Patterning the Axes: A Lesson from the Root



1
Department of Biology, University of Pisa, via L. Ghini, 13-56126 Pisa, Italy
2
Dipartimento di Biologia e Biotecnologie, Laboratory of Functional Genomics and Proteomics of Model Systems, Università di Roma “Sapienza”, via dei Sardi, 70-00185 Rome, Italy
*
Author to whom correspondence should be addressed.
Plants 2019, 8(1), 8; https://doi.org/10.3390/plants8010008
Submission received: 22 November 2018
/
Revised: 19 December 2018
/
Accepted: 24 December 2018
/
Published: 31 December 2018
(This article belongs to the Special Issue Plant Development and Organogenesis: From Basic Principles to Applied Research)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:How the body plan is established and maintained in multicellular organisms is a central question in developmental biology. Thanks to its simple and symmetric structure, the root represents a powerful tool to study the molecular mechanisms underlying the establishment and maintenance of developmental axes. Plant roots show two main axes along which cells pass through different developmental stages and acquire different fates: the root proximodistal axis spans longitudinally from the hypocotyl junction (proximal) to the root tip (distal), whereas the radial axis spans transversely from the vasculature tissue (centre) to the epidermis (outer). Both axes are generated by stereotypical divisions occurring during embryogenesis and are maintained post-embryonically. Here, we review the latest scientific advances on how the correct formation of root proximodistal and radial axes is achieved.
1. Introduction
One of the most intriguing questions in developmental biology is how the body plan is established. To answer this question, for decades scientists have focused on the formation of developmental axes, utilizing different model systems. Most of our knowledge on axes formation derives from studies on vertebrate limb development [1,2]. However, these systems present several limitations due to their complex structure that limits analysis at a single cell resolution. On the contrary, plant roots display a simple and organized structure, where cell lineages are easily distinguishable by shape and position [3,4]. Furthermore, due to the presence of the cell wall, plant cells do not migrate; hence, cell fate and identity can be easily followed during different stages of organ development [3,4,5]. For these reasons, roots represent a powerful tool to study the molecular mechanisms on how developmental axes are established and maintained. Roots can be represented as a series of concentric cylinders, where epidermis is the outermost tissue while the vasculature bundles lie in the centre [4] (Figure 1). Roots display two main developmental axes: the proximodistal axis, extending longitudinally from the root–shoot junction (proximal) to the root apex (distal); the radial axis, spreading transversally from the vasculature bundles to the epidermis [4] (Figure 1). Like other animal model systems, root axes are established during embryogenesis and maintained post-embryonically by the activity of meristems [3,4,5,6]. Meristems are localized structures that sustain post embryonic indeterminate plant organ growth due to the activity of stem cell niches (SCNs) [3,4,5]. In the Arabidopsis root meristem, there are five sets of stem cells (initials) that give rise to all root tissues: epidermis and lateral root cap initials (EPI LRC STEM CELLS), cortex and endodermis initial (CEI), pericycle initials, vasculature initials and distally columella initials (Figure 1) [3,4]. These sets of stem cells surround the QC (Quiescent Center) which maintains, by contacting them, their stem cell identity (Figure 1) [7,8]. The stem cells divide asymmetrically and anticlinally generating daughter cells (Figure 1) that generates both the proximodistal and radial axes through stereotypical cell divisions. Along the proximodistal axis, the stem cell daughters divide anticlinally a fixed number of times, generating the division zone of the meristem. In the proximal area of the meristem, those cells cease to divide when they reach a boundary called the transition zone (TZ). Here they start to elongate and differentiate, generating the elongation/differentiation zone [3,5,9,10] (Figure 1). In this zone, cells acquire characteristic differentiation features such as root hairs for the epidermis or tracheids for the vascular cells [11,12]. The position of the TZ is fundamental for proximodistal axis specification, as it marks the boundary between undifferentiated and differentiated cells [9,13].
Radially, most of the stem cells daughters divide periclinally, giving rise to two tissues with different identities. For example, cortex and endodermis are derived from the periclinal division of the daughter of the cortex and endodermis initial (CEI), whereas epidermis and lateral root cap originate from the EPI LRC initial [14,15,16]. The control of the asymmetric divisions occurring in the stem cell daughters is key for the correct patterning of the radial axis. Indeed, alteration of the position and timing of those divisions causes the formation of aberrant body plan and shape (Figure 1).
Thanks to the generation of new tools and the improvement of molecular methodologies, several molecular mechanisms underlying the establishment and maintenance of the proximodistal and radial are in the process of being discovered and fully comprehended. In this review, we report the current view on how these two axes are patterned.
2. Root Radial Axis
The root radial axis organization depends on the coordinated activity of periclinal divisions of the stem cell daughters. One of the most studied mechanisms patterning the radial axis is the one controlling the formation of the cortex and the endodermis root tissues. These tissues originate from a single stem cell (CEI) that firstly divides anticlinally, thereby generating a daughter cell (CEID). This cell divides periclinally, generating the cortex and the endodermis that together are called Ground Tissue (GT). GT specification starts in the embryo when a periclinal division at early globular embryonic stage separates the pro-vasculature tissues from the GT precursor cell. Only later, at the heart embryonic stage, a pro-GT division leads to the specification of the cortex and the endodermis [6]. It was recently shown that the establishment of the pro-GT at early embryonic stages depends on the plant hormone auxin. A maximum level of auxin activity driven by the auxin responsive factor MONOPTEROS/AUXIN RESPONSIVE FACTOR 5 (MP/ARF5) in the GT precursor cells is required for GT formation [17]. Indeed, mp null mutants display impaired GT establishment [17].
Two GRAS family transcription factors, SHORTROOT (SHR) and SCARECROW (SCR), are involved in the formation of the cortex and endodermis layers, as they are necessary and sufficient to promote the CEID periclinal division [18,19,20,21]. SHR is a mobile transcription factor expressed in the vasculature. SHR moves toward the CEID, CEI and endodermis via plasmodesmata, where it is sequestered into the nucleus [22,23] (Figure 2 and Figure 3). SHR movements restriction is fundamental for GT patterning, as overexpression of SHR results in additional GT layer formation [24,25]. In the vasculature, SHR is maintained mostly in the cytoplasm by the activity of SCARECROW-LIKE23 (SCL23) [26]. In the CEID, SHR forms a molecular complex with SCR and it is sequestered in the nucleus by the activity of SCR. In the nucleus, SHR/SCR complex sustains the expression of SCR itself and induces the expression of INDETERMINATE DOMAIN C2H2 zinc finger (BIRD) transcription factors such as JACKDAW (JKD), NUTCRACKER (NUC) and MAGPIE (MGP) [21,27,28,29,30,31]. BIRD proteins physically interact with the SHR/SCR complex, restricting SHR movements to the stele [27,29,32]. SHR/SCR complex promotes the expression of the cell cycle regulator CYCLIND6 (CYCD6;1) in the CEID, inducing here a periclinal division [33,34]. Via a combination of mathematical modelling and wet biology, it has been proposed that the SHR/SCR/CYCD6;1 module, together with the cell cycle inhibitor RETINOBLASTOMA-RELATED (RBR) protein, acts via a bistable circuit to regulate the CEID asymmetric division [33,34,35]. In the CEI, the CYCD6;1 together with the CDKB1;1 (CYCLIN DEPENDENT KINASE 1;1) or CDKB1;2 induces the phosphorylation of RBR, reducing its activity in the CEID, thus promoting the periclinal division [34]. Auxin is a key factor for the promotion of this periclinal division. Indeed, an auxin maximum in the CEI promotes CYCD6;1 expression [34]. On the contrary, RBR was shown to directly interact with SCR, reducing its transcriptional activator activity in the endodermis [36]. The RBR and SCR interaction, together with the activity of the RBR regulator CYCD6;1, limits the asymmetric cell division in the SCN, thus allowing the formation of the endodermal and cortical layers (Figure 2). Recently, a sophisticated molecular mechanism was proposed for a SHR/SCR-dependent switching on of the CYCD6;1 involving the RNA POLYMERASE II cofactor Mediator. Depending on the SHR concentration, SCR interacts with the subunit 31 of the Mediator to promote CYCD6;1 expression [37].
Once CEID divides, several factors coordinate the formation of the cortical and endodermal layers. SHR also promotes endodermal fate, as suggested by the loss of endodermis identity in shr mutants [18,28,38,39]. It was shown that BIRD proteins, other than regulating SHR movements, play a key role in determining cortical identity, as multiple mutant combinations of BIRD members show GT with no cortical identity [32]. Therefore, the combined activity of SHR, SCR and BIRD proteins is necessary to pattern the GT. Interestingly SHR and SCR are involved only in the maintenance of GT and not in its establishment. Once MP initiates the ground tissue lineage, it acts upstream of the SHR/SCR module, controlling ground tissue patterning and maintenance.
SCHIZORIZA (SCZ), a member of the Heat Shock Transcription Factor family, is also involved in GT patterning and its activity depends on SHR and SCR [40,41,42]. Interestingly, SCZ is expressed in all root tissues except for the lateral root cap. It was shown that SCZ, together with JKD, MGP, and NUC proteins, promotes cortical identity (Figure 2 and Figure 3) [32]. It must be pointed out that scz mutants present additional tissue layers with mixed cortical, endodermal and epidermal identities, suggesting a role for this gene in tissue fate separation [40,41,42]. The analysis of SCZ target genes will help to establish an understanding of how SCZ patterns the radial axis.
Besides organizing the GT, SHR/SCR complex is involved in vasculature patterning. Arabidopsis root vasculature consists of an inner xylem bundle (metaxylem in the centre, protoxylem aside) with two juxtaposed phloematic bundles [4] (Figure 3). The formation and development of the metaxylem depends on the redundant activity of the Class III Homeodomain Leucine Zipper (HD-ZIPIII) members, a family of five transcription factors targeted by microRNA 165/6 (miR165/6). SHR/SCR promotes in the endodermis the expression of miR165/6 that, moving toward the stele via plasmodesmata, generates an opposite gradient of the HD-ZIPIII proteins, with a maximum in the metaxylem and a minimum in the endodermis [22,43,44] (Figure 1 and Figure 3). The formation of a radial gradient of HD-ZIPIIIs is sufficient to pattern the xylem fate specification, as high HD-ZPIIIs levels promote metaxylem formation, whereas low ones promote protoxylem [43,44] (Figure 3). In the stele, HD-ZIPIIIs control the biosynthesis and activity of the phytohormone cytokinin, which in turn regulates auxin distribution and signalling [44,45]. This finely regulated mechanism is sufficient to pattern the stele.
It was recently shown that miR165/6 distribution is not only crucial for vasculature development but also for GT patterning [46,47] (Figure 1 and Figure 2). A mir165/6-dependent minimum of HD-ZIPIIIs in the CEI/CEID and endodermis is required to restrict the number of cortical layers, as miR165/6-insensitive HD-ZIPIII mutants show additional cortical layers [47]. HD-ZIPIIIs expression in the GT results in ectopic CYCD6;1 activation, prompting additional GT divisions. Intriguingly, the HD-ZIPIII member PHABULOSA (PHB) indirectly sustains CYCD6;1 expression in a SHR-independent manner, but how PHB triggers periclinal divisions is still not known [46,47,48]. It was recently shown that PHB directly targets MP to pattern the vasculature tissue [49]. Nevertheless, whether PHB/MP circuit is important for GT establishment is not known.
It was recently shown that SHR, together with SCR, also specifies endodermis differentiation. Functional endodermis is characterized by Casparian strips, lignified structures deposited on the radial and transverse side of the endodermal cell wall [24]. SHR directs the formation of the Casparian strips by inducing the MYB DOMAIN transcription factor MYB36 and the receptor-like kinase SGN1 and SGN3 [50,51] (Figure 3). In endodermal cells, MYB36 induces the expression of the transmembrane proteins CASPARIAN STRIP MEMBRANE DOMAIN PROTEIN (CASP) [52,53], which are involved in the recruitment of lignin synthesis enzymes on the plasma-membrane. SGN1/3 position CASP proteins on the plasma membrane (Figure 3). Nonetheless, SHR promotes the formation of a non-functional Caspary band and it requires the activity of vasculature-deriving small peptides, CASPARIAN STRIP INTEGRITY FACTOR (CIF), to generate a functional strip [50,51] (Figure 3). Hence, SHR and SCR constitute an important module to control endodermis differentiation.
3. Root Proximodistal Axis
Different from the radial axis, where most of the cells show different identities but similar developmental stages, along the proximodistal axis cells display different stages of development. Positioning of the TZ plays a key role for patterning the proximodistal axis, as the TZ separates proliferating meristematic cells from the elongated ones [54] (Figure 1 and Figure 4). The position of the TZ depends on the dynamic equilibrium between cell division and cell differentiation; alterations of this equilibrium cause the TZ position to shift toward the distal or the proximal area of the root, thus varying the proximodistal zonation.
Auxin plays a pivotal role in establishing the root proximodistal axis, acting as a local morphogen [55,56]. Already at the globular stage of embryogenesis, a maximum of auxin in the basal pole of the embryo determines the position of the SCN [57]. This auxin maximum is controlled by the activity of the auxin polar transport efflux facilitators PIN FORMED (PINs) that distribute this hormone [58,59,60]. Auxin signalling is necessary for the formation of the SCN [55,61]. Interestingly, mp loss of function mutants or gain of function mutants of its repressor, the AUX/IAA auxin signalling repressor BODENLOS (BDL), display no root formation [61,62,63]. Together with auxin, four AP2 transcription factors, PLETHORA 1,2,3 and 4 (PLT1,2,3 and 4), control stem cell activity and root growth from embryogenesis onwards [64,65]. Multiple combinations of the loss of function mutants plt1,2,3,4 show no root SCN formation, whereas constitutive expressions of PLT genes induces shoot homeotic transformation into root [65]. The GATA transcription factor HANABA TARANU/MONOPOLE (HAN) forms the boundary between embryonic apical and basal pole, confining PLT expression and the auxin maximum to the root precursors domain [66]. PLTs also play also an active role in the repression of the apical pole identity. Indeed, PLT, together with miR165/6, represses the apical embryonic SCN formation by restricting HD-ZIPIIIs expression [67,68]. Lack of this repression leads to the homeotic transformation of the root into shoot, suggesting a master role for PLT in determining the embryonic apical-basal axis [68]. It has been recently demonstrated that PLT regulates the expression of HAN and the synthesis of auxin via direct control of YUCCA3, a gene involved in auxin biosynthesis [69]. One possibility is that PLTs regulate the expression of genes involved in apical fate determination, such as HD-ZIPIII directly acting on HAN or on auxin synthesis. Future studies will clarify this point.
Post-embryonically, PLTs and auxin are required to maintain SCN activity in the root, forming a gradient with a maximum in this zone (Figure 4) [55,64,65,70]. Ectopic inductions of auxin or PLTs maximum in the meristem convert other cell types in stem cells, underlying the importance of these maxima for stem cell specification. Post-embryonically, PLTs mRNAs and auxin are distributed in a gradient along the meristematic proximodistal axis [70]. PLTs and auxin gradients are strictly interconnected. Indeed, auxin promotes PLTs expression, whereas PLTs regulate auxin distribution, controlling PINs expression and auxin biosynthesis [58,64,69]. Different concentrations of auxin or PLTs result in different developmental outputs, i.e., high PLTs and auxin levels are necessary for SCN specification, whereas minimum auxin and PLT levels are necessary to induce cell differentiation at the TZ [55,64,65,70,71] (Figure 4). Recent studies have shown that the PLTs gradient along the proximodistal axis is partially independent from auxin, while the capacity of these proteins to diffuse along this axis plays an important role [65,70].
PIN-dependent polar auxin transport is necessary to position the auxin maximum at the root distal part [55,59] and the manner in which an auxin minimum is positioned at the proximal TZ has recently been elucidated. Indeed, the role of the plant hormone cytokinin in shaping the auxin gradient has been revealed. To position this minimum cytokinin triggers a module that involves the cytokinin receptor AHK3 (ARABIDOPSIS HISTIDINE KINASE 3), the cytokinin-dependent transcription factor ARR1 (ARABIDOPSIS RESPONSE REGULATOR1), the auxin signalling repressor SHY2/IAA3 (SHORT HYPOCOTYL2/INDOLE-3-ACETIC ACID INDUCIBLE 3) and the auxin catabolic enzyme GH3.17 (GRETCHEN HAGEN 3.17) [54,71,72,73] (Figure 4). Cytokinin via AHK3 activates ARR1 directly inducing the expression of SHY2 in the vasculature at the TZ. Here, SHY2 downregulates PINs expression, thus reducing the shoot to root auxin efflux and, hence, cell division activity; auxin instead induces proteasome-dependent SHY2 degradation, supporting PINs expression [54,72,73,74] (Figure 4). At the same time, cytokinin via ARR1 induces the GH3.17 gene in the lateral root cap and epidermis [71], where it mediates auxin degradation (Figure 4). The coordinated regulation of both auxin signalling and catabolism localizes a developmental instructive auxin minimum that positions the TZ [71].
The coordination of SCN activity with cell differentiation process at the TZ is fundamental for proper proximodistal axis patterning. This spatial coordination is controlled by SCR and SHR, which repress cytokinin activity in the SCN, thus controlling auxin production [34,75,76]. In particular, SCR represses ARR1 expression in the QC, which in turn positively regulates the expression of the auxin biosynthesis gene ASB1 (ANTHRANILATE SYNTHASE BETA SUBUNIT 1). Since ARR1 is induced by auxin at the TZ SCR, by regulating auxin biosynthesis in the QC, this controls stem cell division in the SCN and cell differentiation at the TZ [34].
Considering the key role of cytokinin in positioning the TZ, the regulation of cytokinin synthesis is fundamental for proper root patterning [13,24]. ISOPENTENYL TRANSFERASE (IPT) enzymes are key regulators of cytokinin synthesis [24]. In the meristem, the transcription factor PHB promotes cytokinin synthesis via direct induction of IPT1 and IPT7 [77]. The PHB-dependent cytokinin production is sufficient to activate the ARR1/SHY2 module, positioning the TZ. Intriguingly, ARR1 represses both PHB and its repressors miR165/6 expression, triggering a negative incoherent feedforward loop that finely tunes cytokinin levels and prevents meristem from differentiating [77].
Recently, it was demonstrated that cytokinin also promotes the transition of cells from the meristematic zone to the elongation zone, regulating apoplastic acidification and cell expansion. ARR1 directly regulates enzymes involved in cellular expansion such as the α-expansin EXPANSIN1 (EXPA1), controlling cell wall loosening and the plasma membrane H+-ATPases (HA) 1 and 2 (AHA1 and AHA2) that transport protons (H+) out of the cell. Interestingly, expa1 mutants show a shift of the TZ toward the root proximal zone without interfering with the final cell size, suggesting that EXPA1-dependent cell expansion is mostly controlling the timing of cell exit from the division zone more than the final cell size itself [78] (Figure 4).
4. Future Perspectives
In recent years we have increased our understanding of the mechanisms controlling the formation of both radial and proximodistal axes of the root apical meristem. It is interesting to notice that the molecular mechanisms patterning both the radial and the proximodistal axes involve the same main players (i.e., auxin, PHB and SHR/SCR). Future studies will elucidate how the mechanisms controlling the development of these two axes coordinate in order to generate a structured body plan. Moreover, the effectors of the master genes governing the zonation of both those axes are starting to be understood, but several players are still missing and will be discovered in the future. In this optic, with respect to the proximodistal axis, the lists of targets of ARR1 and PLTs were published. This knowledge will allow us to better understand how those genes are interconnected and how they coordinate to ensure continuous growth.
In multicellular organisms, cell elongation is accompanied by endoreduplication, genome duplication in absence of mitosis [79,80]. Similarly, it was shown that cells at the TZ show enhanced the number of genome copies compared to their meristematic progenitors. Moreover, cytokinin is known to promote endoreduplication [81]. It will be interesting in the future to investigate the role of endoreduplication in patterning the root proximodistal axis.
Most of the molecular mechanisms patterning the root axes were discovered in Arabidopsis. In recent years, several variations in those mechanisms were found to be the basis for interspecific variability in plants.
Indeed, root axis structure are largely variable among species. For example, the root radial axis is extremely variable, as number and features of tissue layers is strictly dependent on the species [24,82,83]. One of the tissues along the radial axis that is most variable between species is the cortex [24]. For example, Cardamine hirsuta, a close relative of Arabidopsis, displays two cortex layers. It was recently shown that the second cortical layer of Cardamine emerges from the activity of a developmental domain absent in Arabidopsis, the cortex endodermis mixed identity tissue (CEM). Activity of HD-ZIPIII in this domain is crucial for the formation of the second cortical, as knockdown of these genes results in the loss of this additional layer [47]. How PHB controls the CEM periclinal division is still not known. In Arabidopsis, ectopic expression of PHB indirectly promotes CYCD6;1 expression independently from SHR, therefore PHB might control CEM division acting on this gene. Moreover, whether this mechanism is conserved in other distant relatives with multiple cortical layers or whether this mechanism adds to the one controlled by SHR is still not known. Future research will allow us to uncover the answers to this interesting question. It was shown that SHR and SCR also play a key role in patterning the differences in radial axis anatomy among species. The multi cortical layered species Oryza sativa (rice), indeed, maintains the SHR/SCR interaction, but SHR movements are subject to lower restriction, for promoting multiple cortical layers formation [38,84].
The mechanisms governing root axis formation and maintenance in Arabidopsis might be valid for most of the species but may not be universal. It was shown that in the root of most of the species, auxin controls cell division, whereas cytokinin controls differentiation. However, in the fern Azolla filiculoides, cytokinin promotes cell division, whereas auxin promotes cell differentiation [85]. Analysis of the molecular mechanisms controlling root axes in species other than Arabidopsis will permit us to understand how and when these mechanisms arose and diverged. In this optic, utilization of close relatives of Arabidopsis might allow us to understand this crucial point.
Author Contributions
R.D.M., S.S. and R.D.I. wrote the manuscript and prepared the figures.
Funding
This work was supported by a FIRB2013 (FUTURO IN RICERCA) to R.D.I. and by The European Research Council (grant number 260368) to S.S.
Acknowledgments
We extend our sincere thanks to P. Costantino, N. Svolacchia, E. Salvi, G. Bertolotti and A. Rocchetti for critical comments on the manuscript.
Conflicts of Interest
The authors claim no conflict of interest.
References
- Tabin, C.; Wolpert, L. Rethinking the proximodistal axis of the vertebrate limb in the molecular era. Genes Dev. 2007, 21, 1433–1442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuniga, A. Next generation limb development and evolution: Old questions, new perspectives. Development 2015, 142, 3810–3820. [Google Scholar] [CrossRef]
- Dolan, L.; Janmaat, K.; Willemsen, V.; Linstead, P.; Poethig, S.; Roberts, K.; Scheres, B. Cellular organisation of the Arabidopsis thaliana root. Development 1993, 119, 71–84. [Google Scholar] [PubMed]
- Scheres, B.; Benfey, P.; Dolan, L. Root development. Arabidopsis Book 2002, 1, e0101. [Google Scholar] [CrossRef] [PubMed]
- Heidstra, R.; Sabatini, S. Plant and animal stem cells: Similar yet different. Nat. Rev. Mol. Cell Biol. 2014, 15, 301–312. [Google Scholar] [CrossRef]
- Ten Hove, C.A.; Lu, K.J.; Weijers, D. Building a plant: Cell fate specification in the early Arabidopsis embryo. Development 2015, 142, 420–430. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg, C.; Willemsen, V.; Hendriks, G.; Weisbeek, P.; Scheres, B. Short-range control of cell differentiation in the Arabidopsis root meristem. Nature 1997, 390, 287–289. [Google Scholar]
- Sabatini, S.; Heidstra, R.; Wildwater, M.; Scheres, B. Scarecrow is involved in positioning the stem cell niche in the Arabidopsis root meristem. Genes Dev. 2003, 17, 354–358. [Google Scholar] [CrossRef]
- Perilli, S.; Di Mambro, R.; Sabatini, S. Growth and development of the root apical meristem. Curr. Opin. Plant Biol. 2012, 15, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Di Mambro, R.; Sabatini, S. Developmental analysis of Arabidopsis root meristem. Methods Mol. Biol. 2018, 1761, 33–45. [Google Scholar]
- Drapek, C.; Sparks, E.E.; Benfey, P.N. Uncovering gene regulatory networks controlling plant cell differentiation. Trends Genet. 2017, 33, 529–539. [Google Scholar] [CrossRef]
- Benfey, P.N. Defining the path from stem cells to differentiated tissue. Curr. Top. Dev. Biol. 2016, 116, 35–43. [Google Scholar] [PubMed]
- Dello Ioio, R.; Linhares, F.S.; Sabatini, S. Emerging role of cytokinin as a regulator of cellular differentiation. Curr. Opin. Plant Biol. 2008, 11, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Kumpf, R.P.; Nowack, M.K. The root cap: A short story of life and death. J. Exp. Bot. 2015, 66, 5651–5662. [Google Scholar] [CrossRef] [PubMed]
- Pauluzzi, G.; Divol, F.; Puig, J.; Guiderdoni, E.; Dievart, A.; Perin, C. Surfing along the root ground tissue gene network. Dev. Biol. 2012, 365, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.W.; Lim, J. Control of asymmetric cell divisions during root ground tissue maturation. Mol. Cells 2016, 39, 524–529. [Google Scholar] [CrossRef] [PubMed]
- Moller, B.K.; Ten Hove, C.A.; Xiang, D.; Williams, N.; Lopez, L.G.; Yoshida, S.; Smit, M.; Datla, R.; Weijers, D. Auxin response cell-autonomously controls ground tissue initiation in the early Arabidopsis embryo. Proc. Natl. Acad. Sci. USA 2017, 114, E2533–E2539. [Google Scholar] [CrossRef] [PubMed]
- Heidstra, R.; Welch, D.; Scheres, B. Mosaic analyses using marked activation and deletion clones dissect Arabidopsis scarecrow action in asymmetric cell division. Genes Dev. 2004, 18, 1964–1969. [Google Scholar] [CrossRef]
- Helariutta, Y.; Fukaki, H.; Wysocka-Diller, J.; Nakajima, K.; Jung, J.; Sena, G.; Hauser, M.T.; Benfey, P.N. The short-root gene controls radial patterning of the Arabidopsis root through radial signaling. Cell 2000, 101, 555–567. [Google Scholar] [CrossRef]
- Nakajima, K.; Sena, G.; Nawy, T.; Benfey, P.N. Intercellular movement of the putative transcription factor shr in root patterning. Nature 2001, 413, 307–311. [Google Scholar] [CrossRef]
- Clark, N.M.; Hinde, E.; Winter, C.M.; Fisher, A.P.; Crosti, G.; Blilou, I.; Gratton, E.; Benfey, P.N.; Sozzani, R. Tracking transcription factor mobility and interaction in Arabidopsis roots with fluorescence correlation spectroscopy. Elife 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Vaten, A.; Dettmer, J.; Wu, S.; Stierhof, Y.D.; Miyashima, S.; Yadav, S.R.; Roberts, C.J.; Campilho, A.; Bulone, V.; Lichtenberger, R.; et al. Callose biosynthesis regulates symplastic trafficking during root development. Dev. Cell 2011, 21, 1144–1155. [Google Scholar] [CrossRef] [PubMed]
- Hofhuis, H.F.; Heidstra, R. Transcription factor dosage: More or less sufficient for growth. Curr. Opin. Plant Biol. 2018, 45, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Di Ruocco, G.; Di Mambro, R.; Dello Ioio, R. Building the differences: A case for the ground tissue patterning in plants. Proc. Biol. Sci. 2018, 285, 20181746. [Google Scholar] [CrossRef] [PubMed]
- Sena, G.; Jung, J.W.; Benfey, P.N. A broad competence to respond to short root revealed by tissue-specific ectopic expression. Development 2004, 131, 2817–2826. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Goedhart, J.; Schneijderberg, M.; Terpstra, I.; Shimotohno, A.; Bouchet, B.P.; Akhmanova, A.; Gadella, T.W., Jr.; Heidstra, R.; Scheres, B.; et al. Scarecrow-like23 and scarecrow jointly specify endodermal cell fate but distinctly control short-root movement. Plant J. 2015, 84, 773–784. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Smet, W.; Cruz-Ramirez, A.; Castelijns, B.; de Jonge, W.; Mahonen, A.P.; Bouchet, B.P.; Perez, G.S.; Akhmanova, A.; Scheres, B.; et al. Arabidopsis bird zinc finger proteins jointly stabilize tissue boundaries by confining the cell fate regulator short-root and contributing to fate specification. Plant Cell 2015, 27, 1185–1199. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Levesque, M.P.; Vernoux, T.; Jung, J.W.; Paquette, A.J.; Gallagher, K.L.; Wang, J.Y.; Blilou, I.; Scheres, B.; Benfey, P.N. An evolutionarily conserved mechanism delimiting shr movement defines a single layer of endodermis in plants. Science 2007, 316, 421–425. [Google Scholar] [CrossRef]
- Welch, D.; Hassan, H.; Blilou, I.; Immink, R.; Heidstra, R.; Scheres, B. Arabidopsis jackdaw and magpie zinc finger proteins delimit asymmetric cell division and stabilize tissue boundaries by restricting short-root action. Genes Dev. 2007, 21, 2196–2204. [Google Scholar] [CrossRef]
- Long, Y.; Stahl, Y.; Weidtkamp-Peters, S.; Postma, M.; Zhou, W.; Goedhart, J.; Sanchez-Perez, M.I.; Gadella, T.W.J.; Simon, R.; Scheres, B.; et al. In vivo fret-flim reveals cell-type-specific protein interactions in Arabidopsis roots. Nature 2017, 548, 97–102. [Google Scholar] [CrossRef]
- Long, Y.; Stahl, Y.; Weidtkamp-Peters, S.; Smet, W.; Du, Y.; Gadella, T.W.J., Jr.; Goedhart, J.; Scheres, B.; Blilou, I. Optimizing fret-flim labeling conditions to detect nuclear protein interactions at native expression levels in living Arabidopsis roots. Front. Plant Sci. 2018, 9, 639. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Risueno, M.A.; Sozzani, R.; Yardimci, G.G.; Petricka, J.J.; Vernoux, T.; Blilou, I.; Alonso, J.; Winter, C.M.; Ohler, U.; Scheres, B.; et al. Transcriptional control of tissue formation throughout root development. Science 2015, 350, 426–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sozzani, R.; Cui, H.; Moreno-Risueno, M.A.; Busch, W.; Van Norman, J.M.; Vernoux, T.; Brady, S.M.; Dewitte, W.; Murray, J.A.; Benfey, P.N. Spatiotemporal regulation of cell-cycle genes by shortroot links patterning and growth. Nature 2010, 466, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Perilli, S.; Perez-Perez, J.M.; Di Mambro, R.; Peris, C.L.; Diaz-Trivino, S.; Del Bianco, M.; Pierdonati, E.; Moubayidin, L.; Cruz-Ramirez, A.; Costantino, P.; et al. Retinoblastoma-related protein stimulates cell differentiation in the Arabidopsis root meristem by interacting with cytokinin signaling. Plant Cell 2013, 25, 4469–4478. [Google Scholar] [CrossRef] [PubMed]
- Wildwater, M.; Campilho, A.; Perez-Perez, J.M.; Heidstra, R.; Blilou, I.; Korthout, H.; Chatterjee, J.; Mariconti, L.; Gruissem, W.; Scheres, B. The retinoblastoma-related gene regulates stem cell maintenance in Arabidopsis roots. Cell 2005, 123, 1337–1349. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Ramirez, A.; Diaz-Trivino, S.; Blilou, I.; Grieneisen, V.A.; Sozzani, R.; Zamioudis, C.; Miskolczi, P.; Nieuwland, J.; Benjamins, R.; Dhonukshe, P.; et al. A bistable circuit involving scarecrow-retinoblastoma integrates cues to inform asymmetric stem cell division. Cell 2012, 150, 1002–1015. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhou, W.; Chen, Q.; Fang, M.; Zheng, S.; Scheres, B.; Li, C. Mediator subunit med31 is required for radial patterning of Arabidopsis roots. Proc. Natl. Acad. Sci. USA 2018, 115, E5624–E5633. [Google Scholar] [CrossRef]
- Wu, S.; Lee, C.M.; Hayashi, T.; Price, S.; Divol, F.; Henry, S.; Pauluzzi, G.; Perin, C.; Gallagher, K.L. A plausible mechanism, based upon short-root movement, for regulating the number of cortex cell layers in roots. Proc. Natl. Acad. Sci. USA 2014, 111, 16184–16189. [Google Scholar] [CrossRef]
- Koizumi, K.; Hayashi, T.; Wu, S.; Gallagher, K.L. The short-root protein acts as a mobile, dose-dependent signal in patterning the ground tissue. Proc. Natl. Acad. Sci. USA 2012, 109, 13010–13015. [Google Scholar] [CrossRef]
- Mylona, P.; Linstead, P.; Martienssen, R.; Dolan, L. Schizoriza controls an asymmetric cell division and restricts epidermal identity in the Arabidopsis root. Development 2002, 129, 4327–4334. [Google Scholar]
- Ten Hove, C.A.; Willemsen, V.; de Vries, W.J.; van Dijken, A.; Scheres, B.; Heidstra, R. Schizoriza encodes a nuclear factor regulating asymmetry of stem cell divisions in the Arabidopsis root. Curr. Biol. 2010, 20, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Pernas, M.; Ryan, E.; Dolan, L. Schizoriza controls tissue system complexity in plants. Curr. Biol. 2010, 20, 818–823. [Google Scholar] [CrossRef] [PubMed]
- Carlsbecker, A.; Lee, J.Y.; Roberts, C.J.; Dettmer, J.; Lehesranta, S.; Zhou, J.; Lindgren, O.; Moreno-Risueno, M.A.; Vaten, A.; Thitamadee, S.; et al. Cell signalling by microrna165/6 directs gene dose-dependent root cell fate. Nature 2010, 465, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Muraro, D.; Mellor, N.; Pound, M.P.; Help, H.; Lucas, M.; Chopard, J.; Byrne, H.M.; Godin, C.; Hodgman, T.C.; King, J.R.; et al. Integration of hormonal signaling networks and mobile micrornas is required for vascular patterning in Arabidopsis roots. Proc. Natl. Acad. Sci. USA 2014, 111, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Bishopp, A.; Help, H.; El-Showk, S.; Weijers, D.; Scheres, B.; Friml, J.; Benkova, E.; Mahonen, A.P.; Helariutta, Y. A mutually inhibitory interaction between auxin and cytokinin specifies vascular pattern in roots. Curr. Biol. 2011, 21, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Miyashima, S.; Koi, S.; Hashimoto, T.; Nakajima, K. Non-cell-autonomous microrna165 acts in a dose-dependent manner to regulate multiple differentiation status in the Arabidopsis root. Development 2011, 138, 2303–2313. [Google Scholar] [CrossRef] [PubMed]
- Di Ruocco, G.; Bertolotti, G.; Pacifici, E.; Polverari, L.; Tsiantis, M.; Sabatini, S.; Costantino, P.; Dello Ioio, R. Differential spatial distribution of mir165/6 determines variability in plant root anatomy. Development 2018, 145. [Google Scholar] [CrossRef]
- Miyashima, S.; Hashimoto, T.; Nakajima, K. Argonaute1 acts in Arabidopsis root radial pattern formation independently of the shr/scr pathway. Plant Cell Physiol. 2009, 50, 626–634. [Google Scholar] [CrossRef]
- Muller, C.J.; Valdes, A.E.; Wang, G.; Ramachandran, P.; Beste, L.; Uddenberg, D.; Carlsbecker, A. Phabulosa mediates an auxin signaling loop to regulate vascular patterning in Arabidopsis. Plant Physiol. 2016, 170, 956–970. [Google Scholar] [CrossRef]
- Li, P.; Yu, Q.; Gu, X.; Xu, C.; Qi, S.; Wang, H.; Zhong, F.; Baskin, T.I.; Rahman, A.; Wu, S. Construction of a functional casparian strip in non-endodermal lineages is orchestrated by two parallel signaling systems in Arabidopsis thaliana. Curr. Biol. 2018, 28, 2777.e2.–2786.e2. [Google Scholar] [CrossRef]
- Drapek, C.; Sparks, E.E.; Marhavy, P.; Taylor, I.; Andersen, T.G.; Hennacy, J.H.; Geldner, N.; Benfey, P.N. Minimum requirements for changing and maintaining endodermis cell identity in the Arabidopsis root. Nat. Plants 2018, 4, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, T.; Borghi, M.; Wang, P.; Danku, J.M.; Kalmbach, L.; Hosmani, P.S.; Naseer, S.; Fujiwara, T.; Geldner, N.; Salt, D.E. The myb36 transcription factor orchestrates casparian strip formation. Proc. Natl. Acad. Sci. USA 2015, 112, 10533–10538. [Google Scholar] [CrossRef] [PubMed]
- Roppolo, D.; De Rybel, B.; Denervaud Tendon, V.; Pfister, A.; Alassimone, J.; Vermeer, J.E.; Yamazaki, M.; Stierhof, Y.D.; Beeckman, T.; Geldner, N. A novel protein family mediates casparian strip formation in the endodermis. Nature 2011, 473, 380–383. [Google Scholar] [CrossRef]
- Dello Ioio, R.; Linhares, F.S.; Scacchi, E.; Casamitjana-Martinez, E.; Heidstra, R.; Costantino, P.; Sabatini, S. Cytokinins determine Arabidopsis root-meristem size by controlling cell differentiation. Curr. Biol. 2007, 17, 678–682. [Google Scholar] [CrossRef] [PubMed]
- Sabatini, S.; Beis, D.; Wolkenfelt, H.; Murfett, J.; Guilfoyle, T.; Malamy, J.; Benfey, P.; Leyser, O.; Bechtold, N.; Weisbeek, P.; et al. An auxin-dependent distal organizer of pattern and polarity in the Arabidopsis root. Cell 1999, 99, 463–472. [Google Scholar] [CrossRef]
- Leyser, O. Auxin distribution and plant pattern formation: How many angels can dance on the point of pin? Cell 2005, 121, 819–822. [Google Scholar] [CrossRef]
- Yoshida, S.; Saiga, S.; Weijers, D. Auxin regulation of embryonic root formation. Plant Cell Physiol. 2013, 54, 325–332. [Google Scholar] [CrossRef]
- Blilou, I.; Xu, J.; Wildwater, M.; Willemsen, V.; Paponov, I.; Friml, J.; Heidstra, R.; Aida, M.; Palme, K.; Scheres, B. The pin auxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nature 2005, 433, 39–44. [Google Scholar] [CrossRef]
- Wisniewska, J.; Xu, J.; Seifertova, D.; Brewer, P.B.; Ruzicka, K.; Blilou, I.; Rouquie, D.; Benkova, E.; Scheres, B.; Friml, J. Polar pin localization directs auxin flow in plants. Science 2006, 312, 883. [Google Scholar] [CrossRef]
- Friml, J.; Vieten, A.; Sauer, M.; Weijers, D.; Schwarz, H.; Hamann, T.; Offringa, R.; Jurgens, G. Efflux-dependent auxin gradients establish the apical-basal axis of Arabidopsis. Nature 2003, 426, 147–153. [Google Scholar] [CrossRef]
- Weijers, D.; Schlereth, A.; Ehrismann, J.S.; Schwank, G.; Kientz, M.; Jurgens, G. Auxin triggers transient local signaling for cell specification in Arabidopsis embryogenesis. Dev. Cell 2006, 10, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Hamann, T.; Mayer, U.; Jurgens, G. The auxin-insensitive bodenlos mutation affects primary root formation and apical-basal patterning in the Arabidopsis embryo. Development 1999, 126, 1387–1395. [Google Scholar] [PubMed]
- Hamann, T.; Benkova, E.; Baurle, I.; Kientz, M.; Jurgens, G. The Arabidopsis bodenlos gene encodes an auxin response protein inhibiting monopteros-mediated embryo patterning. Genes Dev. 2002, 16, 1610–1615. [Google Scholar] [CrossRef] [PubMed]
- Aida, M.; Beis, D.; Heidstra, R.; Willemsen, V.; Blilou, I.; Galinha, C.; Nussaume, L.; Noh, Y.S.; Amasino, R.; Scheres, B. The plethora genes mediate patterning of the Arabidopsis root stem cell niche. Cell 2004, 119, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Galinha, C.; Hofhuis, H.; Luijten, M.; Willemsen, V.; Blilou, I.; Heidstra, R.; Scheres, B. Plethora proteins as dose-dependent master regulators of Arabidopsis root development. Nature 2007, 449, 1053–1057. [Google Scholar] [CrossRef] [PubMed]
- Nawy, T.; Bayer, M.; Mravec, J.; Friml, J.; Birnbaum, K.D.; Lukowitz, W. The gata factor hanaba taranu is required to position the proembryo boundary in the early Arabidopsis embryo. Dev. Cell 2010, 19, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Grigg, S.P.; Galinha, C.; Kornet, N.; Canales, C.; Scheres, B.; Tsiantis, M. Repression of apical homeobox genes is required for embryonic root development in Arabidopsis. Curr. Biol. 2009, 19, 1485–1490. [Google Scholar] [CrossRef]
- Smith, Z.R.; Long, J.A. Control of Arabidopsis apical-basal embryo polarity by antagonistic transcription factors. Nature 2010, 464, 423–426. [Google Scholar] [CrossRef]
- Santuari, L.; Sanchez-Perez, G.F.; Luijten, M.; Rutjens, B.; Terpstra, I.; Berke, L.; Gorte, M.; Prasad, K.; Bao, D.; Timmermans-Hereijgers, J.L.; et al. The plethora gene regulatory network guides growth and cell differentiation in Arabidopsis roots. Plant Cell 2016, 28, 2937–2951. [Google Scholar] [CrossRef]
- Mahonen, A.P.; Ten Tusscher, K.; Siligato, R.; Smetana, O.; Diaz-Trivino, S.; Salojarvi, J.; Wachsman, G.; Prasad, K.; Heidstra, R.; Scheres, B. Plethora gradient formation mechanism separates auxin responses. Nature 2014, 515, 125–129. [Google Scholar] [CrossRef]
- Di Mambro, R.; De Ruvo, M.; Pacifici, E.; Salvi, E.; Sozzani, R.; Benfey, P.N.; Busch, W.; Novak, O.; Ljung, K.; Di Paola, L.; et al. Auxin minimum triggers the developmental switch from cell division to cell differentiation in the Arabidopsis root. Proc. Natl. Acad. Sci. USA 2017, 114, E7641–E7649. [Google Scholar] [CrossRef] [PubMed]
- Dello Ioio, R.; Nakamura, K.; Moubayidin, L.; Perilli, S.; Taniguchi, M.; Morita, M.T.; Aoyama, T.; Costantino, P.; Sabatini, S. A genetic framework for the control of cell division and differentiation in the root meristem. Science 2008, 322, 1380–1384. [Google Scholar] [CrossRef] [PubMed]
- Moubayidin, L.; Perilli, S.; Dello Ioio, R.; Di Mambro, R.; Costantino, P.; Sabatini, S. The rate of cell differentiation controls the Arabidopsis root meristem growth phase. Curr. Biol. 2010, 20, 1138–1143. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.B.; Wang, X.J.; Hagen, G.; Guilfoyle, T.J. Aux/IAA proteins are active repressors, and their stability and activity are modulated by auxin. Plant Cell 2001, 13, 2809–2822. [Google Scholar] [CrossRef] [PubMed]
- Moubayidin, L.; Di Mambro, R.; Sozzani, R.; Pacifici, E.; Salvi, E.; Terpstra, I.; Bao, D.; van Dijken, A.; Dello Ioio, R.; Perilli, S.; et al. Spatial coordination between stem cell activity and cell differentiation in the root meristem. Dev. Cell 2013, 26, 405–415. [Google Scholar] [CrossRef]
- Salvi, E.; Di Mambro, R.; Pacifici, E.; Dello Ioio, R.; Costantino, P.; Moubayidin, L.; Sabatini, S. Scarecrow and shortroot control the auxin/cytokinin balance necessary for embryonic stem cell niche specification. Plant Signal Behav. 2018, 13, e1507402. [Google Scholar]
- Dello Ioio, R.; Galinha, C.; Fletcher, A.G.; Grigg, S.P.; Molnar, A.; Willemsen, V.; Scheres, B.; Sabatini, S.; Baulcombe, D.; Maini, P.K.; et al. A phabulosa/cytokinin feedback loop controls root growth in Arabidopsis. Curr. Biol. 2012, 22, 1699–1704. [Google Scholar] [CrossRef]
- Pacifici, E.; Di Mambro, R.; Dello Ioio, R.; Costantino, P.; Sabatini, S. Acidic cell elongation drives cell differentiation in the Arabidopsis root. EMBO J. 2018, 37, e99134. [Google Scholar] [CrossRef]
- Vuolo, F.; Kierzkowski, D.; Runions, A.; Hajheidari, M.; Mentink, R.A.; Gupta, M.D.; Zhang, Z.; Vlad, D.; Wang, Y.; Pecinka, A.; et al. Lmi1 homeodomain protein regulates organ proportions by spatial modulation of endoreduplication. Genes Dev. 2018, 32, 1361–1366. [Google Scholar] [CrossRef]
- Hayashi, K.; Hasegawa, J.; Matsunaga, S. The boundary of the meristematic and elongation zones in roots: Endoreduplication precedes rapid cell expansion. Sci. Rep. 2013, 3, 2723. [Google Scholar] [CrossRef]
- Takahashi, N.; Kajihara, T.; Okamura, C.; Kim, Y.; Katagiri, Y.; Okushima, Y.; Matsunaga, S.; Hwang, I.; Umeda, M. Cytokinins control endocycle onset by promoting the expression of an apc/c activator in Arabidopsis roots. Curr. Biol. 2013, 23, 1812–1817. [Google Scholar] [CrossRef] [PubMed]
- Esau’s Plant Anatomy: Meristems, Cells, and Tissues of the Plant Body: Their Structure, Function, and Development, 3rd ed.; Wiley Interscience, John Wiley & Sons: Hoboken, NJ, USA, 2006; p. 601. ISBN 978-0-471-73843-5.
- Ron, M.; Dorrity, M.W.; de Lucas, M.; Toal, T.; Hernandez, R.I.; Little, S.A.; Maloof, J.N.; Kliebenstein, D.J.; Brady, S.M. Identification of novel loci regulating interspecific variation in root morphology and cellular development in tomato. Plant Physiol. 2013, 162, 755–768. [Google Scholar] [CrossRef] [PubMed]
- Henry, S.; Dievart, A.; Divol, F.; Pauluzzi, G.; Meynard, D.; Swarup, R.; Wu, S.; Gallagher, K.L.; Perin, C. Shr overexpression induces the formation of supernumerary cell layers with cortex cell identity in rice. Dev. Biol. 2017, 425, 1–7. [Google Scholar] [CrossRef] [PubMed]
- De Vries, J.; Fischer, A.M.; Roettger, M.; Rommel, S.; Schluepmann, H.; Brautigam, A.; Carlsbecker, A.; Gould, S.B. Cytokinin-induced promotion of root meristem size in the fern azolla supports a shoot-like origin of euphyllophyte roots. New Phytol. 2016, 209, 705–720. [Google Scholar] [CrossRef] [PubMed]
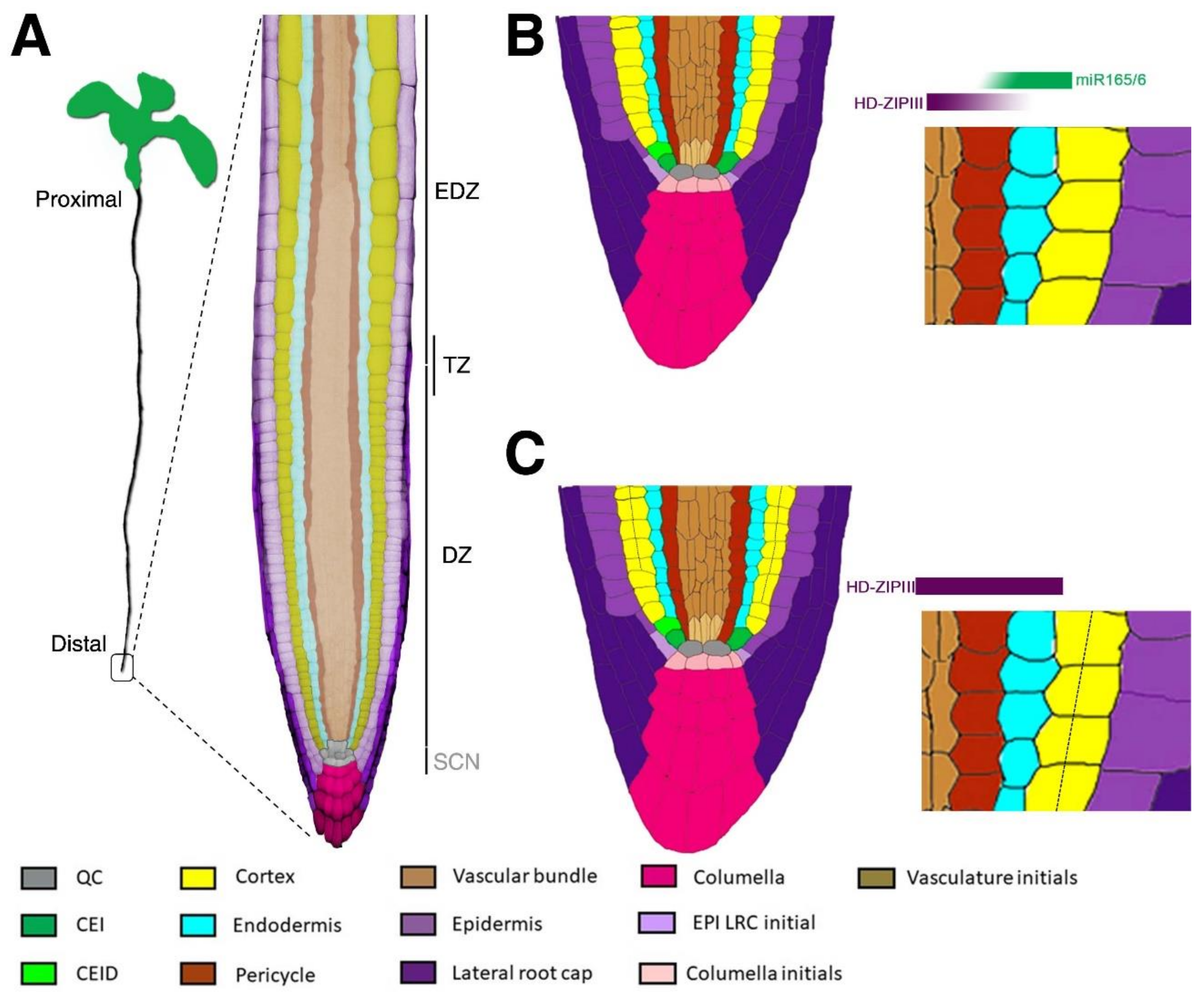
Figure 1.
Arabidopsis root structure. (A) Representation of an Arabidopsis seedling where the proximodistal axis is indicated. In the blow up, a representation of the Arabidopsis root apex is shown where false colours highlight the different tissues. Root zonation: stem cell niche, SCN; division zone, DZ; elongation/differentiation zone, EDZ; transition zone, TZ. (B) Cartoon reporting the longitudinal section of a wild type (Wt) Arabidopsis root stem cell niche. Different colours represent root tissues and initials, as indicated in the legend. The blow up highlights the typical ground tissue (GT) architecture (one layer of endodermis and one layer of cortex) resulting from the opposite graded distribution of miR165/6 and Class III Homeodomain Leucine Zipper (HD-ZIPIII) (triangle shapes above blow up). In particular, miR165/6 (green) presents low expression in the vascular bundle and high expression in the endodermis, constraining HD-ZIPIII expression. As a result, HD-ZIPIII (red) present high expression in the vascular bundle and low expression in the endodermis. (C) Cartoon reporting the longitudinal section of an Arabidopsis stem cell niche lacking miR165/6 expression. The blow up highlights the HD-ZIPIII expanded expression in the whole ground tissue (GT). This results in the formation of an extra layer of the cortical tissue (dashed line). QC, quiescent centre; CEI, cortex and endodermis initial; CEID, cortex and endodermis initial daughter cell; EPI, epidermis; LRC, lateral root cap.
Figure 1.
Arabidopsis root structure. (A) Representation of an Arabidopsis seedling where the proximodistal axis is indicated. In the blow up, a representation of the Arabidopsis root apex is shown where false colours highlight the different tissues. Root zonation: stem cell niche, SCN; division zone, DZ; elongation/differentiation zone, EDZ; transition zone, TZ. (B) Cartoon reporting the longitudinal section of a wild type (Wt) Arabidopsis root stem cell niche. Different colours represent root tissues and initials, as indicated in the legend. The blow up highlights the typical ground tissue (GT) architecture (one layer of endodermis and one layer of cortex) resulting from the opposite graded distribution of miR165/6 and Class III Homeodomain Leucine Zipper (HD-ZIPIII) (triangle shapes above blow up). In particular, miR165/6 (green) presents low expression in the vascular bundle and high expression in the endodermis, constraining HD-ZIPIII expression. As a result, HD-ZIPIII (red) present high expression in the vascular bundle and low expression in the endodermis. (C) Cartoon reporting the longitudinal section of an Arabidopsis stem cell niche lacking miR165/6 expression. The blow up highlights the HD-ZIPIII expanded expression in the whole ground tissue (GT). This results in the formation of an extra layer of the cortical tissue (dashed line). QC, quiescent centre; CEI, cortex and endodermis initial; CEID, cortex and endodermis initial daughter cell; EPI, epidermis; LRC, lateral root cap.

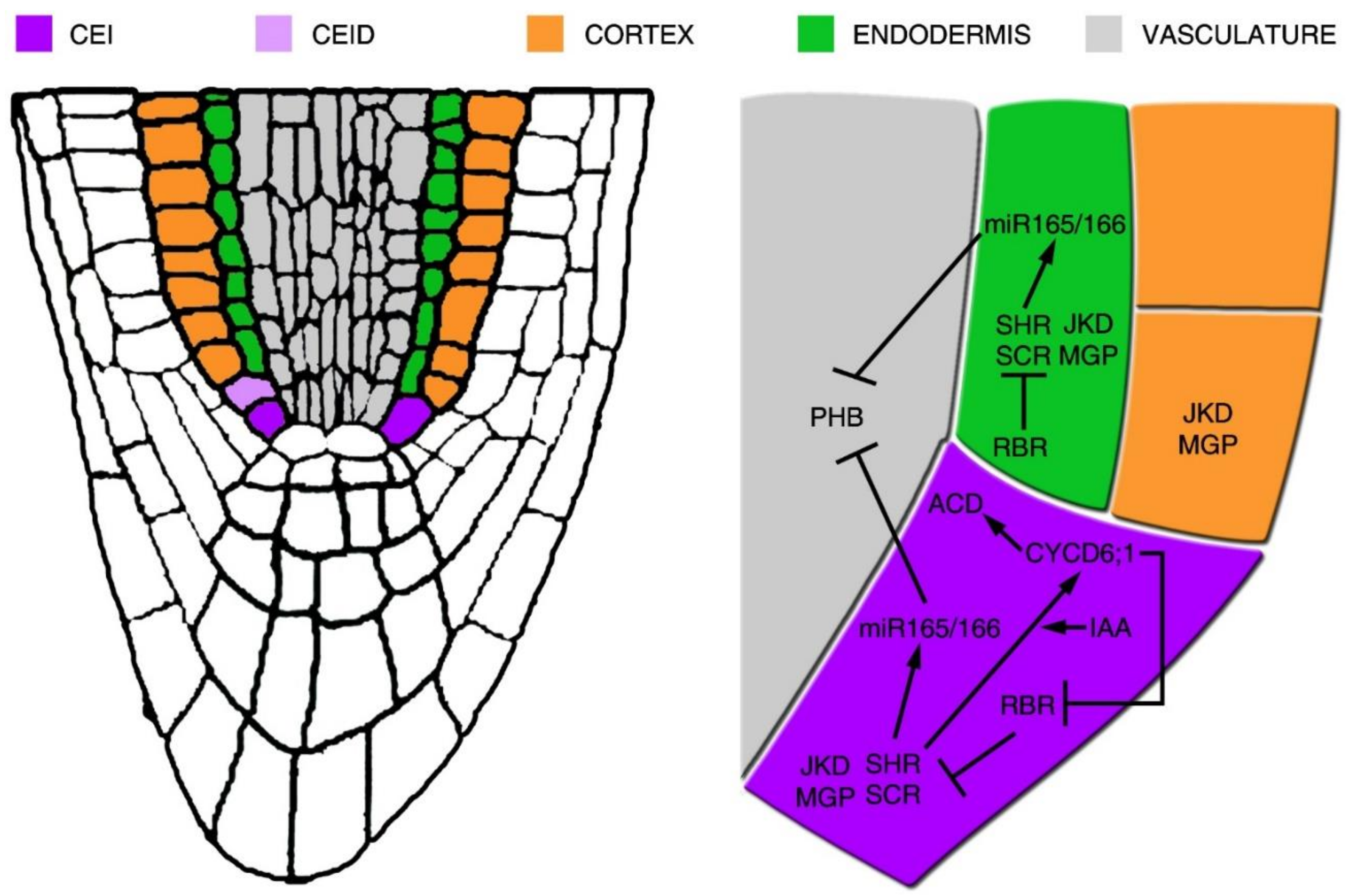
Figure 2.
Schematic representation of the gene regulatory network acting for the cortex and endodermis initial periclinal division. On the left, representation is provided of the Arabidopsis root tip, where the cortex and endodermis initial (CEI) and its daughter cell (CEID), cortex, endodermis and vascular tissues are depicted in colour. In the blow up, the gene regulatory network supporting the CEI asymmetric cell division (ACD) is shown. In the CEI, the SHR/SCR complex sustains the expression of SCR and promotes the expressions of CYCD1;6 and of JKD and MGP. CYCD6;1 expression is also sustained by high levels of auxin (IAA) in the CEI. CYCD6;1 represses RBR activity, which in turn regulates negatively the ACD by a direct repression of SCR activity. SHR/SCR complex also promotes the expression of miR165/6, thus restricting PHB expression in the vascular tissue.
Figure 2.
Schematic representation of the gene regulatory network acting for the cortex and endodermis initial periclinal division. On the left, representation is provided of the Arabidopsis root tip, where the cortex and endodermis initial (CEI) and its daughter cell (CEID), cortex, endodermis and vascular tissues are depicted in colour. In the blow up, the gene regulatory network supporting the CEI asymmetric cell division (ACD) is shown. In the CEI, the SHR/SCR complex sustains the expression of SCR and promotes the expressions of CYCD1;6 and of JKD and MGP. CYCD6;1 expression is also sustained by high levels of auxin (IAA) in the CEI. CYCD6;1 represses RBR activity, which in turn regulates negatively the ACD by a direct repression of SCR activity. SHR/SCR complex also promotes the expression of miR165/6, thus restricting PHB expression in the vascular tissue.

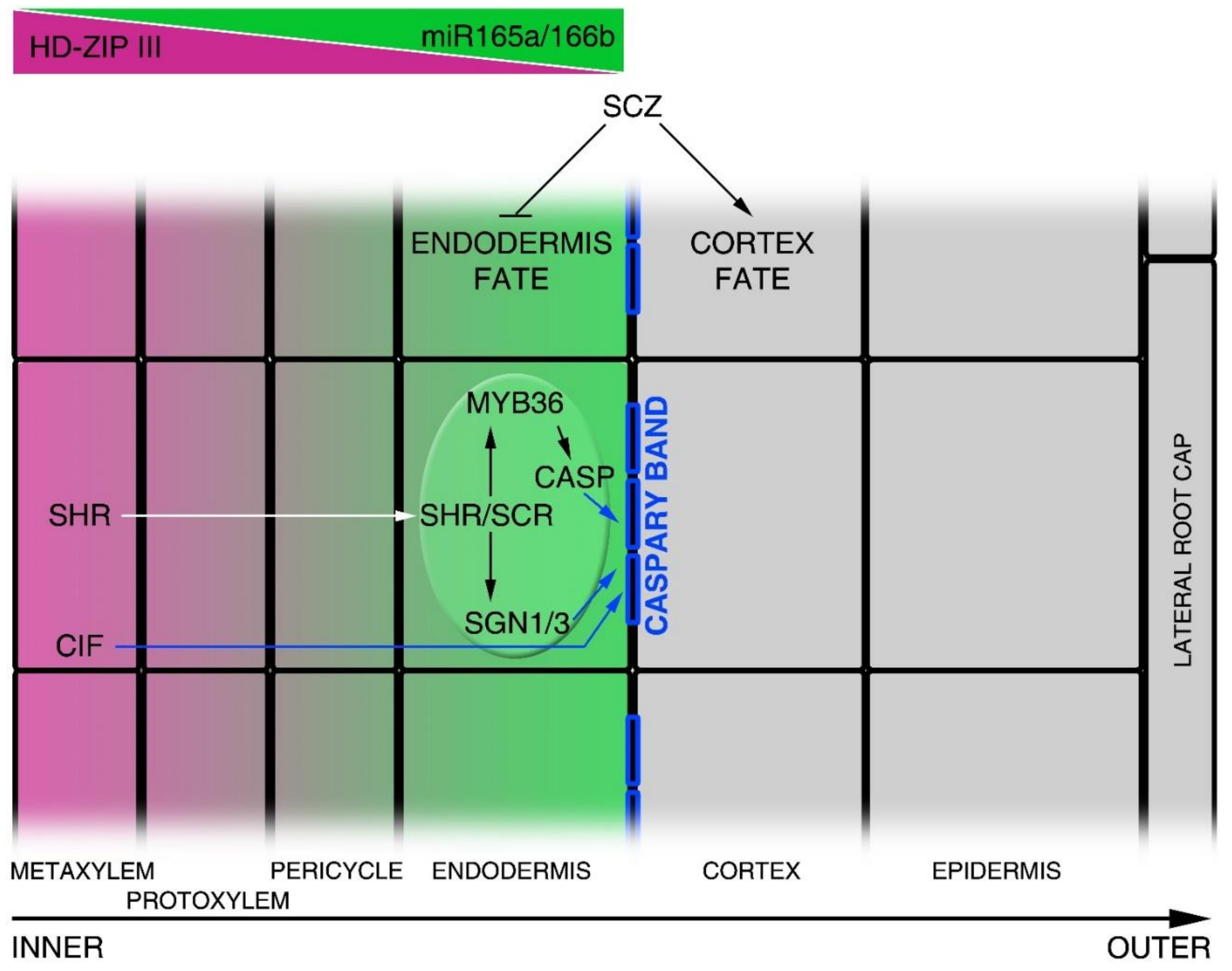
Figure 3.
Image showing the molecular mechanisms controlling radial axis patterning. The figure shows a half radial section of the Arabidopsis root. Each square file corresponds to a different tissue layer, where the central file (inner) corresponds to the metaxylem and the outer file to the lateral root cap, as indicated in the scheme. Class III Homeodomain Leucine Zipper III (HD-ZIPIII) and miR165a/6b (opposite gradients) are indicated in purple and green, respectively. Blue squares on endodermal cells represent Casparian strips. White arrow indicates SHR protein movement from the vascular tissue into endodermal cell nucleus. Blue arrows indicate the CIF-, CASP- and SGN1-dependent regulation of Caspary band formation. SCZ promotes the cortical identity supporting tissue fate separation.
Figure 3.
Image showing the molecular mechanisms controlling radial axis patterning. The figure shows a half radial section of the Arabidopsis root. Each square file corresponds to a different tissue layer, where the central file (inner) corresponds to the metaxylem and the outer file to the lateral root cap, as indicated in the scheme. Class III Homeodomain Leucine Zipper III (HD-ZIPIII) and miR165a/6b (opposite gradients) are indicated in purple and green, respectively. Blue squares on endodermal cells represent Casparian strips. White arrow indicates SHR protein movement from the vascular tissue into endodermal cell nucleus. Blue arrows indicate the CIF-, CASP- and SGN1-dependent regulation of Caspary band formation. SCZ promotes the cortical identity supporting tissue fate separation.

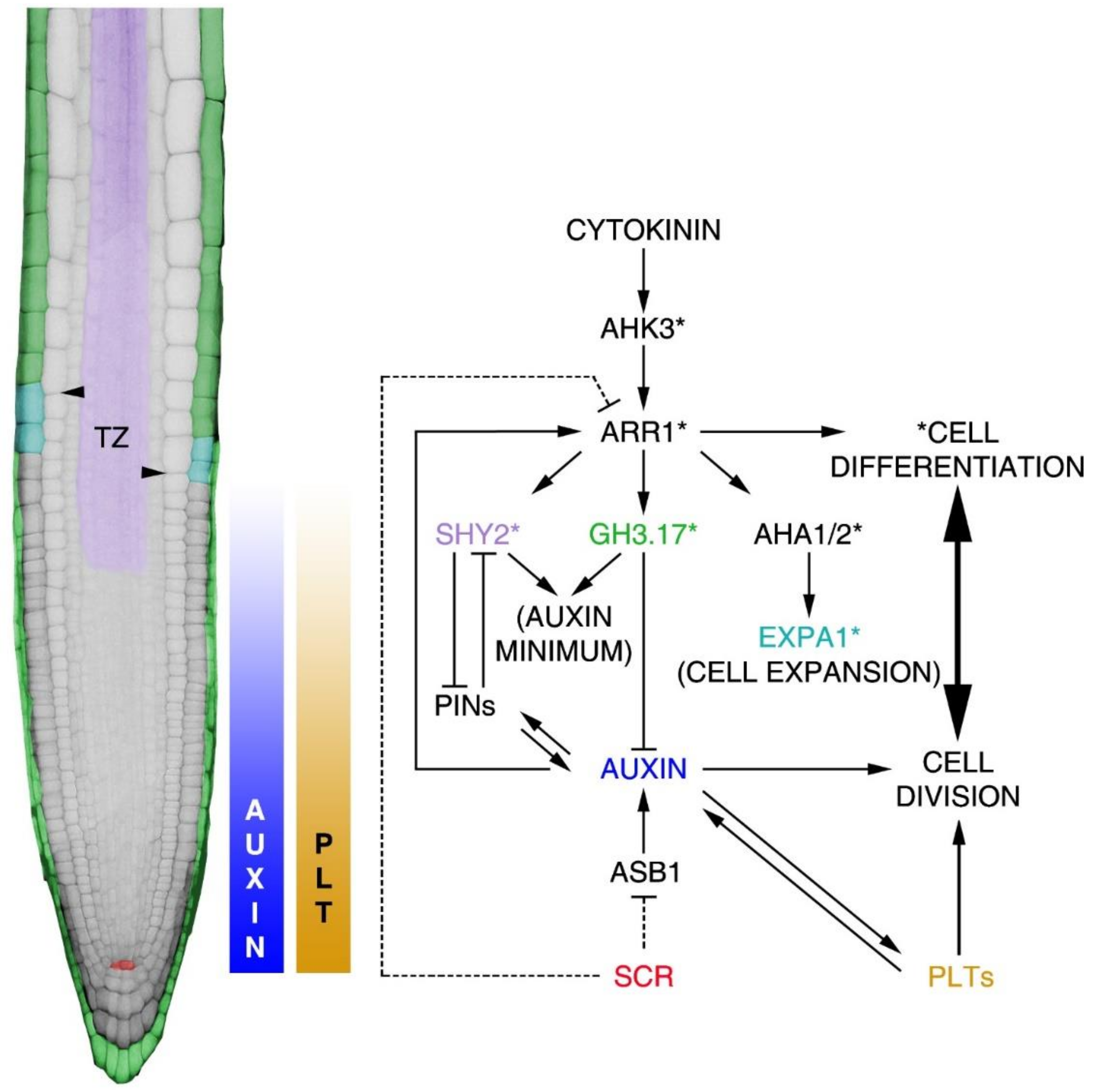
Figure 4.
Molecular mechanisms leading to the root proximodistal axis patterning. On the left, representation of the Arabidopsis root. False colours indicate the activity domain of genes involved in the regulation of the equilibrium between cell division and cell differentiation processes established in the root. Arrowheads indicate the position of the TZ. Auxin and PLT-graded distributions are indicated in the piecewise colour bar, where the maximum is in the SCN and the minimum at the TZ. The molecular mechanisms acting in the control of meristem activity are indicated in the diagram. The QC SCR domain is represented by red; in green, the root cap and differentiated epidermis GH3.17 domain; in purple, the vascular bundles SHY2 domain; and in light blue, the first elongating epidermal cells EXPA1 domain. Asterisks represent genes involved in the regulation of the cell differentiation process mediated by cytokinin activity. Dashed lines indicate SCR-dependent indirect regulation of ASB1 and ARR1.
Figure 4.
Molecular mechanisms leading to the root proximodistal axis patterning. On the left, representation of the Arabidopsis root. False colours indicate the activity domain of genes involved in the regulation of the equilibrium between cell division and cell differentiation processes established in the root. Arrowheads indicate the position of the TZ. Auxin and PLT-graded distributions are indicated in the piecewise colour bar, where the maximum is in the SCN and the minimum at the TZ. The molecular mechanisms acting in the control of meristem activity are indicated in the diagram. The QC SCR domain is represented by red; in green, the root cap and differentiated epidermis GH3.17 domain; in purple, the vascular bundles SHY2 domain; and in light blue, the first elongating epidermal cells EXPA1 domain. Asterisks represent genes involved in the regulation of the cell differentiation process mediated by cytokinin activity. Dashed lines indicate SCR-dependent indirect regulation of ASB1 and ARR1.

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Di Mambro, R.; Sabatini, S.; Dello Ioio, R. Patterning the Axes: A Lesson from the Root. Plants 2019, 8, 8. https://doi.org/10.3390/plants8010008
AMA Style
Di Mambro R, Sabatini S, Dello Ioio R. Patterning the Axes: A Lesson from the Root. Plants. 2019; 8(1):8. https://doi.org/10.3390/plants8010008
Chicago/Turabian StyleDi Mambro, Riccardo, Sabrina Sabatini, and Raffaele Dello Ioio. 2019. "Patterning the Axes: A Lesson from the Root" Plants 8, no. 1: 8. https://doi.org/10.3390/plants8010008
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.