
Responses of Ecological Stoichiometric Characteristics of Carbon, Nitrogen, and Phosphorus to Periodic Submergence in Mega-Reservoir: Growth of Taxodium distichum and Taxodium ascendens


 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Effects of Submergence Treatment on the Growth
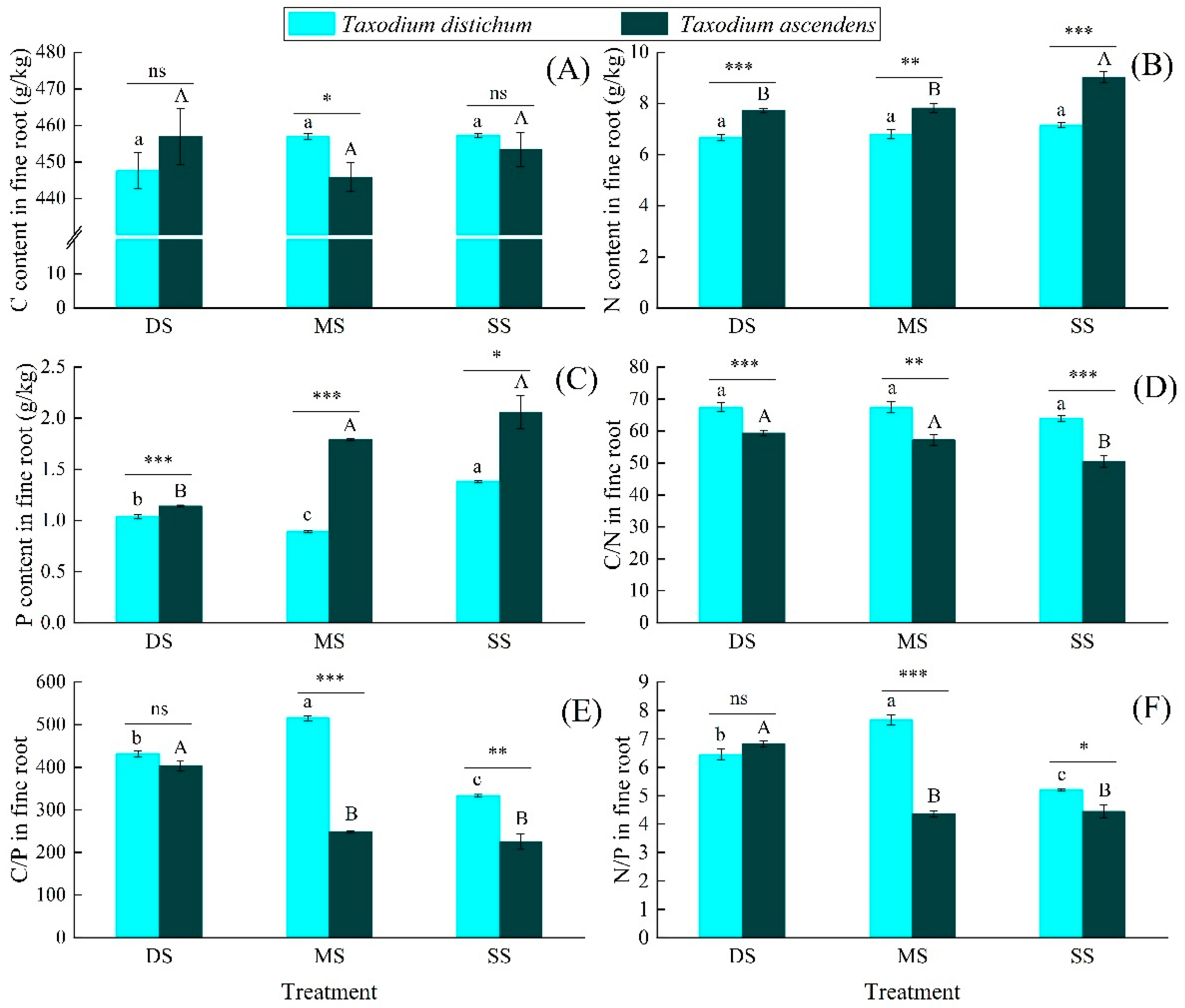
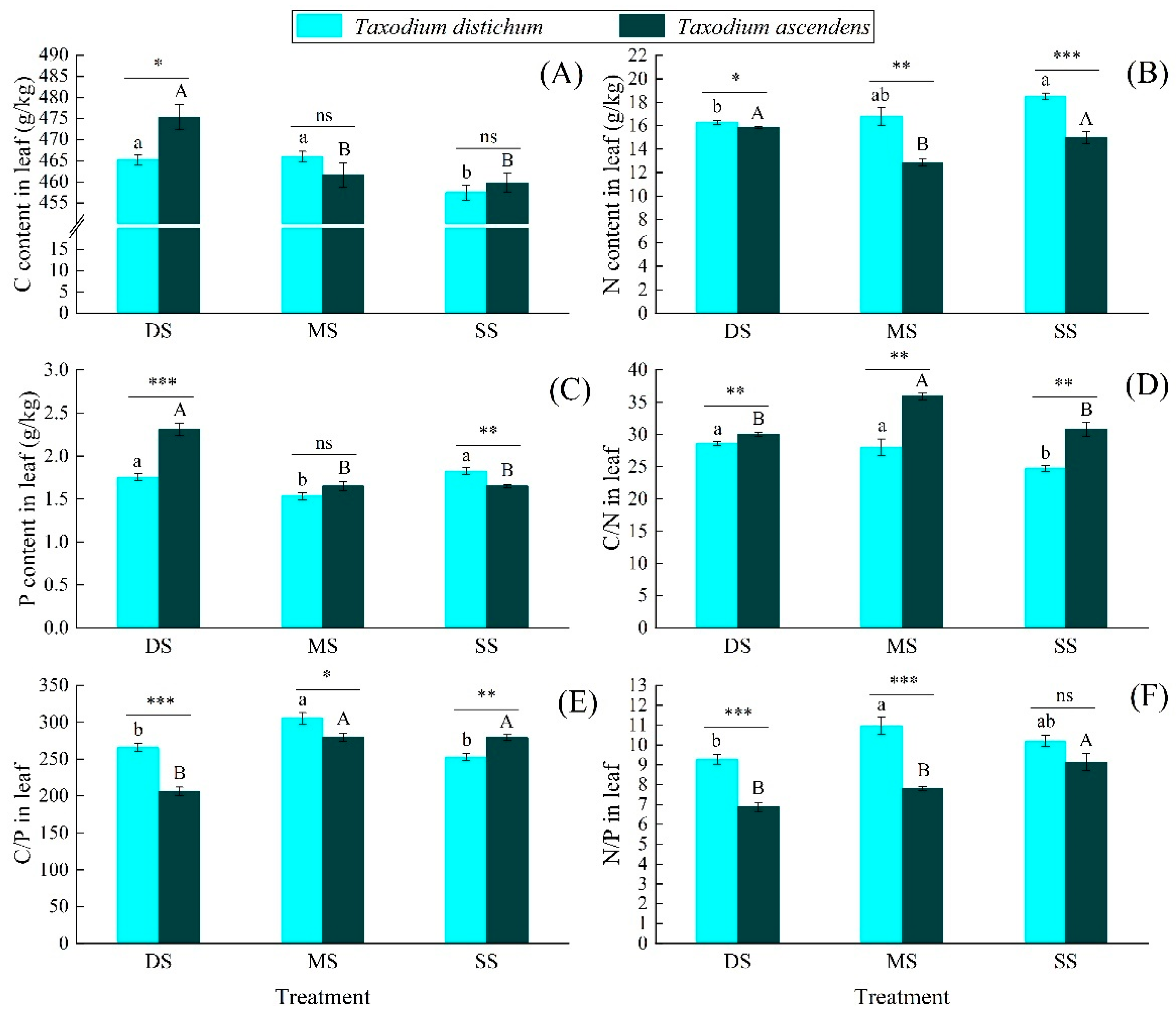
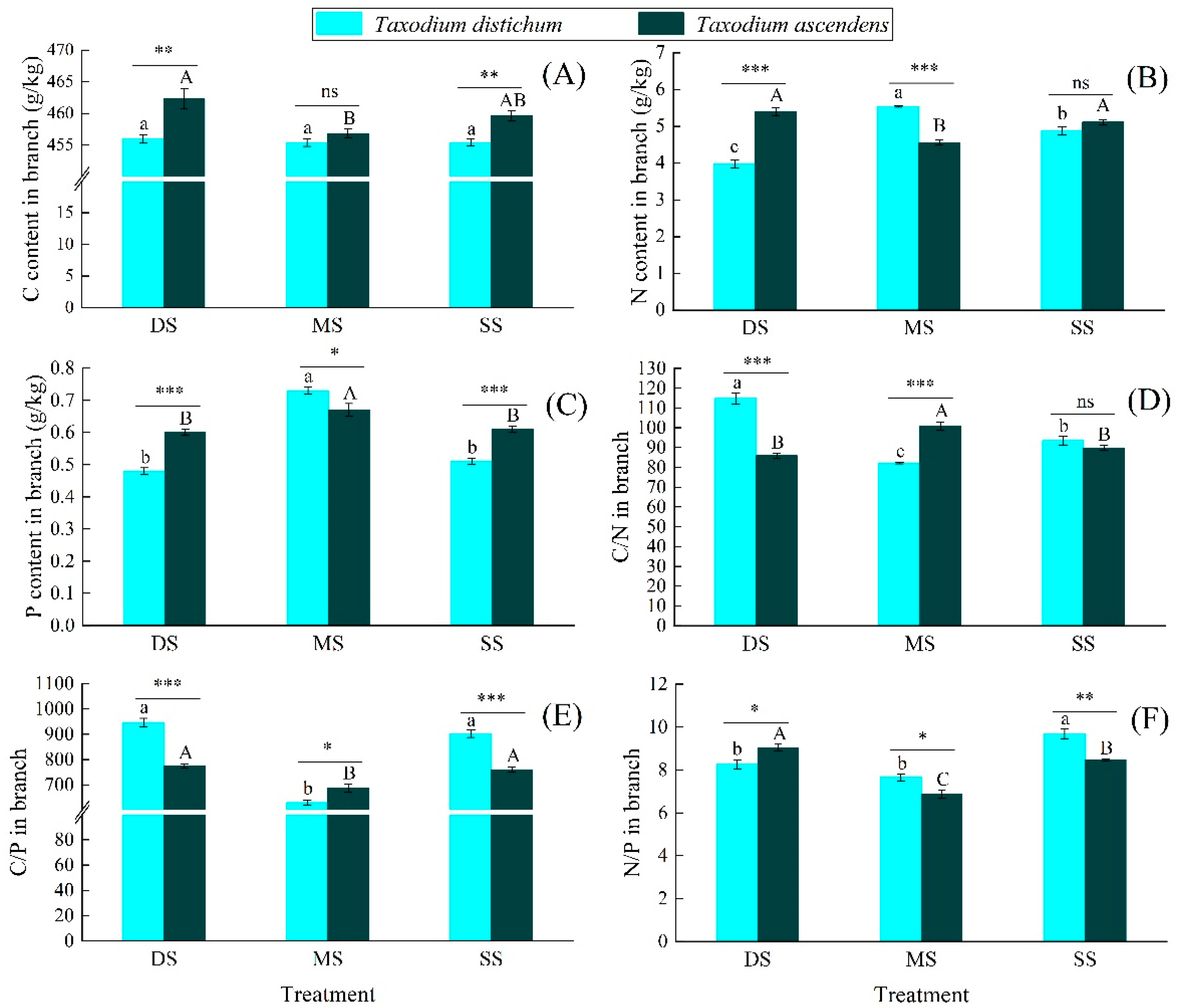
2.2. Effects of Submergence Treatment on the C, N, and P Ecological Stoichiometric Characteristics
2.3. Correlation Analysis of the Growth Conditions and the C, N, and P contents of T. distichum and T. ascendens
3. Discussion
3.1. The Response of Growth to Submergence
3.2. The Responses of C, N, and P Ecological Stoichiometry Characteristics to Submergence
3.3. The Relationship between Growth and C, N, and P Contents
4. Materials and Methods
4.1. Study Site and Experimental Materials
4.2. Sample Collection
4.3. Measurement of Element Content
4.4. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arif, M.; Zheng, J.; Wokadala, C.; Zhang, S.L.; Yuan, Z.X.; Chen, Z.T.; Li, C.X. Assessing riparian zone changes under the influence of stress factors in higher-order streams and tributaries: Implications for the management of massive dams and reservoirs. Sci. Total Environ. 2021, 776, 146011. [Google Scholar] [CrossRef]
- Zheng, J.; Arif, M.; Zhang, S.L.; Yuan, Z.X.; Zhang, L.M.; Dong, Z.; Li, C.X. The convergence of species composition along the drawdown zone of the Three Gorges Dam Reservoir, China: Implications for restoration. Environ. Sci. Pollut. Res. 2021, 28, 42609–42621. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.T.; Arif, M.; Wang, C.Y.; Chen, X.M.; Li, C.X. Effects of hydrological regime on foliar decomposition and nutrient release in the riparian zone of the Three Gorges Reservoir, China. Front. Plant Sci. 2021, 12, 661865. [Google Scholar] [CrossRef]
- He, X.R.; Wang, T.; Wu, K.J.; Wang, P.; Qi, Y.C.; Arif, M.; Wei, H. Responses of swamp cypress (Taxodium distichum) and Chinese willow (Salix matsudana) roots to periodic submergence in mega-reservoir: Changes in organic acid concentration. Forests 2021, 12, 203. [Google Scholar] [CrossRef]
- Arif, M.; Zhang, S.L.; Zheng, J.; Wokadala, C.; Sanelisiwe, M.P.; Li, C.X. Evaluating the effects of pressure indicators on riparian zone health conditions in the three gorges dam reservoir, China. Forests 2020, 11, 214. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Fu, C.; Li, Z.; Fang, F.; Ouyang, W.J.; Guo, J.S. Effect of dissolved organic matters on adsorption and desorption behavior of heavy metals in a water-level-fluctuation zone of the Three Gorges Reservoir, China. Ecotoxicol. Environ. Saf. 2019, 185, 109695. [Google Scholar] [CrossRef]
- Ye, C.; Chen, C.R.; Butler, O.M.; Rashti, M.R.; Esfandbod, M.; Du, M.; Zhang, Q.F. Spatial and temporal dynamics of nutrients in riparian soils after nine years of operation of the Three Gorges Reservoir, China. Sci. Total Environ. 2019, 664, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Li, X.L.; Wang, X.S.; Xiang, L.; Yang, J.; Min, Q.F.; Wang, G.X. Growth and respiratory metabolic adaptation strategies of riparian plant Distylium chinense to submergence by the field study and controlled experiments. Plant Physiol. Biochem. 2020, 157, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Li, C.X.; Zhong, Z.C.; Geng, Y.H.; Schneider, R. Comparative studies on physiological and biochemical adaptation of Taxodium distichum and Taxodium ascendens seedlings to different soil water regimes. Plant Soil 2010, 329, 481–494. [Google Scholar] [CrossRef]
- Wang, C.Y.; Li, C.X.; Wei, H.; Xie, Y.Z.; Han, W.J. Effects of long-term periodic submergence on photosynthesis and growth of Taxodium distichum and Taxodium ascendens saplings in the hydro-fluctuation zone of the Three Gorges Reservoir of China. PLoS ONE 2016, 11, e0162867. [Google Scholar] [CrossRef]
- Wang, C.Y.; Xie, Y.Z.; Ren, Q.S.; Li, C.X. Leaf decomposition and nutrient release of three tree species in the hydro-fluctuation zone of the Three Gorges Dam Reservoir, China. Environ. Sci. Pollut. Res. 2018, 25, 23261–23275. [Google Scholar] [CrossRef]
- Wang, T.; Wei, H.; Ma, W.C.; Zhou, C.; Chen, H.C.; Li, R.; Li, S. Response of Taxodium distichum to winter submergence in the water-level-fluctuating zone of the Three Gorges Reservoir region. J. Freshw. Ecol. 2019, 34, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.Q.; Chen, Z.B.; Yan, X.Y.; Bai, L.Y. Stoichiometric mechanisms of Dicranopteris dichotoma growth and resistance to nutrient limitation in the Zhuxi watershed in the red soil hilly region of China. Plant Soil 2016, 398, 367–379. [Google Scholar] [CrossRef]
- Shang, B.; Feng, Z.Z.; Li, P.; Calatayud, V. Elevated ozone affects C, N and P ecological stoichiometry and nutrient resorption of two poplar clones. Environ. Pollut. 2018, 234, 136–144. [Google Scholar] [CrossRef]
- Sperfeld, E.; Wagner, N.D.; Halvorson, H.M.; Malishev, M.; Raubenheimer, D. Bridging ecological stoichiometry and nutritional geometry with homeostasis concepts and integrative models of organism nutrition. Funct. Ecol. 2016, 31, 286–296. [Google Scholar] [CrossRef]
- Tang, Z.Y.; Xu, W.T.; Zhou, G.Y.; Bai, Y.F.; Li, J.X.; Tang, X.L.; Xie, Z.Q. Patterns of plant carbon, nitrogen, and phosphorus concentration in relation to productivity in China’s terrestrial ecosystems. Proc. Natl. Acad. Sci. USA 2018, 115, 4033–4038. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.F.; Zhang, W.L.; Zhang, L.H.; Chen, X.Y.; Lin, W.; Zeng, C.S.; Tong, C. Stoichiometric characteristics of nitrogen and phosphorus in major wetland vegetation of China. Chin. J. Plant Ecol. 2014, 38, 1041–1052. [Google Scholar] [CrossRef]
- Cao, Y.; Li, Y.N.; Zhang, G.Q.; Zhang, J.; Chen, M. Fine root C:N:P stoichiometry and its driving factors across forest ecosystems in northwestern China. Sci. Total Environ. 2020, 737, 140299. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Peng, Q.W.; Li, K.H.; Gong, Y.M.; Liu, Y.Y.; Han, W.X. Patterns of nitrogen and phosphorus stoichiometry among leaf, stem and root of desert plants and responses to climate and soil factors in Xinjiang, China. Catena 2021, 199, 105100. [Google Scholar] [CrossRef]
- Hou, S.L.; Lü, X.T.; Yin, J.X.; Yang, J.J.; Hu, Y.Y.; Wei, H.W.; Han, X.G. The relative contributions of intra- and inter-specific variation in driving community stoichiometric responses to nitrogen deposition and mowing in a grassland. Sci. Total Environ. 2019, 666, 887–893. [Google Scholar] [CrossRef]
- Dibar, D.T.; Zhang, K.; Yuan, S.Q.; Zhang, J.Y.; Zhou, Z.Z.; Ye, X.X. Ecological stoichiometric characteristics of Carbon (C), Nitrogen (N) and Phosphorus (P) in leaf, root, stem, and soil in four wetland plants communities in Shengjin Lake, China. PLoS ONE 2020, 15, e0230089. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.S.; Reich, P.B.; Chen, H.Y.H.; Xiang, Y.Z.; Luo, Y.P.; Shen, Y.; Niu, S.L. Global changes alter plant multi-element stoichiometric coupling. New Phytol. 2019, 221, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.; Yan, Z.B.; Niklas, K.J.; Han, W.X.; Kattge, J.; Reich, P.B.; Fang, J.Y. Global leaf nitrogen and phosphorus stoichiometry and their scaling exponent. Natl. Sci. Rev. 2018, 5, 728–739. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Liu, Q.; Yin, H.J.; Zhao, C.Z.; Zhang, L.; Zhou, G.Y.; Xie, Z.Q. C:N:P stoichiometry of Ericaceae species in shrubland biomes across Southern China: Influences of climate, soil and species identity. J. Plant Ecol. 2019, 12, 346–357. [Google Scholar] [CrossRef]
- Zhang, K.; Li, M.M.; Su, Y.Z.; Yang, R. Stoichiometry of leaf carbon, nitrogen, and phosphorus along a geographic, climatic, and soil gradients in temperate desert of Hexi Corridor, northwest China. J. Plant Ecol. 2020, 13, 114–121. [Google Scholar] [CrossRef]
- Han, W.X.; Fang, J.Y.; Guo, D.L.; Zhang, Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol. 2005, 168, 377–385. [Google Scholar] [CrossRef]
- Minden, V.; Kleyer, M. Internal and external regulation of plant organ stoichiometry. Plant Biol. 2014, 16, 897–907. [Google Scholar] [CrossRef]
- Elser, J.J.; Fagan, W.F.; Kerkhoff, A.J.; Swenson, N.G.; Enquist, B.J. Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change. New Phytol. 2010, 186, 593–608. [Google Scholar] [CrossRef] [Green Version]
- Kleyer, M.; Minden, V. Why functional ecology should consider all plant organs: An allocation-based perspective. Basic Appl. Ecol. 2015, 16, 1–9. [Google Scholar] [CrossRef]
- Yan, Z.B.; Li, P.; Chen, Y.H.; Han, W.X.; Fang, J.Y. Nutrient allocation strategies of woody plants: An approach from the scaling of nitrogen and phosphorus between twig stems and leaves. Sci. Rep. 2016, 6, 20099. [Google Scholar] [CrossRef] [Green Version]
- Lugli, L.F.; Rosa, J.S.; Andersen, K.M.; Ponzio, R.D.; Almeida, R.V.; Pires, M.; Hartley, I.P. Rapid responses of root traits and productivity to phosphorus and cation additions in a tropical lowland forest in Amazonia. New Phytol. 2021, 230, 116–128. [Google Scholar] [CrossRef]
- Colmer, T.D.; Voesenek, L.J.F.P.B. Flooding tolerance: Suites of plant traits in variable environments. Funct. Plant Biol. 2009, 36, 665–681. [Google Scholar] [CrossRef] [PubMed]
- McAlhaney, A.L.; Keim, R.F.; Allen, S.T. Species-specific growth capacity for floodplain forest trees inferred from sapwood efficiency and individual tree competition. For. Ecol. Manag. 2020, 476, 118427. [Google Scholar] [CrossRef]
- Oram, N.J.; De Deyn, G.B.; Bodelier, P.L.E.; Cornelissen, J.H.C.; Groenigen, J.W.; Abalos, D. Plant community flood resilience in intensively managed grasslands and the role of the plant economic spectrum. J. Appl. Ecol. 2020, 57, 1524–1534. [Google Scholar] [CrossRef]
- Zhou, W.G.; Chen, F.; Meng, Y.J.; Chandrasekaran, U.; Luo, X.F.; Yang, W.Y.; Shu, K. Plant waterlogging/flooding stress responses: From seed germination to maturation. Plant Physiol. Biochem. 2020, 148, 228–236. [Google Scholar] [CrossRef]
- Conserva, A.; Camargo, J.L.C.; Santana, D.G.D.; Piedade, M.T.F. Germinative behaviour of ten tree species in white-water floodplain forests in central Amazonia. Folia Geobot. 2017, 53, 89–101. [Google Scholar] [CrossRef]
- Jackson, M.B. Ethylene-promoted elongation: An adaptation to submergence stress. Ann. Bot. 2008, 101, 229–248. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef]
- Jia, W.; Ma, M.; Chen, J.; Wu, S. Plant Morphological, Physiological and Anatomical Adaption to Flooding Stress and the Underlying Molecular Mechanisms. Int. J. Mol. Sci. 2021, 22, 1088. [Google Scholar] [CrossRef]
- Hong, J.T.; Wang, X.D.; Wu, J.B. Stoichiometry of root and leaf nitrogen and phosphorus in a dry alpine steppe on the Northern Tibetan Plateau. PLoS ONE 2014, 9, e109052. [Google Scholar] [CrossRef]
- Liu, L.; Ge, J.L.; Shu, H.W.; Zhao, C.M.; Xu, W.T.; Shen, G.Z.; Xie, Z.Q. C, N and P stoichiometric ratios in mixed evergreen and deciduous broadleaved forests in Shennongjia, China. Chin. J. Plant Ecol. 2019, 43, 482–489. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, B.R.; An, S.S. Ecological stoichiometry in leaves, roots, litters and soil among different plant communities in a desertified region of Northern China. Catena 2018, 166, 328–338. [Google Scholar] [CrossRef]
- He, M.Z.; Zhang, K.; Tan, H.J.; Hu, R.; Wang, J.; Li, X.R. Nutrient levels within leaves, stems, and roots of the xeric species Reaumuria soongorica in relation to geographical, climatic, and soil conditions. Ecol. Evol. 2015, 5, 1494–1503. [Google Scholar] [CrossRef]
- Hong, J.T.; Wang, X.D.; Wu, J.B. Variation in carbon, nitrogen and phosphorus partitioning between above- and belowground biomass along a precipitation gradient at Tibetan Plateau. J. Mt. Sci. 2016, 13, 661–671. [Google Scholar] [CrossRef]
- Li, A.; Guo, D.L.; Wang, Z.Q.; Liu, H.Y. Nitrogen and phosphorus allocation in leaves, twigs, and fine roots across 49 temperate, subtropical and tropical tree species: A hierarchical pattern. Funct. Ecol. 2010, 24, 224–232. [Google Scholar] [CrossRef]
- Elser, J.J.; Bracken, M.E.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Liu, W.C.; Xu, M.P.; Deng, J.; Han, X.H.; Yang, G.H.; Ren, G.X. Response of forest growth to C:N:P stoichiometry in plants and soils during Robinia pseudoacacia afforestation on the Loess Plateau, China. Geoderma 2019, 337, 280–289. [Google Scholar] [CrossRef]
- Ågren, G.I. Stoichiometry and Nutrition of Plant Growth in Natural Communities. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 153–170. [Google Scholar] [CrossRef]
- Niklas, K.J.; Owens, T.; Reich, P.B.; Cobb, E.D. Nitrogen/phosphorus leaf stoichiometry and the scaling of plant growth. Ecol. Lett. 2005, 8, 636–642. [Google Scholar] [CrossRef]
- Duan, Y.F.; Zhou, Q.Q.; Lu, X.Y.; Wang, J.F.; Li, X.W.; Yang, J.L. Leaf Stoichiometric Characteristics of Different Vegetation Types on the Eastern Slope of Helan Mountains. Acta Bot. Boreali-Occident. Sin. 2020, 40, 113–120. [Google Scholar] [CrossRef]
- Liu, J.X.; Fang, X.; Tang, X.L.; Wang, W.T.; Han, S.J.; Ma, Y.X. Patterns and controlling factors of plant nitrogen and phosphorus stoichiometry across China’s forests. Biogeochemistry 2019, 143, 191–205. [Google Scholar] [CrossRef]
- Hu, M.J.; Peñuelas, J.; Sardans, J.; Sun, Z.G.; Wilson, B.J.; Huang, J.F.; Tong, C. Stoichiometry patterns of plant organ N and P in coastal herbaceous wetlands along the East China Sea: Implications for biogeochemical niche. Plant Soil. 2018, 431, 273–288. [Google Scholar] [CrossRef]
- Dusenge, M.E.; Duarte, A.G.; Way, D.A. Plant carbon metabolism and climate change: Elevated CO2 and temperature impacts on photosynthesis, photorespiration and respiration. New Phytol. 2019, 221, 32–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acosta, D.J.B.S. Responses of photosynthetic and stoichiometric traits to aridity in species and functional types of two sonoran desert plant communities. Bot. Sci. 2021, 99, 257–278. [Google Scholar]
- Tian, D.; Li, P.; Fang, W.J.; Xu, J.; Luo, Y.K.; Yan, Z.B.; Fang, J.Y. Growth responses of trees and understory plants to nitrogen fertilization in a subtropical forest in China. Biogeosciences 2017, 14, 3461–3469. [Google Scholar] [CrossRef] [Green Version]
- Domingues, T.F.; Meir, P.; Feldpausch, T.R.; Saiz, G.; Veenendaal, E.M.; Schrodt, F.; Lloyd, J. Co-limitation of photosynthetic capacity by nitrogen and phosphorus in West Africa woodlands. Plant Cell Environ. 2010, 33, 959–980. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, J.N.; Wang, J.Y.; Guo, Z.W.; Wang, G.G.; Zeng, D.H.; Wu, T.G. Tree stoichiometry and nutrient resorption along a chronosequence of Metasequoia glyptostroboides forests in coastal China. For. Ecol. Manag. 2018, 430, 445–450. [Google Scholar] [CrossRef]
- Reef, R.; Ball, M.C.; Feller, I.C.; Lovelock, C.E. Relationships among RNA:DNA ratio, growth and elemental stoichiometry in mangrove trees. Func. Ecol. 2010, 24, 1064–1072. [Google Scholar] [CrossRef]
- Kerkhoff, A.J.; Fagan, W.F.; Elser, J.J.; Enquist, B.J. Phylogenetic and growth form variation in the scaling of nitrogen and phosphorus in the seed plants. Am. Nat. 2006, 168, E103–E122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Z.Y.; Chen, H.Y.; Reich, P.B. Global-scale latitudinal patterns of plant fine-root nitrogen and phosphorus. Nat. Commun. 2011, 2, 344. [Google Scholar] [CrossRef] [Green Version]
- Fujita, H.; Hayashi-Tsugane, M.; Kawaguchi, M. Spatial regulation of resource allocation in response to nutritional availability. J. Theor. Biol. 2020, 486, 110078. [Google Scholar] [CrossRef] [PubMed]
- Arif, M.; Tahir, M.; Zheng, J.; Li, C.X. Impacts of riparian width and stream channel width on ecological networks in main waterways and tributaries. Sci. Total Environ. 2021, 792, 148457. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Ni, X.; Arif, M.; Dong, Z.; Zhang, L.; Tan, X.; Li, C.X. Transcriptomic analysis of the photosynthetic, respiration and aerenchyma adaptation strategies in bermudagrass (Cynodon dactylon) under different submergence stress. Int. J. Mol. Sci. 2021, 22, 7905. [Google Scholar] [CrossRef]
- Jiajia, L.; Lijuan, L.; Arif, M.; Dongdong, D.; Xin, H.; Jie, Z.; Zhongxun, Y.; Changxiao, L. Artificial plantation responses to periodic submergence in massive dam and reservoir riparian zones: Changes in soil properties and bacterial community characteristics. Biology 2021, 10, 819. [Google Scholar] [CrossRef]
- Zheng, J.; Arif, M.; Zhang, S.; Yuan, Z.; Zhang, L.; Li, J.; Li, C. Dam inundation simplifies the plant community composition. Sci. Total Environ. 2021, 801, 149827. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Parameters | Taxodium distichum | Taxodium ascendens | ||||
|---|---|---|---|---|---|---|
| SS | MS | DS | SS | MS | DS | |
| Height (m) | 6.87 ± 0.07 a | 5.59 ± 0.01 b | 5.41 ± 0.01 c | 6.27 ± 0.04 a | 5.73 ± 0.03 b | 5.46 ± 0.04 c |
| Crown area (m2) | 8.01 ± 0.17 a | 6.45 ± 0.11 b | 4.79 ± 0.28 c | 7.23 ± 0.20 a | 5.81 ± 0.21 b | 5.03 ± 0.08 c |
| Basal diameter (mm) | 104.95 ± 0.99 a | 94.58 ± 1.91 b | 86.51 ± 1.71 c | 100.16 ± 3.22 a | 94.35 ± 2.20 ab | 86.98 ± 2.34 b |
| DBH (mm) | 73.12 ± 1.99 a | 65.00 ± 0.56 b | 56.85 ± 0.21 c | 79.37 ± 2.30 a | 71.58 ± 2.78 ab | 66.35 ± 2.74 b |
| Indexes | Submergence | Organ | Submergence × Organ | |||
|---|---|---|---|---|---|---|
| F | p | F | p | F | p | |
| C content | 2.539 | 0.093 | 18.826 | 0.000 *** | 5.955 | 0.001 *** |
| N content | 12.563 | 0.000 *** | 1493.660 | 0.000 *** | 5.331 | 0.002 ** |
| P content | 44.706 | 0.000 *** | 1545.718 | 0.000 *** | 60.428 | 0.000 *** |
| C/N ratio | 46.386 | 0.000 *** | 1563.755 | 0.000 *** | 37.627 | 0.000 *** |
| C/P ratio | 37.580 | 0.000 *** | 2585.621 | 0.000 *** | 186.738 | 0.000 *** |
| N/P ratio | 7.390 | 0.002 ** | 177.072 | 0.000 *** | 24.142 | 0.000 *** |
| Indexes | Submergence | Organ | Submergence × Organ | |||
|---|---|---|---|---|---|---|
| F | p | F | p | F | p | |
| C content | 6.005 | 0.006 ** | 10.217 | 0.000 *** | 1.134 | 0.356 |
| N content | 32.213 | 0.000 *** | 1417.414 | 0.000 *** | 14.366 | 0.000 *** |
| P content | 1.748 | 0.189 | 361.708 | 0.000 *** | 49.614 | 0.000 *** |
| C/N ratio | 29.012 | 0.000 *** | 1607.768 | 0.000 *** | 12.423 | 0.000 *** |
| C/P ratio | 22.472 | 0.000 *** | 1996.119 | 0.000 *** | 49.289 | 0.000 *** |
| N/P ratio | 31.091 | 0.000 *** | 190.933 | 0.000 *** | 38.294 | 0.000 *** |
| Growth Parameters | C in Fine Root | N in Fine Root | P in Fine Root | C in Leaf | N in Leaf | P in Leaf | C in Branch | N in Branch | P in Branch |
|---|---|---|---|---|---|---|---|---|---|
| Height | 0.373 | 0.588 * | 0.904 ** | −0.785 ** | 0.712 ** | 0.499 | −0.077 | 0.203 | −0.315 |
| Crown area | 0.435 | 0.516 * | 0.642 ** | −0.678 ** | 0.685 ** | 0.163 | −0.221 | 0.505 | 0.069 |
| Basal diameter | 0.465 | 0.685 ** | 0.649 ** | −0.578 * | 0.553 * | 0.289 | −0.139 | 0.480 | 0.024 |
| DBH | 0.537 * | 0.524 * | 0.627 * | −0.598 * | 0.669 ** | 0.217 | −0.087 | 0.533 * | 0.082 |
| Growth Parameters | C in Fine Root | N in Fine Root | P in Fine Root | C in Leaf | N in Leaf | P in Leaf | C in Branch | N in Branch | P in Branch |
|---|---|---|---|---|---|---|---|---|---|
| Height | 0.053 | 0.785 ** | 0.767 ** | −0.534 * | −0.080 | −0.662 ** | −0.248 | −0.115 | −0.134 |
| Crown area | 0.045 | 0.697 ** | 0.746 ** | −0.629 * | −0.087 | −0.687 ** | −0.262 | −0.185 | −0.066 |
| Basal diameter | −0.201 | 0.469 | 0.627 * | −0.594 * | −0.314 | −0.699 ** | −0.051 | −0.196 | 0.188 |
| DBH | −0.080 | 0.457 | 0.549 * | −0.427 | −0.188 | −0.620 * | −0.198 | −0.094 | 0.032 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, D.; Liu, M.; Arif, M.; Yuan, Z.; Li, J.; Hu, X.; Zheng, J.; Li, C. Responses of Ecological Stoichiometric Characteristics of Carbon, Nitrogen, and Phosphorus to Periodic Submergence in Mega-Reservoir: Growth of Taxodium distichum and Taxodium ascendens. Plants 2021, 10, 2040. https://doi.org/10.3390/plants10102040
Ding D, Liu M, Arif M, Yuan Z, Li J, Hu X, Zheng J, Li C. Responses of Ecological Stoichiometric Characteristics of Carbon, Nitrogen, and Phosphorus to Periodic Submergence in Mega-Reservoir: Growth of Taxodium distichum and Taxodium ascendens. Plants. 2021; 10(10):2040. https://doi.org/10.3390/plants10102040
Chicago/Turabian StyleDing, Dongdong, Minghui Liu, Muhammad Arif, Zhongxun Yuan, Jiajia Li, Xin Hu, Jie Zheng, and Changxiao Li. 2021. "Responses of Ecological Stoichiometric Characteristics of Carbon, Nitrogen, and Phosphorus to Periodic Submergence in Mega-Reservoir: Growth of Taxodium distichum and Taxodium ascendens" Plants 10, no. 10: 2040. https://doi.org/10.3390/plants10102040