
Taxonomic Revision of Hook-Forming Acrosorium (Delesseriaceae, Rhodophyta) from the Northwestern Pacific Based on Morphology and Molecular Data


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
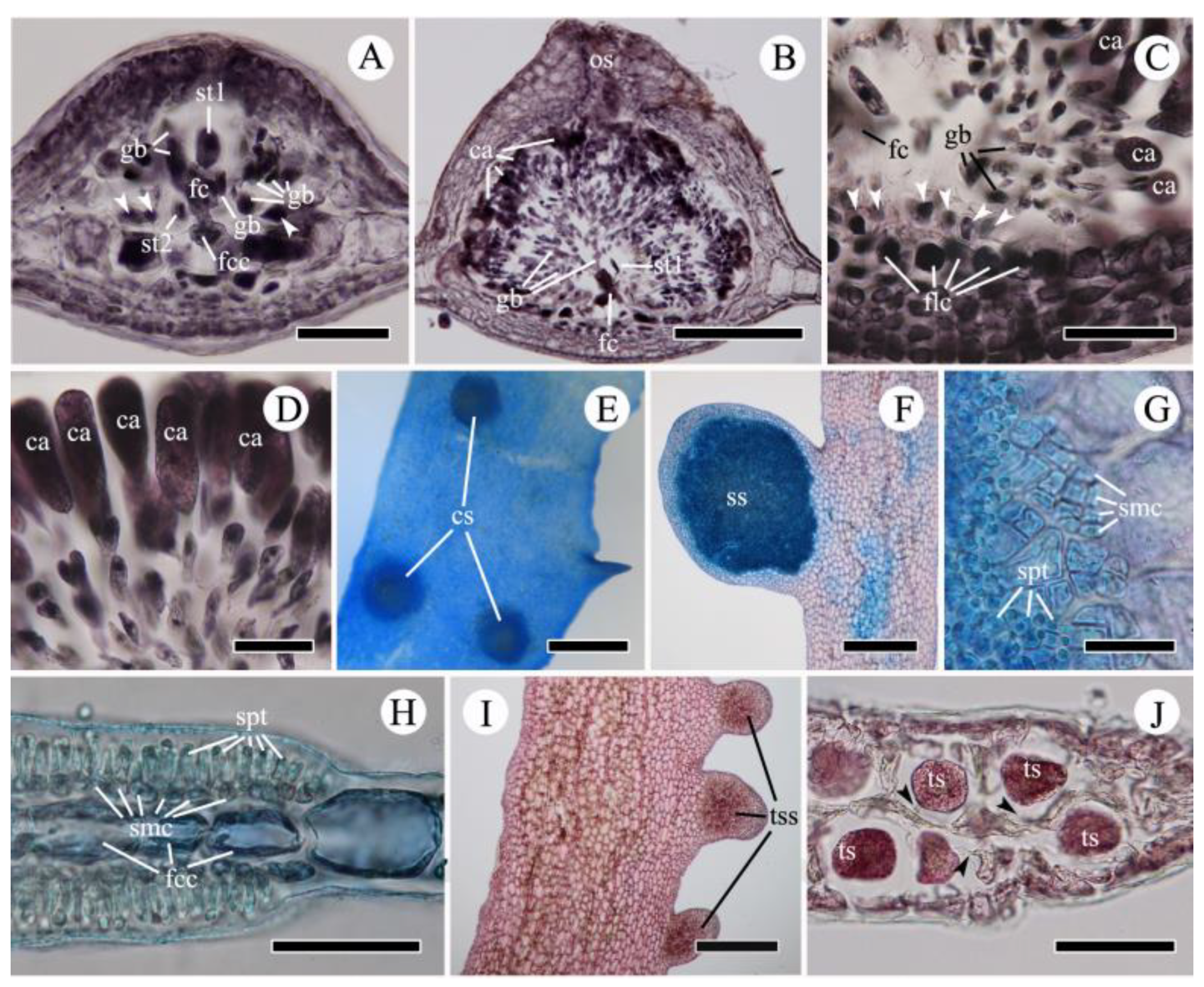{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
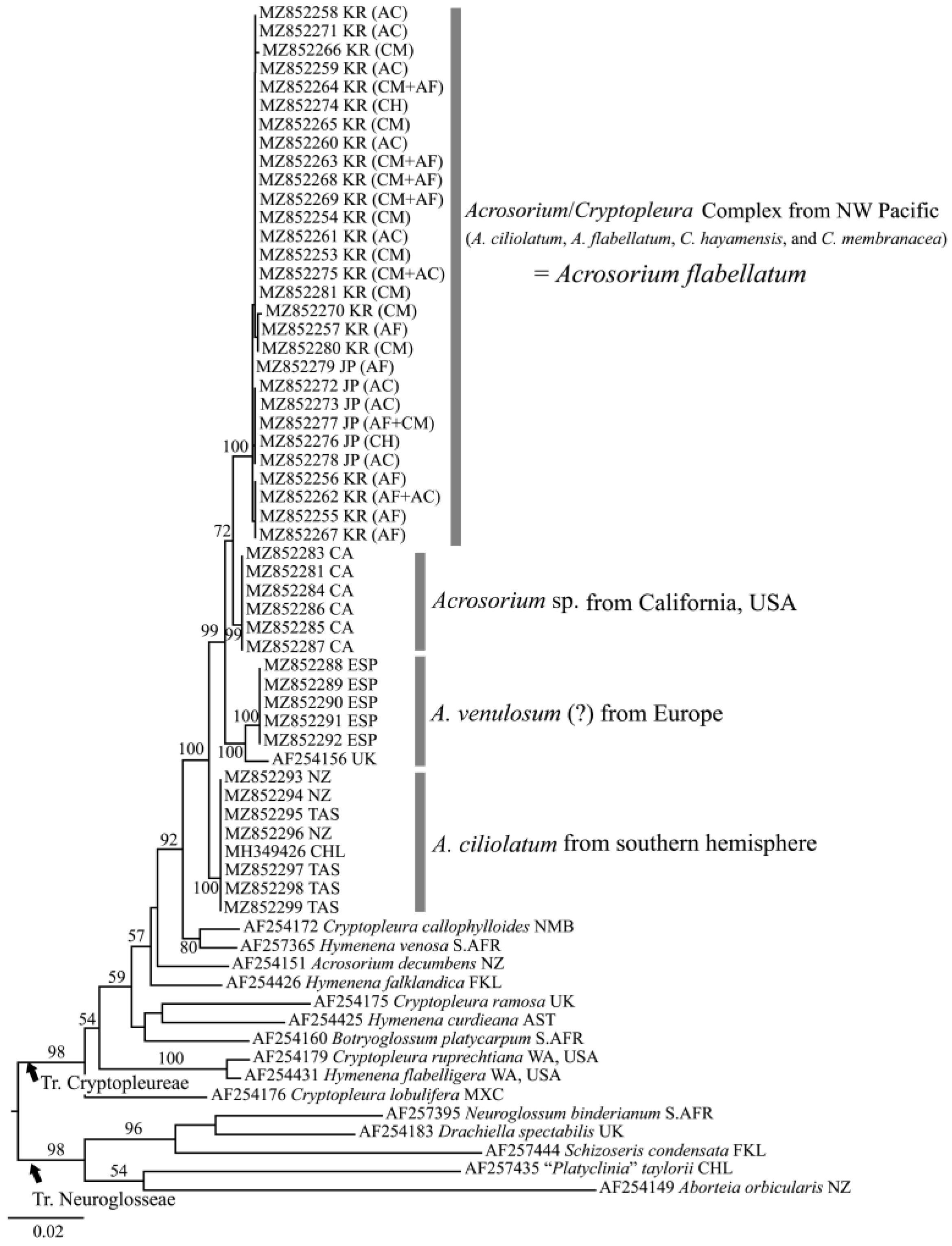
2.1. Molecular Analyses
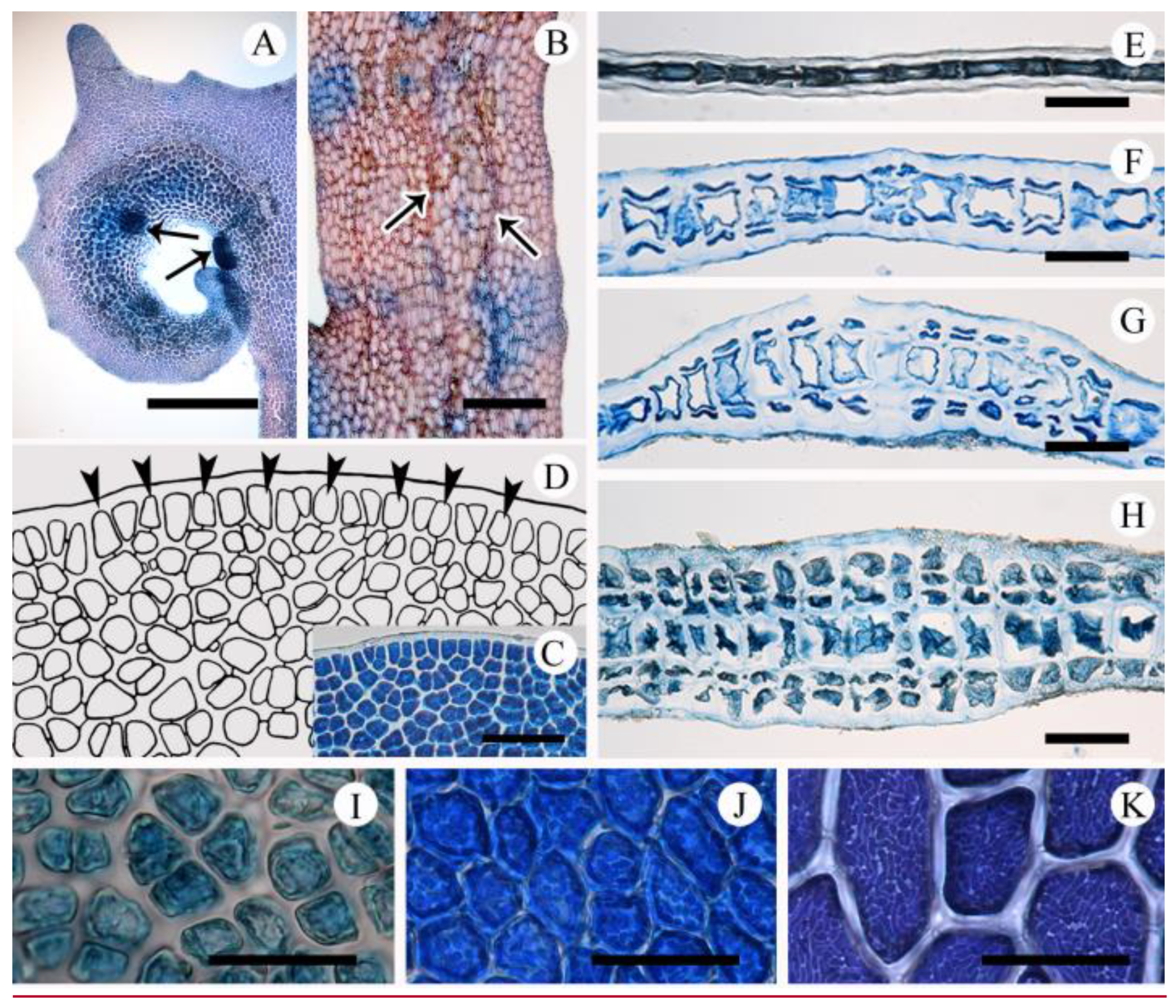
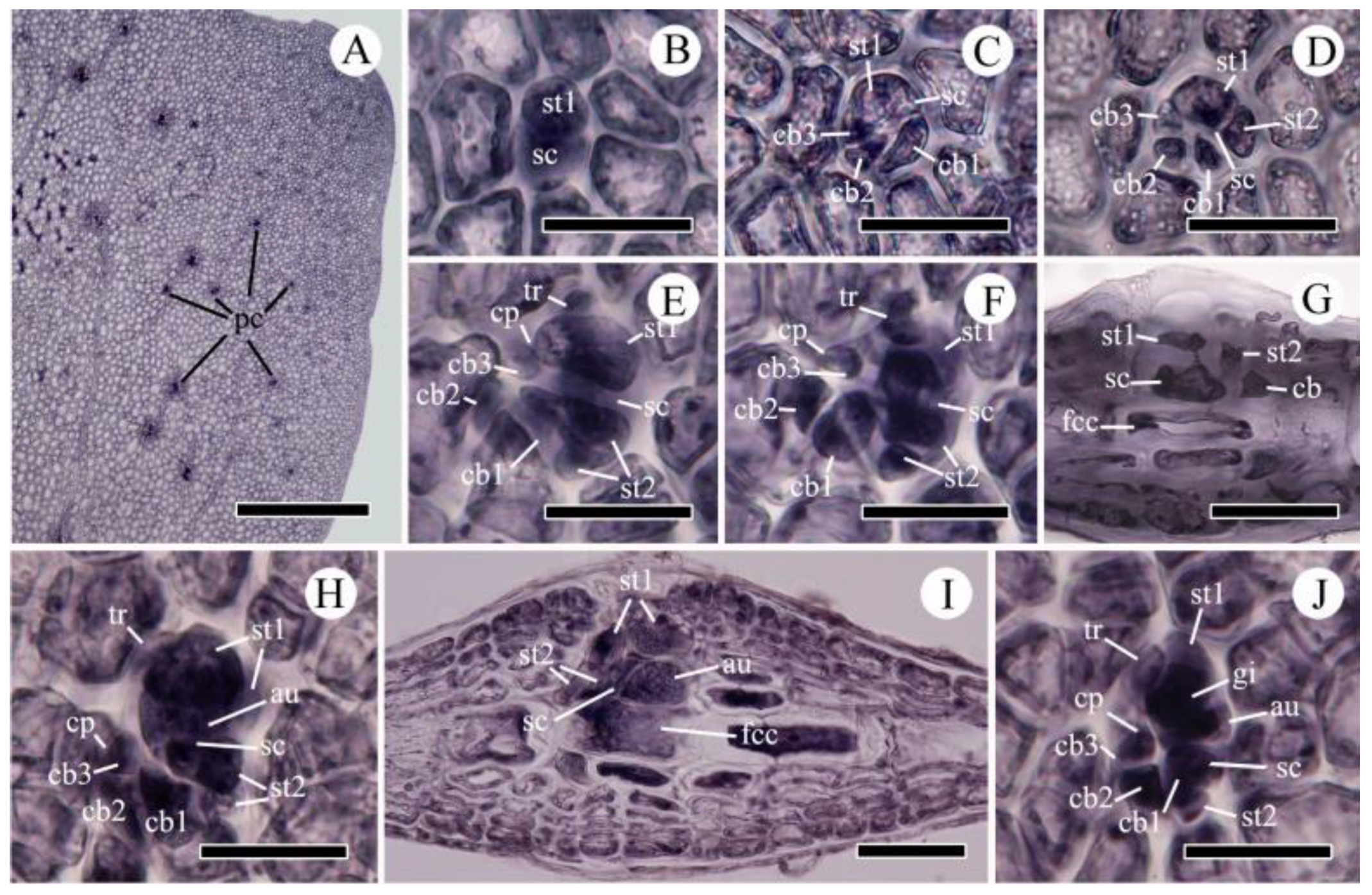
2.2. Morphological Observations
2.2.1. Acrosorium flabellatum Yamada 1930:31
2.2.2. Acrosorium ciliolatum from Beyond the NW Pacific
- Specimens examined (Figure 7)
- Habit (Figure 7)
3. Discussion
4. Taxonomic Conclusions
5. Materials and Methods
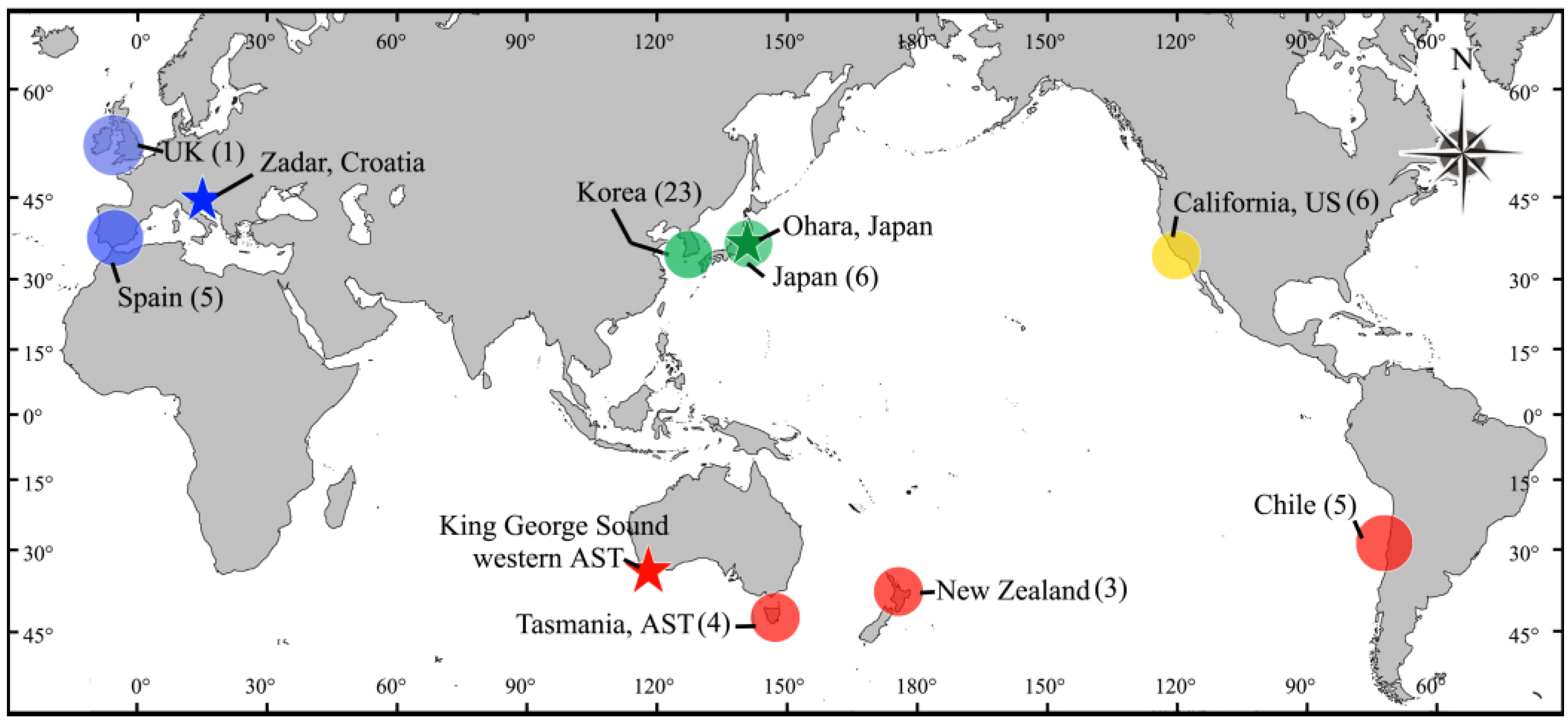
5.1. Collections
5.2. Classifying Specimens with Phenotype
5.3. Molecular Analysis
5.4. Morphological Observation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kützing, F.T. Tabulae phycologicae; oder, Abbildungen der Tange; Gedruckt auf Kosten des Verfassers: Nordhausen, Germany, 1869; Volume 19, p. 4. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. 2021. Available online: http://www.algaebase (accessed on 12 August 2021).
- Kylin, H. Studien über die Delesseriaceen. Lunds Univ. Årsskr. N.F. Avd. 1924, 20, 5–111. [Google Scholar]
- Maggs, C.A.; Hommersand, M.H. Seaweeds of the British Isles. Vol. 1. Rhodophyta. Part 3A. Ceramiales; HMSO Books: London, UK, 1993; pp. 245–257. [Google Scholar]
- Womersley, H.B.S. The Marine Benthic Flora of Southern Australia. Rhodophyta. Part IIID: Ceramiales–Delesseriaceae, Sarcomeniaceae, Rhodomelaceae; Australian Biological Resources Study, Canberra and the State Herbarium of South Australia: Adelaide, Australia, 2003; pp. 129–133.
- Wynne, M.J. Towards the resolution of taxonomic and nomenclatural problems concerning the typification of Acrosorium uncinatum (Delesseriaceae: Rhodophyta). Brit. Phycol. J. 1989, 24, 245–252. [Google Scholar] [CrossRef]
- Harvey, W.H. Some account of the marine botany of the colony of western Australia. T. Roy. Irish Acad. 1855, 22, 525–566. [Google Scholar]
- Zanardini, G. Scelta di Ficee nuove o più rare dei mari Mediterraneo ed Adriatico. [Decade VII]. Mem. R. Ist. Veneto Sci. Lett. Arti 1866, 13, 141–176, pls. XLIX−LVI. [Google Scholar]
- Turner, D. Fuci Sive Plantarum Fucorum Generi a Botanicis Ascriptarum Icones Descriptiones et Historia.; Typis J. M’Creery, impensis J. et A. Arch in vico Cornhill dicto: London, UK, 1808; Volume 1, pp. 151–152, pl. 8. [Google Scholar]
- Newton, L. A Handbook of the British Seaweeds; The Trustees of the British Museum (Natural History): London, UK, 1931; p. 332. [Google Scholar]
- Howe, M.A.; Taylor, W.R. Notes on new or little-known marine algae from Brazil. Brittonia 1931, 1, 7–33. [Google Scholar] [CrossRef]
- Jaasund, E. Marine algae in Tanzania, V. Bot. Mar. 1977, 20, 333–338. [Google Scholar] [CrossRef]
- Papenfuss, G.F. The development of the reproductive organs of Acrosorium acrospermum. Bot. Notiser. 1939, 1939, 11–20. [Google Scholar]
- Papenfuss, G.F. Notes on South African marine algae. I. Bot. Notiser. 1940, 200–226, 16 figs. [Google Scholar]
- Papenfuss, G.F. Notes on South African marine algae. II. J. S. Afr. Bot. 1943, 9, 79−92. [Google Scholar]
- Taylor, W.R. Pacific Marine algae of the Allan Hancock Expeditions to the Galapagos Islands. Allan Hancock Pacific expeditions Vol. 12; The University of Southern California Press: Los Angeles, CA, USA, 1945; pp. 284–286, pl. 95. [Google Scholar]
- Wynne, M.J. The Red Algal Families Delesseriaceae and Sarcomeniaceae; Koeltz Scientific Books: Königstein. Germany, 2014; pp. 157–160. [Google Scholar]
- Okamura, K. Nippon Sorui-Meii [Book Listing Japanese Algae]; Keigyosha: Tokyo, Japan, 1902; p. 49. (In Japanese) [Google Scholar]
- Boo, S.M.; Ko, Y.D. Marine Plants from Korea; Marine & Extreme Genome Research Centre Program: Seoul, Korea, 2012; p. 132. (In Korean) [Google Scholar]
- Nam, K.W.; Kang, P.J. Algal Flora of Korea. Volume 4, Number 7 Rhodophyta: Florideophyceae: Ceramiales: Delesseriaceae: 22 Genera including Acrosorium; NIBR: Incheon, Korea, 2012; pp. 57–79. [Google Scholar]
- Titlyanov, E.A.; Titlyanov, T.A.; Xia, B.M.; Bartsch, I. Retrospective analysis of diversity and species composition of marine macroalgae on Hainan Island (China). Ocean Sci. J. 2016, 51, 1–22. [Google Scholar] [CrossRef]
- Tseng, C.K. Seaweeds in Yellow Sea and Bohai Sea of China; Science Press: Beijing, China, 2009; p. 453. (In Chinese) [Google Scholar]
- Yoshida, T. Marine Algae of Japan. Uchida Rokakuho Publishing Co., Ltd.: Tokyo, Japan, 1998; pp. 955–1001. (In Japanese) [Google Scholar]
- Yoshida, T.; Suzuki, M.; Yoshinaga, K. Checklist of marine algae of Japan (Revised in 2015). Jpn. J. Phycol. 2015, 63, 129–189. (In Japanese) [Google Scholar]
- Yamada, Y. Notes on some Japanese algae, I. J. Fac. Sci. Hokkaido Imper. Univ. 1930, 1, 27–36. Available online: http://hdl.handle.net/2115/26201 (accessed on 20 July 2021).
- Yamada, Y. Notes on some Japanese algae, VI. Sci. Pap. Inst. Algol. Res. Fac. Sci. Hokkaido Imper. Univ. 1935, 1, 27–35. Available online: http://hdl.handle.net/2115/48045 (accessed on 20 July 2021).
- Okamura, K. Nippon kaisô shi [Descriptions of Japanese Algae]; Uchida Rokakuho: Tokyo, Japan, 1936; pp. 755–795. (In Japanese) [Google Scholar]
- Yamada, Y. Notes on some Japanese algae IX. Sci. Pap. Inst. Algol. Res. Fac. Sci. Hokkaido Imper. Univ. 1941, 2, 195–215. Available online: http://hdl.handle.net/2115/48066 (accessed on 20 July 2021).
- Noda, M.; Kitami, T. Some species of marine algae from Echigo Province facing the Japan Sea. Sci. Rep. Niigata Univ. Ser. D 1971, 8, 35–52. [Google Scholar]
- Okamura, K. Icones of Japanese Algae; Kazamashobo: Tokyo, Japan, 1908; Volume I, pp. 121–232. [Google Scholar]
- Kang, J.W. Illustrated Encyclopedia of Fauna and Flora of Korea. Vol. 8. Marine Algae; Ministry of Education: Seoul, Korea, 1968; pp. 310–323. (In Korean)
- Kylin, H. Die Delesseriaceen Neu-seelands. Lunds Univ. Arsskr. N. F. Avd. 1929, 2, 10–14. [Google Scholar]
- Gavio, B.; Fredericq, S. Grateloupia turuturu (Halymeniaceae, Rhodophyta) is the correct name of the non-native species in the Atlantic known as Grateloupia doryphora. Eur. J. Phycol. 2002, 37, 349–360. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y. The genus Martensia Hering (Delesseriaceae, Rhodophyta) with M. albida sp. nov. and M. flammifolia sp. nov. on Jeju Island, Korea. Algae 2006, 21, 15–48. [Google Scholar] [CrossRef]
- Yang, M.Y.; Macaya, E.C.; Kim, M.S. Molecular evidence for verifying the distribution of Chondracanthus chamissoi and C. teedei (Gigartinaceae, Rhodophyta). Bot. Mar. 2015, 58, 103–113. [Google Scholar] [CrossRef]
- Couceiro, L.; Cremades, J.; Barreiro, R. Evidence for multiple introductions of the Pacific green alga Ulva australis Areschoug (Ulvales, Chlorophyta) to the Iberian Peninsula. Bot. Mar. 2011, 54, 391–402. [Google Scholar] [CrossRef]
- Kang, J.C.; Yang, M.Y.; Lin, S.-M.; Kim, M.S. Reappraisal of nine species of Martensia (Delesseriaceae, Rhodophyta) reported from Korea based on morphology and molecular analyses. Bot. Mar. 2015, 58, 151–166. [Google Scholar] [CrossRef]
- Kim, S.Y.; Han, E.G.; Kim, M.S.; Park, J.K.; Boo, S.M. Grateloupia jejuensis (Halymeniales, Rhodophyta): A new species previously confused with G. elata and G. cornea in Korea. Algae 2013, 28, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.Y.; Kim, M.S. DNA barcoding of the funoran-producing red algal genus Gloiopeltis (Gigartinales) and description of a new species, Gloiopeltis frutex sp. nov. J. Appl. Phycol. 2017, 30, 1381–1392. [Google Scholar] [CrossRef]
- Sherwood, A.R.; Lin, S.-M.; Wade, R.M.; Spalding, H.L.; Smith, C.M.; Kosaki, R.K. Characterization of Martensia (Delesseriaceae; Rhodophyta) from shallow and mesophotic habitats in the Hawaiian Islands: Description of four new species. Eur. J. Phycol. 2019, 55, 172–185. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.-M.; Hommersand, M.H.; Nelson, W.A. An assessment of Haraldiophyllum (Delesseriaceae, Rhodophyta), including H. crispatum (J.D. Hooker et Harvey) comb. nov. from New Zealand based on rbcL and LSU sequence analysis and morphological evidence. Eur. J. Phycol. 2007, 42, 391–408. [Google Scholar] [CrossRef]
- Kang, J.C.; Yang, M.Y.; Kim, M.S. Neoharaldiophyllum a new genus of Delesseriaceae (Rhodophyta) based on carposporophyte development and molecular data. Bot. Mar. 2017, 60, 515–532. [Google Scholar] [CrossRef]
- Yang, M.Y.; Kim, M.S. Molecular phylogeny of the genus Chondracanthus (Rhodophyta), focusing on the resurrection of C. okamurae and the description of C. cincinnus sp. nov. Ocean Sci. J. 2016, 51, 517–529. [Google Scholar] [CrossRef]
- Lin, S.-M.; Nelson, W.A.; Hommersand, M.W. Hymenenopsis heterophylla gen. et sp. nov. (Delesseriaceae, Rhodophyta) from New Zealand, based on a red alga previously known as Hymenena palmata f. marginata sensu Kylin, with emphasis on its cystocarp development. Phycologia 2012, 51, 62–73. [Google Scholar] [CrossRef]
- Kang, J.C.; Kim, M.S. Transfer of Papenfussia japonica (Delesseriaceae, Rhodophyta) from Korea to the genus Augophyllum. Bot. Mar. 2016, 59, 405–415. [Google Scholar] [CrossRef]
- Lin, S.-M.; Fredericq, S.; Hommersand, M.H. Apoglosseae trib. nov. (Delesseriaceae, Rhodophyta) based on two molecular datasets and cystocarp development. J. Phycol. 2002, 38, 23–24. [Google Scholar] [CrossRef]
- Kang, J.C.; Kim, M.S. A novel species Symphyocladia glabra sp. nov. (Rhodomelaceae, Rhodophyta) from Korea based on morphological and molecular analyses. Algae 2013, 28, 149–160. [Google Scholar] [CrossRef] [Green Version]
- Mikami, H. On Acrosorium uncinatum (Turner) Kylin (Rhodophyceae, Delesseriaceae) from Japan. Jap. J. Phycol. 1980, 28, 113–116. (In Japanese) [Google Scholar]
- Abbott, I.A.; Hollenberg, G.J. Marine Algae of California; Stanford University Press: Stanford, CA, USA, 1976; Volume i−xii, pp. 1–827, 701 figs. [Google Scholar]
- Díaz-Tapia, P.; Boo, S.M.; Geraldino, P.J.L.; Maneiro, I.; Bárbara, I.; Hommersand, M.H. Morphology and systematics of Calliblepharis hypneoides, sp. nov. (Cystocloniaceae, Rhodophyta) from the Atlantic Iberian Peninsula. Eur. J. Phycol. 2013, 48, 380–397. [Google Scholar] [CrossRef]
- Gargiulo, G.M.; Morabito, M.; Manghisi, A. A re-assessment of reproductive anatomy and postfertilization development in the systematics of Grateloupia (Halymeniales, Rhodophyta). Cryptogamie Algol. 2013, 34, 3–35. [Google Scholar] [CrossRef]
- Lee, H.W.; Kim, M.S. Female reproductive structures define the novel genus, Nesoia (Halymeniaceae, Rhodophyta). Eur. J. Phycol. 2019, 54, 66–77. [Google Scholar] [CrossRef]
- Lee, Y.; Kang, S. A catalogue of the Seaweeds in Korea; Cheju National University Press: Jeju, Korea, 2001; Volume 8, pp. 1–662. (In Korean) [Google Scholar]
- Lee, Y. Marine algae of Jeju; Academy Publication: Seoul, Korea, 2008; Volume i−xvi, pp. 1–477. (In Korean) [Google Scholar]
- Agardh, J.G. Species Genera et Ordines Algarum, seu Descriptiones Succinctae Specierum, Generum et Ordinum, Quibus Algarum Regnum Constituitur; C.W.K. Gleerup: Lund, Sweden, 1852; Volume 2, pp. 577–700. [Google Scholar]
- Kim, M.S.; Kim, S.Y.; Nelson, W. Symphocladia lithophila sp. nov. (Rhodomelaceae, Ceramiales), a new Korean red algal species based on morphology and rbcL sequences. Bot. Mar. 2010, 53, 233–241. [Google Scholar] [CrossRef]
- Saunders, G.W.; Moore, T.E. Refinements for the amplification and sequencing of red algal DNA barcode and RedToL phylogenetic markers: A summary of current primers, profiles, and strategies. Algae 2013, 28, 31–43. [Google Scholar] [CrossRef] [Green Version]
- Chun, J. PHYDIT Version 3.1. 2001. Available online: http://plaza.snu.ac.kr/~jchun/phydit/ (accessed on 23 August 2012).
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, W. Aceto-iron-haematoxylin-chloral hydrate for chromosome staining. Stain Technol. 1965, 40, 161–164. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, J.C.; Lin, S.-M.; Miller, K.A.; Kim, M.S. Taxonomic Revision of Hook-Forming Acrosorium (Delesseriaceae, Rhodophyta) from the Northwestern Pacific Based on Morphology and Molecular Data. Plants 2021, 10, 2269. https://doi.org/10.3390/plants10112269
Kang JC, Lin S-M, Miller KA, Kim MS. Taxonomic Revision of Hook-Forming Acrosorium (Delesseriaceae, Rhodophyta) from the Northwestern Pacific Based on Morphology and Molecular Data. Plants. 2021; 10(11):2269. https://doi.org/10.3390/plants10112269
Chicago/Turabian StyleKang, Jeong Chan, Showe-Mei Lin, Kathy Ann Miller, and Myung Sook Kim. 2021. "Taxonomic Revision of Hook-Forming Acrosorium (Delesseriaceae, Rhodophyta) from the Northwestern Pacific Based on Morphology and Molecular Data" Plants 10, no. 11: 2269. https://doi.org/10.3390/plants10112269