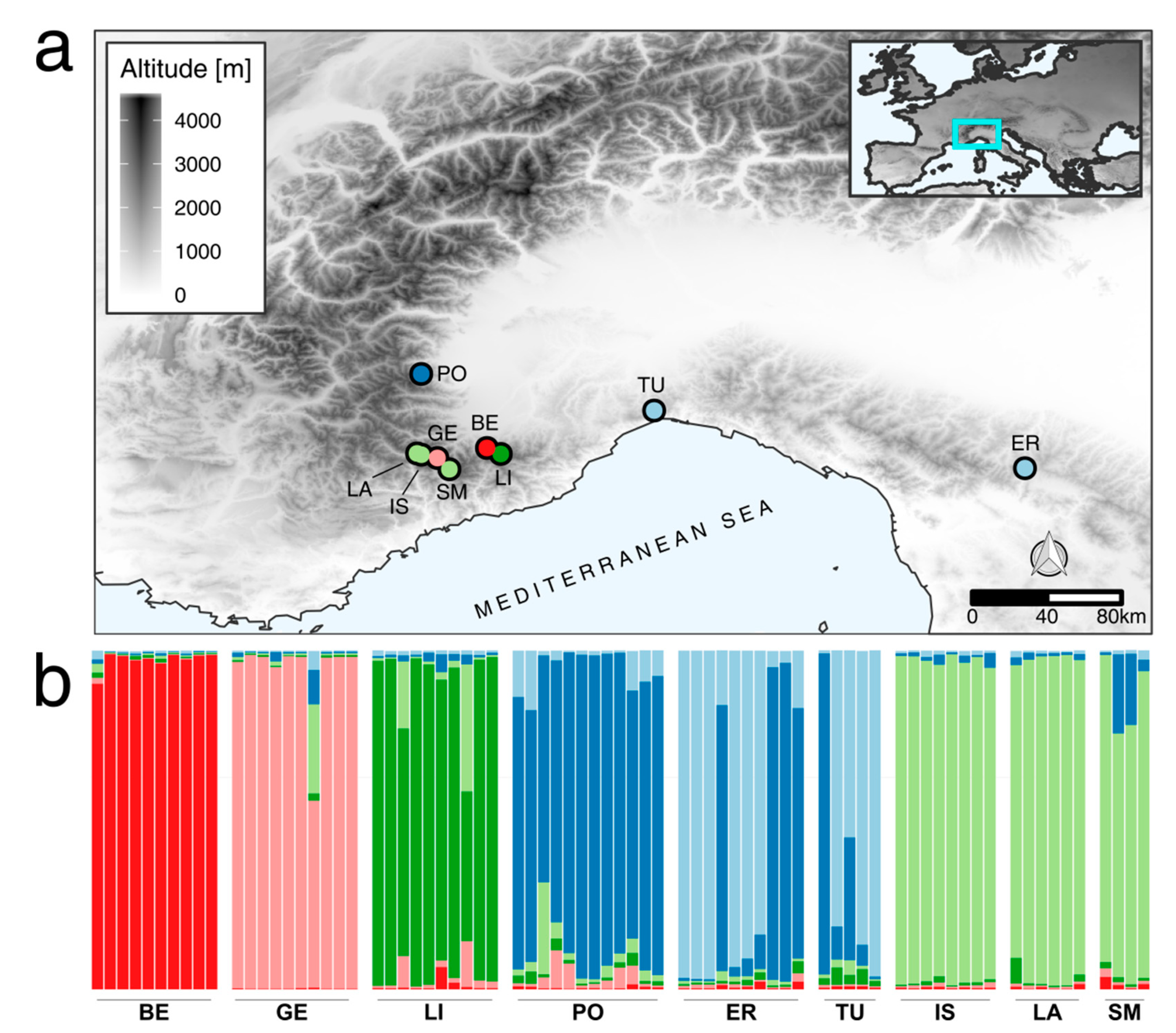
2.1. Genotyping by Fingerprinting
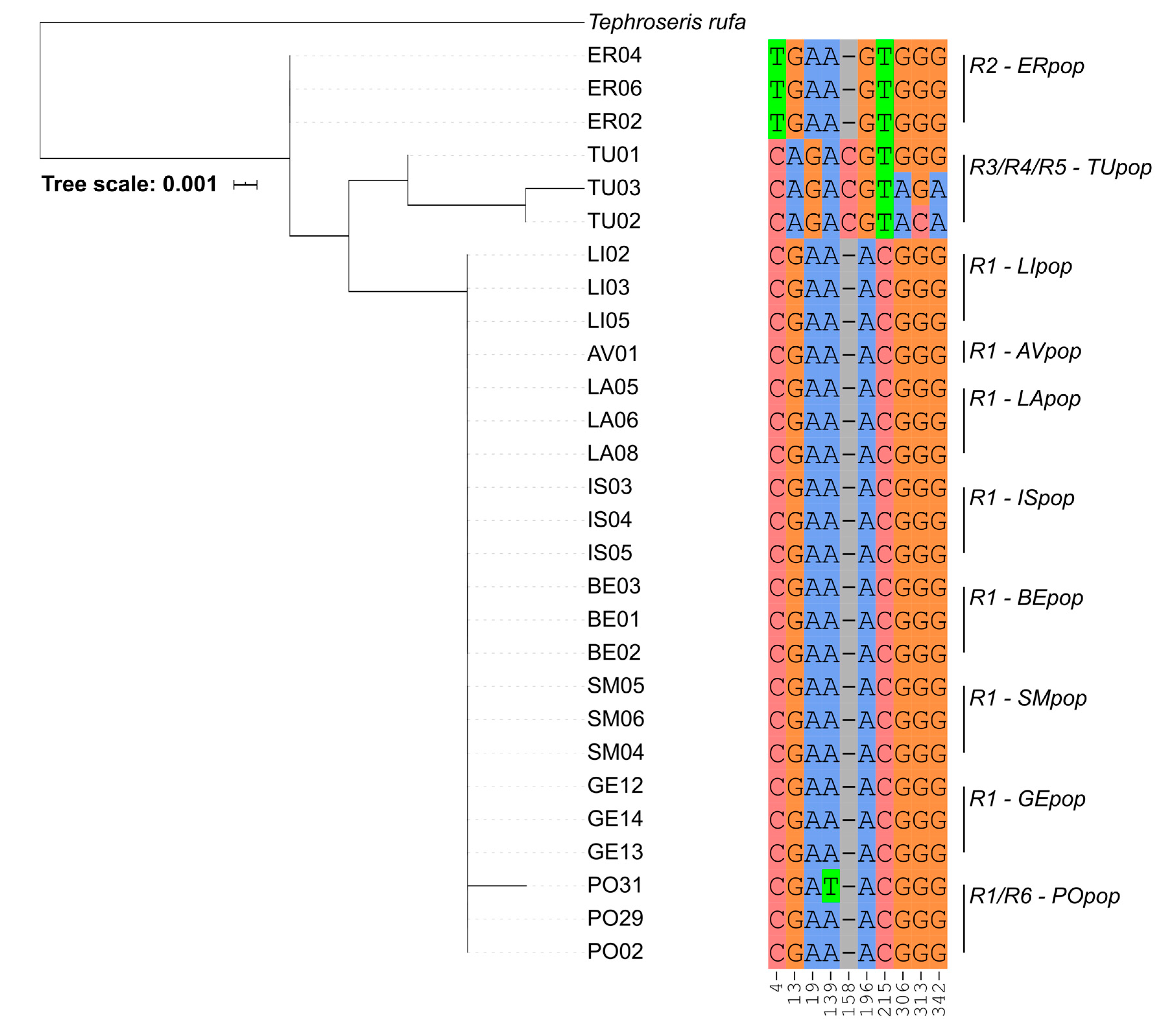
2.2. Phylogenetic Markers
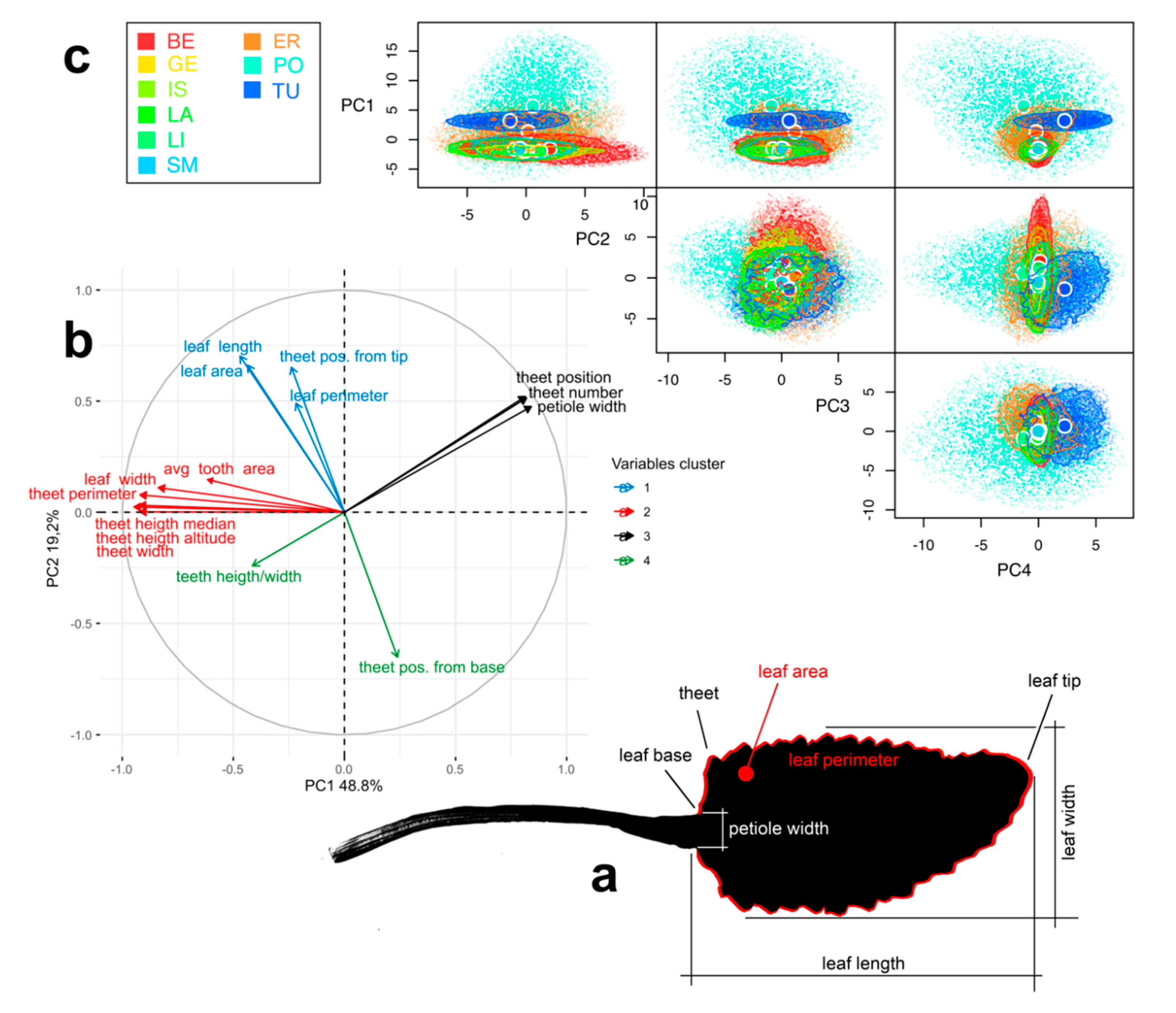
2.3. Leaf Morphology
2.4. Species Distribution Modelling
3. Discussion
3.1. Genetic Analysis
3.2. Morphological Analysis
3.3. Species Distribution Modeling
3.4. Concluding Remarks
4. Materials and Methods
4.1. Sampling Design
4.2. DNA Extraction
4.3. Genotyping by Fingerprinting
4.4. Genotyping by Plastid DNA Markers and ITS-Ribotyping
4.5. Leaf Morphology
4.6. Species Distribution Modeling
4.6.1. Occurrence Data
4.6.2. Calibration Area
4.6.3. Environmental Predictors
4.6.4. Modelling Procedure
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Medail, F.; Quezel, P. Hot-spots analysis for conservation of plant biodiversity in the Mediterranean basin. Ann. Mo. Bot. Gard. 1997, 84, 112. [Google Scholar] [CrossRef]
- Casazza, G.; Barberis, G.; Minuto, L. Ecological characteristics and rarity of endemic plants of the Italian Maritime Alps. Biol. Conserv. 2005, 123, 361–371. [Google Scholar] [CrossRef]
- Médail, F.; Diadema, K. Glacial refugia influence plant diversity patterns in the Mediterranean Basin. J. Biogeogr. 2009, 36, 1333–1345. [Google Scholar] [CrossRef]
- Villemant, C.; Daugeron, C.; Gargominy, O.; Isaia, M.; Deharveng, L.; Judson, M.L.I. The Mercantour/Alpi Marittime all taxa biodiversity inventory (ATBI): Achievements and prospects. Zoosystema 2015, 37, 667–679. [Google Scholar] [CrossRef] [Green Version]
- Mammola, S.; Milano, F.; Vignal, M.; Andrieu, J.; Isaia, M. Associations between habitat quality, body size and reproductive fitness in the alpine endemic spider Vesubia Jugorum. Glob. Ecol. Biogeogr. 2019, 28, 1325–1335. [Google Scholar] [CrossRef]
- Kadereit, J.W. The role of in situ species diversification for the evolution of high vascular plant species diversity in the European Alps—A review and interpretation of phylogenetic studies of the endemic flora of the Alps. Perspect. Plant. Ecol. Evol. Syst. 2017, 26, 28–38. [Google Scholar] [CrossRef]
- Flatscher, R.; García, P.E.; Hülber, K.; Sonnleitner, M.; Winkler, M.; Saukel, J.; Schneeweiss, G.M.; Swetter, P.S. Underestimated diversity in one of the world’s best studied mountain ranges: The polyploid complex of Senecio carniolicus (Asteraceae) contains four species in the European Alps. Phytotaxa 2015, 213, 1–21. [Google Scholar] [CrossRef]
- Taberlet, P.; Zimmermann, N.E.; Englisch, T.; Tribsch, A.; Holderegger, R.; Alvarez, N.; Niklfeld, H.; Coldea, G.; Mirek, Z.; Moilanen, A.; et al. Genetic diversity in widespread species is not congruent with species richness in alpine plant communities. Ecol. Lett. 2012, 15, 1439–1448. [Google Scholar] [CrossRef] [PubMed]
- Stewart, L.; Alsos, I.G.; Bay, C.; Breen, A.L.; Brochmann, C.; Boulanger-Lapointe, N.; Broennimann, O.; Bültmann, H.; Bøcher, P.K.; Damgaard, C.; et al. The regional species richness and genetic diversity of Arctic vegetation reflect both past glaciations and current climate. Glob. Ecol. Biogeogr. 2016, 25, 430–442. [Google Scholar] [CrossRef] [Green Version]
- Bruford, M.W.; Davies, N.; Dulloo, M.E.; Faith, D.P.; Walters, M. Monitoring Changes in Genetic Diversity. In The GEO Handbook on Biodiversity Observation Networks; Springer International Publishing: Cham, Switzerland, 2017; pp. 107–128. [Google Scholar]
- Lamy, T.; Laroche, F.; David, P.; Massol, F.; Jarne, P. The contribution of species—Genetic diversity correlations to the understanding of community assembly rules. Oikos 2017, 126, 759–771. [Google Scholar] [CrossRef]
- Marchesini, A.; Vernesi, C.; Battisti, A.; Ficetola, G.F. Deciphering the drivers of negative species-genetic diversity correlation in Alpine amphibians. Mol. Ecol. 2018, 27, 4916–4930. [Google Scholar] [CrossRef] [PubMed]
- Kahilainen, A.; Puurtinen, M.; Kotiaho, J.S. Conservation implications of species-genetic diversity correlations. Glob. Ecol. Conserv. 2014, 2, 315–323. [Google Scholar] [CrossRef] [Green Version]
- Mráz, P.; Ronikier, M. Biogeography of the Carpathians: Evolutionary and spatial facets of biodiversity. Biol. J. Linn. Soc. 2016, 119, 528–559. [Google Scholar] [CrossRef]
- Schönswetter, P.; Tribsch, A.; Barfuss, M.; Niklfeld, H. Several Pleistocene refugia detected in the high alpine plant Phyteuma globulariifolium Sternb. & Hoppe (Campanulaceae) in the European Alps. Mol. Ecol. 2002, 11, 2637–2647. [Google Scholar] [CrossRef] [Green Version]
- Patsiou, T.S.; Conti, E.; Zimmermann, N.E.; Theodoridis, S.; Randin, C.F. Topo-climatic microrefugia explain the persistence of a rare endemic plant in the Alps during the last 21 millennia. Glob. Chang. Biol. 2014, 20, 2286–2300. [Google Scholar] [CrossRef]
- Mammola, S.; Hormiga, G.; Arnedo, M.A.; Isaia, M. Unexpected diversity in the relictual European spiders of the genus Pimoa (Araneae: Pimoidae). Invertebr. Syst. 2016, 30, 566–587. [Google Scholar] [CrossRef] [Green Version]
- Médail, F.; Verlaque, R. Ecological characteristics and rarity of endemic plants from Southeast France and Corsica: Implications for biodiversity conservation. Biol. Conserv. 1997, 80, 269–281. [Google Scholar] [CrossRef]
- Noble, V.; Diadema, K. Place et Originalité De La Flore; Natural Publications: London, UK, 2011. [Google Scholar]
- Bickford, D.; Lohman, D.J.; Sodhi, N.S.; Ng, P.K.L.; Meier, R.; Winker, K.; Ingram, K.K.; Das, I. Cryptic species as a window on diversity and conservation. Trends Ecol. Evol. 2007, 22, 148–155. [Google Scholar] [CrossRef]
- Lega, M.; Fior, S.; Prosser, F.; Bertolli, A.; Li, M.; Varotto, C. Application of the unified species concept reveals distinct lineages for disjunct endemics of the Brassica repanda (Brassicaceae) complex. Biol. J. Linn. Soc. 2012, 106, 482–497. [Google Scholar] [CrossRef]
- Gugerli, F.; Englisch, T.; Niklfeld, H.; Tribsch, A.; Mirek, Z.; Ronikier, M.; Zimmermann, N.E.; Holderegger, R.; Taberlet, P. Relationships among levels of biodiversity and the relevance of intraspecific diversity in conservation—A project synopsis. Perspect. Plant. Ecol. Evol. Syst. 2008, 10, 259–281. [Google Scholar] [CrossRef]
- De Queiroz, K. Different species problems and their resolution. BioEssays 2005, 27, 1263–1269. [Google Scholar] [CrossRef] [PubMed]
- De Queiroz, K. Species concepts and species delimitation. Syst. Biol. 2007, 56, 879–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlick-Steiner, B.C.; Steiner, F.M.; Seifert, B.; Stauffer, C.; Christian, E.; Crozier, R.H. Integrative Taxonomy: A Multisource Approach to Exploring Biodiversity. Annu. Rev. Entomol. 2010, 55, 421–438. [Google Scholar] [CrossRef] [PubMed]
- Ciccarelli, N.; von Hardenberg, J.; Provenzale, A.; Ronchi, C.; Vargiu, A.; Pelosini, R. Climate variability in north-western Italy during the second half of the 20th century. Glob. Planet. Chang. 2008, 63, 185–195. [Google Scholar] [CrossRef]
- Janišová, M.; Škodová, I.; Hegedüšová, K. Reproductive biology of Tephroseris longifolia subsp. moravica, an endemic taxon of European importance. Seed Sci. Res. 2012, 22, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Biondi, E.; Blasi, C.; Burrascano, S.; Casavecchia, S.; Copiz, R.; Del Vico, E.; Galdenzi, D.; Gigante, D.; Lasen, C.; Spampinato, G.; et al. Manuale italiano di interpretazione degli habitat (Direttiva 92/43/CEE). Available online: https://www.minambiente.it/sites/default/files/archivio/biblioteca/protezione_natura/manuale_interpretazione_habitat_it.pdf (accessed on 7 September 2020).
- Pignatti, S. Flora d’Italia, 1st ed.; Edagricole: Bologna, Italy, 1982. [Google Scholar]
- Bartolucci, F.; Domina, G.; Ardenghi, N.M.G.; Banfi, E.; Bernardo, L.; Bonari, G.; Buccomino, G.; Calvia, G.; Carruggio, F.; Cavallaro, V.; et al. Notulae to the Italian native vascular flora: 5. Ital. Bot. 2018, 5, 71–81. [Google Scholar] [CrossRef]
- Bonafede, F.; Vigonelli, M.; Alessandrini, A. Tephroseris balbisiana (DC.) Holub. In Actaplantarum Notes 1; ArabAFenice: Cuneo, Italy, 2013; p. 82. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Medrano, M.; López-Perea, E.; Herrera, C.M. Population genetics methods applied to a species delimitation problem: Endemic trumpet daffodils (Narcissus section pseudonarcissi) from the southern Iberian Peninsula. Int. J. Plant Sci. 2014, 175, 501–517. [Google Scholar] [CrossRef] [Green Version]
- Naciri, Y.; Linder, H.P. Species delimitation and relationships: The dance of the seven veils. Taxon 2015, 64, 3–16. [Google Scholar] [CrossRef]
- Jackson, N.D.; Carstens, B.C.; Morales, A.E.; O’Meara, B.C. Species delimitation with gene flow. Syst. Biol. 2017, 66, 799–812. [Google Scholar] [CrossRef]
- Habel, J.C.; Zachos, F.E.; Dapporto, L.; Rödder, D.; Radespiel, U.; Tellier, A.; Schmitt, T. Population genetics revisited—Towards a multidisciplinary research field. Biol. J. Linn. Soc. 2015, 115, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia, 2nd ed.; Edagricole-New Business Media: Bologna, Italy, 2017; ISBN 978-88-506-5242-6. [Google Scholar]
- Olšavská, K.; Šingliarová, B.; Kochjarová, J.; Labdíková, Z.; Škodová, I.; Hegedüšová, K.; Janišová, M. Exploring patterns of variation within the central-European Tephroseris longifolia agg.: Karyological and morphological study. Preslia 2015, 87, 163–194. [Google Scholar]
- Skokanová, K.; Šingliarová, B.; Kochjarová, J.; Paule, J. Nuclear ITS and AFLPs provide surprising implications for the taxonomy of Tephroseris longifolia agg. and the endemic status of T. longifolia subsp. moravica. Plant Syst. Evol. 2019, 305, 865–884. [Google Scholar] [CrossRef]
- Nicotra, A.B.; Leigh, A.; Boyce, C.K.; Jones, C.S.; Niklas, K.J.; Royer, D.L.; Tsukaya, H. The evolution and functional significance of leaf shape in the angiosperms. Funct. Plant. Biol. 2011, 38, 535–552. [Google Scholar] [CrossRef] [Green Version]
- Royer, D.L.; Wilf, P.; Janesko, D.A.; Kowalski, E.A.; Dilcher, D.L. Correlations of climate and plant ecology to leaf size and shape: Potential proxies for the fossil record. Am. J. Bot. 2005, 92, 1141–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atkin, O.K.; Loveys, B.R.; Atkinson, L.J.; Pons, T.L. Phenotypic plasticity and growth temperature: Understanding interspecific variability. J. Exp. Bot. 2006, 57, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Royer, D.L.; Meyerson, L.A.; Robertson, K.M.; Adams, J.M. Phenotypic plasticity of leaf shape along a temperature gradient in Acer rubrum. PLoS ONE 2009, 4, e7653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finlayson, C.M.; Davies, G.T.; Moomaw, W.R.; Chmura, G.L.; Natali, S.M.; Perry, J.E.; Roulet, N.; Sutton-Grier, A.E. The Second Warning to Humanity–Providing a Context for Wetland Management and Policy. Wetlands 2019, 39, 1–5. [Google Scholar] [CrossRef]
- Chase, M.W.; Hills, H.H. Silica gel: An ideal material for field preservation of leaf samples for DNA studies. Taxon 1991, 40, 215–220. [Google Scholar] [CrossRef]
- Collard, B.C.Y.; Mackill, D.J. Start Codon Targeted (SCoT) Polymorphism: A simple, novel DNA marker technique for generating gene-targeted markers in plants. Plant. Mol. Biol. Report. 2009, 27, 86–93. [Google Scholar] [CrossRef]
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2014, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Dominant markers and null alleles. Mol. Ecol. Notes 2007, 7, 574–578. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Earl, D.A.; VonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Stucky, B.J. Seqtrace: A graphical tool for rapidly processing DNA sequencing chromatograms. J. Biomol. Tech. 2012, 23, 90–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Gouy, M.; Guindon, S.; Gascuel, O. Sea view version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. Mrbayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Biot, E.; Cortizo, M.; Burguet, J.; Kiss, A.; Oughou, M.; Maugarny-Calès, A.; Gonçalves, B.; Adroher, B.; Andrey, P.; Boudaoud, A.; et al. Multiscale quantification of morphodynamics: Morpholeaf software for 2D shape analysis. Development 2016, 143, 3417–3428. [Google Scholar] [CrossRef] [Green Version]
- Andrey, P.; Maurin, Y. Free-D: An integrated environment for three-dimensional reconstruction from serial sections. J. Neurosci. Methods 2005, 145, 233–244. [Google Scholar] [CrossRef]
- Hutchinson, G.E. Concluding Remarks. Cold Spring Harb. Symp. Quant. Biol. 1957, 22, 415–427. [Google Scholar] [CrossRef]
- Blonder, B. Hypervolume concepts in niche- and trait-based ecology. Ecography 2018, 41, 1441–1455. [Google Scholar] [CrossRef]
- Blonder, B.; Morrow, C.B.; Maitner, B.; Harris, D.J.; Lamanna, C.; Violle, C.; Enquist, B.J.; Kerkhoff, A.J. New approaches for delineating n-dimensional hypervolumes. Methods Ecol. Evol. 2018, 9, 305–319. [Google Scholar] [CrossRef] [Green Version]
- Mammola, S. Assessing similarity of n-dimensional hypervolumes: Which metric to use? J. Biogeogr. 2019, 46, 2012–2023. [Google Scholar] [CrossRef]
- Carvalho, J.C.; Cardoso, P. Decomposing the causes for niche differentiation between species using hypervolumes. BioRxiv 2018, 1–22. [Google Scholar] [CrossRef]
- Mammola, S.; Cardoso, P. Functional diversity metrics using kernel density n-dimensional hypervolumes. BioRxiv Ecol. 2020. [Google Scholar] [CrossRef]
- Cardoso, P.; Rigal, F.; Carvalho, J.C. BAT—Biodiversity Assessment Tools, an R package for the measurement and estimation of alpha and beta taxon, phylogenetic and functional diversity. Methods Ecol. Evol. 2015, 6, 232–236. [Google Scholar] [CrossRef]
- Cardoso, P. RED-IUCN Redlisting Tools—University of Helsinki. Available online: https://researchportal.helsinki.fi/en/publications/red-iucn-redlisting-tools (accessed on 24 February 2020).
- Barve, N.; Barve, V.; Jiménez-Valverde, A.; Lira-Noriega, A.; Maher, S.P.; Peterson, A.T.; Soberón, J.; Villalobos, F. The crucial role of the accessible area in ecological niche modeling and species distribution modeling. Ecol. Model. 2011, 222, 1810–1819. [Google Scholar] [CrossRef]
- Owens, H.L.; Campbell, L.P.; Dornak, L.L.; Saupe, E.E.; Barve, N.; Soberón, J.; Ingenloff, K.; Lira-Noriega, A.; Hensz, C.M.; Myers, C.E.; et al. Constraints on interpretation of ecological niche models by limited environmental ranges on calibration areas. Ecol. Model. 2013, 263, 10–18. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Hengl, T.; De Jesus, J.M.; Heuvelink, G.B.M.; Gonzalez, M.R.; Kilibarda, M.; Blagotić, A.; Shangguan, W.; Wright, M.N.; Geng, X.; Bauer-Marschallinger, B.; et al. SoilGrids250m: Global gridded soil information based on machine learning. PLoS ONE 2017, 12, e0169748. [Google Scholar] [CrossRef] [Green Version]
- Braunisch, V.; Coppes, J.; Arlettaz, R.; Suchant, R.; Schmid, H.; Bollmann, K. Selecting from correlated climate variables: A major source of uncertainty for predicting species distributions under climate change. Ecography 2013, 36, 971–983. [Google Scholar] [CrossRef]
- Mammola, S.; Goodacre, S.L.; Isaia, M. Climate change may drive cave spiders to extinction. Ecography 2018, 41, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Hastie, T.; Tibshirani, R. Generalized Additive Models. In Wiley StatsRef: Statistics Reference Online; John Wiley & Sons, Ltd.: Chichester, UK, 2014. [Google Scholar]
- Elith, J.; Leathwick, J.R.; Hastie, T. A working guide to boosted regression trees. J. Anim. Ecol. 2008, 77, 802–813. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Schapire, E.R. [Internet] Maxent Software for Modeling Species Niches and Distributions (Version 3.4.1). Available online: http://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 1 January 2020).
- Di Cola, V.; Broennimann, O.; Petitpierre, B.; Breiner, F.T.; D’Amen, M.; Randin, C.; Engler, R.; Pottier, J.; Pio, D.; Dubuis, A.; et al. ecospat: An R package to support spatial analyses and modeling of species niches and distributions. Ecography 2017, 40, 774–787. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Araújo, M.B.; New, M. Ensemble forecasting of species distributions. Trends Ecol. Evol. 2007, 22, 42–44. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Sigma | % | Phi | p Value |
|---|---|---|---|---|
| Variations Between Populations | 1.132 | 7.693 | 0.077 | 0.040 |
| Variations Between samples within Populations | 5.382 | 36.570 | 0.396 | 0.001 |
| Variations within samples | 8.203 | 55.736 | 0.443 | 0.001 |
| Total variations | 14.718 | 100.000 |
| Measure | BE | ER | GE | LA | LI | PO | SM | TU | IS |
|---|---|---|---|---|---|---|---|---|---|
| Leaf length (mm) | 177.1 ± 36.5 | 160.4 ± 37.6 | 121.3 ± 16.7 | 119.0 ± 27.9 | 163.6 ± 17.1 | 1419 ± 24.4 | 143.9 ± 24.3 | 118.3 ± 39.9 | 120.0 ± 31.7 |
| Leaf width (mm) | 16.2 ± 5.5 | 41.9 ± 26.1 | 13.9 ± 4.9 | 16.4 ± 4.3 | 66.9 ± 5.7 | 62.2 ± 72.4 | 45.2 ± 3.4 | 35.5 ± 23.8 | 14.6 ± 6.0 |
| Leaf area [mm2] | 883.0 ± 231.4 | 918.1 ± 245.9 | 708.1 ± 115.6 | 656.3 ± 148.8 | 926.9 ± 100.3 | 839.3 ± 228.7 | 716.3 ± 118.4 | 639.1 ± 186.9 | 699.9 ± 183.7 |
| Leaf perimeter (mm) | 258 ± 92 | 256 ± 118 | 174 ± 53 | 192 ± 105 | 208 ± 96 | 205 ± 34 | 279 ± 69 | 225 ± 102 | 174 ± 86 |
| Petiole width (mm) | 0.14 ± 0.04 | 0.07 ± 0.03 | 0.14 ± 0.02 | 0.12 ± 0.03 | 0.09 ± 0.02 | 0.07 ± 0.02 | 0.06 ± 0.02 | 0.09 ± 0.01 | 0.13 ± 0.02 |
| Teeth number | 66 ± 22 | 34 ± 12 | 62 ± 9 | 56 ± 12 | 45 ± 7 | 29 ± 7 | 29 ± 10 | 40 ± 5 | 62 ± 10 |
| Avg tooth position (mm) | 0.07 ± 0.02 | 0.04 ± 0.01 | 0.07 ± 0.01 | 0.06 ± 0.01 | 0.05 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.05 ± 0.01 | 0.07 ± 0.01 |
| Avg tooth width (mm) | 106 ± 11 | 155 ± 53 | 93 ± 8 | 95 ± 12 | 252 ± 4 | 317 ± 269 | 246 ± 12 | 202 ± 75 | 88 ± 10 |
| Avg tooth height altitude (mm) | 19 ± 4 | 59 ± 14 | 17 ± 4 | 13 ± 6 | 37 ± 2 | 48 ± 34 | 40 ± 5 | 30 ± 12 | 15 ± 3 |
| Avg tooth height median (mm) | 28 ± 4 | 76 ± 24 | 25 ± 4 | 21 ± 6 | 71 ± 2 | 71 ± 57 | 55 ± 6 | 40 ± 13 | 22 ± 4 |
| Tooth heigth width ratio | 0.0007 ± 0.0001 | 0.0017 ± 0.0003 | 0.0007 ± 0.0001 | 0.0006 ± 0.0002 | 0.0006 ± 0.0001 | 0.0008 ± 0.0002 | 0.0185 ± 0.0001 | 0.0106 ± 0.0085 | 0.0007 ± 0.0001 |
| Avg tooth area [mm2] | 30.4 ± 9.9 | 118.5 ± 72.3 | 26.1 ± 6.7 | 16.6 ± 11.2 | 29.1 ± 3.3 | 76.3 ± 15.4 | 11.8 ± 11.0 | 12.1 ± 3.2 | 16.1 ± 4.7 |
| Avg tooth perimeter (mm) | 12.8 ± 1.4 | 23.6 ± 6.8 | 11.4 ± 1.1 | 11.1 ± 1.8 | 30.8 ± 0.6 | 39.5 ± 33.5 | 29.0 ± 1.7 | 23.8 ± 7.8 | 10.6 ± 1.3 |
| Avg tooth position from tip (mm) | 0.23 ± 0.01 | 0.2 ± 0.03 | 0.22 ± 0.01 | 0.21 ± 0.01 | 0.22 ± 0.01 | 0.22 ± 0.04 | 0.21 ± 0.01 | 0.21 ± 0.02 | 0.22 ± 0.01 |
| Avg tooth position from base (mm) | 0.21 ± 0.01 | 0.23 ± 0.03 | 0.21 ± 0.01 | 0.22 ± 0.01 | 0.21 ± 0.01 | 0.21 ± 0.04 | 0.22 ± 0.01 | 0.22 ± 0.02 | 0.21 ± 0.01 |
| Morphospace volume | 725.68 | 4597.89 | 52.79 | 89.72 | 294.76 | 31.16 | 41056.2 | 142.77 | 119.43 |
| Morphospace dispersion | 5.54 | 6.84 | 3.17 | 3.92 | 2.62 | 11.24 | 3.39 | 5.21 | 4.10 |
| Variable | Generalized Additive Model | Generalized Boosted Regression Model | MaxEnt |
|---|---|---|---|
| Precipitation seasonality | edf = 5.26; χ2 = 45.46; p < 0.001 | 20.84 | 15.9 |
| Daily temperature range | edf = 2.48; χ2 = 9.27; p = 0.03 | 9.44 | 7.9 |
| Solar Radiation | 0.02 ± 0.04; p < 0.001 | 48.43 | 73.8 |
| Organic material content | –0.03 ± 0.02; p = 0.11 | 7.93 | 0.5 |
| pH | –0.11 ± 0.08; p = 0.15 | 13.34 | 1.9 |
| Pop ID | Site | Locality, Province, Country | Date | Number of Individuals | SCoTs Genotyping | Leaf Morphology | Ribotyping |
|---|---|---|---|---|---|---|---|
| AV | Lago Aver Superiore | Vinadio, CN, IT | 17 July | - | - | - | 1 |
| BE | Vallone S. Bernardo | Limone P.te, CN, IT | 19 June | 30 | 10 | 9 | 3 |
| ER | Fontana Bura | San Benedetto Val Sambro, BO, IT | 19 June | 100 | 10 | 19 | 3 |
| GE | Pian della Casa | Valdieri, CN, IT | 19 July | 12 | 10 | 12 | 3 |
| IS | Isola 2000 | Isola, 06, FR | 19 September | 15 | 8 | 11 | 3 |
| LA | La Lausetta | Isola, 06, FR | 19 September | 15 | 6 | 9 | 3 |
| LI | Porta Sestrera | Chiusa di Pesio, CN, IT | 19 July | 50 | 10 | 8 | 3 |
| PO | Rio Ratisin | Crissolo, CN, IT | 19 July | 100 | 12 | 9 | 3 |
| SM | La Pointe | Saint-Martin Vesubie, 06, FR | 19 September | 15 | 4 | 7 | 3 |
| TU | Passo del Turchino | Masone, GE, IT | 19 July | 20 | 5 | 11 | 3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adamo, M.; Mammola, S.; Noble, V.; Mucciarelli, M. Integrating Multiple Lines of Evidence to Explore Intraspecific Variability in a Rare Endemic Alpine Plant and Implications for Its Conservation. Plants 2020, 9, 1160. https://doi.org/10.3390/plants9091160
Adamo M, Mammola S, Noble V, Mucciarelli M. Integrating Multiple Lines of Evidence to Explore Intraspecific Variability in a Rare Endemic Alpine Plant and Implications for Its Conservation. Plants. 2020; 9(9):1160. https://doi.org/10.3390/plants9091160
Chicago/Turabian StyleAdamo, Martino, Stefano Mammola, Virgile Noble, and Marco Mucciarelli. 2020. "Integrating Multiple Lines of Evidence to Explore Intraspecific Variability in a Rare Endemic Alpine Plant and Implications for Its Conservation" Plants 9, no. 9: 1160. https://doi.org/10.3390/plants9091160