
The Effects of Hydro-Priming and Colonization with Piriformospora indica and Azotobacter chroococcum on Physio-Biochemical Traits, Flavonolignans and Fatty Acids Composition of Milk Thistle (Silybum marianum) under Saline Conditions


 ,
, 
Abstract
:1. Introduction
2. Results
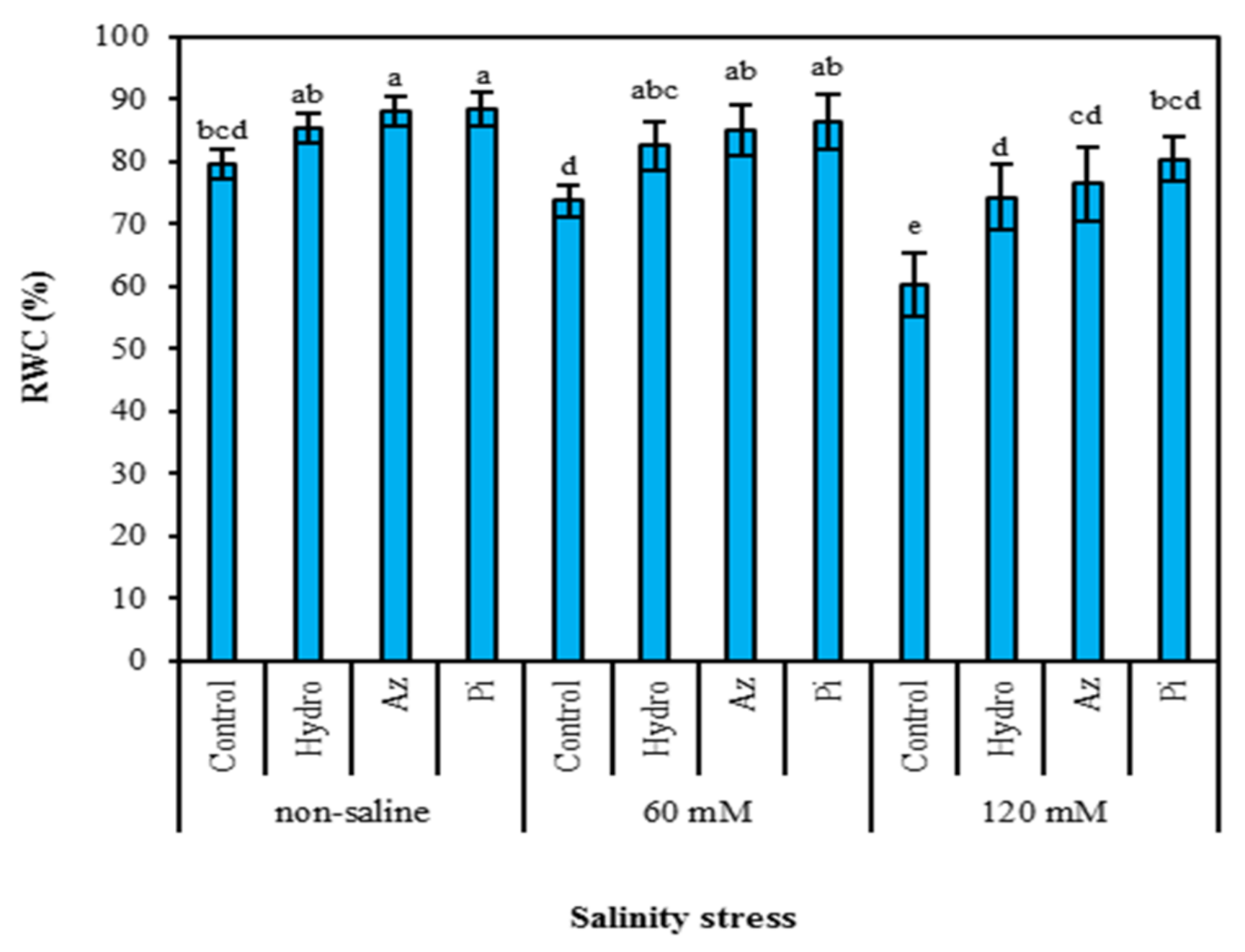
2.1. Relative Water Content (RWC)
2.2. Membrane Stability Index (MSI)
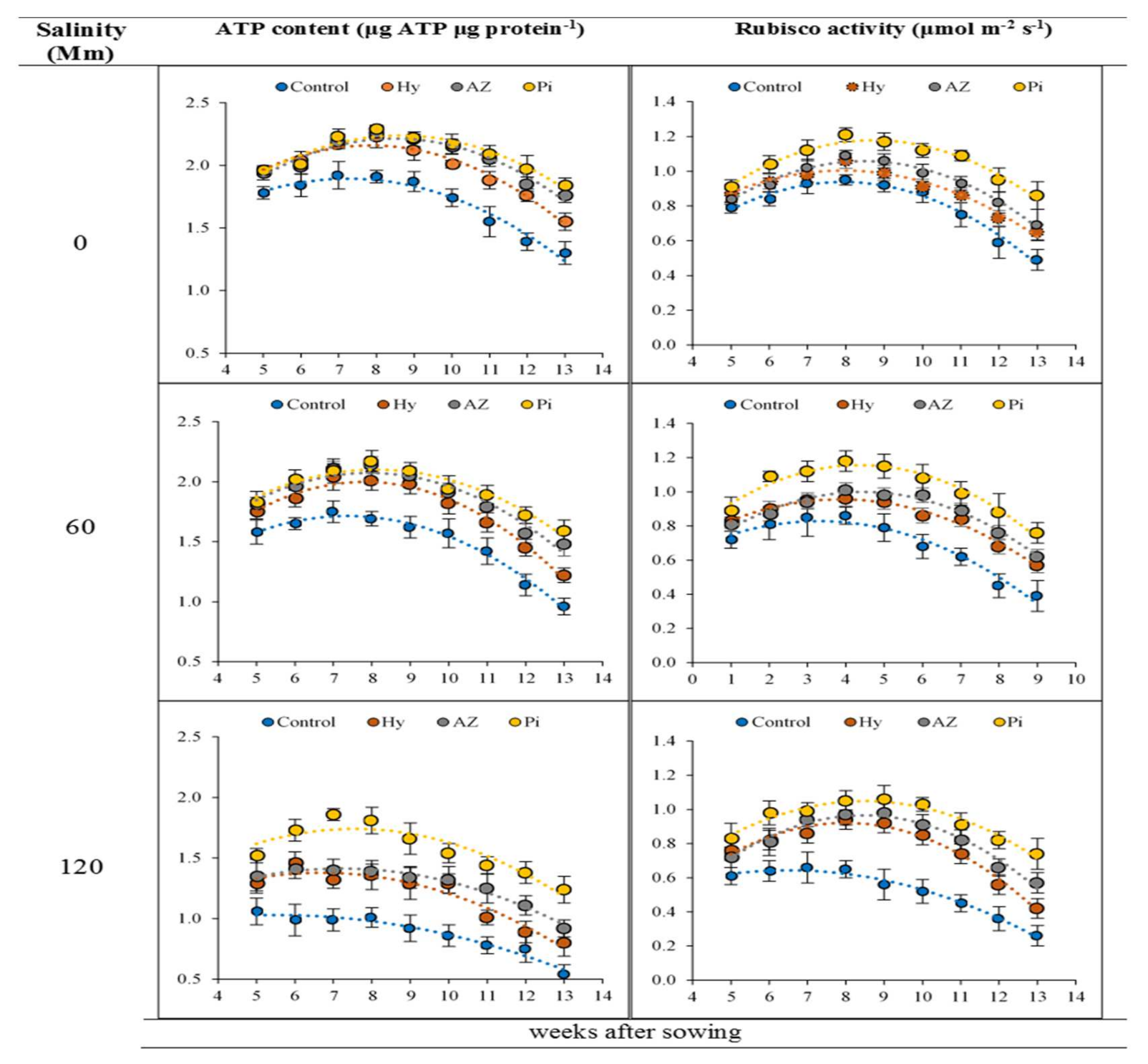
2.3. Adenosine Triphosphate (ATP) Content
2.4. Rubisco Activity
2.5. Malondialdehyde (MDA) Concentration
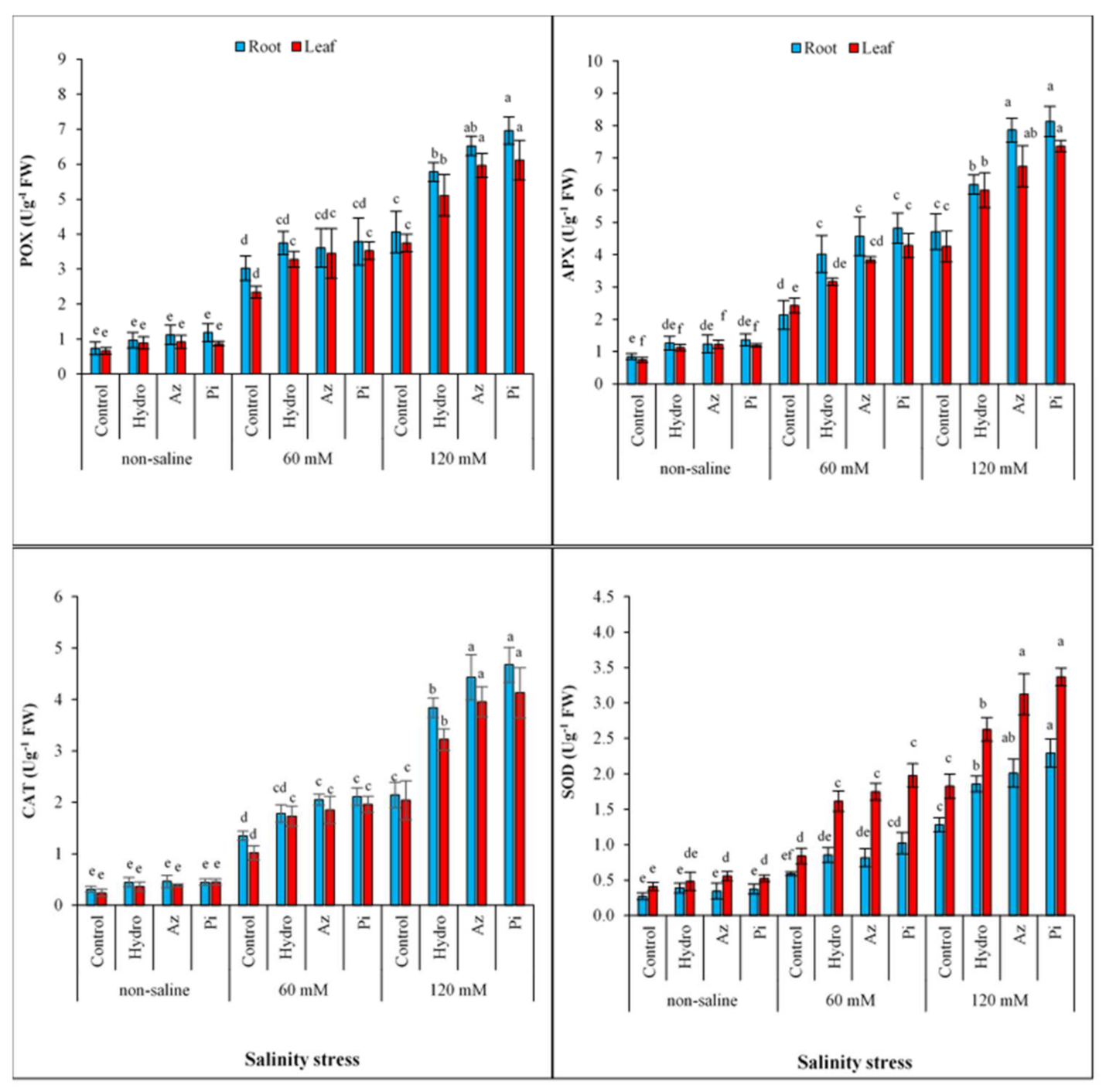
2.6. Antioxidant Enzyme Activities
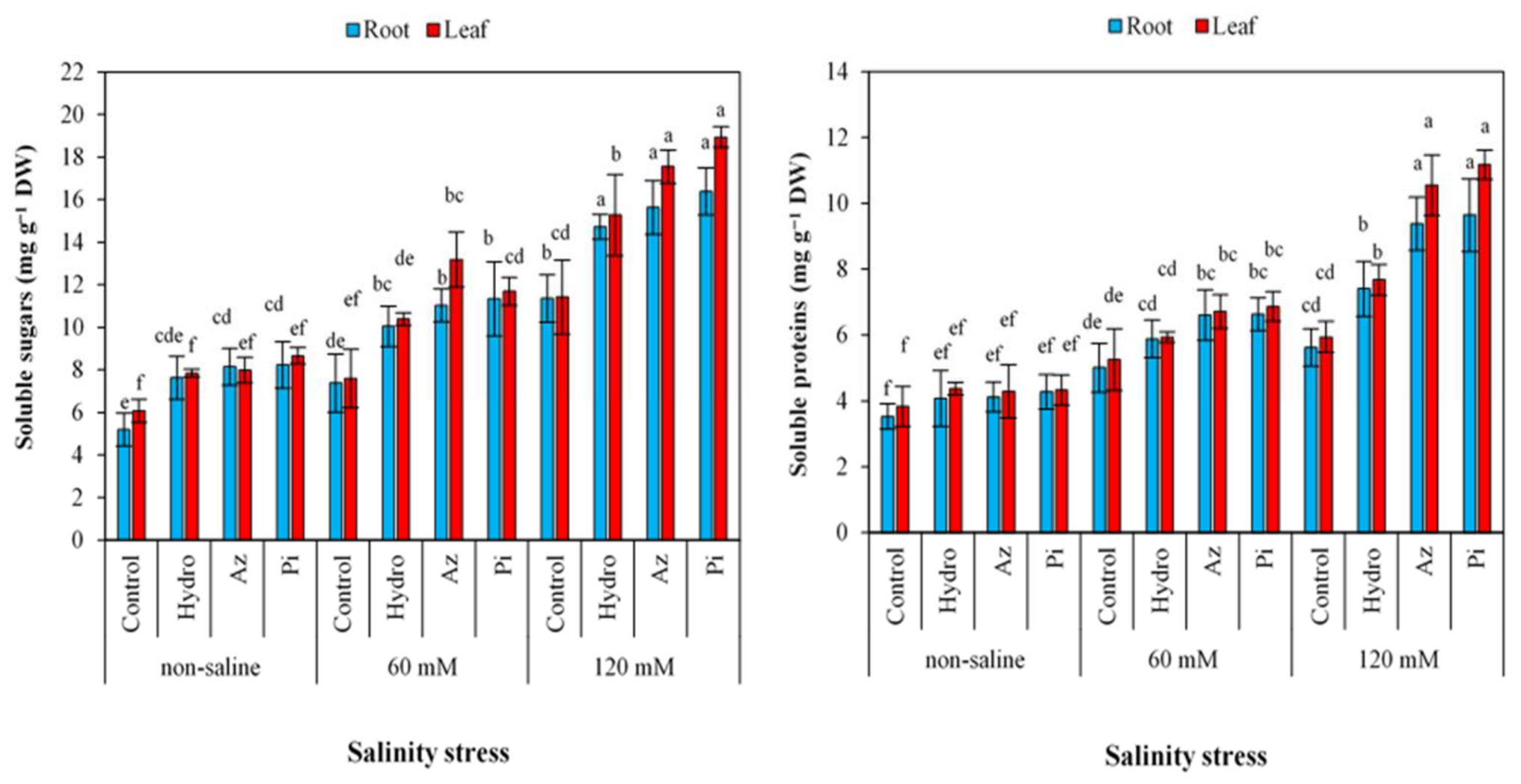
2.7. Soluble Sugar and Proteins
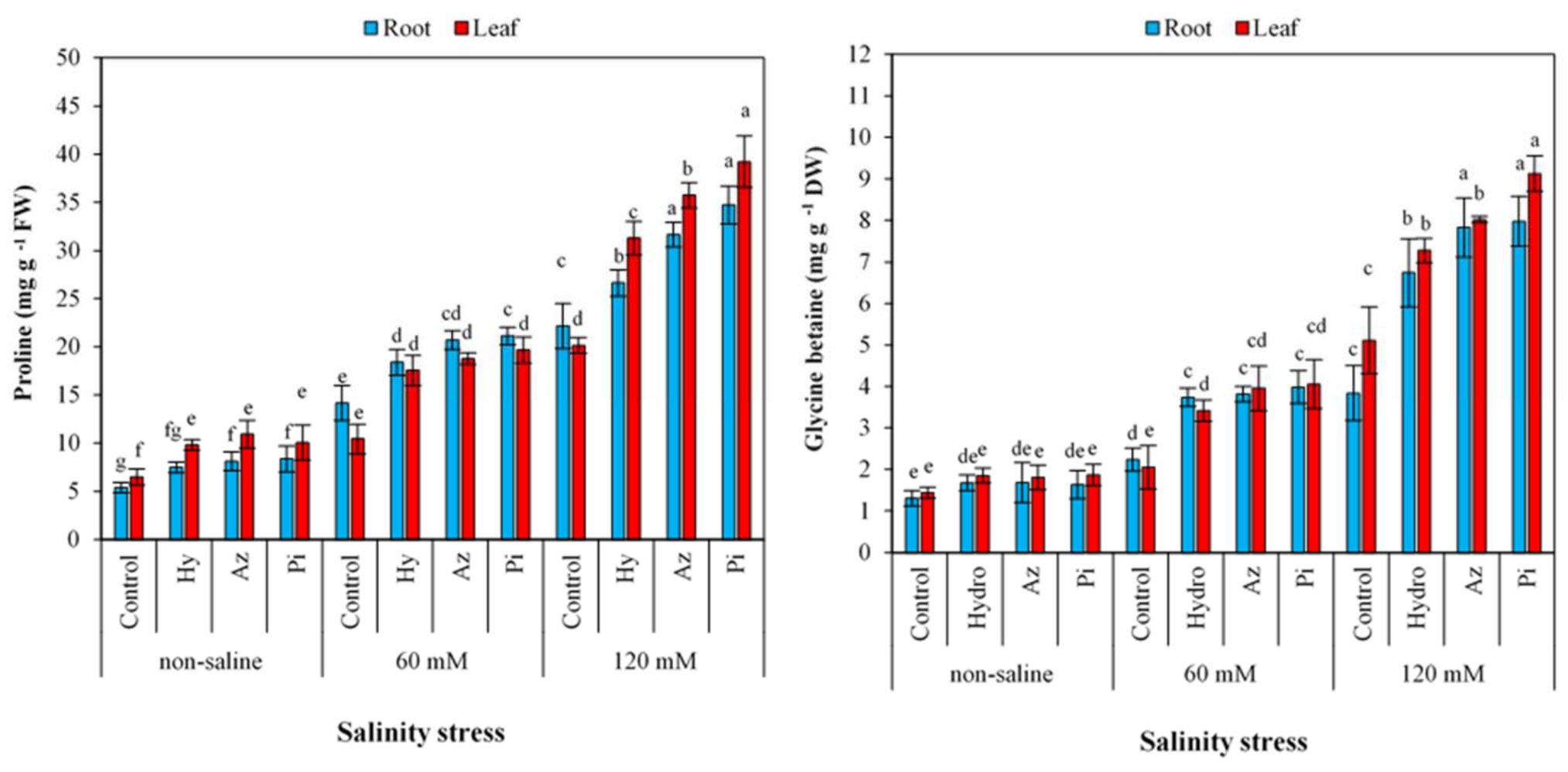
2.8. Proline and Glycine Betaine Contents
2.9. Flavonolignans in Milk Thistle Seed
2.10. Oil Content and Fatty Acids Composition
3. Discussion
4. Materials and Methods
4.1. Experimental Conditions
4.2. Relative Water Content (RWC) and Membrane Stability Index (MSI)
4.3. Adenosine Triphosphate (ATP) Content and Rubisco Activity
4.4. Malondialdehyde (MDA) Concentration
4.5. Determination of Antioxidant Enzymes Activities
4.6. Soluble Sugar and Proteins
4.7. Proline Content and Glycine Betaine Content
4.8. Determination of Flavonolignans
4.9. Oil Extraction and Fatty Acids Composition
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ramasamy, K.; Agarwal, R. Multitargeted therapy of cancer by silymarin. Cancer Lett. 2008, 269, 352–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, R.; Agarwal, C.; Ichikawa, H.; Singh, R.P.; Aggarwal, B.B. Anticancer potential of silymarin: From bench to bed side. Anticancer Res. 2006, 26, 4457–4498. [Google Scholar]
- Hadolin, M.; Škerget, M.; Knez, Z.E.; Bauman, D. High pressure extraction of vitamin E-rich oil from Silybum marianum. Food Chem. 2001, 74, 355–364. [Google Scholar] [CrossRef]
- Glevitzky, I.; Dumitrel, G.A.; Glevitzky, M.; Pasca, B.; Otrisal, P.; Bungau, S.; Cioca, G.; Pantis, C.; Popa, M. Statistical analysis of the relationship between antioxidant activity and the structure of flavonoid compounds. Rev. Chim. 2019, 70, 3103–3107. [Google Scholar] [CrossRef]
- Tedesco, D.; Tava, A.; Galletti, S.; Tameni, M.; Varisco, G.; Costa, A.; Steidler, S. Effects of silymarin, a natural hepatoprotector, in periparturient dairy cows. J. Dairy Sci. 2004, 87, 2239–2247. [Google Scholar] [CrossRef] [Green Version]
- Saller, R.; Melzer, J.; Reichling, J.; Brignoli, R.; Meier, R. An updated systematic review of the pharmacology of silymarin. Complement. Med. Res. 2007, 14, 70–80. [Google Scholar] [CrossRef]
- Mamnabi, S.; Nasrollahzadeh, S.; Ghassemi-Golezani, K.; Raei, Y. Improving yield-related physiological characteristics of spring rapeseed by integrated fertilizer management under water deficit conditions. Saudi J. Biol. Sci. 2020, 27, 797–804. [Google Scholar] [CrossRef]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef]
- Hamdy, A.; Abdel-Dayem, S.; Abu-Zeid, M. Saline water management for optimum crop production. Agric. Water Manag. 1993, 24, 189–203. [Google Scholar] [CrossRef]
- Blokhina, O.; Virolainen, E.; Fagerstedt, K.V. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef] [Green Version]
- Jubany-Marí, T.; Munné-Bosch, S.; Alegre, L. Redox regulation of water stress responses in field-grown plants. Role of hydrogen peroxide and ascorbate. Plant Physiol. Biochem. 2010, 48, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Ozkur, O.; Ozdemir, F.; Bor, M.; Turkan, I. Physiochemical and antioxidant responses of the perennial xerophyte Capparis ovata Desf. to drought. Environ. Exp. Bot. 2009, 66, 487–492. [Google Scholar] [CrossRef]
- Bungau, S.; Behl, T.; Aleya, L.; Bourgeade, P.; Aloui-Sossé, B.; Purza, A.L.; Abid, A.; Samuel, A.D. Expatiating the impact of anthropogenic aspects and climatic factors on long-term soil monitoring and management. Environ. Sci. Pollut. Res. 2021, 28, 30528–30550. [Google Scholar] [CrossRef] [PubMed]
- Asghari, B.; Khademian, R.; Sedaghati, B. Plant growth promoting rhizobacteria (PGPR) confer drought resistance and stimulate biosynthesis of secondary metabolites in pennyroyal (Mentha pulegium L.) under water shortage condition. Sci. Hortic. 2020, 263, 109132. [Google Scholar] [CrossRef]
- Bharti, N.; Barnawal, D.; Awasthi, A.; Yadav, A.; Kalra, A. Plant growth promoting rhizobacteria alleviate salinity induced negative effects on growth, oil content and physiological status in Mentha arvensis. Acta Physiol. Plant. 2014, 36, 45–60. [Google Scholar] [CrossRef]
- Farooq, M.; Siddique, K.H.; Rehman, H.; Aziz, T.; Lee, D.-J.; Wahid, A. Rice direct seeding: Experiences, challenges and opportunities. Soil Tillage Res. 2011, 111, 87–98. [Google Scholar] [CrossRef]
- Bourioug, M.; Ezzaza, K.; Bouabid, R.; Alaoui-Mhamdi, M.; Bungau, S.; Bourgeade, P.; Alaoui-Sossé, L.; Alaoui-Sossé, B.; Aleya, L. Influence of hydro-and osmo-priming on sunflower seeds to break dormancy and improve crop performance under water stress. Environ. Sci. Pollut. Res. 2020, 27, 13215–13226. [Google Scholar] [CrossRef]
- Ghassemi-Golezani, K.; Yaghoubian, I.; Raei, Y. The impact of hydro-priming duration on seed invigoration and field emergence of milk thistle. J. Biodivers. Environ. Sci. 2016, 9, 229–234. [Google Scholar]
- Farooq, M.; Barsa, S.M.; Wahid, A. Priming of field-sown rice seed enhances germination, seedling establishment, allometry and yield. Plant Growth Regul. 2006, 49, 285–294. [Google Scholar] [CrossRef]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil beneficial bacteria and their role in plant growth promotion: A review. Ann. Microbiol. 2010, 60, 579–598. [Google Scholar] [CrossRef]
- Gouda, S.; Kerry, R.G.; Das, G.; Paramithiotis, S.; Shin, H.-S.; Patra, J.K. Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture. Microbiol. Res. 2018, 206, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Antar, M.; Gopal, P.; Msimbira, L.A.; Naamala, J.; Nazari, M.; Overbeek, W.; Backer, R.; Smith, D.L. Inter-organismal signaling in the rhizosphere. In Rhizosphere Biology: Interactions Between Microbes and Plants; Springer: Berlin/Heidelberg, Germany, 2021; pp. 255–293. [Google Scholar]
- Varma, A.; Kost, G.; Oelmüller, R. Piriformospora Indica: Sebacinales and Their Biotechnological Applications; Springer: Berlin, Germany, 2013; Volume 33. [Google Scholar]
- Dos Santos Garcia, J.; Dalmolin, Â.C.; França, M.G.C.; Mangabeira, P.A.O. Different salt concentrations induce alterations both in photosynthetic parameters and salt gland activity in leaves of the mangrove Avicennia schaueriana. Ecotoxicol. Environ. Saf. 2017, 141, 70–74. [Google Scholar] [CrossRef]
- Ansari, M.; Gill, S.; Tuteja, N. Piriformospora indica a powerful tool for crop improvement. Proc. Indian Natl. Sci. Acad. 2014, 80, 317–324. [Google Scholar] [CrossRef]
- Mensah, R.A.; Li, D.; Liu, F.; Tian, N.; Sun, X.; Hao, X.; Lai, Z.; Cheng, C. Versatile Piriformospora indica and its potential applications in horticultural crops. Hortic. Plant J. 2020, 6, 111–121. [Google Scholar] [CrossRef]
- Dolatabadi, H.K.; Goltapeh, E.M.; Moieni, A.; Jaimand, K.; Sardrood, B.P.; Varma, A. Effect of Piriformospora indica and Sebacina vermifera on plant growth and essential oil yield in Thymus vulgaris in vitro and in vivo experiments. Symbiosis 2011, 53, 29–35. [Google Scholar] [CrossRef]
- Saravi, H.B.; Gholami, A.; Pirdashti, H.; Firouzabadi, M.B.; Asghari, H. The response of stevia (Stevia rebaudiana Bertoni) photosystem II photochemistry to fungi symbiosis and spermidine application under saline water irrigation. Russ. Agric. Sci. 2021, 47, 32–36. [Google Scholar] [CrossRef]
- Ghassemi-Golezani, K.; Yaghoubian, I.; Raei, Y. Influence of hydro-priming duration on field performance of milk thistle under water stress. Adv. Biores. 2016, 7, 61–65. [Google Scholar] [CrossRef]
- Mosavikia, A.A.; Mosavi, S.G.; Seghatoleslami, M.; Baradaran, R. Chitosan nanoparticle and pyridoxine seed priming improves tolerance to salinity in milk thistle seedling. Not. Bot. Horti Agrobot. Cluj-Napoca. 2020, 48, 221–233. [Google Scholar] [CrossRef] [Green Version]
- Suriya-arunroj, D.; Supapoj, N.; Toojinda, T.; Vanavichit, A. Relative leaf water content as an efficient method for evaluating rice cultivars for tolerance to salt stress. ScienceAsia 2004, 30, 411–415. [Google Scholar] [CrossRef]
- Negrão, S.; Schmöckel, S.; Tester, M. Evaluating physiological responses of plants to salinity stress. Ann. Bot. 2017, 119, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Keyvan, S. The effects of drought stress on yield, relative water content, proline, soluble carbohydrates and chlorophyll of bread wheat cultivars. Anim. Plant Sci. 2010, 8, 1051–1060. [Google Scholar]
- Oelmüller, R.; Sherameti, I.; Tripathi, S.; Varma, A. Piriformospora indica, a cultivable root endophyte with multiple biotechnological applications. Symbiosis 2009, 49, 1–17. [Google Scholar] [CrossRef]
- Varma, A.; Bakshi, M.; Lou, B.; Hartmann, A.; Oelmueller, R. Piriformospora indica: A novel plant growth-promoting mycorrhizal fungus. Agric. Res. 2012, 1, 117–131. [Google Scholar] [CrossRef]
- Hosseinzadeh-Mahootchi, A.; Ghassemi-Golezani, K. The impact of seed priming and aging on physiological performance of chickpea under different irrigation treatments. Plant Breed. Seed Sci. 2012, 67, 13–26. [Google Scholar] [CrossRef]
- Ghassemi, S.; Farhangi-Abriz, S.; Faegi-Analou, R.; Ghorbanpour, M.; Lajayer, B.A.; Nutrition, P. Monitoring cell energy, physiological functions and grain yield in field-grown mung bean exposed to exogenously applied polyamines under drought stress. J. Soil Sci. Plant Nutr. 2018, 18, 1108–1125. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.; Jiang, D.; Liu, F.; Dai, T.; Jing, Q.; Cao, W. Effects of salt and waterlogging stresses and their combination on leaf photosynthesis, chloroplast ATP synthesis, and antioxidant capacity in wheat. Plant Sci. 2009, 176, 575–582. [Google Scholar] [CrossRef]
- Sfakianaki, M.; Sfichi, L.; Kotzabasis, K. The involvement of LHCII-associated polyamines in the response of the photosynthetic apparatus to low temperature. J. Photochem. Photobiol. B. 2006, 84, 181–188. [Google Scholar] [CrossRef]
- Fermer, E.; Muller, M. Reactive oxygen species mediated lipid peroxidation and RES activated signalling. Annu. Rev. Plant Biol. 2013, 4, 429–450. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Mohammadi, H.; Ghorbanpour, M.; Brestic, M. Exogenous putrescine changes redox regulations and essential oil constituents in field-grown Thymus vulgaris L. under well-watered and drought stress conditions. Ind. Crops Prod. 2018, 122, 119–132. [Google Scholar] [CrossRef]
- Amooaghaie, R. The effect of hydro and osmopriming on alfalfa seed germination and antioxidant defenses under salt stress. Afr. J. Biotechnol. 2011, 10, 6269–6275. [Google Scholar] [CrossRef]
- Kamithi, K.; Wachira, F.; Kibe, A. Effects of different priming methods and priming durations on enzyme activities in germinating chickpea (Cicer arietinum L.). Am. J. Appl. Sci. 2016, 1, 1–9. [Google Scholar]
- El-Araby, M.; Hegazi, A. Responses of tomato seeds to hydro-and osmo-priming, and possible relations of some antioxidant enzymes and endogenous polymine fractions. Egypt J. Biol. 2004, 6, 81–93. [Google Scholar]
- Pryor, W.A.; Stanley, J. Suggested mechanism for the production of malonaldehyde during the autoxidation of polyunsaturated fatty acids. Nonenzymic production of prostaglandin endoperoxides during autoxidation. J. Org. Chem. 1975, 40, 3615–3617. [Google Scholar] [CrossRef]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef]
- Sun, C.; Johnson, J.M.; Cai, D.; Sherameti, I.; Oelmüller, R.; Lou, B. Piriformospora indica confers drought tolerance in Chinese cabbage leaves by stimulating antioxidant enzymes, the expression of drought-related genes and the plastid-localized CAS protein. J. Plant Physiol. 2010, 167, 1009–1017. [Google Scholar] [CrossRef] [PubMed]
- Shah, T.; Latif, S.; Saeed, F.; Ali, I.; Ullah, S.; Alsahli, A.A.; Jan, S.; Ahmad, P. Seed priming with titanium dioxide nanoparticles enhances seed vigor, leaf water status, and antioxidant enzyme activities in maize (Zea mays L.) under salinity stress. J. King Saud Univ. Sci. 2021, 33, 101207. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, P.; Hashem, A.; Abd-Allah, E.F.; Alqarawi, A.; John, R.; Egamberdieva, D.; Gucel, S. Role of Trichoderma harzianum in mitigating NaCl stress in Indian mustard (Brassica juncea L) through antioxidative defense system. Front. Plant Sci. 2015, 6, 868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bose, J.; Rodrigo-Moreno, A.; Shabala, S. ROS homeostasis in halophytes in the context of salinity stress tolerance. J. Exp. Bot. 2014, 65, 1241–1257. [Google Scholar] [CrossRef]
- Siadat, S.A.; Moosavi, A.; Sharafizadeh. Effects of seed priming on antioxidant activity and germination characteristics of maize seeds under different ageing treatment. Res. J. Seed Sci. 2012, 5, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Kerepesi, I.; Galiba, G. Osmotic and salt stress-induced alteration in soluble carbohydrate content in wheat seedlings. Crop Sci. 2000, 40, 482–487. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Ghassemi-Golezani, K. How can salicylic acid and jasmonic acid mitigate salt toxicity in soybean plants? Ecotoxicol. Environ. Saf. 2018, 147, 1010–1016. [Google Scholar] [CrossRef] [PubMed]
- Capron, I.; Corbineau, F.; Dacher, F.; Job, C.; Côme, D.; Job, D. Sugarbeet seed priming: Effects of priming conditions on germination, solubilization of 11-S globulin and accumulation of LEA proteins. Seed Sci. Res. 2000, 10, 243–254. [Google Scholar] [CrossRef]
- Verma, J.; Srivastava, A. Physiological basis of salt stress resistance in pigeon pea (Cajanus cajan L.)-II. Pre-sowing seed soaking treatment in regulating early seedling metabolism during seed germination. Plant Physiol. Biochem. 1998, 25, 89–94. [Google Scholar]
- Khademian, R.; Asghari, B.; Sedaghati, B.; Yaghoubian, Y. Plant beneficial rhizospheric microorganisms (PBRMs) mitigate deleterious effects of salinity in sesame (Sesamum indicum L.): Physio-biochemical properties, fatty acids composition and secondary metabolites content. Ind. Crops Prod. 2019, 136, 129–139. [Google Scholar] [CrossRef]
- Bagheri, A.A.; Saadatmand, S.; Niknam, V.; Nejadsatari, T.; Babaeizad, V. Effect of endophytic fungus, Piriformospora indica, on growth and activity of antioxidant enzymes of rice (Oryza sativa L.) under salinity stress. Int. J. Adv. Biol. Biomed. Res. 2013, 1, 1337–1350. [Google Scholar]
- Afshar, R.K.; Chaichi, M.R.; Jovini, M.A.; Jahanzad, E.; Hashemi, M. Accumulation of silymarin in milk thistle seeds under drought stress. Planta 2015, 242, 539–543. [Google Scholar] [CrossRef]
- Laoué, J.; Fernandez, C.; Ormeño, E. Plant Flavonoids in Mediterranean Species: A Focus on Flavonols as Protective Metabolites under Climate Stress. Plants 2022, 11, 172. [Google Scholar] [CrossRef]
- Selmar, D.; Kleinwächter, M. Influencing the product quality by deliberately applying drought stress during the cultivation of medicinal plants. Ind. Crops Prod. 2013, 42, 558–566. [Google Scholar] [CrossRef]
- Sarmoum, R.; Haid, S.; Biche, M.; Djazouli, Z.; Zebib, B.; Merah, O. Effect of salinity and water stress on the essential oil components of rosemary (Rosmarinus officinalis L.). Agronomy 2019, 9, 214. [Google Scholar] [CrossRef] [Green Version]
- Mahfouz, S.; Sharaf-Eldin, M. Effect of mineral vs. biofertilizer on growth, yield, and essential oil content of fennel (Foeniculum vulgare Mill). Int. Agrophys. 2007, 21, 361–366. [Google Scholar]
- Banchio, E.; Bogino, P.C.; Zygadlo, J.; Giordano, W. Plant growth promoting rhizobacteria improve growth and essential oil yield in Origanum majorana L. Biochem. Syst. Ecol. 2008, 36, 766–771. [Google Scholar] [CrossRef]
- Khademian, R.; Ghassemi, S.; Asghari, B. Bio-fertilizer improves physio-biochemical characteristics and grain yield of safflower (Carthamus tinctorius L.) under drought stress. Russ. Agric. Sci. 2019, 45, 458–463. [Google Scholar] [CrossRef]
- Kapoor, R.; Chaudhary, V.; Bhatnagar, A. Effects of arbuscular mycorrhiza and phosphorus application on artemisinin concentration in Artemisia annua L. Mycorrhiza 2007, 17, 581–587. [Google Scholar] [CrossRef]
- Mandal, S.; Upadhyay, S.; Singh, V.P.; Kapoor, R. Enhanced production of steviol glycosides in mycorrhizal plants: A concerted effect of arbuscular mycorrhizal symbiosis on transcription of biosynthetic genes. Plant Physiol. Biochem. 2015, 89, 100–106. [Google Scholar] [CrossRef]
- Baldi, A.; Dixit, V. Yield enhancement strategies for artemisinin production by suspension cultures of Artemisia annua. Bioresour. Technol. 2008, 99, 4609–4614. [Google Scholar] [CrossRef]
- Baldi, A.; Farkya, S.; Jain, A.; Gupta, N.; Mehra, R.; Datta, V.; Srivastava, A.K.; Bisaria, V.S. Enhanced production of podophyllotoxins by co-culture of transformed Linum album cells with plant growth-promoting fungi. Pure Appl. Chem. 2010, 82, 227–241. [Google Scholar] [CrossRef]
- Farkya, S.; Baldi, A.; Kumar, V.; Datta, V.; Mehra, R.; Gupta, N.; Jain, A.; Srivastava, A.; Bisaria, V. Impact of symbiotic fungi on production of secondary metabolites by plant cell culture. Asia Pac. J. Mol. Biol. Biotechnol. 2010, 18, 51–53. [Google Scholar]
- Yaghoubian, I.; Msimbira, L.A.; Smith, D.L. Cell-free supernatant of Bacillus strains (CFS) can improve seed vigor index of corn under salinity stress. Front. Sustain. Food Syst. 2022, 92, 857643. [Google Scholar] [CrossRef]
- Saravi, H.B.; Gholami, A.; Pirdashti, H.; Firouzabadi, M.B.; Asghari, H.; Yaghoubian, Y. Products. Improvement of salt tolerance in Stevia rebaudiana by co-application of endophytic fungi and exogenous spermidine. Ind. Crops Prod. 2022, 177, 114443. [Google Scholar] [CrossRef]
- Yaghoubian, I.; Ghassemi, S.; Nazari, M.; Raei, Y.; Smith, D.L. Response of physiological traits, antioxidant enzymes and nutrient uptake of soybean to Azotobacter Chroococcum and zinc sulfate under salinity. S. Afr. J. Bot. 2021, 143, 42–51. [Google Scholar] [CrossRef]
- Yaghoubian, Y.; Siadat, S.A.; Telavat, M.R.M.; Pirdashti, H.; Yaghoubian, I. Bio-removal of cadmium from aqueous solutions by filamentous fungi: Trichoderma spp. and Piriformospora indica. Environ. Sci. Pollut. Res. 2019, 26, 7863–7872. [Google Scholar] [CrossRef] [PubMed]
- Ghassemi-Golezani, K.; Ghassemi, S.; Yaghoubian, I. Salicylic ccid regulate physiological performance of milk thistle (Silybum marianum L.) under water stress. Adv. Biores. 2016, 7, 34–40. [Google Scholar] [CrossRef]
- Ghassemi, S.; Yaghoubian, I.; Yaghoubian, Y. Hydro-priming effects on safflower under water limitation: Some physiological traits, grain and oil yields. J. Biodivers. Environ. Sci. 2016, 9, 367–375. [Google Scholar]
- Larsson, C.-M.; Olssonl, T. Firefly assay of adenine nucleotides from algae: Comparison of extraction methods. Plant Cell Physiol. 1979, 20, 145–155. [Google Scholar]
- Lobo, A.K.M.; de Oliveira Martins, M.; Neto, M.C.L.; Machado, E.C.; Ribeiro, R.V.; Silveira, J.A.G. Exogenous sucrose supply changes sugar metabolism and reduces photosynthesis of sugarcane through the down-regulation of Rubisco abundance and activity. J. Plant Physiol. 2015, 179, 113–121. [Google Scholar] [CrossRef]
- Rao, K.M.; Sresty, T. Antioxidative parameters in the seedlings of pigeonpea (Cajanus cajan (L.) Millspaugh) in response to Zn and Ni stresses. Plant Sci. 2000, 157, 113–128. [Google Scholar]
- Dzung, N.A.; Khanh, V.T.P.; Dzung, T.T. Research on impact of chitosan oligomers on biophysical characteristics, growth, development and drought resistance of coffee. Carbohydr. Polym. 2011, 84, 751–755. [Google Scholar] [CrossRef]
- Grieve, C.; Grattan, S. Rapid assay for determination of water soluble quaternary ammonium compounds. Plant Soil 1983, 70, 303–307. [Google Scholar] [CrossRef]
- Sitaramaraju, Y.; Riadi, A.; D’Autry, W.; Wolfs, K.; Hoogmartens, J.; Van Schepdael, A.; Adams, E. Evaluation of the European pharmacopoeia method for control of residual solvents in some antibiotics. J. Pharm. Biomed. Anal. 2008, 48, 113–119. [Google Scholar] [CrossRef] [PubMed]
- AOCS. Methods and Recommended Practices of the American Oil Chemists’ Society; AOCS Press: Champaign, IL, USA, 1993; pp. 573–577. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Salinity | Seed Priming | Silymarin | Silybin | Isosilybin | Silydianin | Taxidolin | Silychristin |
|---|---|---|---|---|---|---|---|
| Non-saline | Control | 16.28 ± 1.88 e | 9.11 ± 0.22 e | 2.21 ± 0.28 h | 1.02 ± 0.08 f | 0.68 ± 0.10 e | 0.96 ± 0.12 d |
| Hydro | 18.18 ± 0.39 cde | 10.13 ± 0.87 de | 2.82 ± 0.22 fg | 1.12 ± 0.10 f | 0.74 ± 0.08 de | 0.01 ± 0.10 cd | |
| Az | 17.49 ± 1.66 de | 10.08 ± 0.81 de | 2.72 ± 0.35 gh | 1.28 ± 0.16 def | 0.77 ± 0.03 cde | 1.08 ± 0.05 cd | |
| Pi | 18.94 ± 0.98 cde | 10.49 ± 0.69 cde | 2.96 ± 0.21 efg | 1.41 ± 0.19 def | 0.83 ± 0.05 bcd | 1.18 ± 0.17 bcd | |
| 60 mM | Control | 18.14 ± 2.17 cde | 10.65 ± 0.75 cde | 2.66 ± 0.10 gh | 1.24 ± 0.04 | 0.75 ± 0.04 de | 1.04 ± 0.15 cd |
| Hydro | 20.63 ± 1.97 bcd | 12.06 ± 0.91 bcd | 3.84 ± 0.11 cd | 1.45 ± 0.21 def | 0.95 ± 0.09 bc | 1.16 ± 0.11 bcd | |
| Az | 20.96 ± 0.79 bcd | 12.18 ± 1.85 bcd | 3.51 ± 0.31 de | 1.66 ± 0.35 cde | 0.91 ± 0.12 bcd | 1.24 ± 0.21 bcd | |
| Pi | 21.38 ± 2.90 bc | 12.82 ± 0.67 b | 3.76 ± 0.33 cd | 1.82 ± 0.27 c | 1.02 ± 0.11 b | 1.37 ± 0.13 b | |
| 120 mM | Control | 22.31 ± 1.26 b | 12.47 ± 0.87 bc | 3.38 ± 0.54 def | 1.72 ± 0.32 cd | 0.98 ± 0.04 b | 1.27 ± 0.12 bc |
| Hydro | 27.44 ± 1.92 a | 16.01 ± 1.48 a | 4.24 ± 0.14 bc | 2.36 ± 0.31 b | 1.37 ± 0.11 a | 1.68 ± 0.25 a | |
| Az | 28.05 ± 2.16 a | 16.43 ± 2.07 a | 4.56 ± 0.37 ab | 2.71 ± 0.29 ab | 1.41 ± 0.14 a | 1.79 ± 0.06 a | |
| Pi | 29.84 ± 2.64 a | 17.52 ± 1.71 a | 4.83 ± 0.45 a | 2.84 ± 0.36 a | 1.52 ± 0.22 a | 1.94 ± 0.20 a |
| Salinity | Seed Priming | Oil Content | Palmitic Acid | Stearic Acid | Oleic Acid | Linoleic Acid | Linolenic Acid | UI |
|---|---|---|---|---|---|---|---|---|
| Non-saline | Control | 30.14 ± 3.12 cd | 7.82 ± 0.35 b | 5.68 ± 0.14 c | 29.62 ± 0.62 def | 50.68 ± 0.57 bc | 0.45 ± 0.03 ab | 1.32 ± 0.03 ab |
| Hydro | 31.27 ± 2.02 cd | 7.71 ± 0.27 b | 5.52 ± 0.41 c | 29.02 ± 0.52 ef | 51.13 ± 0.88 ab | 0.47 ± 0.05 ab | 1.33 ± 0.03 ab | |
| Az | 34.49 ± 2.62 bc | 7.75 ± 0.25 b | 5.41 ± 0.20 d | 28.11 ± 1.08 f | 51.95 ± 0.43 ab | 0.49 ± 0.06 a | 1.33 ± 0.02 ab | |
| Pi | 34.83 ± 1.29 bc | 7.81 ± 0.41 b | 5.47 ± 0.34 cd | 28.04 ± 1.09 f | 52.34 ± 0.55 a | 0.49 ± 0.07 a | 1.34 ± 0.02 a | |
| 60 mM | Control | 32.20 ± 3.29 c | 8.02 ± 0.47 ab | 5.97 ± 0.64 bc | 30.71 ± 0.31 bc | 48.46 ± 0.94 d | 0.41 ± 0.03 abc | 1.28 ± 0.01 b |
| Hydro | 37.45 ± 2.71 a | 7.92 ± 0.27 b | 5.73 ± 0.35 c | 29.51 ± 0.85 def | 50.51 ± 1.22 bc | 0.44 ± 0.02 ab | 1.32 ± 0.02 ab | |
| Az | 38.84 ± 3.55 a | 7.81 ± 0.15 b | 5.52 ± 0.17 c | 28.48 ± 0.69 ef | 50.62 ± 0.44 bc | 0.45 ± 0.04 ab | 1.31 ± 0.02 ab | |
| Pi | 39.18 ± 2.16 a | 7.75 ± 0.53 b | 5.54 ± 0.45 c | 28.22 ± 0.94 f | 50.45 ± 0.44 bc | 0.47 ± 0.04 ab | 1.30 ± 0.03 ab | |
| 120 mM | Control | 26.28 ± 0.88 d | 8.64 ± 0.32 a | 6.83 ± 0.26 a | 35.86 ± 1.22 a | 43.04 ± 1.74 e | 0.35 ± 0.02 c | 1.23 ± 0.02 c |
| Hydro | 37.05 ± 1.54 ab | 8.11 ± 0.42 ab | 6.40 ± 0.35 ab | 31.54 ± 1.28 b | 48.72 ± 0.51 d | 0.40 ± 0.03 bc | 1.30 ± 0.02 ab | |
| Az | 37.51 ± 1.89 a | 8.02 ± 0.30 ab | 5.98 ± 0.15 bc | 30.13 ± 0.36 bcd | 49.48 ± 0.37 cd | 0.43 ± 0.04 ab | 1.30 ± 0.03 ab | |
| Pi | 38.11 ± 2.19 a | 7.93 ± 0.29 b | 5.86 ± 0.16 bc | 29.98 ± 0.33 cde | 50.02 ± 1.02 cd | 0.48 ± 0.03 ab | 1.31 ± 0.03 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yaghoubian, I.; Antar, M.; Ghassemi, S.; Modarres-Sanavy, S.A.M.; Smith, D.L. The Effects of Hydro-Priming and Colonization with Piriformospora indica and Azotobacter chroococcum on Physio-Biochemical Traits, Flavonolignans and Fatty Acids Composition of Milk Thistle (Silybum marianum) under Saline Conditions. Plants 2022, 11, 1281. https://doi.org/10.3390/plants11101281
Yaghoubian I, Antar M, Ghassemi S, Modarres-Sanavy SAM, Smith DL. The Effects of Hydro-Priming and Colonization with Piriformospora indica and Azotobacter chroococcum on Physio-Biochemical Traits, Flavonolignans and Fatty Acids Composition of Milk Thistle (Silybum marianum) under Saline Conditions. Plants. 2022; 11(10):1281. https://doi.org/10.3390/plants11101281
Chicago/Turabian StyleYaghoubian, Iraj, Mohammed Antar, Saeid Ghassemi, Seyed Ali Mohammad Modarres-Sanavy, and Donald L. Smith. 2022. "The Effects of Hydro-Priming and Colonization with Piriformospora indica and Azotobacter chroococcum on Physio-Biochemical Traits, Flavonolignans and Fatty Acids Composition of Milk Thistle (Silybum marianum) under Saline Conditions" Plants 11, no. 10: 1281. https://doi.org/10.3390/plants11101281