
Biochemical and Molecular Characterization of the Rice Chalcone Isomerase Family


Department of Genetics and Biotechnology, Kyung Hee University, Yongin 17104, Korea
*
Authors to whom correspondence should be addressed.
†
These authors equally contributed to this work.
Plants 2021, 10(10), 2064; https://doi.org/10.3390/plants10102064
Submission received: 27 August 2021
/
Revised: 24 September 2021
/
Accepted: 28 September 2021
/
Published: 30 September 2021
(This article belongs to the Special Issue Rice Genetics and Breeding)
Abstract
:Chalcone isomerase (CHI) is a key enzyme in flavonoid biosynthesis. In plants, CHIs occur in multigene families, and they are divided into four types, types I–IV. Type I and II CHIs are bona fide CHIs with CHI activity, and type III and IV CHIs are non-catalytic members with different functions. Rice contains seven CHI family genes (OsCHIs). Molecular analysis suggested that OsCHI3 is a type I CHI, and the other OsCHIs were classified into types III and IV. To elucidate their biochemical functions, OsCHI1, OsCHI3, OsCHI6, and OsCHI7 were expressed in Escherichia coli, and the recombinant OsCHI proteins were purified. An activity assay of recombinant OsCHIs showed that OsCHI3 catalyzed the isomerization of naringenin chalcone and isoliquiritigenin, whereas the other recombinant OsCHIs had no CHI activity. OsCHI3 also exhibited a strong preference to naringenin chalcone compared to isoliquiritigenin, which agrees well with the catalytic properties of type I CHIs. These results ascertain OsCHI3 to be a bona fide CHI in rice. OsCHI3 and the other OsCHIs were expressed constitutively throughout the rice growth period and different tissues. OsCHI3 expression was induced immediately in response to ultra-violet (UV) stress, suggesting its involvement in the biosynthesis of sakuranetin, a flavonoid phytoalexin in rice.
1. Introduction
Flavonoids are a major group of plant secondary metabolites that play a role in pigmentation, development, and reproduction as well as participate in defense against biotic and abiotic stresses as UV-protectants, antioxidants, and antimicrobial agents [1,2,3]. They are also health-beneficial phytonutrients in the human diet, with diverse biological activities, including hepatoprotective, anti-inflammatory, antibacterial, and anticancer properties [4,5].
Flavonoid biosynthesis is branched from the general phenylpropanoid pathway [1,2]. Chalcone synthase (CHS) is the first committed enzyme for flavonoid biosynthesis and catalyzes the formation of chalcones from one p-coumaroyl-CoA and three malonyl-CoAs [6,7,8,9]. Chalcone isomerase (CHI) sequentially catalyzes the stereospecific isomerization of chalcones into the corresponding (2S)-flavanones, providing basic backbones for a wide array of flavonoids [10,11]. In plants, CHI genes exist in multigene families and are divided into four types, types I–IV [12,13,14,15]. Types I and II CHIs are bona fide CHIs, having catalytic activities, and type III and IV CHIs are non-catalytic members with different physiological functions in plants [11,12,13,16,17,18,19]. In most plant species, CHS catalyzes the formation of 4,2′,4′,6′-tetrahydroxychalcone (naringenin chalcone) from p-coumaroyl-CoA and malonyl-CoAs. Naringenin chalcone is then isomerized to naringenin (5,7,4′-trihydroxyflavanone) by type I CHI, providing a basic backbone for general flavonoids [12,15]. In legumes, 4,2′,4′-trihydroxychalcone (isoliquiritigenin) is formed by CHS in conjunction with chalcone reductase [20,21]. Type II CHIs then isomerize isoliquiritigenin to liquiritigenin (7,4′-dihyrroxyflavanone), leading to isoflavonoid biosynthesis [11,12,14]. Isoflavonoids play an important role in legumes as phytoalexins, antimicrobial secondary metabolites produced in plants in response to pathogen attacks [3].
Rice produces a diverse variety of diterpenoid and phenolic phytoalexins in response to pathogen infections and UV stress [9,22,23,24,25,26,27]. Sakuranetin is a well-known rice phytoalexin and is a 7-methylated form of the flavanone naringenin, which is isolated from UV-treated rice leaves and exhibits antimicrobial activity against rice blast fungus (Magnaporthe grisea) [23]. Further research demonstrated that sakuranetin also has antimicrobial activity against several fungal and bacterial pathogens in rice, such as brown spot fungus (Bipolaris oryzae), grain rot bacterium (Burkholderia glumae), and blight bacterium (Xanthomonas oryzae pv. oryzae) [23,26]. Moreover, phenylpropanoid pathway and CHS genes are upregulated by UV irradiation and participate in sakuranetin synthesis under UV stress [9,25,27].
The rice genome contains seven CHI family members (OsCHIs). To delineate their physiological roles, we carried out molecular and biochemical characterization of the OsCHI family. The CHI activity assay showed that OsCHI3 is the only catalytic CHI, and the other OsCHIs are non-catalytic. Such catalytic activity and phylogenetic relationships indicate that OsCHI3 is a bona fide type I CHI in rice. In addition, the UV-induced expression of OsCHI3 suggested its involvement in sakuranetin biosynthesis. Expression patterns of six non-catalytic OsCHI family members in different tissues and developmental stages were examined, and their possible role was discussed.
2. Results
2.1. The CHI Family in Rice
A search of the MSU Rice Genome Annotation Project (RGAP) database [28] indicated that the rice CHI family is comprised of seven members designated as OsCHI1–OsCHI7 (Table 1). The open reading frames (ORFs) and amino acids lengths of bona fide CHIs of Arabidopsis, alfalfa, pea, soybean, and maize are 657 to 741 nucleotides and 218 to 246 amino acids long, respectively [12,13,14,29]. Of the OsCHIs, OsCHI1, OsCHI3, OsCHI5, OsCHI6, and OsCHI7 showed comparable ORF sizes (627–843 nucleotides) to bona fide CHIs, which encode polypeptides of 208–280 amino acids long (Table 1). The RGAP database suggests two splicing variants (Os03g60509.1 and Os03g60509.2) of OsCHI3. One variant (Os03g60509.1) has a slightly long ORF (888 nucleotides), and the other variant (Os03g60509.2) shows a similar ORF size to typical plant CHIs. Despite extensive attempts to clone OsCHI3 cDNA from diverse rice tissues, only a smaller variant was cloned from rice leaves. These pieces of evidence suggest that the splicing variant Os03g60509.2 is an expressing form of OsCHI3. The ORF lengths of OsCHI2 and OsCHI4 are longer than those of other CHIs (Table 1) because of their large N-terminal extension (Figure S1).
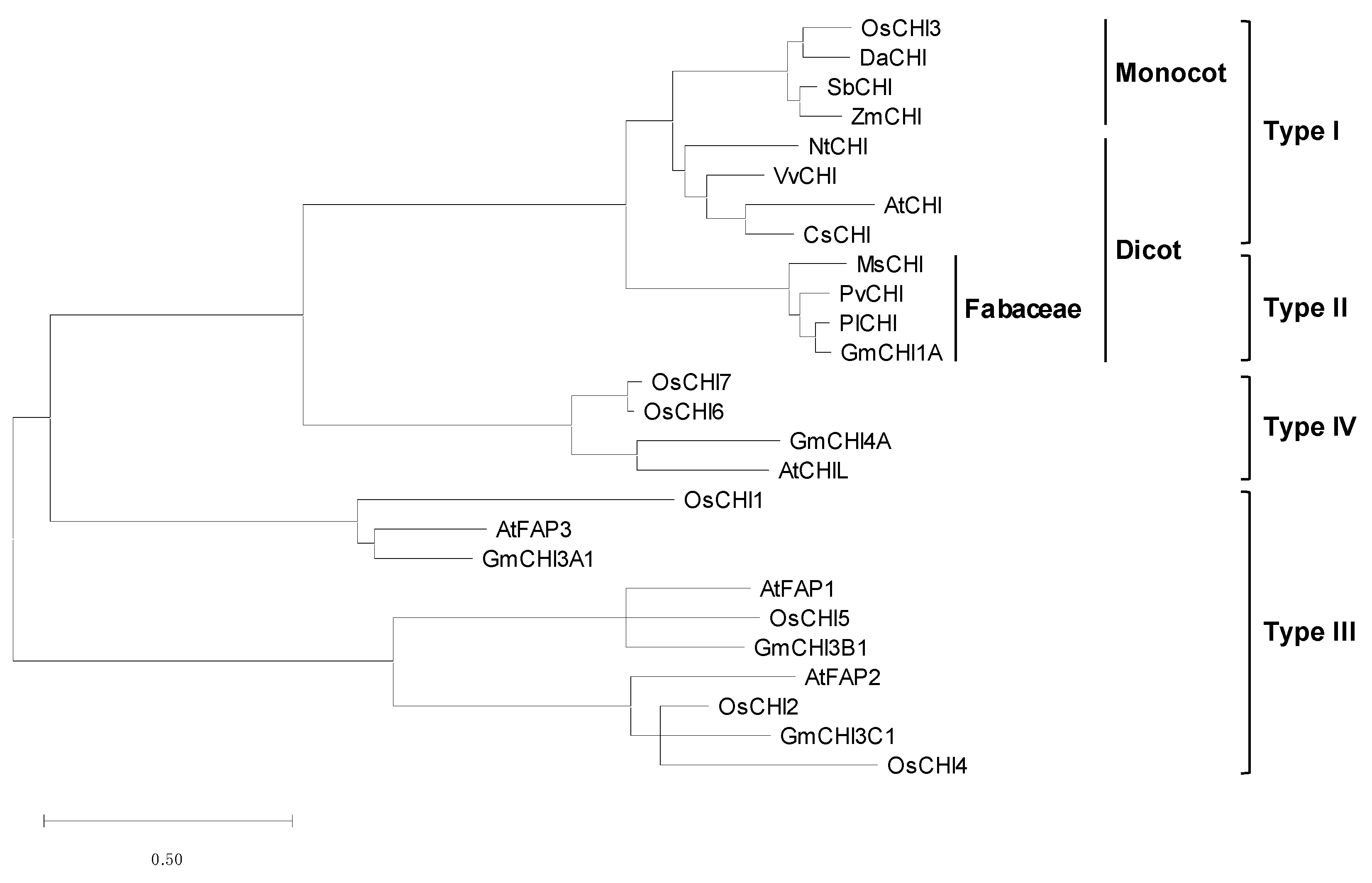
Amino acid sequence homology and phylogenetic relationships between OsCHIs and other plant CHIs were analyzed to classify the OsCHI family (Figure 1 and Table S1). Among the OsCHIs, OsCHI3 is highly homologous to bona fide CHIs, showing 44.0–77.8% sequence identity (Table S1). The other OsCHI members showed 6.0–30.4% identity with bona fide CHIs (Table S1). Phylogenetic analysis revealed that OsCHI3 is classified as type I CHI (Figure 1). Along with type II CHIs, type I CHIs are bona fide CHIs with catalytic activity [11,12,15]. Type II CHIs are homologous to each other, with more than 70% peptide sequence identity, whereas the identity between types I and II is about 50% [11]. Similarly, OsCHI3 showed 44.0–48.9% identity with leguminous type II CHIs, such as Medicago sativa CHI (MsCHI), Phaseolus vulgaris CHI (PvCHI), Pueraria lobata CHI (PlCHI), and Glycine max CHI1A (GmCHI1A). Within type I CHIs, OsCHI3 exhibited greater homology with monocot CHIs, such as Deschampsia antarctica CHI (DaCHI), Zea mays CHI (ZmCHI), and Sorghum bicolor CHI (SbCHI), with 77.8%, 73.6%, and 76.5% identities, respectively, than to dicot CHIs (51.4–58.9% identity), such as Arabidopsis thaliana CHI (AtCHI, also known as TT5), Vitis vinifera CHI (VvCHI), Nicotiana tabacum CHI (NtCHI), and Citrus sinensis CHI (CsCHI) (Figure 1 and Table S1). Phylogenetic analysis also showed that OsCHI3 forms a separate branch with other monocot type I CHIs (Figure 1). Phylogenetic analysis showed that there is no type II CHI in the rice CHI family.
OsCHI1, OsCHI2, OsCHI4, and OsCHI5 are grouped into the type III CHI subfamily (Figure 1). It has been suggested that type III CHIs are distributed widely in land plants and green algae [12,13]. Previous studies have shown that CHI is evolved from fatty acid-binding proteins (FAPs) [13,14]. In the AtCHI family, three members (AtFAP1–3) were found to encode FAPs, categorized as type III CHIs [13]. OsCHI1 is closely related to AtFAP3 and forms one subgroup in the type III CHI subfamily (Figure 1). AtFAP1 and AtFAP2 form the other subgroup, which includes OsCHI 2, OsCHI4, and OsCHI5 (Figure 1). The peptide length of AtFAP2 is longer than that of typical CHIs and is related closely to GmCHI3C1, OsCHI2, and OsCHI4 (Figure 1) [14]. The peptide lengths of these CHI members are about 400 amino acids long (Table 1) [14]. Unlike the type III CHIs, type IV CHIs are found only in land plants [12,13]. OsCHI6 and OsCHI7 are classified as type IV CHIs accompanied by AtCHI-like (AtCHIL) and GmCHI4A (Figure 1) [14].
2.2. Analysis of Conserved Residues within CHIs regarding Substrate-Binding and Catalysis
Three-dimensional structures of MsCHI, AtCHI, and DaCHI were resolved by X-ray crystallography [13,29,30]. The crystal structures revealed the residues forming the substrate-binding clefts of the CHIs and the active site residues participating in the hydrogen-bond networks with substrates and water molecules for catalysis [29,30]. These substrate-binding and catalytic residues are well-conserved in both type I and type II CHIs (Figure 2) [11,12,14,29,30].
The substrate-binding cleft of type II MsCHI is largely apolar and consists of Arg 36, Gly 37, Leu 38, Phe 47, Thr 48, Ile 50, Tyr 106, Lys 109, Val 110, Asn 113, Thr 190, and Met 191 [29]. Most of these residues are conserved in type I DaCHI (Figure 2) [30]. In addition to these residues, Ile 38, Ile 43, Val 94, Met 96, and Leu 100 are involved in the formation of the substrate-binding cleft in DaCHI [30]. Among OsCHIs, the residues forming the substrate-binding cleft are conserved highly in OsCHI3, and they are largely variable in the other OsCHIs (Figure 2 and Figure S1). Type I CHIs show a strong preference for naringenin chalcone, whereas type II CHIs exhibit a broader substrate preference, catalyzing the isomerization of both naringenin chalcone and isoliquiritigenin [11,12,14,15]. The Thr 190 and Met 191 residues of MsCHI have been suggested to participate in substrate preference, and they are conserved among type II CHIs (Figure 2) [29]. These Thr and Met residues are substituted mostly to Ser and Ile, respectively, in type I CHIs, including OsCHI3 (Figure 2). The conserved Val residue (Val 94 in DaCHI) in type I CHIs is replaced with Gly or Arg in type II CHIs (Figure 2). Multiple sequence alignments of bona fide CHIs showed that Leu 38 in MsCHI is conserved in type II CHIs and dicot type I CHIs, whereas it is substituted with Met, Val, and Ile in monocot DaCHI, ZmCHI, and SbCHI, respectively (Figure 2). In OsCHI3, this residue is changed to Val, similar to ZmCHI (Figure 2). The conserved Ile residue (Ile 43 in DaCHI) in monocot type I and type II CHIs is substituted with Val in dicot type I CHIs (Figure 2).
In MsCHI, Thr 48, Lys 97, Tyr 106, Tyr 152, and Thr 190 are involved in the hydrogen-bond networks with substrates and water molecules [29,31]. Of these residues, Thr 48 and Tyr 106 are conserved in all bona fide CHIs (Figure 2) [29,31]. They form hydrogen bonds with catalytic water molecules [29,30,31]. Lys 97 is mostly substituted with Met, and Tyr 152 is changed to Phe in type I CHIs, including OsCHI3 and DaCHI (Figure 2) [31]. As mentioned above, Thr 190 is substituted to Ser in type I CHIs (Figure 2).
2.3. Cloning and Heterologous Expression of OsCHIs
To elucidate the biochemical functions of OsCHIs, we tried to clone OsCHI genes from rice tissues. The cDNAs of OsCHIs were cloned successfully from rice leaves. Regarding the production of recombinant OsCHI proteins, each OsCHI cDNA was inserted into the expression vector pET-28a. Heterologous expression of the N-terminal His-tagged OsCHI proteins in Escherichia coli BL21(DE3) and Rosetta 2(DE3) strains was attempted under various induction conditions. OsCHI3 was expressed successfully in E. coli Rosetta 2 cells as a soluble protein by 0.5 mM isopropyl β-D-thiogalactopyranoside (IPTG) at an 18 °C growth temperature (Figure S2). OsCHI6 and OsCHI7 were expressed as soluble forms in E. coli BL21 cells by 0.5 mM IPTG under an induction temperature of 18 °C (Figure S2). The heterologous expression of OsCHI1 was attempted in both E. coli strains, and only a small amount was produced in E. coli Rosetta 2 cells as a soluble protein at an 18 °C growth temperature following induction with 0.5 mM IPTG (Figure S2). OsCHI2, OsCHI4, and OsCHI5 were not expressed as a soluble form in both E. coli strains.
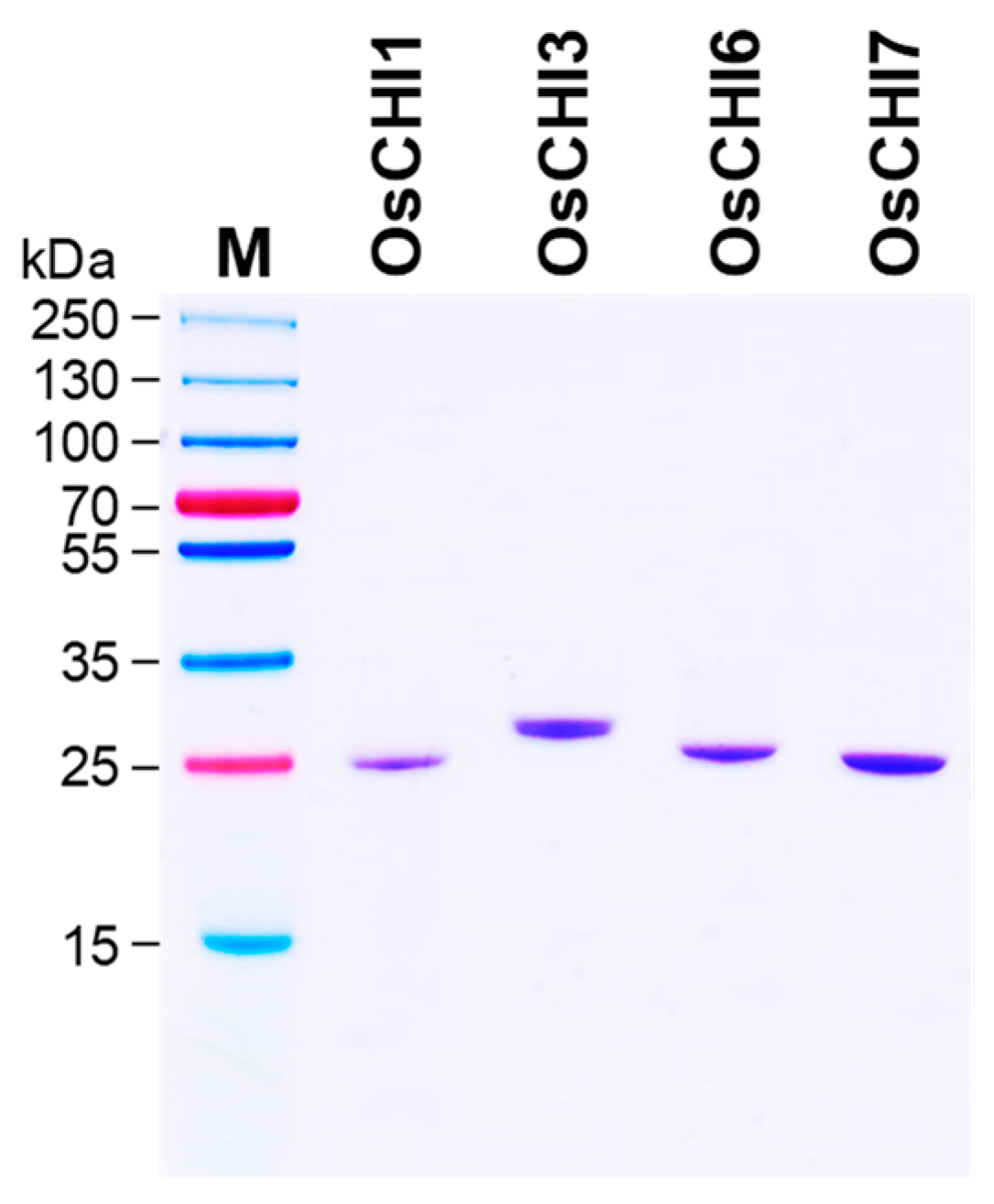
Purification of recombinant OsCHI1, OsCHI3, OsCHI6, and OsCHI7 was attempted with Ni2+-affinity chromatography. OsCHI3, OsCHI6, and OsCHI7 were purified to apparent homogeneity by affinity chromatography, whereas the OsCHI1 fraction contained a large amount of non-specific proteins (Figure 3 and Figure S2). The affinity chromatography fraction was subjected to further cation-exchange chromatography to remove the unwanted proteins from the OsCHI1 preparation, leading to a successful purification of recombinant OsCHI1 to apparent homogeneity (Figure 3 and Figure S2). The purified OsCHI1, OsCHI3, OsCHI6, and OsCHI7 exhibited molecular masses of 25, 28, 25, and 25 kDa on SDS-PAGE, respectively, which are consistent with their theoretical molecular masses (Figure 3).
2.4. CHI Activity and Kinetic Properties of OsCHIs
Plant CHI families include bona fide CHIs and non-catalytic members [11,12,13,14,15]. The catalytic activity of recombinant OsCHI1, OsCHI3, OsCHI6, and OsCHI7 was examined with two substrates, naringenin chalcone and isoliquiritigenin. As expected, type I OsCHI3 showed CHI activity for naringenin chalcone, indicating that it is a bona fide CHI in rice (Figure S3), while the other recombinant OsCHIs showed no detectable CHI activity (Figure S4). As mentioned above, these OsCHIs have large substitutions in the conserved amino acid residues participating in the formation of their substrate-binding clefts and hydrogen bond networks (Figure S1), which can lead to loss of catalytic activity. Type I CHIs have long been thought to be a naringenin chalcone-specific enzyme [11,12,32]. A recent study showed that type I DaCHI also used isoliquiritigenin as a substrate, with a much lesser efficiency [30]. CHI activity for isoliquiritigenin was not detected under the same assay condition (0.1 µg of OsCHI3/mL) as for naringenin chalcone, and an enzyme reaction with a large amount (440 µg/mL) of OsCHI3 showed very slow isomerization of isoliquiritigenin to liquiritigenin (Figure S3).
The kinetic parameters of recombinant OsCHI3 towards naringenin chalcone and isoliquiritigenin were determined (Table 2). The KM values of OsCHI3 for naringenin chalcone and isoliquiritigenin were 11.60 µM and 50.95 µM, respectively (Table 2). These values are comparable to those of other bona fide CHIs, ranging from 1 µM to 112 µM [10,11,12,30,32,33,34,35,36,37]. The kcat values of OsCHI3 for the two examined substrates were very different, with 69.35 s−1 and 9.214 × 10−5 s−1 for naringenin chalcone and isoliquiritigenin, respectively (Table 2). The kcat/KM value of OsCHI for naringenin chalcone was 5.978 × 106 M−1 s−1 (Table 2). CHIs have been appeared to be perfect enzymes [10,11,12,30,33,34,35,36,37]. The kcat/KM values of type I CHIs range from 4.62 × 105 M−1 s−1 to 1.92 × 108 M−1 s−1 [12,30,32,37]. The isomerization efficiency (kcat/KM value of 1.809 M−1 s−1) of OsCHI3 for isoliquiritigenin was extremely lower than that for naringenin chalcone. The difference in catalytic efficiencies of OsCHI3 for naringenin and isoliquiritigenin was much bigger than that (about 1000-fold) of DaCHI [30].
2.5. Analysis of OsCHI Gene Expression
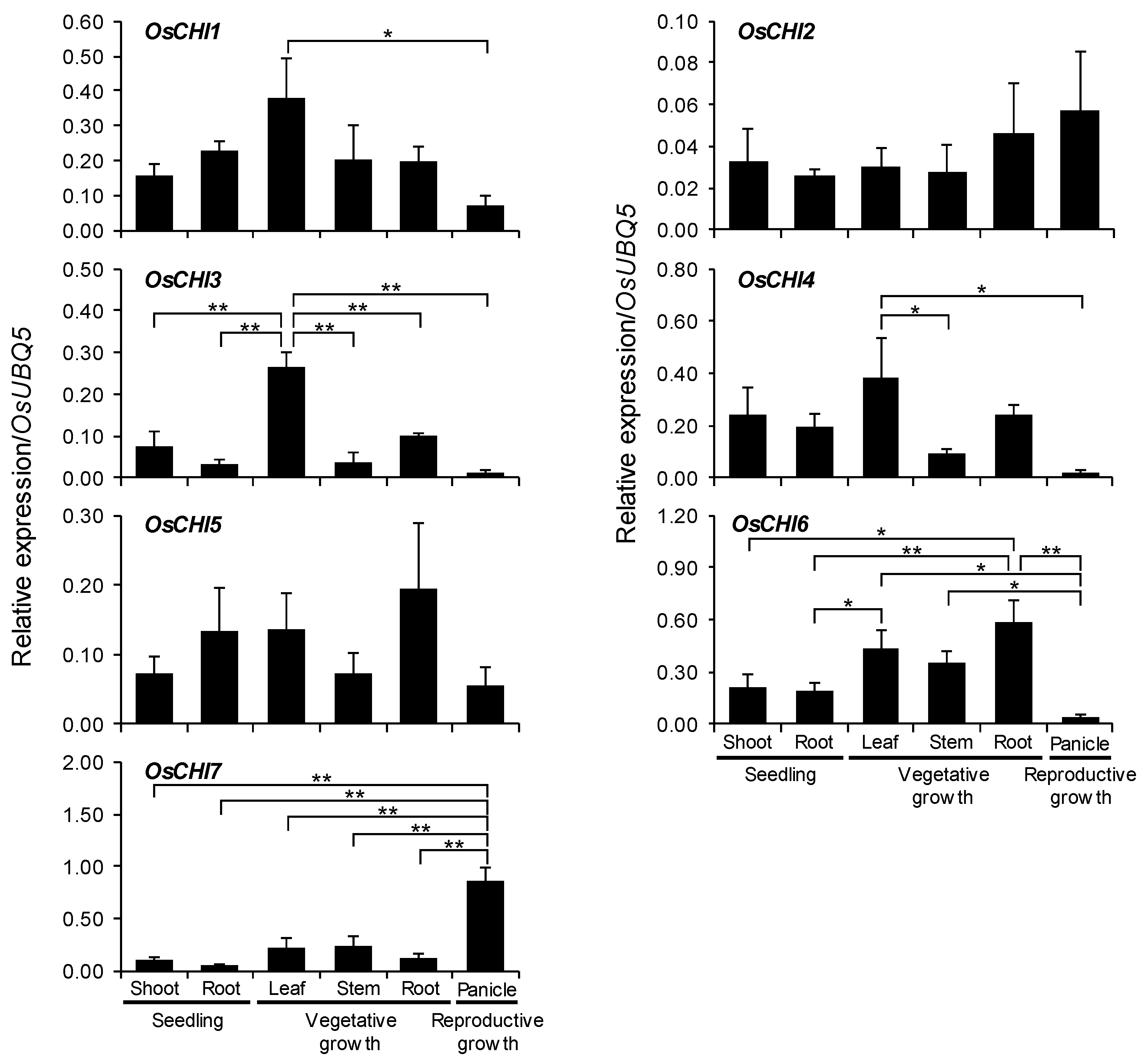
The spatial and temporal expression levels of OsCHIs in rice plants were examined through quantitative real-time polymerase chain reaction (qRT-PCR) analysis. The results showed that all OsCHIs are expressed constitutively throughout the growth period of rice plants (Figure 4 and Figure S5). At the seedling stage, the expression levels of each OsCHI were comparable in both shoots and roots (Figure 4 and Figure S5). At the vegetative growth stage, the bona fide OsCHI3 was expressed highly in leaves compared to stems and roots, exhibiting the lowest expression level in stems (Figure 4 and Figure S5). OsCHI4 expression was higher in leaves than in stems (Figure 4 and Figure S5). OsCHI2, OsCHI6, and OsCHI7 were expressed similarly in leaves, stems, and roots in adult plants (Figure 4). In panicles, expression of OsCHI3, OsCHI4, and OsCHI6 was very low compared to other tissues, while OsCHI7 showed much higher expression, with about 4-fold higher expression than those in leaves and stems (Figure 4 and Figure S5).
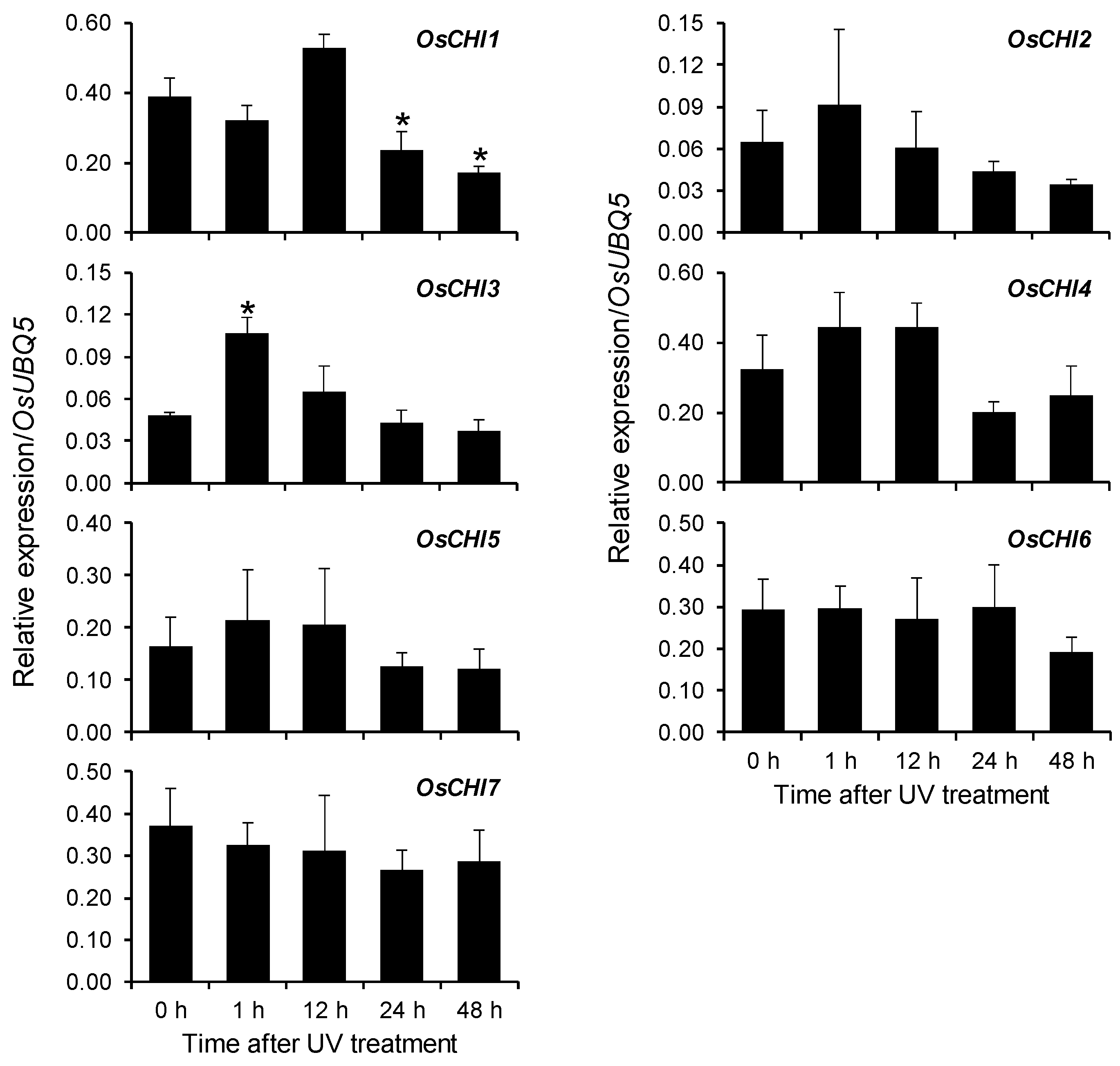
Previous studies have shown that UV stress stimulates the biosynthesis of the flavonoid phytoalexin sakuranetin in rice leaves [9,23,25,26]. High performance liquid chromatography (HPLC) analysis also showed the accumulation of sakuranetin in UV-treated rice leaves (Figure S6). Prior to sakuranetin accumulation, the expression of phenylpropanoid and flavonoid pathway genes, including CHS was upregulated in rice leaves in response to UV irradiation [9,25,27]. The expression of OsCHI3 was increased immediately by UV irradiation, with the highest level at 1 h after UV treatment (Figure 5 and Figure S7). This UV-stimulated expression pattern is similar to that of OsCHS8, which encode a functional rice CHS isozyme [9]. The other OsCHIs showed no significant change in expression in response to UV treatment (Figure 5 and Figure S7).
3. Discussion
3.1. OsCHI3 Is a Bona Fide CHI in the OsCHI Family
In plants, the CHI families consist of several genes [13,14,38]. Soybean contains 12 CHI members [14], and five genes comprise the CHI family in Arabidopsis [13]. Eleven CHI family genes were identified from Dracaena cambodiana [38]. Likewise, seven OsCHI genes were found in the rice genome and were classified into three types, types I, III, and IV (Table 1 and Figure 1). Among family members, a few genes encode bona fide type I and type II CHIs [12,14,38]. In Arabidopsis, only one of five CHI members appears to be a bona fide type I CHI [13,39]. Soybeans have one type I CHI and three type II CHIs among 12 CHI family members [12,14]. Type I CHIs are ubiquitous in vascular plants and participate in naringenin formation, leading to flavonoid biosynthesis [11,12,14]. Type II CHIs have been thought to be legume specific and participate in biosynthesis of isoflavonoids [11,12,14]. Recent studies demonstrated that type II CHIs also exist in basal land plants, such as liverworts, Selaginella moellendorffii, and ferns [36,37]. Phylogenetic analysis predicted that the OsCHI family contains only one type I CHI, OsCHI3 (Figure 1). The OsCHI3 gene was isolated previously from a bacterial artificial chromosome library of rice cv. Nipponbare based on sequence homology to ZmCHI, which was found to complement the tt5 mutation of Arabidopsis [40,41]. An insertion mutation in the 5′-untranslated region of OsCHI3 was reported to exhibit a reddish-brown pigmentation in rice hulls and internodes [42]. With the strict conservation of amino acid residues being important in substrate binding, recombinant OsCHI3 showed CHI activity to naringenin chalcone and isoliquiritigenin (Figure 2 and Table 2). Recombinant OsCHI1, OsCHI6, and OsCHI7 showed no detectable CHI activity (Figure S4), which agrees well with the non-catalytic nature of type III and IV CHIs [11,12,13,14,15]. Kinetic study showed that the kcat/KM value of OsCHI3 for naringenin chalcone is comparable to that of other catalytic CHIs [12,30,32,37]. The catalytic efficiency of OsCHI3 to isoliquiritigenin is extremely lower than that to naringenin chalcone, which agrees well with the property of type I CHIs. These findings ascertain that OsCHI3 is a bona fide CHI in rice.
3.2. Monocot Bona Fide CHIs Categorized into a Separate Branch within Type I CHIs
It has been suggested that bona fide CHIs have strong homology, with more than 70% peptide sequence identity within the types (Table S1) [11]. Monocot type I CHIs, OsCHI3, DaCHI, ZmCHI, and SbCHI, were strongly homologous to each other with 73.6–88.3% identities (Table S1). However, the sequence identities between monocot and dicot type I CHIs were only 51.4–61.6% (Table S1), which resulted in these two groups being categorized into separate branches within type I CHIs (Figure 1). Monocot and dicot type I CHIs also showed a few mismatches in the residues forming the substrate-binding cleft. Leu 38 in MsCHI is conserved in type II CHIs and dicot type I CHIs, whereas it is substituted with other hydrophobic residues, such as Met, Val, and Ile, in monocot OsCHI3, DaCHI, ZmCHI, and SbCHI (Figure 2). This substitution of the Leu residue (Leu 38 in MsCHI) with other nonpolar residues is likely a characteristic change in monocot type I CHIs. The conserved Ile residue (Ile 43 in DaCHI) in monocot type I and type II CHIs is substituted to Val in dicot type I CHIs (Figure 2). These changes likely contributed to the subdivision of type I CHIs into monocot and dicot groups (Figure 1).
3.3. OsCHI3 Participates in UV-Induced Sakuranetin Synthesis in Rice Leaves
Pathogen infections and UV stress stimulate the production of phenolic phytoalexins, including sakuranetin, in rice [25,26,43]. Under UV stress, phenylpropanoid pathway genes have been shown to be upregulated prior to accumulation of phenolic phytoalexins in rice leaves [9,25,27]. A previous study demonstrated that rice contains two catalytic CHS isozymes, OsCHS8 and OsCHS24 [9]. Expression of both OsCHS8 and OsCHS24 appeared to be upregulated by UV irradiation, and they were shown to be redundantly involved in UV-induced sakuranetin synthesis in rice leaves [9]. Of these OsCHSs, OsCHS8 expression was reported to be immediately induced by UV irradiation and subsequently decrease to non-UV-treated levels [9]. Likewise, OsCHI3 is a bona fide CHI, and its expression was increased by UV irradiation, reaching its peak at 1 h after UV treatment (Figure 5 and Figure S7). UV-stimulated expression of the only catalytic OsCHI3 suggests that it contributes to UV-induced sakuranetin synthesis in rice leaves.
3.4. Prospective Role of Non-Catalytic OsCHI Members
Type III and IV CHIs are non-catalytic members of the CHI families, and their physiological functions have long been elusive. Of the AtCHI family members, three type III CHI genes were shown to encode FAPs [13]. A recent study showed that loss-of-function mutations in the AtCHIL gene encoding type IV CHI led to strong reduction of proanthocyanin and flavonol levels in seeds, suggesting that it is an enhancer of the flavonoid pathway [17]. Similarly, mutations in the enhancer of flavonoid production gene encoding a type IV CHI protein resulted in reduction of flavonols and anthocyanins in the flower petals of Ipomoea nil [16]. Humulus lupulus CHI-like 2, a type IV CHI, increased the level of demethylxanthohumol (DMX), a prenylated chalcone, in the engineered yeast harboring all genes required for DMX synthesis [18]. Several studies demonstrated that Type IV CHIs enhance flavonoid production through physical interactions with CHSs [18,19,37].
Constitutive expression of types III and IV OsCHIs throughout growth periods and tissues (Figure 4 and Figure S5) implies that they play a role in rice. However, little is known about the role of types III and IV OsCHIs. Very recently, transgenic rice plants expressing type III OsCHI4 under the control of a stress-inducible promoter were reported to show improved tolerance to abiotic stresses, such as drought, salinity, and cold [44]. Proteomic analysis showed that type IV OsCHI6 and other related enzymes in phytoalexin biosynthesis were involved in the interaction between rice and X. oryzae pv. Oryzae [45]. qRT-PCR analysis showed that type IV OsCHI7 was expressed highly in panicles compared to other rice tissues (Figure 4 and Figure S5). It has been known that CHI participates in seed coloration. The tt5 mutation of Arabidopsis led to produce yellow seeds because of no accumulation of flavonoid pigments [39]. The proanthocyanin levels in seeds were remarkably decreased in the atchil mutants [17]. In this regard, the strong expression of OsCHI7 in panicles suggests its possible involvement in the coloration of rice grains. Therefore, the physiological role of type III and IV OsCHIs, especially their stress-protective roles, need to be elucidated.
4. Materials and Methods
4.1. Plant Growth, UV Treatment, HPLC Analysis, and Materials
Wild-type rice (Oryza sativa L. subsp. Japonica cv. Dongjin) seeds were sterilized with 50% bleach for 30 min and germinated on Murachige and Skoog medium (Duchefa Biochemie, Haarlem, The Netherlands) in a growth chamber with a 16 h light/8 h dark photoperiod at 28 °C for two weeks. The rice seedlings were transferred to soil and grown in a greenhouse at 28 °C during the day. Shoot and root samples were collected from two-week-old rice seedlings. Leaves, stems, and roots were obtained from eight-week-old rice plants during the vegetative growth period. Panicles were harvested from 14-week-old rice plants. Eight-week-old rice plants were irradiated with UV light to examine the UV stress response. UV treatment and HPLC analysis were carried out according to the methods described by Park et al. [25].
Naringenin chalcone and isoliquiritigenin were purchased from Sigma-Aldrich (St. Louis, MO, USA). E. coli BL21(DE3) and Rosetta 2(DE3) strains were obtained from Thermo Fisher Scientific (Waltham, MA, USA). Ni-NTA Agarose beads and CM Sepharose Fast Flow resin were purchased from Qiagen (Hilden, Germany) and Cytiva (Marlborough, MA, USA), respectively. Restriction enzymes were bought from New England Biolabs (Ipswich, MA, USA) and Enzynomics (Daejeon, Korea). Reagents for buffers and media were obtained from Sigma-Aldrich and Duchefa Biochemie.
4.2. Multiple Sequence Alignment and Phylogenetic Analysis of CHIs
Amino acid sequences of OsCHIs and other plant CHIs were obtained from the MSU RGAP database (http://rice.plantbiology.msu.edu/ accessed on 9 May 2021) and the National Center for Biotechnological Information (https://www.ncbi.nlm.nih.gov/ accessed on 28 April 2021) database, respectively. Multiple alignment of the amino acid sequences was conducted with Clustal W [46]. Phylogenetic analysis was performed with the maximum likelihood method, and Whelan and Goldman model using MEGA X [47,48]. Reliability of phylogeny was tested by the bootstrap method, with 100 of bootstrap replications.
4.3. Cloning of OsCHIs
Total RNA was extracted from eight-week-old wild-type rice leaves with RNAiso (Takara, Shiga, Japan). The first-strand cDNA was synthesized from the total RNA according to the methods described by Park et al. [9]. OsCHIs were amplified individually through PCR from the first-strand cDNA. The primer sets used for cloning of OsCHIs and PCR conditions are summarized in Table S2. The amplified PCR products were subcloned into the pJET1.2/blunt vector (Thermo Fisher Scientific). After sequence confirmation, each OsCHI was cut with suitable restriction enzymes, and inserted into the pET-28a(+) vector (Novagen, Madison, WI, USA). The resulting OsCHI/pET-28a(+) constructs were transformed into E. coli BL21(DE3) and Rosetta 2(DE3) strains for the OsCHI expression.
4.4. Production and Purification of Recombinant OsCHIs
The E. coli transformants bearing each OsCHI/pET-28a(+) construct were grown in LB medium supplemented with the appropriate antibiotics at 37 °C. When an OD600 of ~0.6 was reached, different concentrations of IPTG (0.1–1 mM) were added into the cell culture to induce the production of OsCHI proteins, followed by an additional growth period of 16–18 h at various temperatures ranging from 16–37 °C. After induction, the cells were harvested by centrifugation (5000× g for 15 min), and the resulting cell pellets were resuspended with phosphate-buffered saline (10 mM Na2HPO4, 2 mM KH2PO4, 137 mM NaCl, 2.7 mM KCl,) supplemented with lysozyme (1 mg/mL) and phenylmethylsulfonyl fluoride (1 mM). The resuspended cells were lysed by sonication on ice, and the cell debris was removed by centrifugation at 15,000× g for 15 min. Recombinant OsCHI proteins were purified from the crude extract with Ni-NTA agarose chromatography according to the methods described by Park et al. [9]. The recombinant OsCHIs were eluted with 20–150 mM imidazole in Tris buffer (50 mM Tris, pH 8.0, 300 mM NaCl). The affinity chromatography fractions of recombinant OsCHI1 were applied to the CM-Sepharose column equilibrated with sodium phosphate buffer (50 mM, pH 7.0). The recombinant OsCHI1 protein was eluted with 200–300 mM NaCl in the equilibration buffer. The purified OsCHI proteins were analyzed with SDS-PAGE.
4.5. CHI Activity Assay
CHI activity was assayed by monitoring the decrease in absorbance at 390 nm (A390), which results from the conversion of chalcones to their corresponding flavanone [10]. A standard reaction mixture contained 50 µM of naringenin chalcone or isoliquiritigenin and the purified recombinant OsCHI protein (0.1 µg for OsCHI3, 100 µg for OsCHI1, and 500 µg for OsCHI6 and OsCHI7) in Tris buffer (50 mM, pH 7.5) to a total volume of 1 mL. Decreases in A390 were monitored using a Cary 300 Bio UV/Vis spectrophotometer (Varian, Mulgrave, Victoria, Australia). Due to spontaneous isomerization of substrates, the CHI activities of OsCHIs were determined by subtraction of un-catalyzed reactions from the OsCHI-catalyzed reactions. In the determination of kinetic parameters, the amounts of OsCHI3 protein used were 0.1 µg for naringenin chalcone and 500 µg for isoliquiritigenin, and substrate concentrations were 1–50 µM. Enzyme assays were performed in triplicate.
4.6. qRT- PCR Analysis of OsCHIs
Total RNAs from different rice tissues and UV-treated rice leaves were extracted using the Total RNA Prep Kit (Biofact, Daejeon, Korea), and cDNA was synthesized using SuPrimeScript RT premix, and OligodT (GeNet Bio, Daejeon, Korea). qRT-PCR was performed using Prime Q-Mastermix (GeNet Bio, Daejeon, Korea) on a Rotor-Gene Q real-time PCR cycler with Q-Rex Software (Qiagen, Hilden, Germany). Transcript levels were normalized to that of rice ubiquitin 5 (OsUBQ5, Os01g22490) and rice ubiquitin 1 (OsUBQ1, Os03g13170) transcripts as controls. The ΔCt method was applied to calculate OsCHI expression levels, and the primer specificity was assessed with a single peak in the melting curve. The primer sequences and annealing temperature for qRT-PCR are listed in Table S3. Gene expression analysis was performed on triplicated biological samples.
4.7. Statistical Analysis
Data are presented as mean ± standard deviation of three independent experiments. Multiple comparison of OsCHI expressions in rice seedlings and different tissues were performed with one-way ANOVA and Tukey’s HSD test. Significant differences of OsCHI3 expression between UV-treated and untreated samples were determined by t-test. A p-value of <0.05 was considered as statistically significant.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/plants10102064/s1, Figure S1: Multiple alignment of OsCHIs with other CHIs, Figure S2: Purification of recombinant OsCHSs expressed in E. coli, Figure S3: CHI activity assay of recombinant OsCHI3, Figure S4: CHI activity assay of recombinant OsCHI1, OsCHI6, and OsCHI7, Figure S5: qRT-PCR analysis of OsCHI gene expression in rice seedlings and different tissues, Figure S6: HPLC analysis of UV-treated (a) and untreated (b) rice leaves, Figure S7: Expression of OsCHIs in rice leaves in response to UV treatment was analyzed using qRT-PCR, Table S1: Amino acid homology of OsCHIs and bona fide CHIs, Table S2: Primer sequences and PCR conditions for OsCHI cloning, Table S3: Primer sequences and PCR conditions for quantitative real-time PCR analysis.
Author Contributions
Conceptualization, M.-H.C., S.-H.B. and S.-W.L.; investigation, S.-I.P., H.-L.P. and M.-H.C.; writing—original draft preparation, M.-H.C., S.-I.P. and H.-L.P.; writing—review and editing, M.-H.C., H.-L.P., S.-H.B. and S.-W.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Mid-career Researcher Program (NRF-2019R1A2B5B01070202) through NRF grant funded by the Ministry of Education, Science and Technology, Republic of Korea.
Data Availability Statement
The data presented in this study are available within this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Winkel-Shirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol. 2002, 5, 18–223. [Google Scholar] [CrossRef]
- Buer, C.S.; Imin, N.; Djordjevic, M.A. Flavonoids: New roles for old molecules. J. Integr. Plant Biol. 2010, 52, 98–111. [Google Scholar] [CrossRef]
- Ahuja, I.; Kissen, R.; Bones, A.M. Phytoalexins in defense against pathogens. Trends Plant Sci. 2012, 17, 73–90. [Google Scholar] [CrossRef]
- Williams, C.A.; Grayer, R.J. Anthocyanins and other flavonoids. Nat. Prod. Rep. 2004, 21, 539–573. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austin, M.B.; Noel, J. The chalcone synthase superfamily of type III polyketide synthase. Nat. Prod. Rep. 2003, 20, 79–110. [Google Scholar] [CrossRef] [PubMed]
- Abe, I.; Morita, H. Structure and function of the chalcone synthase superfamily of plant type III polyketide synthases. Nat. Prod. Rep. 2010, 27, 809–838. [Google Scholar] [CrossRef] [PubMed]
- Dao, T.T.H.; Linthorst, H.J.M.; Verpoorte, R. Chalcone synthase and its functions in plant resistance. Phytochem. Rev. 2011, 10, 397–412. [Google Scholar] [CrossRef] [Green Version]
- Park, H.L.; Yoo, Y.; Bhoo, S.H.; Lee, T.H.; Lee, S.W.; Cho, M.H. Two chalcone isozymes participate redundantly in UV-induced sakuranetin synthesis in rice. Int. J. Mol. Sci. 2020, 21, 3777. [Google Scholar] [CrossRef]
- Bednar, R.A.; Hadcock, J.R. Purification and characterization of chalcone isomerase from Soybeans. J. Biol. Chem. 1988, 263, 9582–9588. [Google Scholar] [CrossRef]
- Shimada, N.; Aoki, T.; Sato, S.; Nakamura, Y.; Tabata, S.; Ayabe, S. A cluster of genes encodes the two types of chalcone isomerase involved in the biosynthesis of general flavonoids and legume-specific 5-deoxy(iso)flavonoids in Lotus japonicus. Plant Physiol. 2003, 131, 941–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ralston, L.; Subramanian, S.; Matsuno, M.; Yu, O. Partial reconstruction of flavonoid and isoflavonoid biosynthesis in yeast using soybean type I and type II chalcone isomerases. Plant Physiol. 2005, 137, 1375–1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngaki, M.N.; Louie, G.V.; Philippe, R.N.; Manning, G.; Pojer, F.; Bowman, M.E.; Li, L.; Larsen, E.; Wurtele, E.S.; Noel, J.P. Evolution of the chalcone-isomerase fold from fatty-acid binding to stereospecific catalysis. Nature 2012, 485, 530–533. [Google Scholar] [CrossRef] [Green Version]
- Dastmalchi, M.; Dhaubhadel, S. Soybean chalcone isomerase: Evolution of the fold, and the differential expression and localization of the gene family. Planta 2015, 241, 507–523. [Google Scholar] [CrossRef]
- Yin, Y.C.; Zhang, X.D.; Gao, Z.Q.; Hu, T.; Liu, Y. The research progress of chalcone isomerase (CHI) in plants. Mol. Biotechnol. 2019, 61, 32–52. [Google Scholar] [CrossRef]
- Morita, Y.; Takagi, K.; Fukuchi-Mizutani, M.; Ishiguro, K.; Tanaka, Y.; Nitasaka, E.; Nakayama, M.; Saito, N.; Kagami, T.; Hoshino, A.; et al. A chalcone isomerase-like protein enhances flavonoid production and flower pigmentation. Plant J. 2014, 78, 294–304. [Google Scholar] [CrossRef]
- Jiang, W.; Yin, Q.; Wu, R.; Zheng, G.; Liu, J.; Dixon, R.A.; Pang, Y. Role of a chalcone isomerase-like protein in flavonoid biosynthesis in Arabidopsis thaliana. J. Exp. Bot. 2015, 66, 7165–7179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ban, Z.; Qin, H.; Mitchell, A.J.; Liu, B.; Zhang, F.; Weng, J.-K.; Dixon, R.A.; Wang, G. Noncatalytic chalcone isomerase-fold proteins in Humulus lupulus are auxiliary components in prenylated flavonoid biosynthesis. Proc. Natl. Acad. Sci. USA 2018, 115, E5223–E5232. [Google Scholar] [CrossRef] [Green Version]
- Waki, T.; Mameda, R.; Nakano, T.; Yamada, S.; Terashita, M.; Ito, K.; Tenma, N.; Li, Y.; Fujino, N.; Uno, K.; et al. A conserved strategy of chalcone isomerase-like protein to rectify promiscuous chalcone synthase specificity. Nat. Commun. 2020, 11, 870. [Google Scholar] [CrossRef]
- Aoki, T.; Akashi, T.; Ayabe, S. Flavonoids of leguminous plants: Structure, biological activity, and biosynthesis. J. Plant Res. 2000, 113, 475–488. [Google Scholar] [CrossRef]
- Wang, X. Structure, function, and engineering of enzymes in isoflavonoid biosynthesis. Funct. Integr. Genomics 2011, 11, 13–22. [Google Scholar] [CrossRef]
- Cartwright, D.W.; Langcake, P.; Pryce, R.J.; Leworthy, D.P.; Ride, J.P. Isolation and characterization of two phytoalexins from rice as momilactones A and B. Phytochemistry 1981, 20, 535–537. [Google Scholar] [CrossRef]
- Kodama, O.; Miyakawa, J.; Akatsuka, T.; Kiyosawa, S. Sakuranetin, a flavanone phytoalexin from ultraviolet-irradiated rice leaves. Phytochemistry 1992, 31, 3807–3809. [Google Scholar] [CrossRef]
- Peters, R.J. Uncovering the complex metabolic network underlying diterpenoid phytoalexin biosynthesis in rice and other cereal crop plants. Phytochemistry 2006, 67, 2307–2317. [Google Scholar] [CrossRef]
- Park, H.L.; Lee, S.W.; Jung, K.H.; Hahn, T.R.; Cho, M.H. Transcriptomic analysis of UV-treated rice leaves reveals UV-induced phytoalexin biosynthetic pathways and their regulatory networks in rice. Phytochemistry 2013, 96, 57–71. [Google Scholar] [CrossRef]
- Park, H.L.; Yoo, Y.; Hahn, T.R.; Bhoo, S.H.; Lee, S.W.; Cho, M.H. Antimicrobial activity of UV-induced phenylamides from rice leaves. Molecules 2014, 19, 18139–18151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, M.H.; Lee, S.W. Phenolic phytoalexins in rice: Biological functions and biosynthesis. Int. J. Mol. Sci. 2015, 16, 29120–29133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawahara, Y.; De la Bastide, M.; Hamilton, J.P.; Kanamori, H.; McCombie, W.R.; Ouyang, S.; Schwartz, D.C.; Tanaka, T.; Wu, J.; Zhou, S.; et al. Improvement of the Oryza sativa Nipponbare reference genome using next generation sequence and optical map data. Rice 2013, 6, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jez, J.M.; Bowman, M.E.; Dixon, R.A.; Noel, J.P. Structure and mechanism of the evolutionarily unique plant enzyme chalcone isomerase. Nat. Struct. Biol. 2000, 7, 786–791. [Google Scholar] [PubMed]
- Park, S.H.; Lee, C.W.; Cho, S.M.; Lee, H.; Park, H.; Lee, J.; Lee, J.H. Crystal structure and enzymatic properties of chalcone isomerase from the Antarctic vascular plant Deschampsia antarctica Desv. PLoS ONE 2018, 13, e0192415. [Google Scholar] [CrossRef] [Green Version]
- Jez, J.M.; Bowman, M.E.; Noel, J.P. Role of hydrogen bonds in the reaction mechanism of chalcone isomerase. Biochemistry 2002, 41, 5168–5176. [Google Scholar] [CrossRef]
- Fouché, S.D.; Dubery, I.A. Chalcone isomerase from Citrus sinensis: Purification and characterization. Phytochemistry 1994, 37, 127–132. [Google Scholar] [CrossRef]
- Dixon, R.A.; Richard Blyden, E.; Robbins, M.P.; Van Tunen, A.J.; Mol, J.N. Comparative biochemistry of chalcone isomerases. Phytochemistry 1988, 27, 2801–2808. [Google Scholar] [CrossRef]
- Jez, J.M.; Noel, J.P. Reaction mechanism of chalcone isomerase. pH dependence, diffusion control, and product binding differences. J. Biol. Chem. 2002, 277, 1361–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, J.C.; Zhu, L.; Gao, M.J.; Wu, X.; Pan, H.Y.; Zhang, Y.S.; Li, X. Cloning and functional characterization of a chalcone isomerase from Trigonella foenum-graecum L. Planta Med. 2011, 77, 765–770. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.X.; Zhang, X.; Han, X.J.; Zhang, Y.Y.; Gao, S.; Liu, C.J.; Lou, H.X. Identification of chalcone isomerase in the basal land plants reveals an ancient evolution of enzymatic cyclization activity for synthesis of flavonoids. New Phytol. 2018, 217, 909–924. [Google Scholar] [CrossRef] [Green Version]
- Ni, R.; Zhu, T.T.; Zhang, X.S.; Wang, P.Y.; Sun, C.J.; Qiao, Y.N.; Lou, H.X.; Cheng, A.X. Identification and evolutionary analysis of chalcone isomerase-fold proteins in ferns. J. Exp. Bot. 2020, 71, 290–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Zhao, W.; Li, R.; Guo, D.; Li, H.; Wang, Y.; Mei, W.; Peng, S. Identification and characterization of chalcone isomerase genes involved in flavonoid production in Dracaena cambodiana. Front. Plant Sci. 2021, 12, 616396. [Google Scholar] [CrossRef]
- Shirley, B.W.; Kubasek, W.L.; Storz, G.; Bruggemann, E.; Koornneef, M.; Ausubel, F.M.; Goodman, H.M. Analysis of Arabidopsis mutants deficient in flavonoid biosynthesis. Plant J. 1995, 8, 651–671. [Google Scholar] [CrossRef]
- Druka, A.; Kudrna, D.; Rostoks, N.; Brueggeman, R.; Wettstein, D.; Kleinhofs, A. Chalcone isomerase gene from rice (Oryza sativa) and barley (Hordeum vulgare): Physical, genetic and mutation mapping. Gene 2003, 302, 171–178. [Google Scholar] [CrossRef]
- Shih, C.H.; Chu, H.; Tang, L.K.; Sakamoto, W.; Maekawa, M.; Chu, I.K.; Wang, M.; Lo, C. Functional characterization of key structural genes in rice flavonoid biosynthesis. Planta 2008, 228, 1043–1054. [Google Scholar] [CrossRef]
- Hong, L.; Qian, Q.; Tang, D.; Wang, K.; Li, M.; Cheng, Z. A mutation in the rice chalcone isomerase gene causes the golden hull and internode 1 phenotype. Planta 2012, 236, 141–151. [Google Scholar] [CrossRef]
- Ishihara, A.; Hashimoto, Y.; Tanaka, C.; Dubouzet, J.G.; Nakao, T.; Matsuda, F.; Nishioka, T.; Miyagawa, H.; Wakasa, K. The tryptophan pathway is involved in the defense responses of rice against pathogenic infection via serotonin production. Plant J. 2008, 54, 481–495. [Google Scholar] [CrossRef]
- Jayaraman, K.; Raman, K.V.; Sevanthi, A.M.; Sivakumar, S.R.; Viswanathan, C.; Mohapatra, T.; Mandal, P.K. Stress-inducible expression of chalcone isomerase2 gene improves accumulation of flavonoid and imparts enhanced abiotic stress tolerance. Environ. Exp. Bot. 2021, 190, 104582. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, F.; Huang, L.; Zeng, D.; Cruz, C.V.; Li, Z.; Zhou, Y. Comparative proteomic analysis reveals novel insights into the interaction between rice and Xanthomonas oryzae pv. Oryzae. BMC Plant Biol. 2020, 20, 563. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whelan, S.; Goldman, N. A general empirical model of protein evolution derived from multiple protein families using a maximum-likelihood approach. Mol. Biol. Evol. 2001, 18, 691–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Phylogenetic analysis of OsCHIs and other plant CHIs. Amino acid sequences were aligned using Clustal W, and the phylogenetic tree was constructed by the maximum likelihood method using MEGA X. Scale bar denotes branch lengths measured in the number of substitutions per site. Amino acid sequences of plant CHIs used were DaCHI (CBX44252), ZmCHI (Q08704), SbCHI (XP_002463631), VvCHI (NP_001268033), AtCHI (P41088), NtCHI (NP_001312216), MsCHI (P28012), PlCHI (Q43056) GmCHI1A (NP_001235219), PvCHI (P14298), AtCHL (AY063786), AtFAP1 (Q9M1x2), AtFAP2 (Q84RK2), AtFAP3 (Q9C8L2), GmCHI3A1(NP_001238390), GmCHI3B1 (NP_001351383), GmCHI3C1 (Q43056), and GmCHI4A (AAT94362).
Figure 1.
Phylogenetic analysis of OsCHIs and other plant CHIs. Amino acid sequences were aligned using Clustal W, and the phylogenetic tree was constructed by the maximum likelihood method using MEGA X. Scale bar denotes branch lengths measured in the number of substitutions per site. Amino acid sequences of plant CHIs used were DaCHI (CBX44252), ZmCHI (Q08704), SbCHI (XP_002463631), VvCHI (NP_001268033), AtCHI (P41088), NtCHI (NP_001312216), MsCHI (P28012), PlCHI (Q43056) GmCHI1A (NP_001235219), PvCHI (P14298), AtCHL (AY063786), AtFAP1 (Q9M1x2), AtFAP2 (Q84RK2), AtFAP3 (Q9C8L2), GmCHI3A1(NP_001238390), GmCHI3B1 (NP_001351383), GmCHI3C1 (Q43056), and GmCHI4A (AAT94362).

Figure 2.
Multiple alignment of OsCHI3 with bona fide CHIs from other plant species. Identical and similar amino acid residues are shaded in black and grey, respectively. Blue circles above the residues indicate the conserved residues forming the substrate-binding cleft in MsCHI and DaCHI. Red triangles indicate the residues participating in the hydrogen-bond networks for catalysis.
Figure 2.
Multiple alignment of OsCHI3 with bona fide CHIs from other plant species. Identical and similar amino acid residues are shaded in black and grey, respectively. Blue circles above the residues indicate the conserved residues forming the substrate-binding cleft in MsCHI and DaCHI. Red triangles indicate the residues participating in the hydrogen-bond networks for catalysis.

Figure 3.
Purification of recombinant OsCHIs expressed in E. coli. The His-tagged recombinant OsCHI1, OsCHI3, OsCHI6, and OsCHI7 proteins were expressed as a soluble form in E. coli. The recombinant OsCHI3, OsCHI6, and OsCHI7 proteins were purified with Ni2+-affinity chromatography. OsCHI1 was purified with Ni2+-affinity chromatography followed by cation-exchange chromatography. M, Molecular weight marker.
Figure 3.
Purification of recombinant OsCHIs expressed in E. coli. The His-tagged recombinant OsCHI1, OsCHI3, OsCHI6, and OsCHI7 proteins were expressed as a soluble form in E. coli. The recombinant OsCHI3, OsCHI6, and OsCHI7 proteins were purified with Ni2+-affinity chromatography. OsCHI1 was purified with Ni2+-affinity chromatography followed by cation-exchange chromatography. M, Molecular weight marker.

Figure 4.
qRT-PCR analysis of OsCHI gene expression in rice seedlings and different tissues. A ubiquitin gene (OsUBQ5) was amplified using specific primers and was used as an internal control. Expression level of each OsCHI gene is presented as the relative expression compared to the OsUBQ5 mRNA level. Asterisks indicate significant differences (* p < 0.05, ** p < 0.005, Tukey’s HSD-test). qRT-PCR analysis was performed on triplicate biological samples.
Figure 4.
qRT-PCR analysis of OsCHI gene expression in rice seedlings and different tissues. A ubiquitin gene (OsUBQ5) was amplified using specific primers and was used as an internal control. Expression level of each OsCHI gene is presented as the relative expression compared to the OsUBQ5 mRNA level. Asterisks indicate significant differences (* p < 0.05, ** p < 0.005, Tukey’s HSD-test). qRT-PCR analysis was performed on triplicate biological samples.

Figure 5.
Expression of OsCHIs in rice leaves in response to UV treatment was analyzed using qRT-PCR. Rice plants treated with UV irradiation were collected at the designated time points and used to examine the expression of OsCHIs. A ubiquitin gene (OsUBQ5) was amplified using specific primers and used as an internal control. The expression level of each OsCHI gene was presented as the relative expression compared to the OsUBQ5 mRNA level. Asterisks indicate significant differences (p < 0.05, t-test) between UV-untreated and UV-treated samples. qRT-PCR analysis was performed on the triplicated biological samples.
Figure 5.
Expression of OsCHIs in rice leaves in response to UV treatment was analyzed using qRT-PCR. Rice plants treated with UV irradiation were collected at the designated time points and used to examine the expression of OsCHIs. A ubiquitin gene (OsUBQ5) was amplified using specific primers and used as an internal control. The expression level of each OsCHI gene was presented as the relative expression compared to the OsUBQ5 mRNA level. Asterisks indicate significant differences (p < 0.05, t-test) between UV-untreated and UV-treated samples. qRT-PCR analysis was performed on the triplicated biological samples.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Rice chalcone isomerase (CHI) gene family.
| Name | Locus ID | Gene Description in the RGAP Database | ORF (bp) 1 | Protein Size (aa 2) | CHI Type |
|---|---|---|---|---|---|
| OsCHI1 | Os02g21520 | chalcone isomerase 3, putative | 843 | 280 | III |
| OsCHI2 | Os02g53810 | expressed protein | 1284 | 427 | III |
| OsCHI3 | Os03g60509 | expressed protein | 702 | 233 | I |
| OsCHI4 | Os06g10210 | expressed protein | 1290 | 429 | III |
| OsCHI5 | Os07g38390 | chalcone isomerase, putative | 792 | 263 | III |
| OsCHI6 | Os11g02440 | chalcone-flavonone isomerase, putative | 651 | 216 | IV |
| OsCHI7 | Os12g02370 | chalcone-flavonone isomerase, putative | 627 | 208 | IV |
1 ORF; Open reading frame, 2 aa; amino acid.
Table 2.
Kinetic parameters of recombinant OsCHI3 1.
| Substrate | KM (µM) | Vmax (nmol s−1 mg−1) | kcat (s−1) | kcat/KM (M−1 s−1) |
|---|---|---|---|---|
| Naringenin chalcone | 11.60 ± 0.78 | 2662 ± 207.9 | 69.35 | 5.978 × 106 |
| Isoliquiritigenin | 50.95 ± 4.83 | 3.54 × 10−3 ± 1.25 × 10−4 | 9.214 × 10−5 | 1.809 |
1 The results represent the mean ± standard deviation of three independent experiments.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Park, S.-I.; Park, H.-L.; Bhoo, S.-H.; Lee, S.-W.; Cho, M.-H. Biochemical and Molecular Characterization of the Rice Chalcone Isomerase Family. Plants 2021, 10, 2064. https://doi.org/10.3390/plants10102064
AMA Style
Park S-I, Park H-L, Bhoo S-H, Lee S-W, Cho M-H. Biochemical and Molecular Characterization of the Rice Chalcone Isomerase Family. Plants. 2021; 10(10):2064. https://doi.org/10.3390/plants10102064
Chicago/Turabian StylePark, Sang-Il, Hye-Lin Park, Seong-Hee Bhoo, Sang-Won Lee, and Man-Ho Cho. 2021. "Biochemical and Molecular Characterization of the Rice Chalcone Isomerase Family" Plants 10, no. 10: 2064. https://doi.org/10.3390/plants10102064
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.