
Photoactivated TiO2 Nanocomposite Delays the Postharvest Ripening Phenomenon through Ethylene Metabolism and Related Physiological Changes in Capsicum Fruit

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
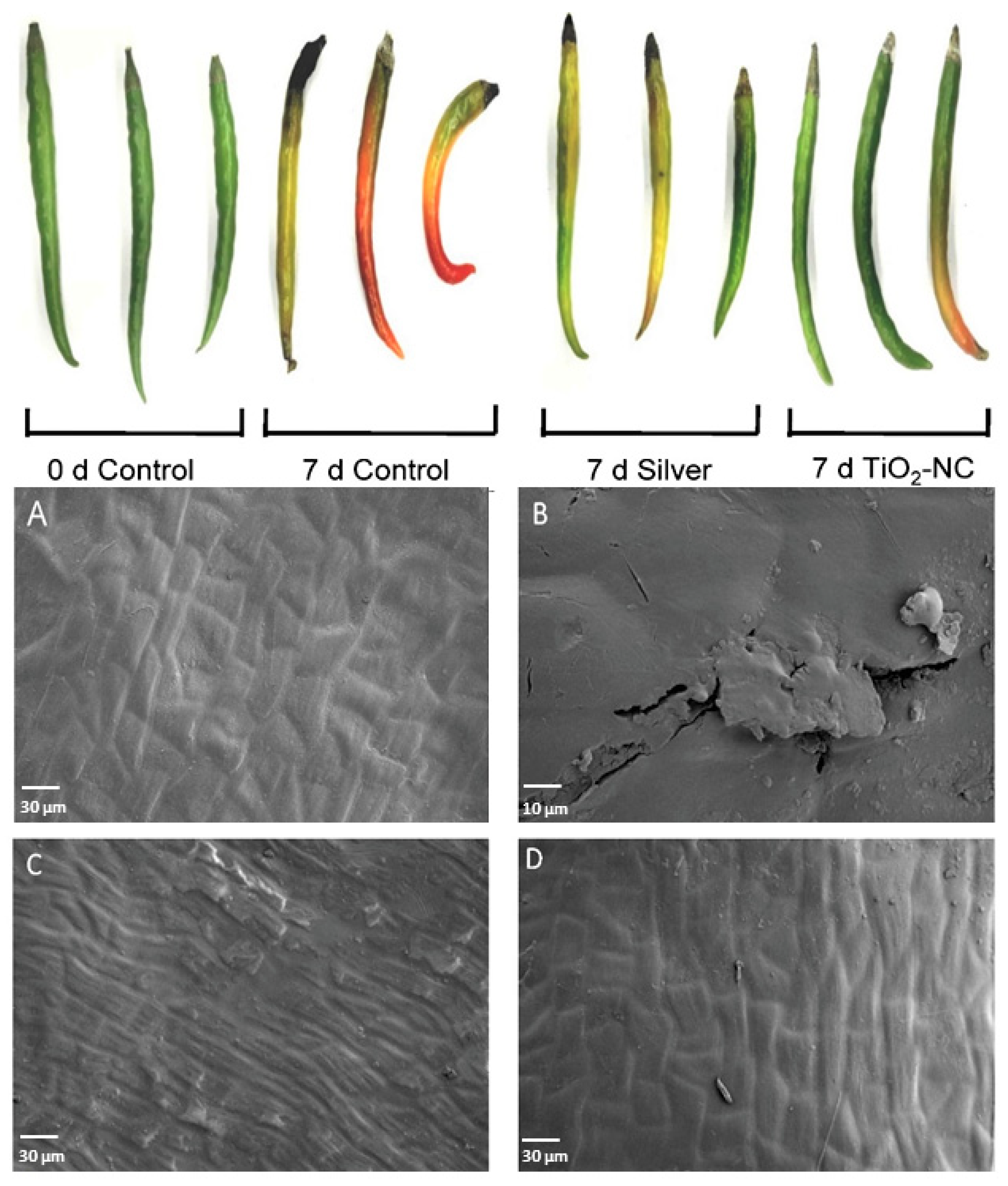
2.2. Study of Fruit Surface Morphology through Scanning Electron Microscopy (SEM)
2.3. Quality Parameters
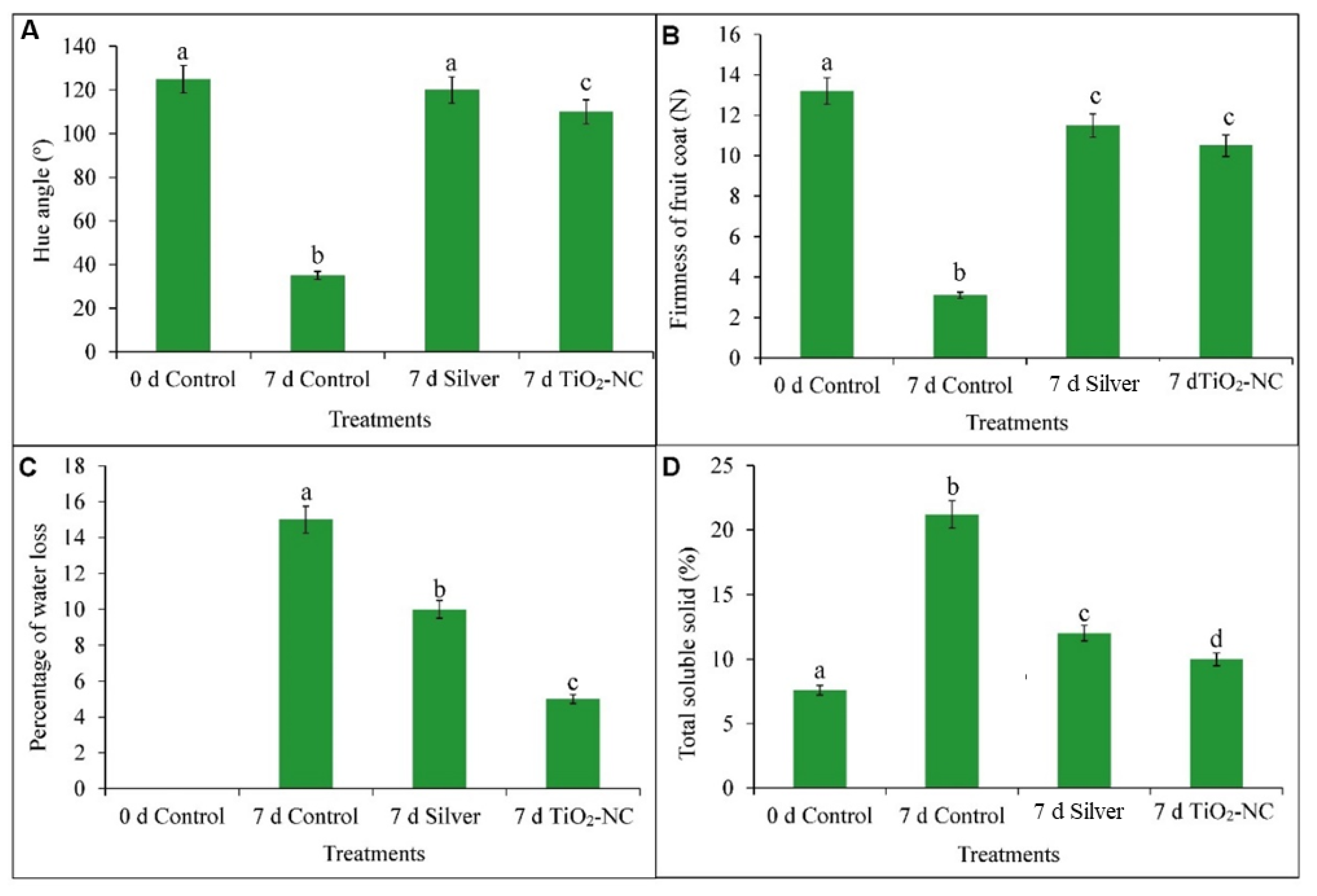
2.3.1. Firmness of Fruit Coat
2.3.2. Color of Fruit Coat
2.3.3. Percentage of Water Loss
2.3.4. Total Soluble Solid Content
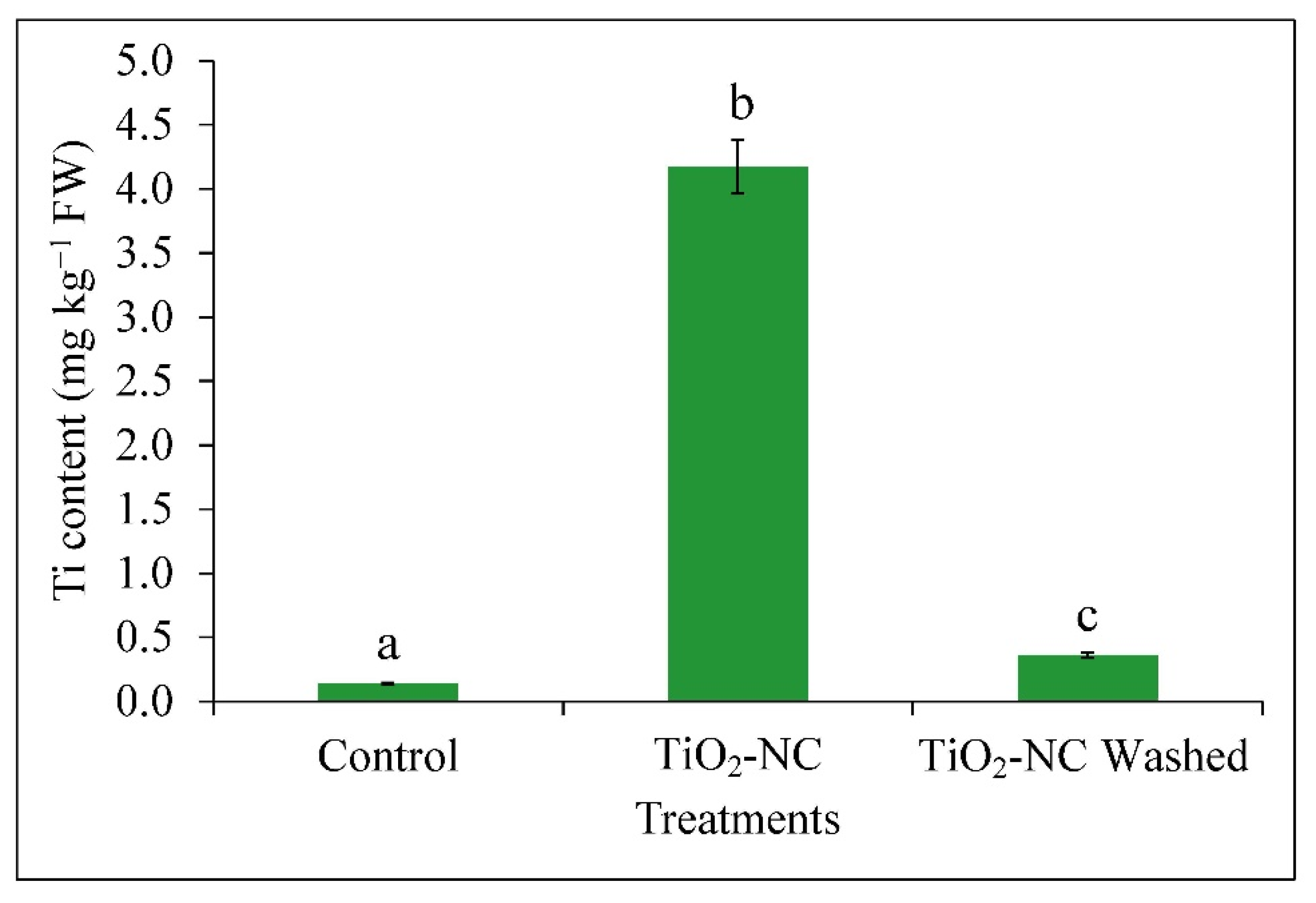
2.3.5. Quantification of Ti in Fruit
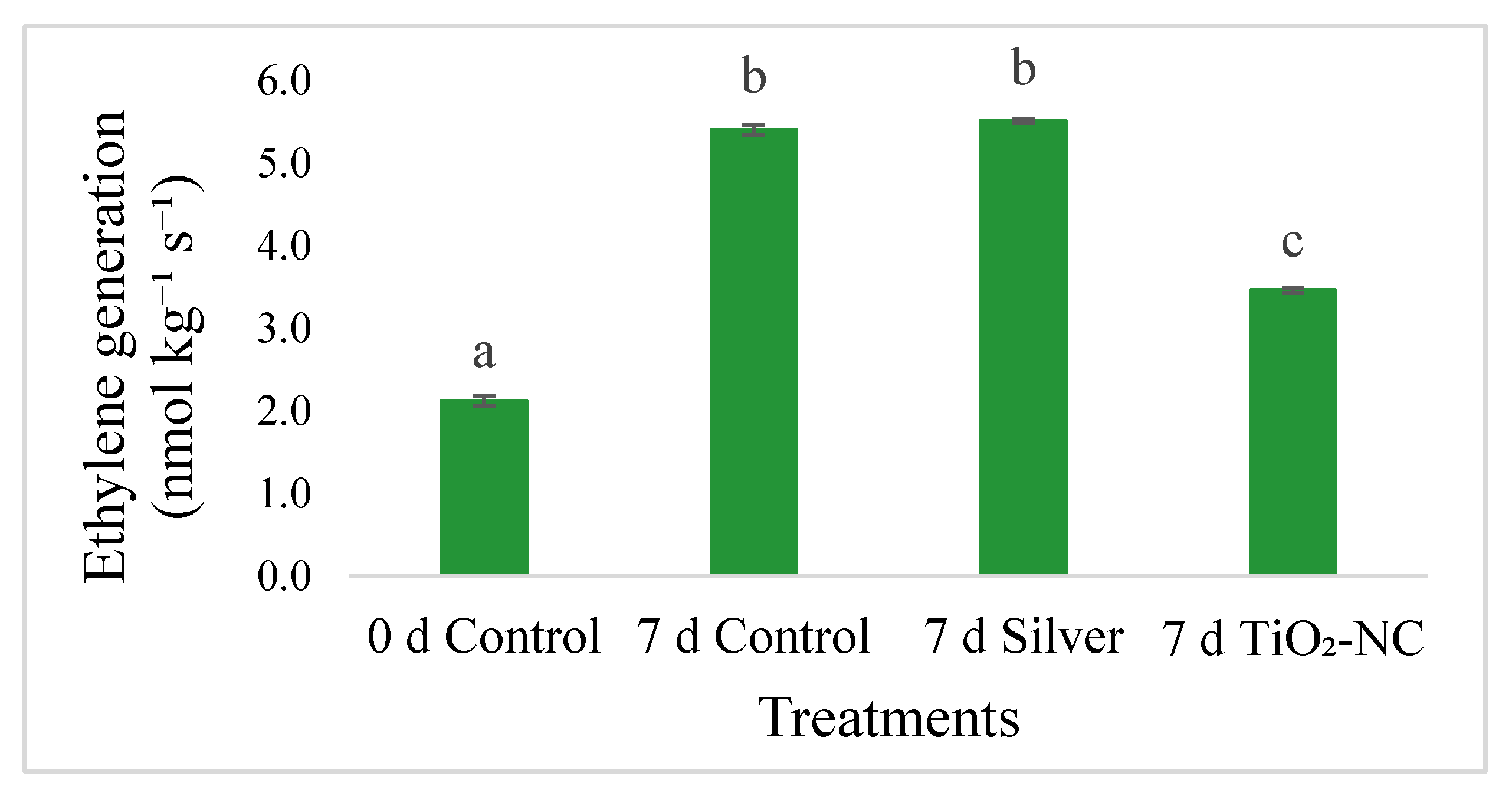
2.4. Ethylene Measurement
2.5. Preparation of RNA
2.6. Semi-Quantitative PCR
2.7. Quantification of ROS
2.7.1. Quantification of H2O2
2.7.2. O2•− Content
2.8. Quantification of Total Capsaicin Content
2.9. Quantification of Polyamine and TLC Separation
2.10. Enzyme Activity Assay
2.10.1. Esterase (EC 3.1.1.1) Activity
2.10.2. Polyamine Oxidase Activity and Diamine Oxidase Activity
2.11. Extraction of Essential Oil
2.12. GC-MS Analysis of Essential Oil
2.13. Characterization of TiO2-NP
2.14. Statistical Analysis
3. Results
3.1. Firmness, Color, Turgidity and Total Soluble Solids of Fruit on TiO2-NC Treatment
3.2. Absorption of Ti and Release of Ethylene from Fruit Tissues
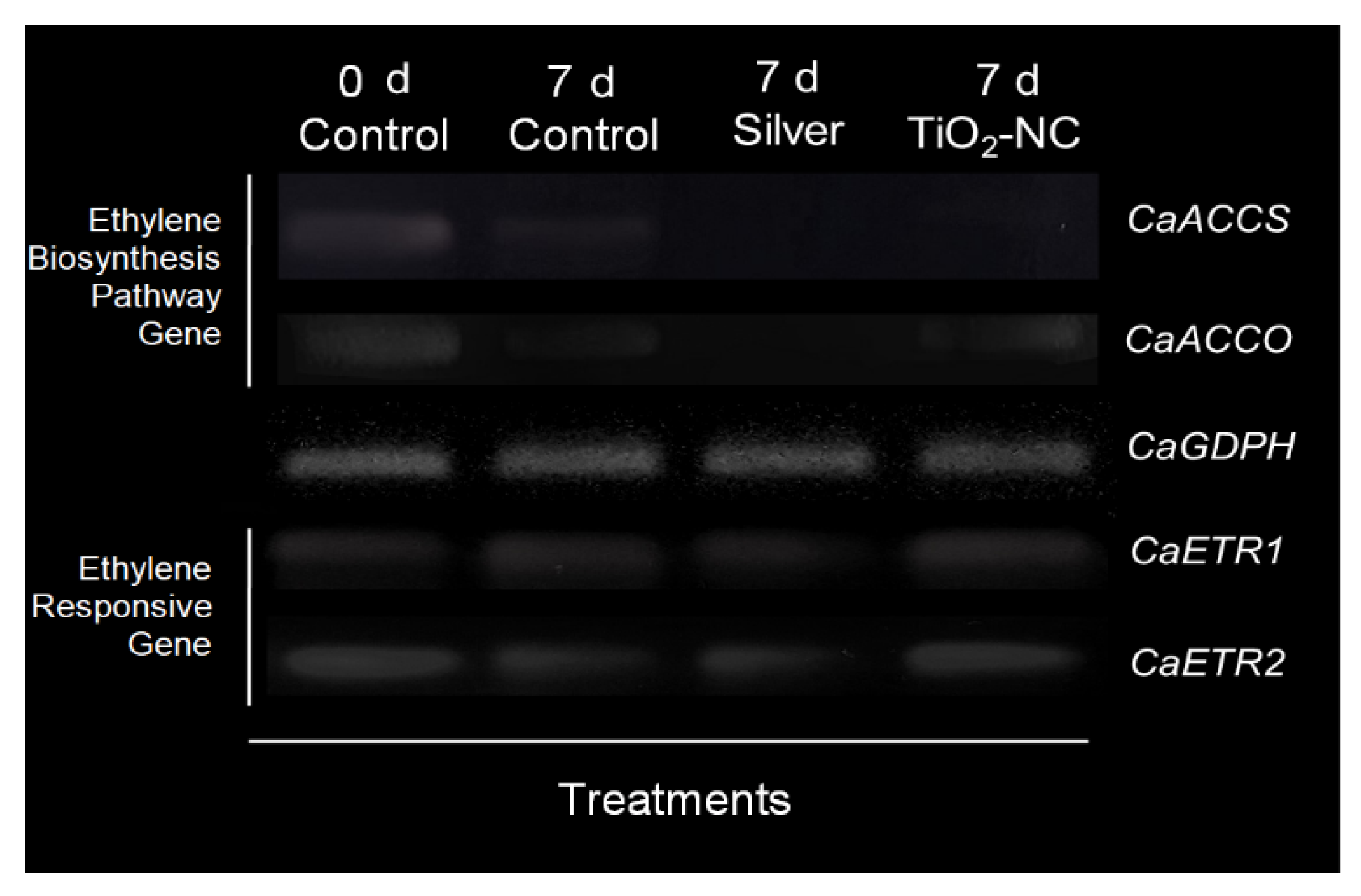
3.3. Expression of Genes for Ethylene Biosynthesis and Signaling
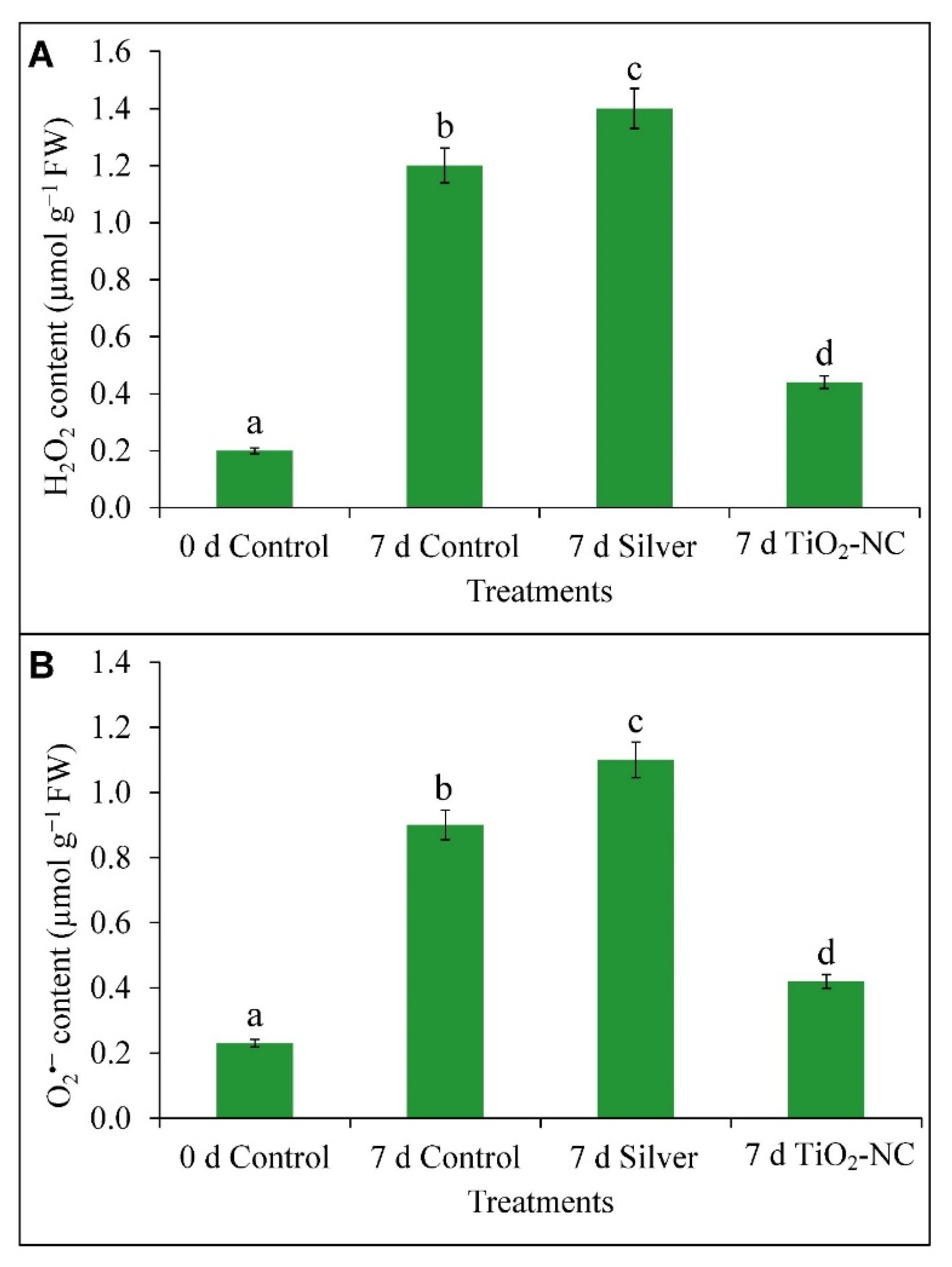
3.4. Accumulation of ROS (H2O2 and O2•−) in Fruit Tissues under Storage
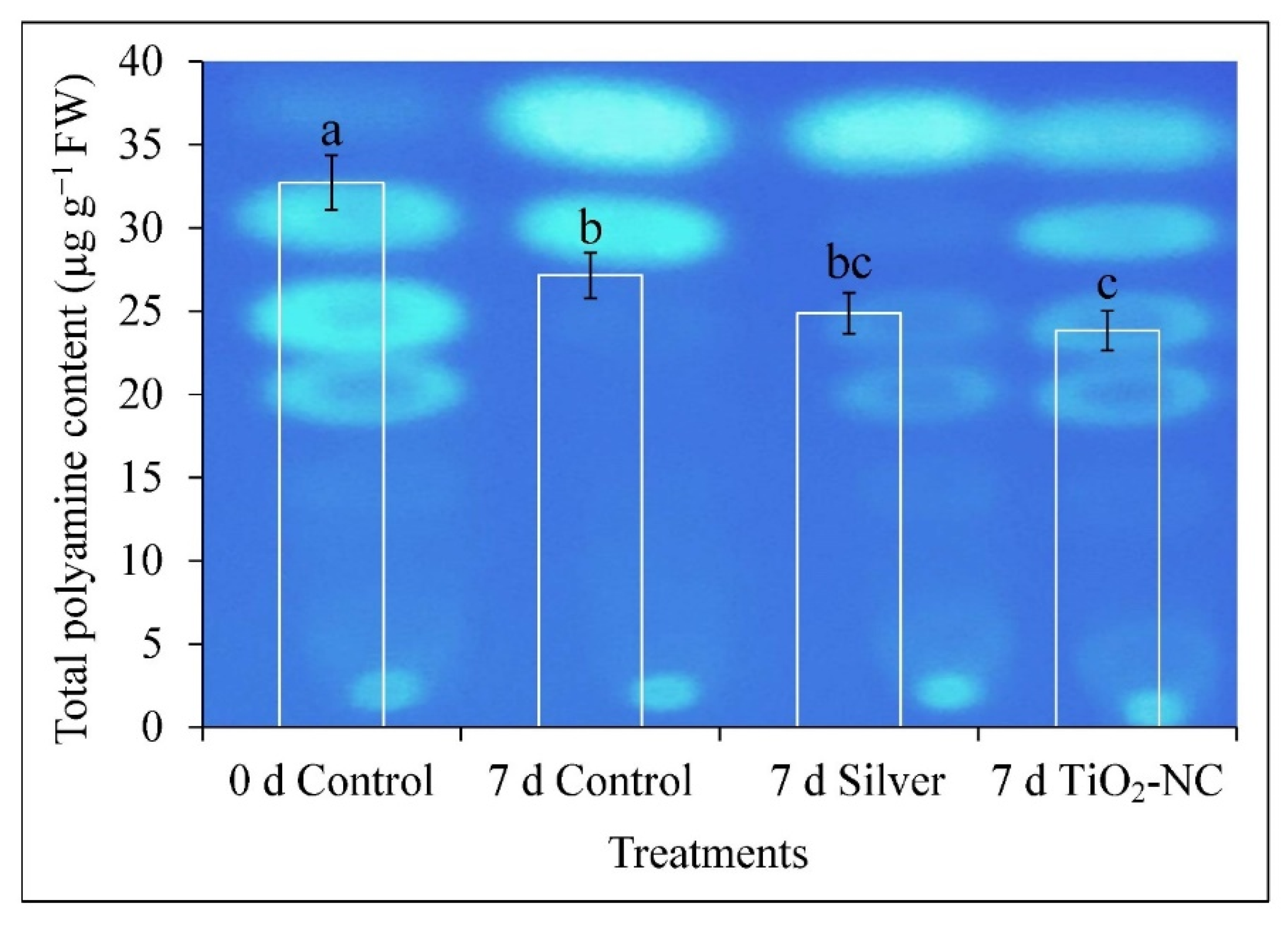
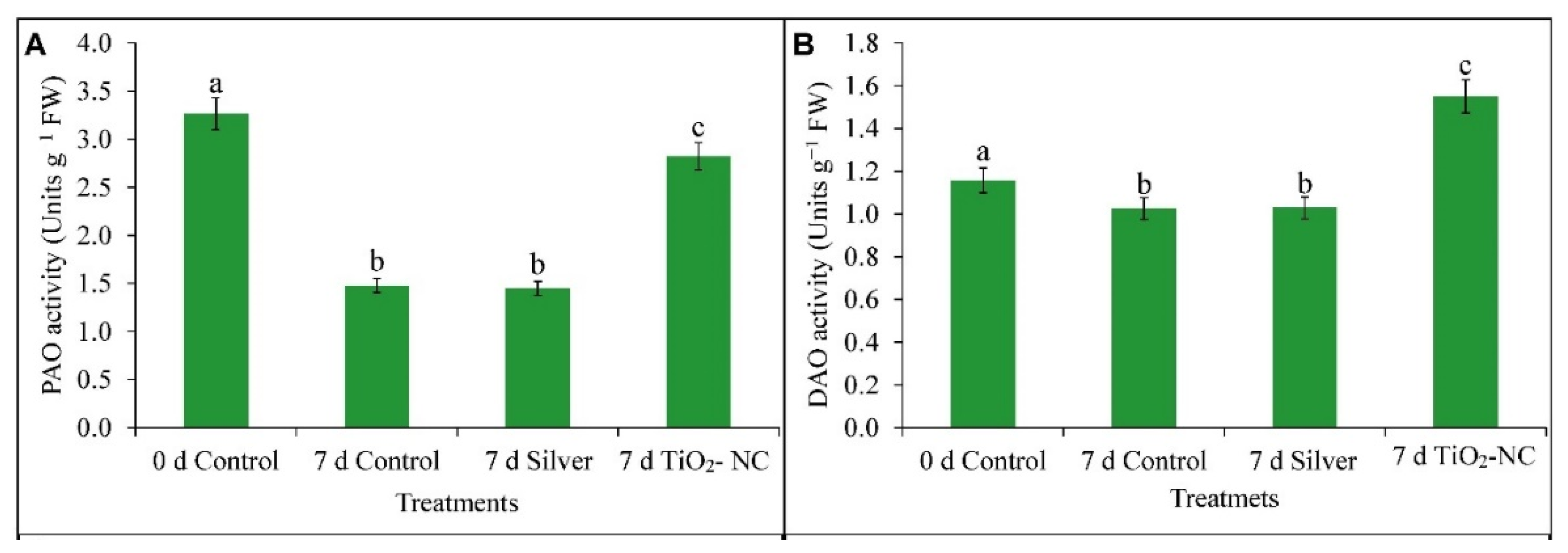
3.5. Catabolism of Polyamine during Ripening on Storage
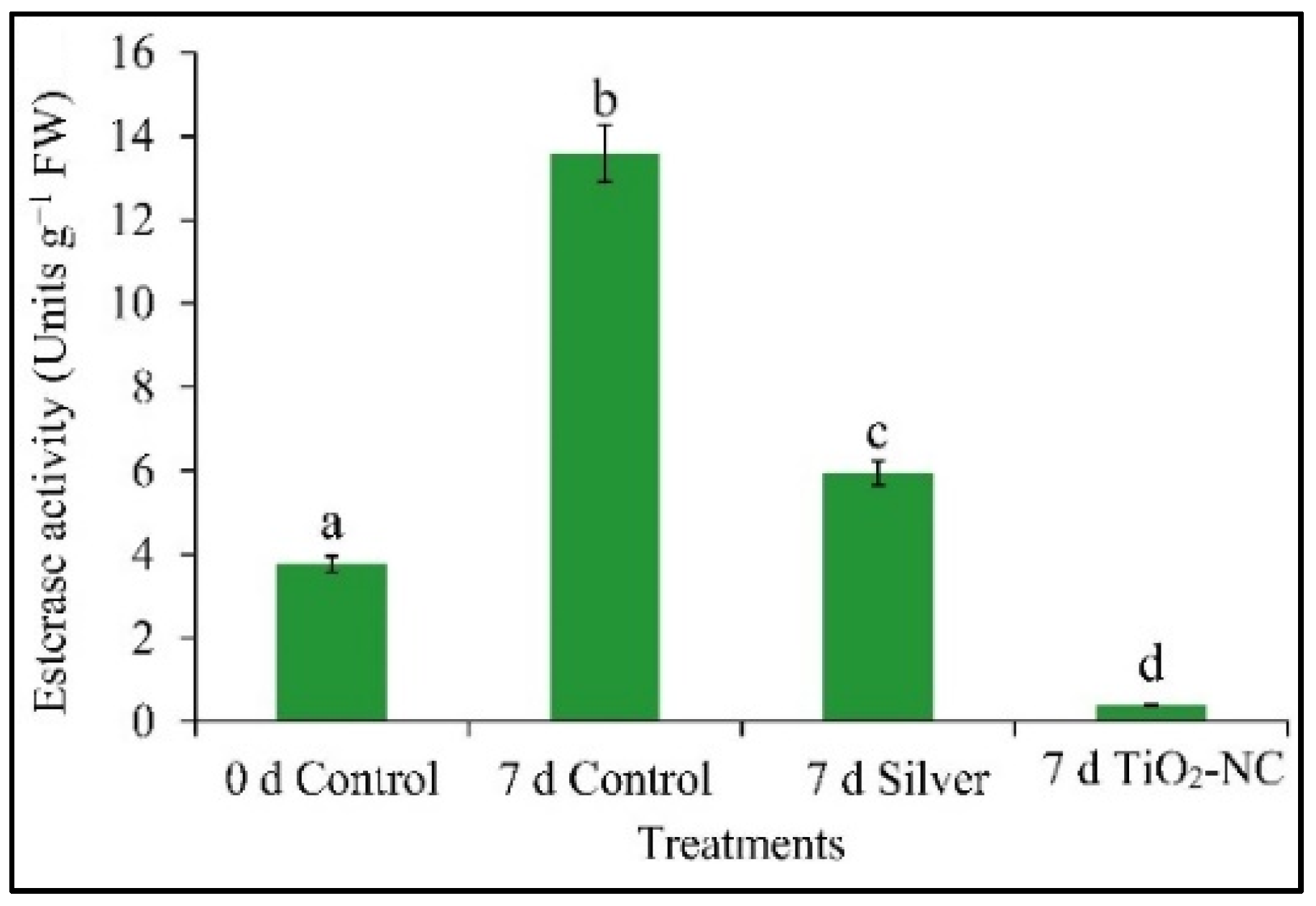
3.6. Esterase Activity and Its Polymorphic Expression on Fruit Softness
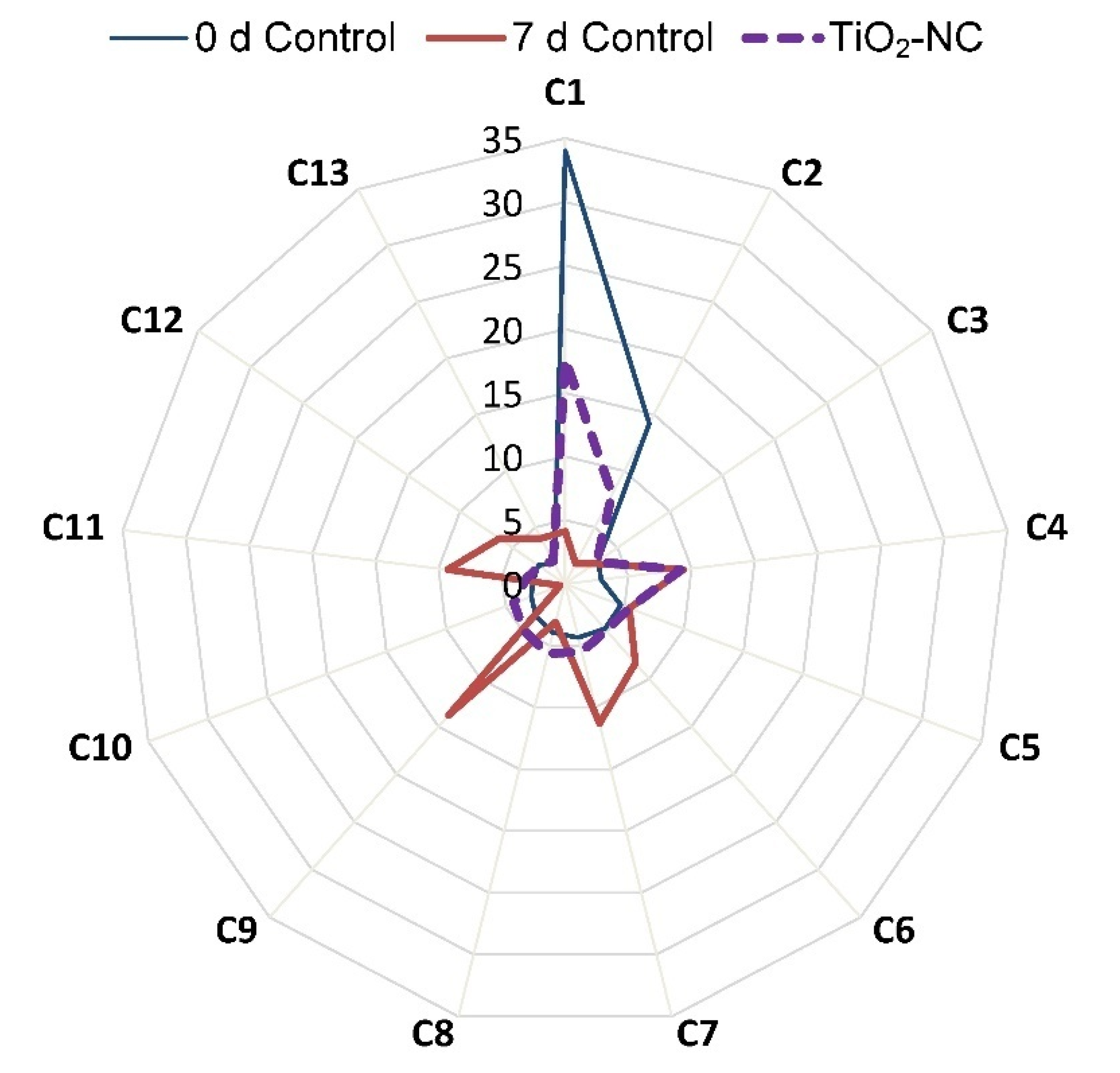
3.7. Changes in Essential Oil Contributing Major Metabolites in Ripening
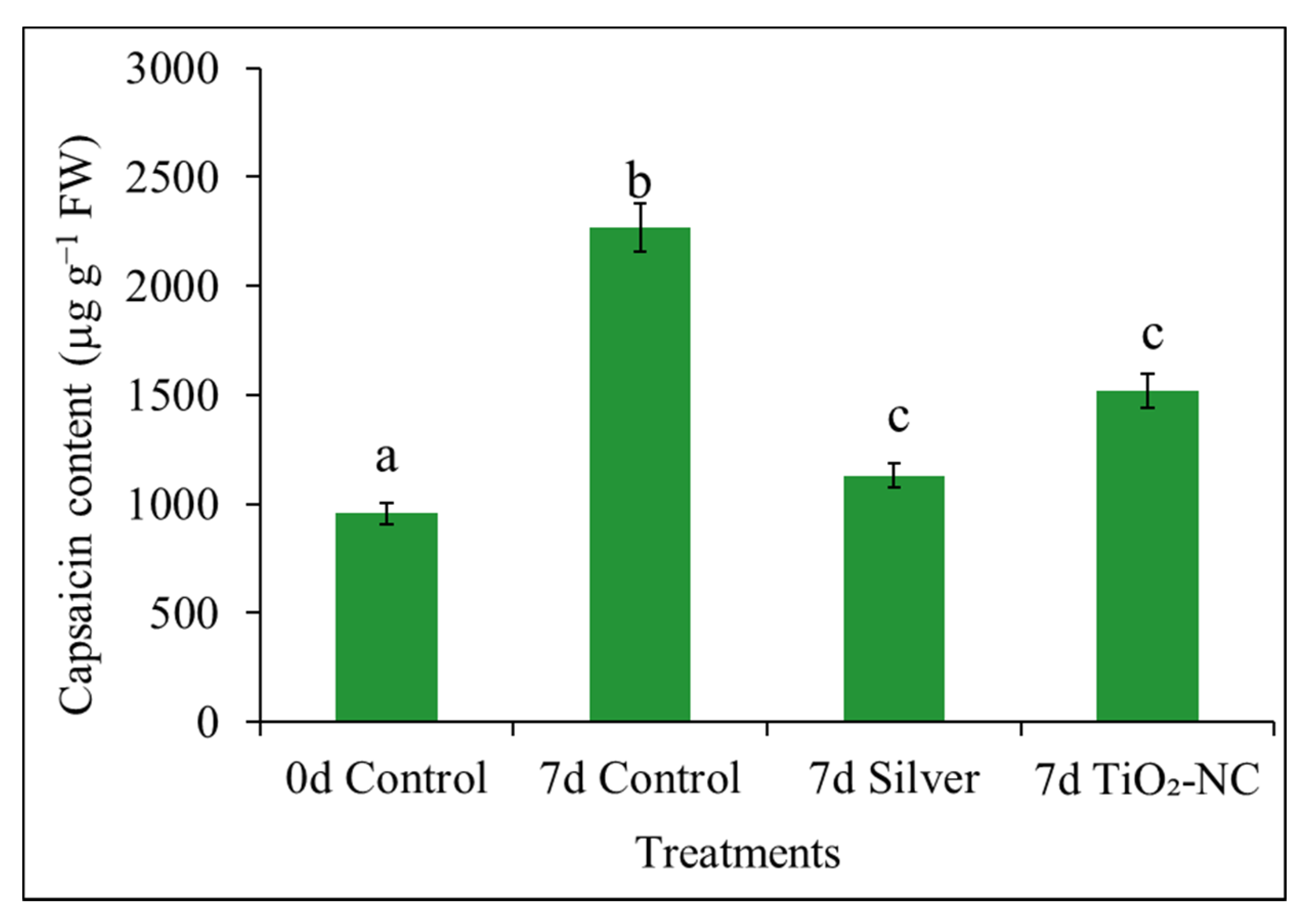
3.8. Capsaicin Content
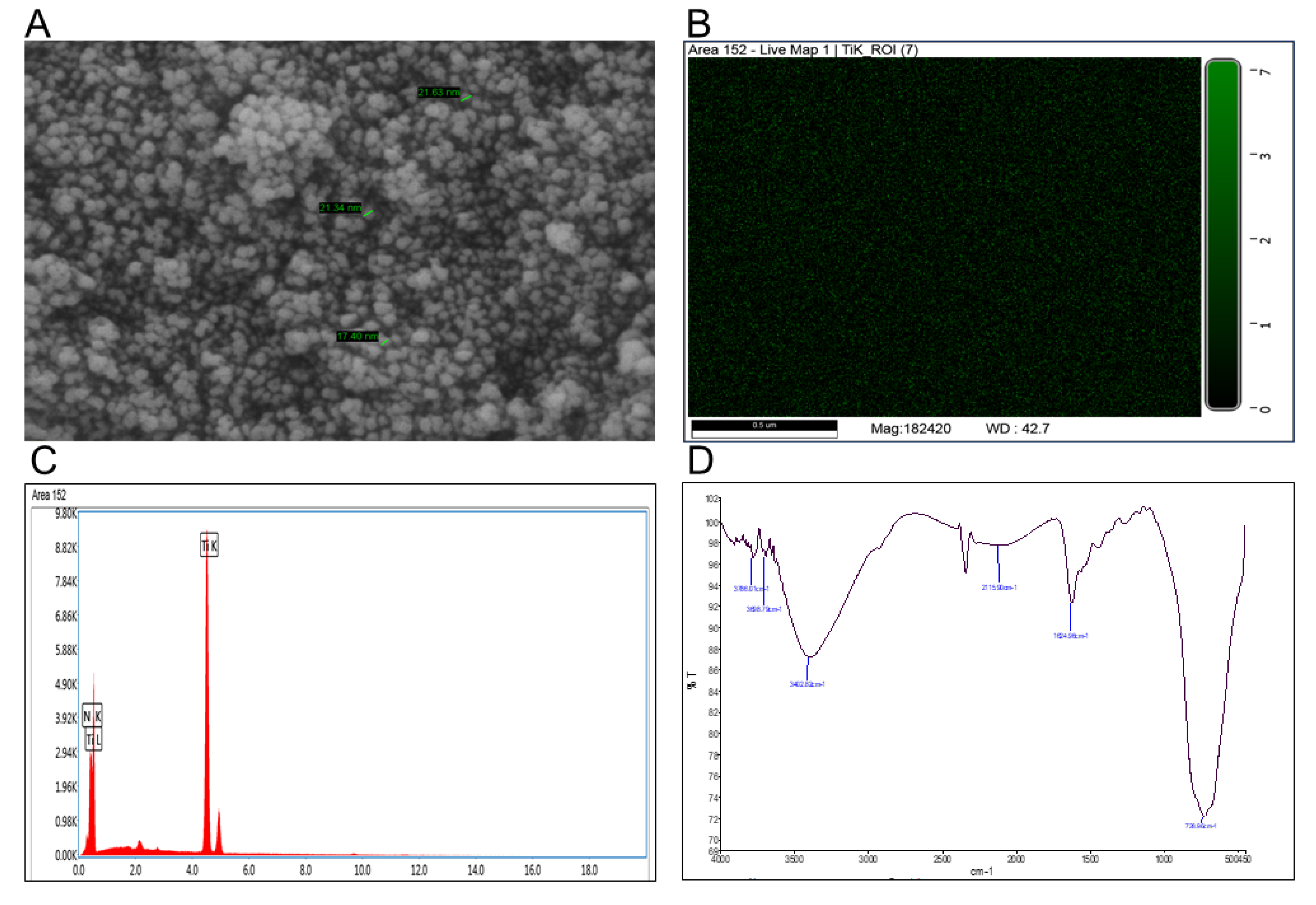
3.9. TiO2-NP and Its Opto-Physico-Chemical Characterizations
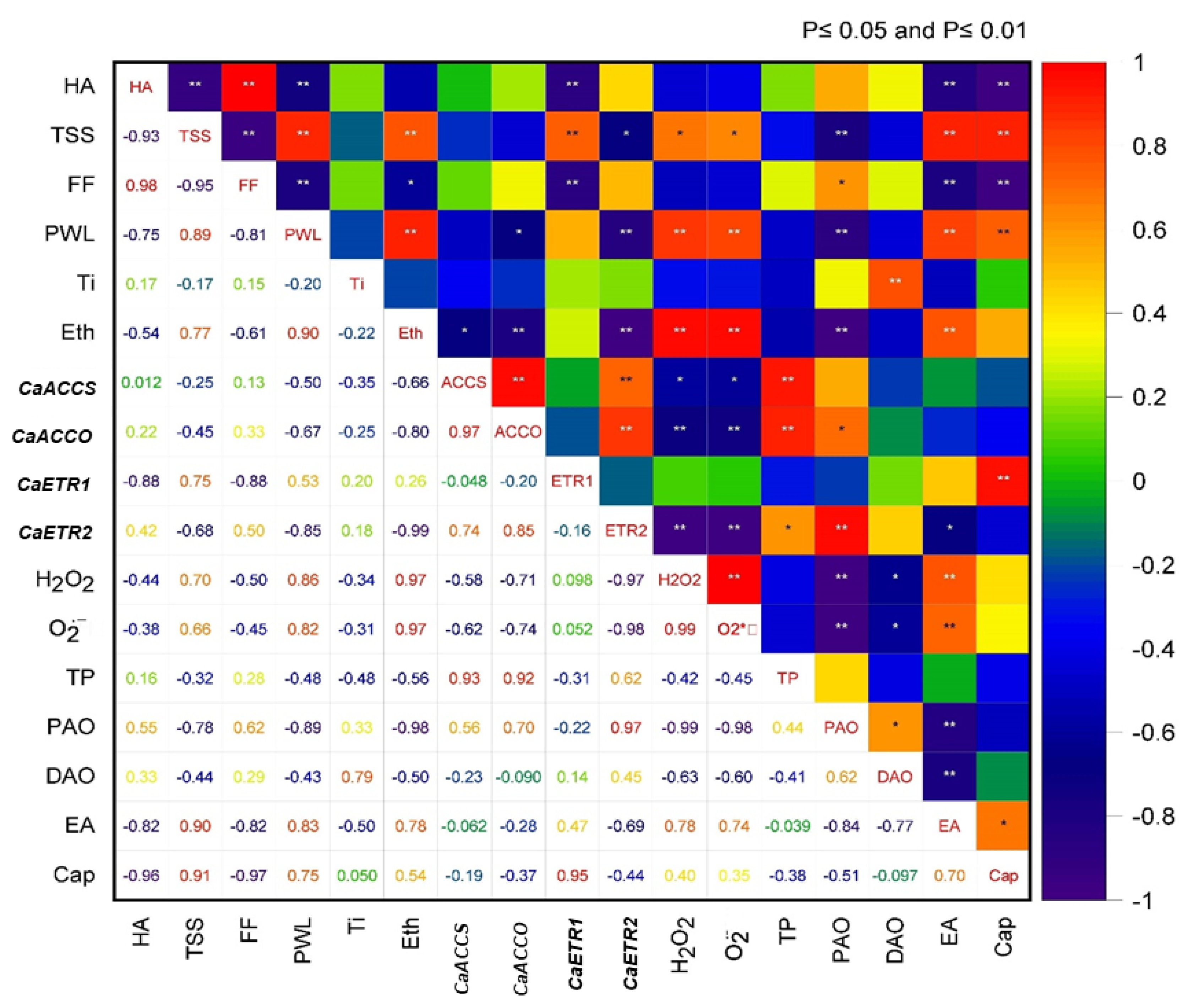
3.10. Correlation among the Parameters
4. Discussion
4.1. Modulation of Quality Characters for Ripening under TiO2 Nanocomposite
4.2. Transcript Dynamics of Ethylene Biosynthesis and Signaling Paths
4.3. ROS Metabolism Is Related to Polyamine Oxidation in Progress of Fruit Ripening at Storage
4.4. Metabolites Profiling: The Abundance of Essential Oils Induced by Nanocomposite
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fuentes, L.; Figueroa, C.R.; Valdenegro, M. Recent advances in hormonal regulation and cross-talk during non-climacteric fruit development and ripening. Horticulturae 2019, 5, 45. [Google Scholar] [CrossRef] [Green Version]
- Prasanna, V.; Prabha, T.N.; Tharanathan, R.N. Fruit ripening phenomena–An overview. Crit. Rev. Food Sci. Nutr. 2007, 47, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Grimplet, J.; David, K.; Castellarin, S.D.; Terol, J.; Wong, D.C.; Luo, Z.; Schaffer, R.; Celton, J.M.; Talon, M.; et al. Ethylene receptors and related proteins in climacteric and non-climacteric fruits. Plant Sci. 2018, 276, 63–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siebeneichler, T.J.; Crizel, R.L.; Camozatto, G.H.; Paim, B.T.; da Silva Messias, R.; Rombaldi, C.V.; Galli, V. The postharvest ripening of strawberry fruits induced by abscisic acid and sucrose differs from their in vivo ripening. Food Chem. 2020, 317, 126407. [Google Scholar] [CrossRef]
- Yan, S.C.; Chen, J.Y.; Yu, W.M.; Kuang, J.F.; Chen, W.X.; Li, X.P.; Lu, W.J. Expression of genes associated with ethylene-signalling pathway in harvested banana fruit in response to temperature and 1-MCP treatment. J. Sci. Food Agric. 2011, 91, 650–657. [Google Scholar] [CrossRef]
- Ghosh, A.; Saha, I.; Dolui, D.; De, A.K.; Sarkar, B.; Adak, M.K. Silver can induce oxidative stress in parallel to other chemical elicitors to modulate the ripening of chili cultivars. Plants 2020, 9, 238. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhu, X.; Mao, J.; Zou, Y.; Fu, D.; Chen, W.; Lu, W. Isolation and characterization of ethylene response factor family genes during development, ethylene regulation and stress treatments in papaya fruit. Plant Physiol. Biochem. 2013, 70, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Giné-Bordonaba, J.; Echeverria, G.; Duaigües, E.; Bobo, G.; Larrigaudière, C. A comprehensive study on the main physiological and biochemical changes occurring during growth and on-tree ripening of two apple varieties with different postharvest behaviour. Plant Physiol. Biochem. 2019, 135, 601–610. [Google Scholar] [CrossRef]
- Farcuh, M.; Rivero, R.M.; Sadka, A.; Blumwald, E. Ethylene regulation of sugar metabolism in climacteric and non-climacteric plums. Postharvest Biol. Technol. 2018, 139, 20–30. [Google Scholar] [CrossRef]
- Dubey, M.; Jaiswal, V.; Rawoof, A.; Kumar, A.; Nitin, M.; Chhapekar, S.S.; Kumar, N.; Ahmad, I.; Islam, K.; Brahma, V.; et al. Identification of genes involved in fruit development/ripening in Capsicum and development of functional markers. Genomics 2019, 111, 1913–1922. [Google Scholar] [CrossRef]
- Cisneros, L.; Gao, F.; Corma, A. Silver nanocluster in zeolites. Adsorption of ethylene traces for fruit preservation. Microporous Mesoporous Mater. 2019, 283, 25–30. [Google Scholar] [CrossRef]
- Strano, A.; Falcone, G.; Nicolò, B.F.; Stillitano, T.; De Luca, A.I.; Nesci, F.S.; Gulisano, G. Eco-profiles and economic performances of a high-value fruit crop in southern Italy: A case study of bergamot (Citrus bergamia Risso). Agroecol. Sustain. Food Syst. 2017, 41, 1124–1145. [Google Scholar]
- Ntsoane, M.L.; Jalali, A.; Römer, J.; Duewel, K.; Göller, C.; Kühn, R.; Mähne, K.; Geyer, M.; Sivakumar, D.; Mahajan, P.V. Performance evaluation of silica gel-water adsorption based cooling system for mango fruit storage in Sub-Saharan Africa. Postharvest Biol. Technol. 2019, 149, 195–199. [Google Scholar] [CrossRef]
- Bailén, G.; Guillén, F.; Castillo, S.; Serrano, M.; Valero, D.; Martínez-Romero, D. Use of activated carbon inside modified atmosphere packages to maintain tomato fruit quality during cold storage. J. Agric. Food Chem. 2006, 54, 2229–2235. [Google Scholar] [CrossRef] [PubMed]
- Pathak, N.; Caleb, O.J.; Geyer, M.; Herppich, W.B.; Rauh, C.; Mahajan, P.V. Photocatalytic and photochemical oxidation of ethylene: Potential for storage of fresh produce—A review. Food Bioproc. Technol. 2017, 10, 982–1001. [Google Scholar] [CrossRef]
- Geng, Q.; Lin, X.; Si, R.; Chen, X.; Dai, W.; Fu, X.; Wang, X. The correlation between the ethylene response and its oxidation over TiO2 under UV irradiation. Sens. Actuators B Chem. 2012, 174, 449–457. [Google Scholar] [CrossRef]
- Xing, Y.; Yang, H.; Guo, X.; Bi, X.; Liu, X.; Xu, Q.; Wang, Q.; Li, W.; Li, X.; Shui, Y.; et al. Effect of chitosan/Nano-TiO2 composite coatings on the postharvest quality and physicochemical characteristics of mango fruits. Sci. Hortic. 2020, 263, 109135. [Google Scholar] [CrossRef]
- Helal, M.; Sami, R.; Khojah, E.; Elhakem, A.; Benajiba, N.; Al-Mushhin, A.A.; Fouda, N. Evaluating the coating process of titanium dioxide nanoparticles and sodium tripolyphosphate on cucumbers under chilling condition to extend the shelf-life. Sci. Rep. 2021, 11, 20312. [Google Scholar] [CrossRef]
- de Matos Fonseca, J.; dos Santos Alves, M.J.; Soares, L.S.; Moreira, R.D.F.P.M.; Valencia, G.A.; Monteiro, A.R. A review on TiO2-based photocatalytic systems applied in fruit postharvest: Set-ups and perspectives. Food Res. Int. 2021, 144, 110378. [Google Scholar] [CrossRef]
- Wonglom, P.; Daengsuwan, W.; Ito, S.I.; Sunpapao, A. Biological control of Sclerotium fruit rot of snake fruit and stem rot of lettuce by Trichoderma sp. T76-12/2 and the mechanisms involved. Physiol. Mol. Plant Pathol. 2019, 107, 1–7. [Google Scholar] [CrossRef]
- Scalisi, A.; O’Connell, M.G. Application of Visible/NIR spectroscopy for the estimation of soluble solids, dry matter and flesh firmness in stone fruits. J. Sci. Food Agric. 2021, 101, 2100–2107. [Google Scholar] [CrossRef] [PubMed]
- Aizat, W.M.; Able, J.A.; Stangoulis, J.C.; Able, A.J. Characterisation of ethylene pathway components in non-climacteric capsicum. BMC Plant Biol. 2013, 13, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, A.; Saha, I.; Debnath, S.C.; Hasanuzzaman, M.; Adak, M.K. Chitosan and putrescine modulate reactive oxygen species metabolism and physiological responses during chili fruit ripening. Plant Physiol. Biochem. 2021, 163, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Sadasivam, S.; Manickam, A. Biochemical Methods, 3rd ed.; New Age International Publishers: New Delhi, India, 2008; pp. 209–210. [Google Scholar]
- Wettlaufer, S.H.; Weinstein, L.H. Quantitation of polyamines using thin-layer chromatography and image analysis. J. Chromatogr. A 1988, 441, 361–366. [Google Scholar] [CrossRef]
- Kumar, D.; Behal, S.; Bhattacharyya, R.; Banerjee, D. Fast blue B produces a light background on the gel surface. Indian J. Clin. Biochem. 2020, 16, 124–125. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Rocha, J.; Andrés, M.F.; Díaz, C.E.; Burillo, J.; González-Coloma, A. Composition and biocidal properties of essential oil from pre-domesticated Spanish Satureja Montana. Ind. Crops Prod. 2020, 145, 111958. [Google Scholar] [CrossRef]
- Femi-Adepoju, A.G.; Dada, A.O.; Otun, K.O.; Adepoju, A.O.; Fatoba, O.P. Green synthesis of silver nanoparticles using terrestrial fern (Gleichenia Pectinata (Willd.) C. Presl.): Characterization and antimicrobial studies. Heliyon 2019, 5, e01543. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Liu, S.; Cao, X.; Wu, S.; Liu, C.; Li, G.; Jiang, W.; Wang, H.; Wang, N.; Ding, W. Preparation and characterization of TiO2 nanoparticles by two different precipitation methods. Ceram. Int. 2020, 46, 15333–15341. [Google Scholar] [CrossRef]
- Bagheri, S.; Shameli, K.; Abd Hamid, S.B. Synthesis and characterization of anatase titanium dioxide nanoparticles using egg white solution via Sol-Gel method. J. Chem. 2013, 1, 848205. [Google Scholar] [CrossRef]
- Lyu, S.; Wei, X.; Chen, J.; Wang, C.; Wang, X.; Pan, D. Titanium as a beneficial element for crop production. Front. Plant Sci. 2017, 8, 597. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.; Zhou, Y.; Ma, C.; Feng, Y.; Hao, Y.; Rui, Y.; Wu, W.; Gui, X.; Han, Y.; Wang, Y.; et al. Jointed toxicity of TiO2 NPs and Cd to rice seedlings: NPs alleviated Cd toxicity and Cd promoted NPs uptake. Plant Physiol. Biochem. 2017, 110, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Jaimun, R.; Sangsuwan, J. Efficacy of chitosan-coated paper incorporated with vanillin and ethylene adsorbents on the control of anthracnose and the quality of Nam Dok Mai mango fruit. Packag. Technol. Sci. 2019, 32, 383–394. [Google Scholar] [CrossRef]
- Xing, Y.; Yang, S.; Xu, Q.; Xu, L.; Zhu, D.; Li, X.; Shui, Y.; Liu, X.; Bi, X. Effect of chitosan/nano-TiO2 composite coating on the postharvest quality of blueberry fruit. Coatings 2021, 11, 512. [Google Scholar] [CrossRef]
- Jang, S.; Park, M.; Lee, D.G.; Lim, J.H.; Jung, J.W.; Kang, B.C. Breeding Capsicum chinense lines with high levels of capsaicinoids and capsinoids in the fruit. Agriculture 2021, 11, 819. [Google Scholar] [CrossRef]
- Samira, A.; Woldetsadik, K.; Workneh, T.S. Postharvest quality and shelf life of some hot pepper varieties. J. Food Sci. Technol. 2013, 50, 842–855. [Google Scholar] [CrossRef] [Green Version]
- Zou, J.; Chen, J.; Tang, N.; Gao, Y.; Hong, M.; Wei, W.; Cao, H.; Jian, W.; Li, N.; Deng, W.; et al. Transcriptome analysis of aroma volatile metabolism change in tomato (Solanum lycopersicum) fruit under different storage temperatures and 1-MCP treatment. Postharvest Biol. Technol. 2018, 135, 57–67. [Google Scholar] [CrossRef]
- Steelheart, C.; Alegre, M.L.; Bahima, J.V.; Senn, M.E.; Simontacchi, M.; Bartoli, C.G.; Grozeff, G.E. Nitric oxide improves the effect of 1-methylcyclopropene extending the tomato (Lycopersicum esculentum L.) fruit postharvest life. Sci. Hortic. 2019, 255, 193–201. [Google Scholar] [CrossRef]
- Kumarihami, H.M.; Kim, J.G.; Kim, Y.H.; Lee, M.; Lee, Y.S.; Kwack, Y.B.; Kim, J. Preharvest application of chitosan improves the postharvest life of ‘Garmrok’ kiwifruit through the modulation of genes related to ethylene biosynthesis, cell wall modification, and lignin metabolism. Foods 2021, 10, 373. [Google Scholar] [CrossRef]
- Massolo, J.F.; Forte, L.G.; Concellón, A.; Viña, S.Z.; Vicente, A.R. Effects of ethylene and 1-MCP on quality maintenance of fresh cut celery. Postharvest Biol. Technol. 2019, 148, 176–183. [Google Scholar] [CrossRef]
- Li, S.; Chen, K.; Grierson, D. A critical evaluation of the role of ethylene and MADS transcription factors in the network controlling fleshy fruit ripening. New Phytol. 2019, 221, 1724–1741. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Ambrocio, A.; Guerrero-Beltrán, J.A.; Aparicio-Fernández, X.; Ávila-Sosa, R.; Hernández-Carranza, P.; Cid-Pérez, S.; Ochoa-Velasco, C.E. Effect of blue and ultraviolet-C light irradiation on bioactive compounds and antioxidant capacity of habanero pepper (Capsicum chinense) during refrigeration storage. Postharvest Biol. Technol. 2018, 135, 19–26. [Google Scholar] [CrossRef]
- Rodríguez-Ruiz, M.; González-Gordo, S.; Cañas, A.; Campos, M.J.; Paradela, A.; Corpas, F.J.; Palma, J.M. Sweet pepper (Capsicum annuum L.) fruits contain an atypical peroxisomal catalase that is modulated by reactive oxygen and nitrogen species. Antioxidants 2019, 8, 374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakya, R.; Singh, N.; Bhatla, S.C. Iron homeostasis regulates maturation of tomato (climacteric) and capsicum (non-climacteric) fruits. J. Plant Biochem. Biotechnol. 2021, 30, 392–395. [Google Scholar] [CrossRef]
- Sharma, S.; Pareek, S.; Sagar, N.A.; Valero, D.; Serrano, M. Modulatory effects of exogenously applied polyamines on postharvest physiology, antioxidant system and shelf life of fruits: A review. Int. J. Mol. Sci. 2017, 18, 1789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Rodríguez, E.; Romero, L.; Ruiz, J.M. Accumulation of free polyamines enhances the antioxidant response in fruits of grafted tomato plants under water stress. J. Plant Physiol. 2016, 190, 72–78. [Google Scholar] [CrossRef]
- Zamljen, T.; Zupanc, V.; Slatnar, A. Influence of irrigation on yield and primary and secondary metabolites in two chilies species, Capsicum annuum L. and Capsicum chinense Jacq. Agric. Water Manag. 2020, 234, 106104. [Google Scholar] [CrossRef]
- Romero, P.; Lafuente, M.T. Abscisic acid deficiency alters epicuticular wax metabolism and morphology that leads to increased cuticle permeability during sweet orange (Citrus sinensis) fruit ripening. Front. Plant Sci. 2020, 11, 1914. [Google Scholar] [CrossRef]
- Spadafora, N.D.; Cocetta, G.; Cavaiuolo, M.; Bulgari, R.; Dhorajiwala, R.; Ferrante, A.; Spinardi, A.; Rogers, H.J.; Müller, C.T. A complex interaction between pre-harvest and post-harvest factors determines fresh-cut melon quality and aroma. Sci. Rep. 2019, 9, 2745. [Google Scholar] [CrossRef]
- Taheri, A.; Behnamian, M.; Dezhsetan, S.; Karimirad, R. Shelf life extension of bell pepper by application of chitosan nanoparticles containing Heracleum persicum fruit essential oil. Postharvest Biol. Technol. 2020, 170, 111313. [Google Scholar] [CrossRef]
- Korkmaz, A.; Atasoy, A.F.; Hayaloglu, A.A. Changes in volatile compounds, sugars and organic acids of different spices of peppers (Capsicum annuum L.) during storage. Food Chem. 2020, 311, 125910. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Name of Compound | Calculated KI | Reported KI | Concentration (%) | ||
|---|---|---|---|---|---|---|
| 0 d Control | 7 d Control | TiO2-NC | ||||
| C1 | Cuben-11-ol | 1595 | 1595 | 34.02 | 4.12 | 17.66 |
| C2 | Spiroether | 1880 | 1879 | 14.22 | 1.78 | 7.94 |
| C3 | 1,3-dihydroxypropan-2-ylhexadecanoate | 2289 | 2289 | 3.15 | 2.79 | 3.02 |
| C4 | Methyl labdanolate | 2382 | 2381 | 2.83 | 9.36 | 9.11 |
| C5 | Tetracosane | 2388 | 2400 | 4.67 | 5.39 | 5.40 |
| C6 | Pentacosane | 2488 | 2500 | 4.74 | 8.32 | 4.88 |
| C7 | Hexacosane | 2588 | 2600 | 4.36 | 11.30 | 5.50 |
| C8 | Nonacosanoicacid | 2673 | 2674 | 3.95 | 3.08 | 5.65 |
| C9 | Heptacosane | 2745 | 2700 | 3.41 | 13.79 | 4.90 |
| C10 | Pentatriacontane-2-one | 2873 | 2873 | 2.85 | 0.40 | 4.31 |
| C11 | Triacontane | 2985 | 3000 | 2.50 | 9.31 | 3.10 |
| C12 | Hentriacontane | 3084 | 3100 | 2.55 | 6.26 | 2.46 |
| C13 | Dotriacontane | 3183 | 3200 | 1.84 | 4.02 | 2.02 |
| Total percentage | 85.10 | 79.93 | 75.94 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghosh, A.; Saha, I.; Fujita, M.; Debnath, S.C.; Hazra, A.K.; Adak, M.K.; Hasanuzzaman, M. Photoactivated TiO2 Nanocomposite Delays the Postharvest Ripening Phenomenon through Ethylene Metabolism and Related Physiological Changes in Capsicum Fruit. Plants 2022, 11, 513. https://doi.org/10.3390/plants11040513
Ghosh A, Saha I, Fujita M, Debnath SC, Hazra AK, Adak MK, Hasanuzzaman M. Photoactivated TiO2 Nanocomposite Delays the Postharvest Ripening Phenomenon through Ethylene Metabolism and Related Physiological Changes in Capsicum Fruit. Plants. 2022; 11(4):513. https://doi.org/10.3390/plants11040513
Chicago/Turabian StyleGhosh, Arijit, Indraneel Saha, Masayuki Fujita, Subhas Chandra Debnath, Alok Kumar Hazra, Malay Kumar Adak, and Mirza Hasanuzzaman. 2022. "Photoactivated TiO2 Nanocomposite Delays the Postharvest Ripening Phenomenon through Ethylene Metabolism and Related Physiological Changes in Capsicum Fruit" Plants 11, no. 4: 513. https://doi.org/10.3390/plants11040513