
Proposal to Consider Chemical/Physical Microenvironment as a New Therapeutic Off-Target Approach



1
Department of Environment and Health, Istituto Superiore di Sanità, 00161 Rome, Italy
2
Department of Oncology and Molecular Medicine, Istituto Superiore di Sanità, 00161 Rome, Italy
*
Author to whom correspondence should be addressed.
Pharmaceutics 2022, 14(10), 2084; https://doi.org/10.3390/pharmaceutics14102084
Submission received: 1 August 2022
/
Revised: 26 September 2022
/
Accepted: 28 September 2022
/
Published: 29 September 2022
(This article belongs to the Section Drug Targeting and Design)
Abstract
:The molecular revolution could lead drug discovery from chance observation to the rational design of new classes of drugs that could simultaneously be more effective and less toxic. Unfortunately, we are witnessing some failure in this sense, and the causes of the crisis involve a wide range of epistemological and scientific aspects. In pharmacology, one key point is the crisis of the paradigm the “magic bullet”, which is to design therapies based on specific molecular targets. Drug repurposing is one of the proposed ways out of the crisis and is based on the off-target effects of known drugs. Here, we propose the microenvironment as the ideal place to direct the off-targeting of known drugs. While it has been extensively investigated in tumors, the generation of a harsh microenvironment is also a phenotype of the vast majority of chronic diseases. The hostile microenvironment, on the one hand, reduces the efficacy of both chemical and biological drugs; on the other hand, it dictates a sort of “Darwinian” selection of those cells armed to survive in such hostile conditions. This opens the way to the consideration of the microenvironment as a convenient target for pharmacological action, with a clear example in proton pump inhibitors.
1. Introduction
Notwithstanding the ever-increasing expenditure in pharmacological research and the claimed ever-increasing and detailed knowledge of biological mechanisms, the number of newly marketed drugs and the ratio between new drugs and expenditure have been rapidly falling since the 1980s [1]. Moreover, despite the promises of a “druggable genome” set forth by the rise of the genomic (and post-genomic) era, the majority of newly marketed drugs interact with “old” receptors discovered well before this era [2].
The crisis encompasses a wide range of epistemological issues related to the general crisis of reductionism in science. This crisis, in the case of drug discovery, has its focus on the paradigm of a “magic bullet” capable of interfering with a supposed “critical step” of a largely deterministic chain of events that goes from the molecular to the organismic layer of organization [3].
The feasibility of drug repurposing stems from the presence of a huge “dark side of pharmacology” that involves the off-target (and, in general, unexpected) effects of known drugs coming from the presence of intermingled interaction networks preventing any effort of developing drug molecules endowed with a single molecular mechanism of action [4,5].
In the first part of this article, we briefly set out why a strategy rooted in the quest for the “correct receptor to hit” cannot work in the case of intermingled interaction networks [6], with a particular emphasis on cancer [7,8]. In this section, we outline the essentials of the complex network paradigm and the consequent acquiring of a conscious approach in facing complexity [9]. The network paradigm indicates the emergence of “bio-dynamic interfaces” [10] as the only way to mediate the interaction between complex systems, such as the organism and its environment, at all layers of biological organization. In contrast, the network pharmacology approach is strictly linked to rational drug re-purposing [11] by promoting a process-focused disease description over the usual phenotype-based one.
In the second section of the article, the prevailing role of context (microenvironmental cues and physical forces) with respect to receptor-based mechanistic rules is demonstrated as one of the main drivers of cancer treatment failure [12,13]. Other relevant issues, such as the dependence of the microenvironment on the successes and failures of antibiotic therapies [14] and in the management of gut functionality [15], are also outlined.
The last part of the review is devoted to the analysis of diverse successful cases of drug repurposing spanning different application fields, with a particular emphasis on non-canonical pharmacodynamics [16,17] and newly proposed computational hypothesis testing approaches [18,19,20].
The term microenvironment indicates the entire context that the biological system is involved in, even the chemical/physical forces impinging on (and, in turn, being modified by) the system, and throughout this work we concentrate on these forces. This choice was dictated by both the impossibility of exhaustively taking into consideration all the intricacies of microenvironment factors and of highlighting a still largely unexplored avenue of drug action different from the usual molecular organization layer taken into consideration by pharmacological research.
2. The Network Paradigm
The multi-level organization of nature is self-evident: at the very basic level of biological organization, proteins interact among themselves to give rise to an organized metabolism, while, at the same time, each protein (a single node of such an interaction network) is itself a network of interacting amino acid residues. Similar pictures can be drawn for the structure and function of cells, organs, tissues, and ecological systems. The network paradigm indicates a circular causation pattern in which bottom-up (the perturbation of more basic layers influences higher level layers) and top-down (the global architecture of the network impinges on the behavior of a single node) causation models are both relevant for the same phenomenon.
This evidence is in sharp contrast to the usual way of approaching pharmacology by an exclusive bottom-up quasi-deterministic approach: causally relevant events originate from the lower level (the molecular one) in the form of perturbations that “climb up” the hierarchy, reaching the ultimate layer of macroscopic behavior (e.g., causing a specific disease) [21]. The rising interest in complex network studies allows scientists to further the pure qualitative observation of the existence of both non-linear and non-bottom-up processes, and to uncover the deep nature of multi-level organization. As pointed out by Nicosia et al. [22], “Networks are the fabric of complex systems”. Only a network organization, essentially consisting of a set of nodes (elementary players) linked by edges (mutual interactions), can give rise to the classical attributes of living matter: the presence of multiple equilibrium states, robustness, and the possibility to adapt to an ever-changing environment [23]. In a network system, the single players do not work in isolation: the “passage of scale” from the microscopic level (e.g., single node perturbation) to the global response of the system (e.g., the organism outcome) emerges from the re-arrangement of the entire network in which the activities of the single nodes are influenced by local perturbation. In the majority of cases, this perturbation is buffered by means of the rapid extinction of the initial stimulus [23], which dissipates while spreading across the network. This phenomenon is both the basis of biological system resilience and of the missed promise of a “druggable genome”. Being only a strict minority and, in most cases, already “drugged” in past decades [2], the nodes whose stimulation provokes a persistent perturbation are able to initiate a regime shift of the entire system. The recognition of this fact fostered two new pharmacological lines of research: an approach mimicking allostery, the most evident “global reorganization” of a network system by an effective stimulus [24], and the search for “network drugs” that, instead of targeting a single molecular entity, act as “weak binders” at multiple sites of the interaction network [25].
Biological networks are embedded into an ever-changing microenvironment that, in turn, can be considered a network of mutual interactions among its constituents. This embedding is far from being a uni-directional relationship. Rather, it is a continuous bi-directional accommodation of the microenvironment. In these “microcosmo” cells, vesicles, soluble factors, vessels, and chemical and physical factors crosstalk continuously in an unceasing modification of the biological system. This implies the existence of a “field” encompassing both the biological system and its microenvironment. The concept of a “field” is crucial to catch the essence of considering a microenvironment as a target for drug action. From basic physics, we know that a point charge embedded into an electromagnetic field both “senses” (i.e., is influenced by the field) and modifies (i.e., influences) the field. This is exactly what happens with a microenvironment, in which environmental cues influence the biological system, and, consequently, the biological system modifies its microenvironment (e.g., pH modifications triggered by the Warburg effect and the continuous acidification supported by the H+ extracellular elimination by proton pumps). This mutual interaction is made possible by a shared biodynamic interface that changes in both time and space [10].
3. Microenvironment and Diseases
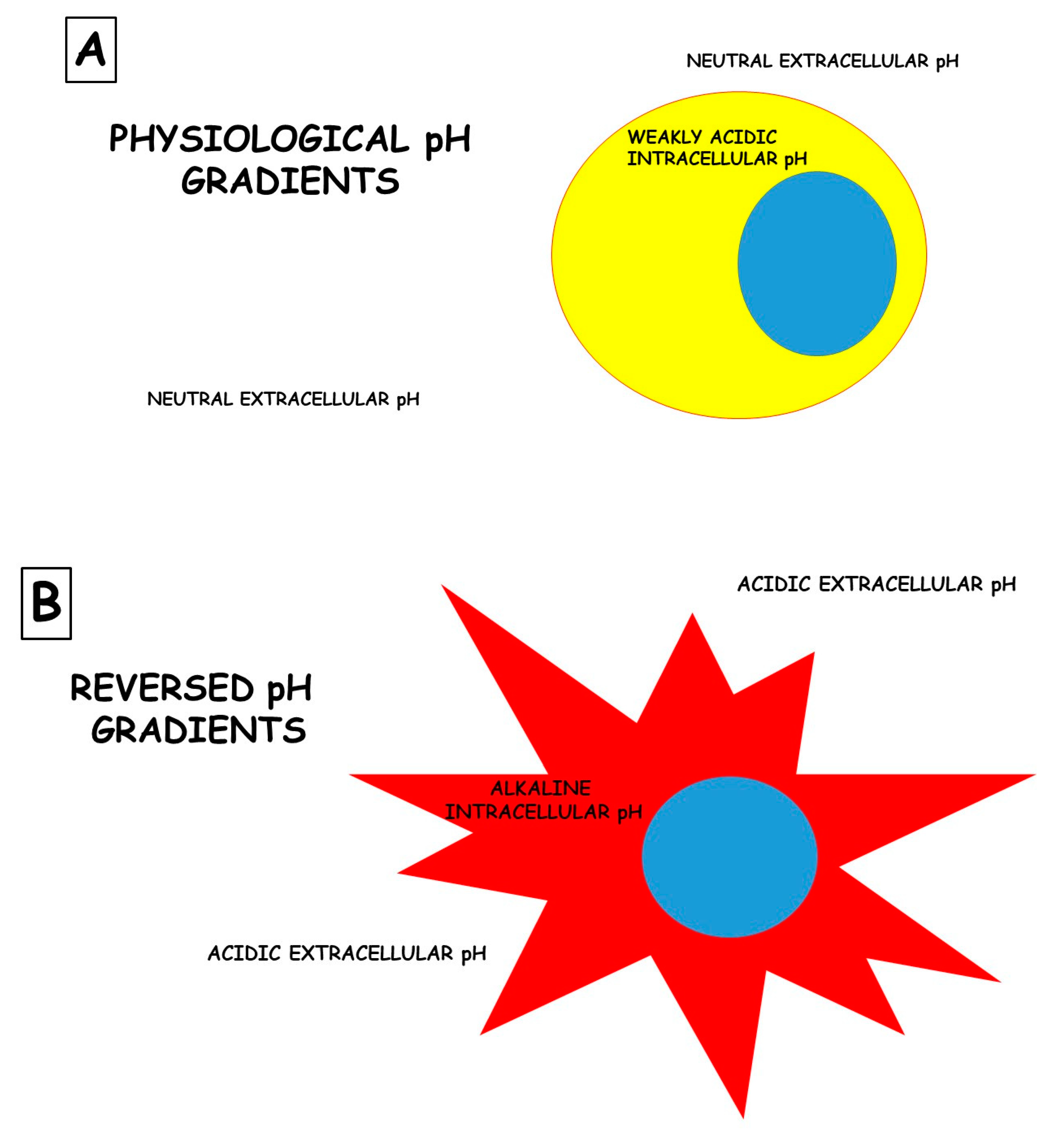
The role of the microenvironment in disease pathogenesis and, most of all, in response to therapies based on drugs is sadly neglected. An example, among many, is cancer. While cancer is commonly described as “a disease of the genes”, it is also associated with massive metabolic re-programming, which is now accepted as a disease hallmark [26,27]. This programming is complex and often involves metabolic cooperativity between cancer cells and their surrounding stroma. Indeed, there is emerging clinical evidence that interrupting a cancer’s metabolic program can improve patients’ outcomes [28,29,30]. The most commonly observed and well-studied metabolic adaptation in cancers is the fermentation of glucose to lactic acid, even in the presence of oxygen, also known as “aerobic glycolysis” or the “Warburg effect”. Much has been written about the mechanisms of the Warburg effect, and this remains a topic of great debate [27,31]. Independently from the mechanism underlying this phenomenon, the ultimate outcome of aerobic glycolysis is the acidification of the tumor microenvironment. Rather than being an epiphenomenon, it is now appreciated that this acidosis is a key player in cancer somatic evolution and progression to malignancy [26,32]. Adaptation to acidosis induces and selects malignant behaviors, such as increased invasion and metastasis, chemoresistance, and the inhibition of immune surveillance. However, the metabolic reprogramming that occurs during the adaptation to acidosis also introduces therapeutic vulnerabilities. Thus, tumor acidosis is a relevant therapeutic target, and there are some reasonable approaches for accomplishing this: (1) neutralizing acid directly with buffers, (2) targeting the metabolic vulnerabilities revealed by acidosis, (3) developing either acid-activatable drugs or acid-sensible nanocarriers, and (4) inhibiting the metabolic processes responsible for generating acids. Several cellular functions are dictated by the pH differences between extracellular and intracellular spaces. These differences are called “pH gradients”, and they are, for instance, crucial for the uptake of chemical drugs by target cells. Figure 1 illustrates a rudimentary portrait of this phenomenon.
However, some strategies have been used to exploit these potentially very powerful approaches [26,33]. These strategies include (i) the use of simple buffers, such as sodium bicarbonate; (ii) more complex buffers, such as some mixes of carbonates and bicarbonates (e.g., Basenpulver); and (iii) the use of a series of inhibitors of ion/proton exchangers. The vast majority of the studies involving either buffers or exchange inhibitors are confined to pre-clinical studies, but proton pump inhibitors (PPIs), which are being extensively used worldwide as anti-acidic drugs, have been combined with chemotherapy in some clinical trials. The results of both pre-clinical and clinical studies have convinced scientists to think about the re-purposing of these classes of drugs as anti-tumor drugs [33,34].
Epidemiological studies have shown that numerous risk factors are shared by diabetes and several cancer sites. Among these primarily are obesity and smoking status, but they also include low physical activity and alcohol consumption. The pathophysiological mechanisms implicated in the association between Type 2 Diabetes (T2D) and cancer have been proposed for colorectal, pancreas, and liver cancers. These include the T2D microenvironment, as represented by advanced glycation end-products; chronic local inflammation; hyperlipidemia; extracellular matrix disorders; and altered microbiota that could predispose the development of colorectal cancer. However, despite the strong epidemiological evidence, the mechanistic bases of the association between diabetes and cancer are still not understood [35].
The major sub-type of T2D is peripheral insulin resistance associated with obesity and central adiposity, leading to hyperinsulinemia and chronic inflammation, both of which have the potential to exacerbate the risk of cancer. Hyperinsulinemia, together with hyperglycemia, also contributes to the accumulation of keto acids, leading to chronic systemic acidosis, which is compensated for by reducing HCO3 and by reducing the metabolic interstitial buffering capacity, making interstitial pH more fragile. Hyperinsulinemia is also associated with increased circulating levels of insulin-like growth factor-1 (IGF1), which is a potent mitogenic factor for neoplastic epithelial cells. The binding of IGF1 to its receptor triggers the activation of the PI3K➔Akt➔mTOR pathway, inducing metabolic activation and mitogenesis [35].
What cancer and diabetes have in common is the increased reliance on glucose fermentation. Continuous glucose fermentation leads to lactate production and significant local acidosis in both diabetic peripheral tissues and tumors. Acidosis is exacerbated if combined with decreased perfusion, which can be a consequence of inflammation, peripheral vascular resistance, and dysangiogenesis, all common syndromes in cancer and diabetes. There is significant evidence, presented below, that this local acidosis in cancer can promote tissue re-modeling, local invasion, metastasis, and the inhibition of immune surveillance. In diabetes, local and systemic acidosis reduces insulin’s affinity for its receptor, exacerbating the spiral of peripheral insulin resistance. Consequently, targeting acidosis is an important therapeutic approach in both T2D and cancer, as discussed below.
The same principles hold true for another major health treatment: the rising resistance to antibiotic therapy. In [14], the authors explicitly take into account the consideration of a microenvironment as a target of antibiotic treatment in terms of the modification of microenvironmental cues so as to inhibit the emergence of biofilms, organized colonies of bacterial cells, which are very difficult to eradicate with antibiotics. It is worth noting that antibiotic efficacy is routinely assessed by antibiograms, in which the sensitivity of the bacterial strains responsible for an infection in a given patient is tested in vitro. The puzzle resides in the fact that bacteria behave substantially differently in standard laboratory conditions from actual infections. The infectious microenvironment imposes changes in growth and metabolic activity that result in increased protection against antibiotics (e.g., biofilm production). Therefore, an improved antibiotic treatment of chronic infections is achievable when antibiotics are recommended based on susceptibility testing in relevant in vitro conditions that resemble actual infectious microenvironments, with a particular focus on fostering direct interventions on the microenvironment that could prevent biofilm formation.
Along a similar vein (after all, we are dealing with ecological systems, and, again, we can safely consider cancer as an ecological threat with a species going out of control), we can posit direct intervention on the microenvironment of microbiota [15]. The microbiome can be considered a complex biocenosis stemming from a network of interactions between thousands of bacterial and yeast species. It is not by chance that microbiome research is based on the same computational and theoretical principles as ecology [36]. The microbiome is involved in the creation of complex biodynamic interfaces with host tissues, e.g., the ability to colonize and thrive within the mucous layer that covers the colon epithelium. These mucosal microbes intimately interact with the intestinal tissue and are important modulators of human health. Embedded in the host-secreted mucous matrix, they form a “mucosal biofilm” with a distinct composition and functionality that, in turn, shapes the mucosal microenvironment. This implies the need to actively consider the mucosal microenvironment as a target for any therapeutic intervention for re-establishing the correct microbial ecology.
4. Rethinking off-Targeting for Treating Microenvironment—An Eco-evolutionary Way of Thinking
In dealing with microenvironments, tumors represent an emblematic issue. As introduced above, tumors survive in extreme conditions due to the upregulation of a series of proton extrusion pumps [34], which release protons and lactate into the extracellular environment; this avoids the acidification of the cytosol, which inevitably kills any cells. Among the proton flux regulators are vacuolar H+-ATPases (V-ATPases), Na+/H+ exchangers (NHE), monocarboxylate transporters (MCTs), carbonic anhydrase IX (CA-IX), and Na+/HCO3 co-transporters (NBCs) [34]. However, it is a common belief that this phenomenon is not the result of transformation but rather the result of “cell clone selection”. In fact, uncontrolled growth, lactic and carbonic acid production, and low blood and nutrient supply contribute to the generation of a tumor microenvironment that is extremely toxic for either normal or more differentiated cells, thus progressively selecting cells capable of surviving in these adverse conditions. This phenomenon occurs independently from the tumor histotype.
Notably, normal cells at a pH ranging from very acidic to weakly acidic inevitably die or are entirely blocked in their functions [35].
For these reasons, a therapy based on a combination of existing proton or ion efflux pump inhibitors is under investigation in order to determine whether the implementation of molecules targeting several mechanisms underlying tumor acidification may have a significant effect on tumors. To do this, a series of drugs, whose main target is not tumors, have been considered. This list includes (i) proton pump inhibitors (Lansoprazole, Omeprazole, Esomeprazole, Rabeprazole, and Pantoprazole), whose main purpose is to act as an anti-acidic treatment for gastroprotection; (ii) carbonic anhydrase inhibitors (Acetazolamide), whose main target is glaucoma; (iii) inhibitors of Na+/H+ exchangers, including Amiloride, whose main purpose is to act as anti-hypertensive treatment for their diuretic activity, and Cariporide, whose main target is myocardial ischemia; (iv) monocarboxylate transporters inhibitors with multiple indications, such as the prevention of cardiac graft rejection; and (v) Na+/HCO3 co-transporters inhibitors thought to counteract heart failure.
However, at least as far as tumors are concerned, there is a general hypothesis supporting the progressive establishment of a sort of clonal cell selection induced by the setting of a very hostile microenvironment. This has also been called the “eco-evolutionary theory” of tumors, and it is considered a major actor in dictating the progressive isolation of growing tumors from the rest of the body [37], as well as in leading to an unresponsiveness to therapies [38]. One piece of evidence supporting this hypothesis is the discovery of cannibalism and, more generally, the “cell-in-cell” phenomena in cancers [39], witnessing a sort of primeval behavior in malignant tumor cells. It must also be considered that cannibalism is an almost exclusive phenotype of cells deriving from metastatic lesions [40]. This introduces a new concept in cancer therapy: to cure the microenvironment by re-tuning the eco-evolutionary selection of very malignant cells.
In a similar vein, the targeting of microenvironments has been invoked to deal with bacterial biofilms [41]. Inorganic nanoparticles with intrinsic antibacterial activity and inert nanoparticles assisted by external stimuli, including heat, photons, magnetism, or sound, have been found to be efficient against persistent infections [42]. These strategies are explicitly designed to target the unique microenvironment of bacterial infections.
Off-target experimentation is strictly connected to the consideration of a microenvironment as a proper therapeutic “target”. Here, we wish to stress the fact that this “target” is substantially different to a receptor molecular entity (e.g., there is no specific receptor to modify pH) and requires the abandoning of a strict reductionist approach to pharmacology.
5. Discussion
Here, we wish to propose a new paradigm in pharmacology that puts together the following well-established and recognized issues: (i) many known drugs have off-targets that very often may be recognized by their side effects [40]; (ii) changes in the extracellular microenvironment may represent one phenotype common to many diseases [34]; (iii) some features of the microenvironment may well represent a target for some known drugs, as has been shown for PPIs [34]; and (iv) the example of proton pump inhibitors should be taken into careful consideration when designing new drugs. In fact, as has been extensively shown for tumors, the vast majority of drugs once ending in a H+-enriched microenvironment are protonated and blocked outside tumor cells. PPIs are administered as prodrugs that are transformed in their active molecule (tetracyclic sulphenamide) only following protonation, and both pre-clinical and clinical evidence suggests that they are extremely active in the tumor microenvironment [34]. In fact, both pre-clinical and clinical investigations have shown that pre-treatment with PPIs allows other drugs, which are mostly ineffective in single therapies, to work [43].
To further emphasize the importance of one of the major microenvironmental phenotypes (i.e., extracellular acidosis) in conditioning the efficacy of past and new therapies, it has been shown that the same cell lines cultured at either a “physiological” (7,4) or acidic (6,5) pH display a very different lipid composition of the cell’s plasma membrane [44], which, together with increasing exosome release [12], reducing chemical drug efficacy [45], and inhibiting immune reaction against tumors [46], make tumor microenvironmental acidity a major determinant in anti-tumor therapy failure.
However, as is clearly emphasized in this article, a harsh microenvironment is a major determinant in the pathogenesis of many diseases [35,47], strongly supporting a new era in which the microenvironment may well represent a major target for new therapeutic molecules, while also considering the off-targeting of old drugs. Scientific evidence supports the role of acidity in impairing the immune response. In fact, systemic buffering improves the immune reaction against tumors [48]. However, the solid literature supports, either directly or indirectly, a key role of microenvironmental acidity not only in tumors but also in a series of pathological conditions involving metabolism, the cardiovascular system, the nervous system, infectious diseases, inflammation and the immune system, renal function, pain, and other pathological conditions. Table 1 summarizes these data with the appropriate references. However, some reviews have emphasized the importance of acidity in tumors, proposing some approaches to counteract it [26,27,49]. Many of the purposes behind newly treated microenvironmental acidity come from the new paradigm of using the off-targeting of old drugs [5], but some interesting and novel approaches to finding new anti-cancer drugs in the panel of available drugs have recently been proposed [50], thus opening a promising future for this issue. Lastly, two recent papers have shown that an anti-acidic treatment may reduce the risk of lung cancer [51] and that urine acidity is associated with poor prognosis in advanced bladder cancers [52], further suggesting that targeting microenvironmental acidity may represent a future strategy in both the prevention and treatment of diseases.
Funding
This article was supported by funds from the Italian Ministry of Health.
Acknowledgments
We thank Rossella Di Raimo for her assistance in the final editing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Scannell, J.W.; Blanckley, A.; Boldon, H.; Warrington, B. Diagnosing the decline in pharmaceutical R&D efficiency. Nat. Rev. Drug Discov. 2012, 11, 191–200. [Google Scholar] [CrossRef]
- Overington, J.P.; Al-Lazikani, B.; Hopkins, A.L. How many drug targets are there? Nat. Rev. Drug Discov. 2006, 5, 993–996. [Google Scholar] [CrossRef]
- Noble, R.; Tasaki, K.; Noble, P.J.; Noble, D. Biological Relativity Requires Circular Causality but Not Symmetry of Causation: So, Where, What and When Are the Boundaries? Front. Physiol. 2019, 10, 827. [Google Scholar] [CrossRef]
- Li, Z.; He, P.; Long, Y.; Yuan, G.; Shen, W.; Chen, Z.; Zhang, B.; Wang, Y.; Yue, D.; Seidl, C.; et al. Drug Repurposing of Pantoprazole and Vitamin C Targeting Tumor Microenvironment Conditions Improves Anticancer Effect in Metastatic Castration-Resistant Prostate Cancer. Front. Oncol. 2021, 11, 660320. [Google Scholar] [CrossRef]
- Campillos, M.; Kuhn, M.; Gavin, A.-C.; Jensen, L.J.; Bork, P. Drug Target Identification Using Side-Effect Similarity. Science 2008, 321, 263–266. [Google Scholar] [CrossRef]
- Tun, K.; Menghini, M.; D’Andrea, L.; Dhar, P.; Tanaka, H.; Giuliani, A. Why so Few Drug Targets: A Mathematical Explanation? CAD 2011, 7, 206–213. [Google Scholar] [CrossRef]
- Joyner, M.J.; Paneth, N.; Ioannidis, J.P.A. What Happens When Underperforming Big Ideas in Research Become Entrenched? JAMA 2016, 316, 1355–1356. [Google Scholar] [CrossRef]
- Baker, S.G. A Cancer Theory Kerfuffle Can Lead to New Lines of Research. JNCI J. Natl. Cancer Inst. 2014, 107, dju405. [Google Scholar] [CrossRef]
- Csermely, P.; Korcsmáros, T.; Kiss, H.J.M.; London, G.; Nussinov, R. Structure and dynamics of molecular networks: A novel paradigm of drug discovery. Pharmacol. Ther. 2013, 138, 333–408. [Google Scholar] [CrossRef]
- Arora, M.; Giuliani, A.; Curtin, P. Biodynamic Interfaces Are Essential for Human–Environment Interactions. BioEssays 2020, 42, e2000017. [Google Scholar] [CrossRef]
- Nogales, C.; Mamdouh, Z.M.; List, M.; Kiel, C.; Casas, A.I.; Schmidt, H.H. Network pharmacology: Curing causal mechanisms instead of treating symptoms. Trends Pharmacol. Sci. 2021, 43, 136–150. [Google Scholar] [CrossRef]
- Logozzi, M.; Mizzoni, D.; Angelini, D.; Di Raimo, R.; Falchi, M.; Battistini, L.; Fais, S. Microenvironmental pH and Exosome Levels Interplay in Human Cancer Cell Lines of Different Histotypes. Cancers 2018, 10, 370. [Google Scholar] [CrossRef]
- Choudhury, A.R.; Gupta, S.; Chaturvedi, P.K.; Kumar, N.; Pandey, D. Mechanobiology of Cancer Stem Cells and Their Niche. Cancer Microenviron. 2019, 12, 17–27. [Google Scholar] [CrossRef]
- Bjarnsholt, T.; Whiteley, M.; Rumbaugh, K.P.; Stewart, P.S.; Jensen, P.; Frimodt-Møller, N. The importance of understanding the infectious microenvironment. Lancet Infect. Dis. 2022, 22, e88–e92. [Google Scholar] [CrossRef]
- De Weirdt, R.; Van De Wiele, T. Micromanagement in the gut: Microenvironmental factors govern colon mucosal biofilm structure and functionality. NPJ Biofilms Microbiomes 2015, 1, 15026. [Google Scholar] [CrossRef]
- Bizzarri, M.; Giuliani, A.; Monti, N.; Verna, R.; Pensotti, A.; Cucina, A. Rediscovery of natural compounds acting via multitarget recognition and noncanonical pharmacodynamical actions. Drug Discov. Today 2020, 25, 920–927. [Google Scholar] [CrossRef]
- Fais, S.; Venturi, G.; Gatenby, B. Microenvironmental acidosis in carcinogenesis and metastases: New strategies in prevention and therapy. Cancer Metastasis Rev. 2014, 33, 1095–1108. [Google Scholar] [CrossRef]
- Hopkins, A.L. Network pharmacology. Nat. Biotechnol. 2007, 25, 1110–1111. [Google Scholar] [CrossRef]
- Udrescu, L.; Bogdan, P.; Chiş, A.; Sîrbu, I.O.; Topîrceanu, A.; Văruţ, R.-M.; Udrescu, M. Uncovering New Drug Properties in Target-Based Drug–Drug Similarity Networks. Pharmaceutics 2020, 12, 879. [Google Scholar] [CrossRef]
- An, Q.; Yu, L. A heterogeneous network embedding framework for predicting similarity-based drug-target interactions. Brief. Bioinform. 2021, 22, bbab275. [Google Scholar] [CrossRef]
- Uversky, V.N.; Giuliani, A. Networks of Networks: An Essay on Multi-Level Biological Organization. Front. Genet. 2021, 12, 706260. [Google Scholar] [CrossRef]
- Nicosia, V.; De Domenico, M.; Latora, V. Characteristic exponents of complex networks. Eur. Lett. 2014, 106, 58005. [Google Scholar] [CrossRef]
- Liu, X.; Li, D.; Ma, M.; Szymanski, B.K.; Stanley, H.E.; Gao, J. Network resilience. Phys. Rep. 2022, 971, 1–108. [Google Scholar] [CrossRef]
- Wagner, J.R.; Lee, C.T.; Durrant, J.D.; Malmstrom, R.D.; Feher, V.A.; Amaro, R.E. Emerging Computational Methods for the Rational Discovery of Allosteric Drugs. Chem. Rev. 2016, 116, 6370–6390. [Google Scholar] [CrossRef]
- Csermely, P.; Agoston, V.; Pongor, S. The efficiency of multi-target drugs: The network approach might help drug design. Trends Pharmacol. Sci. 2005, 26, 178–182. [Google Scholar] [CrossRef]
- Pillai, S.R.; Damaghi, M.; Marunaka, Y.; Spugnini, E.P.; Fais, S.; Gillies, R.J. Causes, consequences, and therapy of tumors acidosis. Cancer Metastasis Rev. 2019, 38, 205–222. [Google Scholar] [CrossRef]
- Schwartz, L.; Seyfried, T.; Alfarouk, K.O.; Da Veiga Moreira, J.; Fais, S. Out of Warburg effect: An effective cancer treatment targeting the tumor specific metabolism and dysregulated pH. Semin. Cancer Biol. 2017, 43, 134–138. [Google Scholar] [CrossRef]
- Ferrari, S.; Perut, F.; Fagioli, F.; Del Prever, A.B.; Meazza, C.; Parafioriti, A.; Picci, P.; Gambarotti, M.; Avnet, S.; Baldini, N.; et al. Proton pump inhibitor chemosensitization in human osteosarcoma: From the bench to the patients’ bed. J. Transl. Med. 2013, 11, 268. [Google Scholar] [CrossRef]
- Wang, B.-Y.; Zhang, J.; Wang, J.-L.; Sun, S.; Wang, Z.-H.; Wang, L.-P.; Zhang, Q.-L.; Lv, F.-F.; Cao, E.-Y.; Shao, Z.-M.; et al. Intermittent high dose proton pump inhibitor enhances the antitumor effects of chemotherapy in metastatic breast cancer. J. Exp. Clin. Cancer Res. 2015, 34, 85. [Google Scholar] [CrossRef]
- Falcone, R.; Roberto, M.; D’Antonio, C.; Romiti, A.; Milano, A.; Onesti, C.E.; Marchetti, P.; Fais, S. High-doses of proton pump inhibitors in refractory gastro-intestinal cancer: A case series and the state of art. Dig. Liver Dis. 2016, 48, 1503–1505. [Google Scholar] [CrossRef]
- Warburg, O. On the Origin of Cancer Cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef]
- Gatenby, R.A.; Gawlinski, E.T.; Gmitro, A.F.; Kaylor, B.; Gillies, R.J. Acid-Mediated Tumor Invasion: A Multidisciplinary Study. Cancer Res. 2006, 66, 5216–5223. [Google Scholar] [CrossRef]
- Spugnini, E.P.; Sonveaux, P.; Stock, C.; Sayáns, M.P.; De Milito, A.; Avnet, S.; Garcia-Garcia, A.; Harguindey, S.; Fais, S. Proton channels and exchangers in cancer. Biochim. Biophys. Acta 2015, 1848, 2715–2726. [Google Scholar] [CrossRef] [PubMed]
- Spugnini, E.P.; Fais, S. Drug repurposing for anticancer therapies. A lesson from proton pump inhibitors. Expert Opin. Ther. Pat. 2019, 30, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Gillies, R.J.; Pilot, C.; Marunaka, Y.; Fais, S. Targeting acidity in cancer and diabetes. Biochim. Biophys. Acta Rev. Cancer 2019, 1871, 273–280. [Google Scholar] [CrossRef] [PubMed]
- McKnight, D.T.; Huerlimann, R.; Bower, D.S.; Schwarzkopf, L.; Alford, R.A.; Zenger, K.R. Methods for normalizing microbiome data: An ecological perspective. Methods Ecol. Evol. 2018, 10, 389–400. [Google Scholar] [CrossRef]
- Gillies, R.J.; Brown, J.S.; Anderson, A.R.A.; Gatenby, R.A. Eco-evolutionary causes and consequences of temporal changes in intratumoural blood flow. Nat. Rev. Cancer 2018, 18, 576–585. [Google Scholar] [CrossRef]
- Gillies, R.; Verduzco, D.; Gatenby, R.A. Evolutionary dynamics of carcinogenesis and why targeted therapy does not work. Nat. Rev. Cancer 2012, 12, 487–493. [Google Scholar] [CrossRef]
- Fais, S.; Overholtzer, M. Cell-in-cell phenomena in cancer. Nat. Rev. Cancer 2018, 18, 758–766. [Google Scholar] [CrossRef]
- Lugini, L.; Matarrese, P.; Tinari, A.; Lozupone, F.; Federici, C.; Iessi, E.; Gentile, M.; Luciani, F.; Parmiani, G.; Rivoltini, L.; et al. Cannibalism of Live Lymphocytes by Human Metastatic but Not Primary Melanoma Cells. Cancer Res. 2006, 66, 3629–3638. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Wang, J.; Chai, M.; Li, X.; Deng, Y.; Jin, Q.; Ji, J. Size and Charge Adaptive Clustered Nanoparticles Targeting the Biofilm Microenvironment for Chronic Lung Infection Management. ACS Nano 2020, 14, 5686–5699. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, X.; Duan, Y.; Huang, Y. Infection microenvironment-related antibacterial nanotherapeutic strategies. Biomaterials 2022, 280, 121249. [Google Scholar] [CrossRef] [PubMed]
- Fais, S. Evidence-based support for the use of proton pump inhibitors in cancer therapy. J. Transl. Med. 2015, 13, 368. [Google Scholar] [CrossRef] [PubMed]
- Urbanelli, L.; Buratta, S.; Logozzi, M.; Mitro, N.; Sagini, K.; Di Raimo, R.; Caruso, D.; Fais, S.; Emiliani, C. Lipidomic analysis of cancer cells cultivated at acidic pH reveals phospholipid fatty acids remodelling associated with transcriptional reprogramming. J. Enzym. Inhib. Med. Chem. 2020, 35, 963–973. [Google Scholar] [CrossRef]
- Luciani, F.; Spada, M.; De Milito, A.; Molinari, A.; Rivoltini, L.; Montinaro, A.; Marra, M.; Lugini, L.; Logozzi, M.; Lozupone, F.; et al. Effect of Proton Pump Inhibitor Pretreatment on Resistance of Solid Tumors to Cytotoxic Drugs. JNCI J. Natl. Cancer Inst. 2004, 96, 1702–1713. [Google Scholar] [CrossRef]
- Calcinotto, A.; Filipazzi, P.; Grioni, M.; Iero, M.; De Milito, A.; Ricupito, A.; Cova, A.; Canese, R.; Jachetti, E.; Rossetti, M.; et al. Modulation of Microenvironment Acidity Reverses Anergy in Human and Murine Tumor-Infiltrating T Lymphocytes. Cancer Res. 2012, 72, 2746–2756. [Google Scholar] [CrossRef]
- Fais, S.; Marunaka, Y. The Acidic Microenvironment: Is It a Phenotype of All Cancers? A Focus on Multiple Myeloma and Some Analogies with Diabetes Mellitus. Cancers 2020, 12, 3226. [Google Scholar] [CrossRef]
- Pilon-Thomas, S.; Kodumudi, K.N.; El-Kenawi, A.E.; Russell, S.; Weber, A.M.; Luddy, K.; Damaghi, M.; Wojtkowiak, J.W.; Mulé, J.J.; Ibrahim-Hashim, A.; et al. Neutralization of Tumor Acidity Improves Antitumor Responses to Immunotherapy. Cancer Res. 2016, 76, 1381–1390, Erratum in Cancer Res. 2017, 77, 2552. [Google Scholar] [CrossRef]
- Koltai, T. Targeting the pH Paradigm at the Bedside: A Practical Approach. Int. J. Mol. Sci. 2020, 21, 9221. [Google Scholar] [CrossRef]
- Wu, Y.; Warner, J.L.; Wang, L.; Jiang, M.; Xu, J.; Chen, Q.; Nian, H.; Dai, Q.; Du, X.; Yang, P.; et al. Discovery of Noncancer Drug Effects on Survival in Electronic Health Records of Patients With Cancer: A New Paradigm for Drug Repurposing. JCO Clin. Cancer Inform. 2019, 3, 1–9. [Google Scholar] [CrossRef]
- Go, S.; Lee, D.Y.; Choi, W.I.; Jeong, J. Association between use of antacid medications (proton pump inhibitors and histamine-2 receptor antagonists) and the incidence of lung cancer: A population-based cohort analysis. Medicine 2022, 101, e30399. [Google Scholar] [CrossRef]
- Han, J.H.; Jeong, S.H.; Yuk, H.D.; Jeong, C.W.; Kwak, C.; Ku, J.H. Acidic urine is associated with poor prognosis in patients with bladder cancer undergoing radical cystectomy. Front. Oncol. 2022, 12, 964571. [Google Scholar] [CrossRef] [PubMed]
- Rofstad, E.K.; Mathiesen, B.; Kindem, K.; Galappathi, K. Acidic Extracellular pH Promotes Experimental Metastasis of Human Melanoma Cells in Athymic Nude Mice. Cancer Res. 2006, 66, 6699–6707. [Google Scholar] [CrossRef] [PubMed]
- Azzarito, T.; Venturi, G.; Cesolini, A.; Fais, S. Lansoprazole induces sensitivity to suboptimal doses of paclitaxel in human melanoma. Cancer Lett. 2015, 356, 697–703. [Google Scholar] [CrossRef] [PubMed]
- De Milito, A.; Canese, R.; Marino, M.L.; Borghi, M.; Iero, M.; Villa, A.; Venturi, G.; Lozupone, F.; Iessi, E.; Logozzi, M.; et al. pH-dependent antitumor activity of proton pump inhibitors against human melanoma is mediated by inhibition of tumor acidity. Int. J. Cancer 2009, 127, 207–219. [Google Scholar] [CrossRef]
- Federici, C.; Lugini, L.; Marino, M.L.; Carta, F.; Iessi, E.; Azzarito, T.; Supuran, C.T.; Fais, S. Lansoprazole and carbonic anhydrase IX inhibitors sinergize against human melanoma cells. J. Enzym. Inhib. Med. Chem. 2016, 31, 119–125. [Google Scholar] [CrossRef]
- Federici, C.; Petrucci, F.; Caimi, S.; Cesolini, A.; Logozzi, M.; Borghi, M.; D’Ilio, S.; Lugini, L.; Violante, N.; Azzarito, T.; et al. Exosome release and low pH belong to a framework of resistance of human melanoma cells to cisplatin. PLoS ONE 2014, 9, e88193. [Google Scholar] [CrossRef]
- Moellering, R.; Black, K.C.; Krishnamurty, C.; Baggett, B.K.; Stafford, P.; Rain, M.; Gatenby, R.A.; Gillies, R.J. Acid treatment of melanoma cells selects for invasive phenotypes. Clin. Exp. Metastasis 2008, 25, 411–425. [Google Scholar] [CrossRef]
- Martinez-Zaguilan, R.; Seftor, E.A.; Seftor, R.E.B.; Chu, Y.-W.; Gillies, R.J.; Hendrix, M.J.C. Acidic pH enhances the invasive behavior of human melanoma cells. Clin. Exp. Metastasis 1996, 14, 176–186. [Google Scholar] [CrossRef]
- De Milito, A.; Iessi, E.; Logozzi, M.; Lozupone, F.; Spada, M.; Marino, M.L.; Federici, C.; Perdicchio, M.; Matarrese, P.; Lugini, L.; et al. Proton Pump Inhibitors Induce Apoptosis of Human B-Cell Tumors through a Caspase-Independent Mechanism Involving Reactive Oxygen Species. Cancer Res. 2007, 67, 5408–5417. [Google Scholar] [CrossRef] [Green Version]
- Nakanishi, M.; Morita, Y.; Hata, K.; Muragaki, Y. Acidic microenvironments induce lymphangiogenesis and IL-8 production via TRPV1 activation in human lymphatic endothelial cells. Exp. Cell Res. 2016, 345, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Spugnini, E.P.; Baldi, A.; Buglioni, S.; Carocci, F.; de Bazzichini, G.M.; Betti, G.; Pantaleo, I.; Menicagli, F.; Citro, G.; Fais, S. Lansoprazole as a rescue agent in chemoresistant tumors: A phase I/II study in companion animals with spontaneously occurring tumors. J. Transl. Med. 2011, 9, 221. [Google Scholar] [CrossRef] [PubMed]
- Spugnini, E.P.; Buglioni, S.; Carocci, F.; Francesco, M.; Vincenzi, B.; Fanciulli, M.; Fais, S. High dose lansoprazole combined with metronomic chemotherapy: A phase I/II study in companion animals with spontaneously occurring tumors. J. Transl. Med. 2014, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Li, M.-H.; Liu, J.; Huang, N.; Li, N.; Liu, S.; Liu, Y.; Zhang, T.; Zou, Q.; Li, H. Benzimidazole derivative, BMT-1, induces apoptosis in multiple myeloma cells via a mitochondrial-mediated pathway involving H+/K+-ATPase inhibition. Oncol. Rep. 2014, 31, 2743–2750. [Google Scholar] [CrossRef] [PubMed]
- Canitano, A.; Iessi, E.; Spugnini, E.P.; Federici, C.; Fais, S. Proton pump inhibitors induce a caspase-independent antitumor effect against human multiple myeloma. Cancer Lett. 2016, 376, 278–283. [Google Scholar] [CrossRef]
- Zhao, M.; Liu, Q.; Gong, Y.; Xu, X.; Zhang, C.; Liu, X.; Zhang, C.; Guo, H.; Zhang, X.; Gong, Y.; et al. GSH-dependent antioxidant defense contributes to the acclimation of colon cancer cells to acidic microenvironment. Cell Cycle 2016, 15, 1125–1133. [Google Scholar] [CrossRef]
- Lozupone, F.; Borghi, M.; Marzoli, F.; Azzarito, T.; Matarrese, P.; Iessi, E.; Venturi, G.; Meschini, S.; Canitano, A.; Bona, R.; et al. TM9SF4 is a novel V-ATPase-interacting protein that modulates tumor pH alterations associated with drug resistance and invasiveness of colon cancer cells. Oncogene 2015, 34, 4482–4490. [Google Scholar] [CrossRef]
- Fan, S.; Niu, Y.; Tan, N.; Wu, Z.; Wang, Y.; You, H.; Ke, R.; Song, J.; Shen, Q.; Wang, W.; et al. LASS2 enhances chemosensitivity of breast cancer by counteracting acidic tumor microenvironment through inhibiting activity of V-ATPase proton pump. Oncogene 2013, 32, 1682–1690. [Google Scholar] [CrossRef]
- Glunde, K.; Guggino, S.E.; Solaiyappan, M.; Pathak, A.P.; Ichikawa, Y.; Bhujwalla, Z.M. Extracellular Acidification Alters Lysosomal Trafficking in Human Breast Cancer Cells. Neoplasia 2003, 5, 533–545. [Google Scholar] [CrossRef]
- Udelnow, A.; Kreyes, A.; Ellinger, S.; Landfester, K.; Walther, P.; Klapperstueck, T.; Wohlrab, J.; Henne-Bruns, D.; Knippschild, U.; Würl, P. Omeprazole Inhibits Proliferation and Modulates Autophagy in Pancreatic Cancer Cells. PLoS ONE 2011, 6, e20143. [Google Scholar] [CrossRef] [Green Version]
- Jin, U.-H.; Lee, S.-O.; Pfent, C.; Safe, S. The aryl hydrocarbon receptor ligand omeprazole inhibits breast cancer cell invasion and metastasis. BMC Cancer 2014, 14, 498. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Zaguilán, R.; Raghunand, N.; Lynch, R.M.; Bellamy, W.; Martinez, G.M.; Rojas, B.; Smith, D.; Dalton, W.S.; Gillies, R.J. pH and drug resistance. I. functional expression of plasmalemmal V-type H+-ATPase in drug-resistant human breast carcinoma cell lines. Biochem. Pharmacol. 1999, 57, 1037–1046. [Google Scholar] [CrossRef]
- Robey, I.F.; Martin, N.K. Bicarbonate and dichloroacetate: Evaluating pH altering therapies in a mouse model for metastatic breast cancer. BMC Cancer 2011, 11, 235. [Google Scholar] [CrossRef]
- You, H.; Jin, J.; Shu, H.; Yu, B.; De Milito, A.; Lozupone, F.; Deng, Y.; Tang, N.; Yao, G.; Fais, S.; et al. Small interfering RNA targeting the subunit ATP6L of proton pump V-ATPase overcomes chemoresistance of breast cancer cells. Cancer Lett. 2009, 280, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, Y.; Li, S.J. Lansoprazole induces apoptosis of breast cancer cells through inhibition of intracellular proton extrusion. Biochem. Biophys. Res. Commun. 2014, 448, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-Y.; Jeon, H.-K.; Hong, J.E.; Cho, Y.J.; Ryu, J.Y.; Choi, J.-J.; Lee, S.H.; Yoon, G.; Kim, W.Y.; Do, I.-G.; et al. Proton pump inhibitors enhance the effects of cytotoxic agents in chemoresistant epithelial ovarian carcinoma. Oncotarget 2015, 6, 35040–35050. [Google Scholar] [CrossRef] [PubMed]
- Jin, U.-H.; Kim, S.-B.; Safe, S. Omeprazole Inhibits Pancreatic Cancer Cell Invasion through a Nongenomic Aryl Hydrocarbon Receptor Pathway. Chem. Res. Toxicol. 2015, 28, 907–918. [Google Scholar] [CrossRef]
- Shen, Y.; Chen, M.; Huang, S.; Zou, X. Pantoprazole inhibits human gastric adenocarcinoma SGC-7901 cells by downregulating the expression of pyruvate kinase M2. Oncol. Lett. 2015, 11, 717–722. [Google Scholar] [CrossRef]
- Gu, M.; Zhang, Y.; Zhou, X.; Ma, H.; Yao, H.; Ji, F. Rabeprazole exhibits antiproliferative effects on human gastric cancer cell lines. Oncol. Lett. 2014, 8, 1739–1744. [Google Scholar] [CrossRef]
- Zhang, B.; Ling, T.; Zhaxi, P.; Cao, Y.; Qian, L.; Zhao, D.; Kang, W.; Zhang, W.; Wang, L.; Xu, G.; et al. Proton pump inhibitor pantoprazole inhibits gastric cancer metastasis via suppression of telomerase reverse transcriptase gene expression. Cancer Lett. 2019, 452, 23–30. [Google Scholar] [CrossRef]
- Chen, M.; Huang, S.-L.; Zhang, X.-Q.; Zhang, B.; Zhu, H.; Yang, V.W.; Zou, X.-P. Reversal effects of pantoprazole on multidrug resistance in human gastric adenocarcinoma cells by down-regulating the V-ATPases/mTOR/HIF-1α/P-gp and MRP1 signaling pathway in vitro and in vivo. J. Cell. Biochem. 2012, 113, 2474–2487. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Zou, X.; Luo, H.; Cao, J.; Zhang, X.; Zhang, B.; Liu, W. Effects and mechanisms of proton pump inhibitors as a novel chemosensitizer on human gastric adenocarcinoma (SGC7901) cells. Cell Biol. Int. 2009, 33, 1008–1019. [Google Scholar] [CrossRef] [PubMed]
- Yeo, M.; Kim, D.-K.; Kim, Y.-B.; Oh, T.Y.; Lee, J.-E.; Cho, S.W.; Kim, H.C.; Hahm, K.-B. Selective Induction of Apoptosis with Proton Pump Inhibitor in Gastric Cancer Cells. Clin. Cancer Res. 2004, 10, 8687–8696. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Chen, M.; Ding, X.; Zhang, X.; Zou, X. Proton pump inhibitor selectively suppresses proliferation and restores the chemosensitivity of gastric cancer cells by inhibiting STAT3 signaling pathway. Int. Immunopharmacol. 2013, 17, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Sreevathsa, M.R.; Choudhury, A. Long-term survival in a patient with carcinoid syndrome receiving treatment for Zollinger-Ellison syndrome. Int. Surg. 2009, 94, 20–22. [Google Scholar]
- Ibrahim-Hashim, A.; Cornnell, H.H.; Abrahams, D.; Lloyd, M.; Bui, M.; Gillies, R.J.; Gatenby, R.A. Systemic Buffers Inhibit Carcinogenesis in TRAMP Mice. J. Urol. 2012, 188, 624–631. [Google Scholar] [CrossRef]
- Ihling, A.; Ihling, C.H.; Sinz, A.; Gekle, M. Acidosis-Induced Changes in Proteome Patterns of the Prostate Cancer-Derived Tumor Cell Line AT-1. J. Proteome Res. 2015, 14, 3996–4004. [Google Scholar] [CrossRef]
- Lu, X.; Qin, W.; Li, J.; Tan, N.; Pan, D.; Zhang, H.; Xie, L.; Yao, G.; Shu, H.; Yao, M.; et al. The Growth and Metastasis of Human Hepatocellular Carcinoma Xenografts Are Inhibited by Small Interfering RNA Targeting to the Subunit ATP6L of Proton Pump. Cancer Res. 2005, 65, 6843–6849. [Google Scholar] [CrossRef]
- Xu, J.; Xie, R.; Liu, X.; Wen, G.; Jin, H.; Yu, Z.; Jiang, Y.; Zhao, Z.; Yang, Y.; Ji, B.; et al. Expression and functional role of vacuolar H+-ATPase in human hepatocellular carcinoma. Carcinogenesis 2012, 33, 2432–2440. [Google Scholar] [CrossRef]
- Morimura, T.; Fujita, K.; Akita, M.; Nagashima, M.; Satomi, A. The proton pump inhibitor inhibits cell growth and induces apoptosis in human hepatoblastoma. Pediatr. Surg. Int. 2008, 24, 1087–1094. [Google Scholar] [CrossRef]
- Song, J.; Ge, Z.; Yang, X.; Luo, Q.; Wang, C.; You, H.; Ge, T.; Deng, Y.; Lin, H.; Cui, Y.; et al. Hepatic stellate cells activated by acidic tumor microenvironment promote the metastasis of hepatocellular carcinoma via osteopontin. Cancer Lett. 2015, 356, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Kuchuk, O.; Tuccitto, A.; Citterio, D.; Huber, V.; Camisaschi, C.; Milione, M.; Vergani, B.; Villa, A.; Alison, M.R.; Carradori, S.; et al. pH regulators to target the tumor immune microenvironment in human hepatocellular carcinoma. OncoImmunology 2018, 7, e1445452. [Google Scholar] [CrossRef] [PubMed]
- Avnet, S.; Di Pompo, G.; Lemma, S.; Salerno, M.; Perut, F.; Bonuccelli, G.; Granchi, D.; Zini, N.; Baldini, N. V-ATPase is a candidate therapeutic target for Ewing sarcoma. Biochim. Biophys. Acta 2013, 1832, 1105–1116. [Google Scholar] [CrossRef] [PubMed]
- Coman, D.; Huang, Y.; Rao, J.U.; De Feyter, H.M.; Rothman, D.L.; Juchem, C.; Hyder, F. Imaging the intratumoral-peritumoral extracellular pH gradient of gliomas. NMR Biomed. 2016, 29, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Chakhoyan, A.; Nathanson, D.A.; Yong, W.H.; Salamon, N.; Raymond, C.; Mareninov, S.; Lai, A.; Nghiemphu, P.L.; Prins, R.M.; et al. Metabolic characterization of human IDH mutant and wild type gliomas using simultaneous pH- and oxygen-sensitive molecular MRI. Neuro-Oncology 2019, 21, 1184–1196. [Google Scholar] [CrossRef]
- Hebert, K.A.; Bonnen, M.D.; Ghebre, Y.T. Proton pump inhibitors and sensitization of cancer cells to radiation therapy. Front. Oncol. 2022, 12, 937166. [Google Scholar] [CrossRef]
- Hebert, K.A.; Jaramillo, S.; Yu, W.; Wang, M.; Veeramachaneni, R.; Sandulache, V.C.; Sikora, A.G.; Bonnen, M.D.; Annapragada, A.V.; Corry, D.; et al. Esomeprazole enhances the effect of ionizing radiation to improve tumor control. Oncotarget 2021, 12, 1339–1353. [Google Scholar] [CrossRef]
- Hu, R.-M.; Tan, B.-H.; Tester, D.J.; Song, C.; He, Y.; Dovat, S.; Peterson, B.Z.; Ackerman, M.J.; Makielski, J.C. Arrhythmogenic Biophysical Phenotype for SCN5A Mutation S1787N Depends upon Splice Variant Background and Intracellular Acidosis. PLoS ONE 2015, 10, e0124921. [Google Scholar] [CrossRef]
- Demartini, F.E.; Cannon, P.J.; Stason, W.B.; Laragh, J.H. Lactic Acid Metabolism in Hypertensive Patients. Science 1965, 148, 1482–1484. [Google Scholar] [CrossRef]
- Pattharanitima, P.; Thongprayoon, C.; Kaewput, W.; Qureshi, F.; Qureshi, F.; Petnak, T.; Srivali, N.; Gembillo, G.; O’Corragain, O.A.; Chesdachai, S.; et al. Machine Learning Prediction Models for Mortality in Intensive Care Unit Patients with Lactic Acidosis. J. Clin. Med. 2021, 10, 5021. [Google Scholar] [CrossRef]
- Kim, J.; Kim, K.; Park, J.; Jo, Y.H.; Lee, J.H.; Hwang, J.E.; Ha, C.; Ko, Y.-S.; Jung, E. Sodium bicarbonate administration during ongoing resuscitation is associated with increased return of spontaneous circulation. Am. J. Emerg. Med. 2016, 34, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Marunaka, Y. The Proposal of Molecular Mechanisms of Weak Organic Acids Intake-Induced Improvement of Insulin Resistance in Diabetes Mellitus via Elevation of Interstitial Fluid pH. Int. J. Mol. Sci. 2018, 19, 3244. [Google Scholar] [CrossRef] [PubMed]
- Aoi, W.; Marunaka, Y. Importance of pH Homeostasis in Metabolic Health and Diseases: Crucial Role of Membrane Proton Transport. BioMed Res. Int. 2014, 2014, 598986. [Google Scholar] [CrossRef] [PubMed]
- Hayata, H.; Miyazaki, H.; Niisato, N.; Yokoyama, N.; Marunaka, Y. Lowered extracellular pH is involved in the pathogenesis of skeletal muscle insulin resistance. Biochem. Biophys. Res. Commun. 2014, 445, 170–174. [Google Scholar] [CrossRef] [PubMed]
- de Alwis, N.; Binder, N.K.; Mangwiro, Y.T.M.; Beard, S.; Pritchard, N.; Kadife, E.; Fato, B.R.; Keenan, E.; Brownfoot, F.C.; Kaitu’U-Lino, T.J.; et al. Actions of Esomeprazole on the Maternal Vasculature in Lean and Obese Pregnant Mice with Impaired Nitric Oxide Synthesis: A Model of Preeclampsia. Int. J. Mol. Sci. 2022, 23, 8185. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; More, A.S.; Kraut, J.A.; Wu, D. Interaction of Sodium Bicarbonate and Na+/H+ Exchanger Inhibition in the Treatment of Acute Metabolic Acidosis in Pigs. Crit. Care Med. 2015, 43, e160–e169. [Google Scholar] [CrossRef]
- Marunaka, Y. Roles of interstitial fluid pH in diabetes mellitus: Glycolysis and mitochondrial function. World J. Diabetes 2015, 6, 125–135. [Google Scholar] [CrossRef]
- Maalouf, N.M.; Cameron, M.A.; Moe, O.W.; Sakhaee, K. Metabolic Basis for Low Urine pH in Type 2 Diabetes. Clin. J. Am. Soc. Nephrol. 2010, 5, 1277–1281. [Google Scholar] [CrossRef]
- Jiang, N.; Wu, J.; Leng, T.; Yang, T.; Zhou, Y.; Jiang, Q.; Wang, B.; Hu, Y.; Ji, Y.H.; Simon, R.P.; et al. Region specific contribution of ASIC2 to acidosis-and ischemia-induced neuronal injury. J. Cereb. Blood Flow Metab. 2017, 37, 528–540. [Google Scholar] [CrossRef]
- Nedergaard, M.; Goldman, S.A.; Desai, S.; Pulsinelli, W.A. Acid-induced death in neurons and glia. J. Neurosci. 1991, 11, 2489–2497. [Google Scholar] [CrossRef]
- Huang, L.; Zhao, S.; Lu, W.; Guan, S.; Zhu, Y.; Wang, J.-H. Acidosis-Induced Dysfunction of Cortical GABAergic Neurons through Astrocyte-Related Excitotoxicity. PLoS ONE 2015, 10, e0140324. [Google Scholar] [CrossRef] [PubMed]
- Goldman, S.A.; Pulsinelli, W.A.; Clarke, W.Y.; Kraig, R.P.; Plum, F. The Effects of Extracellular Acidosis on Neurons and Glia in vitro. J. Cereb. Blood Flow Metab. 1989, 9, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Cichy, A.; Ackels, T.; Tsitoura, C.; Kahan, A.; Gronloh, N.; Söchtig, M.; Engelhardt, C.H.; Ben-Shaul, Y.; Müller, F.; Spehr, J.; et al. Extracellular pH Regulates Excitability of Vomeronasal Sensory Neurons. J. Neurosci. 2015, 35, 4025–4039. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, F.; Wang, C.; Su, Z. More sensitivity of cortical GABAergic neurons than glutamatergic neurons in response to acidosis. NeuroReport 2016, 27, 610–616. [Google Scholar] [CrossRef]
- Rajamäki, K.; Nordström, T.; Nurmi, K.; Åkerman, K.E.; Kovanen, P.T.; Öörni, K.; Eklund, K.K. Extracellular Acidosis Is a Novel Danger Signal Alerting Innate Immunity via the NLRP3 Inflammasome. J. Biol. Chem. 2013, 288, 13410–13419. [Google Scholar] [CrossRef]
- Wang, Y.-C.; Li, W.-Z.; Wu, Y.; Yin, Y.-Y.; Dong, L.-Y.; Chen, Z.-W.; Wu, W.-N. Acid-sensing ion channel 1a contributes to the effect of extracellular acidosis on NLRP1 inflammasome activation in cortical neurons. J. Neuroinflam. 2015, 12, 1–12. [Google Scholar] [CrossRef]
- Alkhalil, M.; Al-Hiari, Y.; Kasabri, V.; Arabiyat, S.; Al-Zweiri, M.; Mamdooh, N.; Telfah, A. Selected pharmacotherapy agents as antiproliferative and anti-inflammatory compounds. Drug Dev. Res. 2020, 81, 470–490. [Google Scholar] [CrossRef]
- Kong, X.; Tang, X.; Du, W.; Tong, J.; Yan, Y.; Zheng, F.; Fang, M.; Gong, F.; Tan, Z. Extracellular acidosis modulates the endocytosis and maturation of macrophages. Cell. Immunol. 2013, 281, 44–50. [Google Scholar] [CrossRef]
- Minich, D.M.; Bland, J.S. Acid-alkaline balance: Role in chronic disease and detoxification. Altern. Ther. Health Med. 2007, 13, 62–65. [Google Scholar]
- Nakagawa, Y.; Negishi, Y.; Shimizu, M.; Takahashi, M.; Ichikawa, M.; Takahashi, H. Effects of extracellular pH and hypoxia on the function and development of antigen-specific cytotoxic T lymphocytes. Immunol. Lett. 2015, 167, 72–86. [Google Scholar] [CrossRef]
- Gołębiowski, T.; Kusztal, M.; Konieczny, A.; Kuriata-Kordek, M.; Gawryś, A.; Augustyniak-Bartosik, H.; Letachowicz, K.; Zielińska, D.; Wiśniewska, M.; Krajewska, M. Exhausted Capacity of Bicarbonate Buffer in Renal Failure Diagnosed Using Point of Care Analyzer. Diagnostics 2021, 11, 226. [Google Scholar] [CrossRef] [PubMed]
- Woo, Y.-C.; Park, S.S.; Subieta, A.R.; Brennan, T. Changes in Tissue pH and Temperature after Incision Indicate Acidosis May Contribute to Postoperative Pain. Anesthesiology 2004, 101, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.-Z.; Li, H.; Tao, Y.-Q.; Zhou, X.-P.; Yang, Z.-R.; Li, F.-C.; Chen, Q.-X. Systematic review/Meta-analysis The relationship between low pH in intervertebral discs and low back pain: A systematic review. Arch. Med. Sci. 2012, 6, 952–956. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Li, H.; Tao, Y.; Shen, C.; Li, F.; Shi, Z.; Han, B.; Chen, Q. New hypothesis of chronic back pain: Low pH promotes nerve ingrowth into damaged intervertebral disks. Acta Anaesthesiol. Scand. 2013, 57, 271–277. [Google Scholar] [CrossRef]
- Watanabe, S.M.; Ehrlich, L.S.; Strickland, M.; Li, X.; Soloveva, V.; Goff, A.J.; Stauft, C.B.; Bhaduri-McIntosh, S.; Tjandra, N.; Carter, C. Selective Targeting of Virus Replication by Proton Pump Inhibitors. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef]
- Long, J.; Wright, E.; Molesti, E.; Temperton, N.; Barclay, W. Antiviral therapies against Ebola and other emerging viral diseases using existing medicines that block virus entry. F1000Research 2015, 4, 30. [Google Scholar] [CrossRef]
- Madshus, I.H.; Olsnes, S.; Sandvig, K. Mechanism of entry into the cytosol of poliovirus type 1: Requirement for low pH. J. Cell Biol. 1984, 98, 1194–1200. [Google Scholar] [CrossRef]
- Madshus, I.H.; Olsnes, S.; Sandvig, K. Different pH requirements for entry of the two picornaviruses, human rhinovirus 2 and murine encephalomyocarditis virus. Virology 1984, 139, 346–357. [Google Scholar] [CrossRef]
- Touret, F.; Gilles, M.; Barral, K.; Nougairède, A.; van Helden, J.; Decroly, E.; de Lamballerie, X.; Coutard, B. In vitro screening of a FDA approved chemical library reveals potential inhibitors of SARS-CoV-2 replication. Sci. Rep. 2020, 10, 13093. [Google Scholar] [CrossRef]
- Kane, P.M. Proton Transport and pH Control in Fungi. Adv. Exp. Med. Biol. 2016, 892, 33–68. [Google Scholar] [CrossRef]
- Martínez-Muñoz, G.A.; Kane, P. Vacuolar and Plasma Membrane Proton Pumps Collaborate to Achieve Cytosolic pH Homeostasis in Yeast. J. Biol. Chem. 2008, 283, 20309–20319, Erratum in J. Biol. Chem. 2017, 292, 7743. [Google Scholar] [CrossRef] [PubMed]
- Nobile, C.J.; Ennis, C.L.; Hartooni, N.; Johnson, A.D.; Lohse, M.B. A Selective Serotonin Reuptake Inhibitor, a Proton Pump Inhibitor, and Two Calcium Channel Blockers Inhibit Candida albicans Biofilms. Microorganisms 2020, 8, 756. [Google Scholar] [CrossRef] [PubMed]
- da Rosa, T.F.; Foletto, V.S.; Serafin, M.B.; Bottega, A.; Hörner, R. Anti-infective properties of proton pump inhibitors: Perspectives. Int. Microbiol. 2022, 25, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Sjöström, J.E.; Fryklund, J.; Kühler, T.; Larsson, H. In vitro antibacterial activity of omeprazole and its selectivity for Helicobacter spp. are dependent on incubation conditions. Antimicrob. Agents Chemother. 1996, 40, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Huynh, K.K.; Grinstein, S. Regulation of Vacuolar pH and Its Modulation by Some Microbial Species. Microbiol. Mol. Biol. Rev. 2007, 71, 452–462. [Google Scholar] [CrossRef]
- Ferguson, S.J.; Ingledew, W.J. Energetic problems faced by micro-organisms growing or surviving on parsimonious energy sources and at acidic pH: I. Acidithiobacillus ferrooxidans as a paradigm. Biochim. Biophys. Acta 2008, 1777, 1471–1479. [Google Scholar] [CrossRef]
- Puiac, S.; Negrea, A.; Richter-Dahlfors, A.; Plant, L.; Rhen, M. Omeprazole Antagonizes Virulence and Inflammation in Salmonella enterica -Infected RAW264.7 Cells. Antimicrob. Agents Chemother. 2009, 53, 2402–2409. [Google Scholar] [CrossRef]
- Sjøstrøm, J.E.; Kühler, T.; Larsson, H. Basis for the selective antibacterial activity in vitro of proton pump inhibitors against Helicobacter spp. Antimicrob. Agents Chemother. 1997, 41, 1797–1801. [Google Scholar] [CrossRef]
- Evans, C.E.; Peng, Y.; Zhu, M.M.; Dai, Z.; Zhang, X.; Zhao, Y.-Y. Rabeprazole Promotes Vascular Repair and Resolution of Sepsis-Induced Inflammatory Lung Injury through HIF-1α. Cells 2022, 11, 1425. [Google Scholar] [CrossRef]
- Moriyama, Y.; Hayashi, M.; Yatsushiro, S.; Yamamoto, A. Vacuolar proton pumps in malaria parasite cells. J. Bioenerg. Biomembr. 2003, 35, 367–375. [Google Scholar] [CrossRef]
- Hayashi, M.; Yamada, H.; Mitamura, T.; Horii, T.; Yamamoto, A.; Moriyama, Y. Vacuolar H(+)-ATPase Localized in Plasma Membranes of Malaria Parasite Cells, Plasmodium falciparum, Is Involved in Regional Acidification of Parasitized Erythrocytes. J. Biol. Chem. 2000, 275, 34353–34358. [Google Scholar] [CrossRef] [PubMed]
- Gupta, Y.; Goicoechea, S.; Romero, J.G.; Mathur, R.; Caulfield, T.R.; Becker, D.P.; Durvasula, R.; Kempaiah, P. Repurposing Lansoprazole and Posaconazole to treat Leishmaniasis: Integration of in vitro testing, pharmacological corroboration, and mechanisms of action. J. Food Drug Anal. 2022, 30, 129–150. [Google Scholar] [CrossRef] [PubMed]
- López-Velázquez, G.; Fernández-Lainez, C.; de la Mora-de la Mora, J.I.; Caudillo De La Portilla, D.; Reynoso-Robles, R.; González-Maciel, A.; Ridaura, C.; García-Torres, I.; Gutiérrez-Castrellón, P.; Olivos-García, A.; et al. On the molecular and cellular effects of omeprazole to further support its effectiveness as an antigiardial drug. Sci. Rep. 2019, 9, 8922. [Google Scholar] [CrossRef]
- Tong, S.; Kaitu’U-Lino, T.J.; Hastie, R.; Brownfoot, F.; Cluver, C.; Hannan, N. Pravastatin, proton-pump inhibitors, metformin, micronutrients, and biologics: New horizons for the prevention or treatment of preeclampsia. Am. J. Obstet. Gynecol. 2022, 226, S1157–S1170. [Google Scholar] [CrossRef]
- de Alwis, N.; Binder, N.K.; Beard, S.; Kaitu’U-Lino, T.J.; Tong, S.; Brownfoot, F.; Hannan, N.J. Novel approaches to combat preeclampsia: From new drugs to innovative delivery. Placenta 2020, 102, 10–16. [Google Scholar] [CrossRef]
- Bi, Y.; Might, M.; Vankayalapati, H.; Kuberan, B. Repurposing of Proton Pump Inhibitors as first identified small molecule inhibitors of endo -β- N -acetylglucosaminidase (ENGase) for the treatment of NGLY1 deficiency, a rare genetic disease. Bioorganic Med. Chem. Lett. 2017, 27, 2962–2966. [Google Scholar] [CrossRef]
Figure 1.
Differences in the pH gradients between normal (A) and tumor (B) cells. Notably, the presence of a deranged pH gradient in tumor cells does not allow chemical drugs to enter the cells and carry out their effects; instead, they are entirely protonated and, therefore, blocked outside the cells.
Figure 1.
Differences in the pH gradients between normal (A) and tumor (B) cells. Notably, the presence of a deranged pH gradient in tumor cells does not allow chemical drugs to enter the cells and carry out their effects; instead, they are entirely protonated and, therefore, blocked outside the cells.


{kind=link}
Table 1.
pH and diseases. Direct and indirect evidence that microenvironmental acidity is involved in human diseases.
Table 1.
pH and diseases. Direct and indirect evidence that microenvironmental acidity is involved in human diseases.
| Disease | Histology | Type of Evidence | References |
|---|---|---|---|
| Tumor | Melanoma | Pre-clinical | [45] |
| Pre-clinical | [53] | ||
| Pre-clinical | [48] | ||
| Pre-clinical | [54] | ||
| Pre-clinical | [55] | ||
| Pre-clinical | [56] | ||
| Pre-clinical | [57] | ||
| Pre-clinical | [58] | ||
| Pre-clinical | [59] | ||
| Lymphoma | Pre-clinical | [60] | |
| Pre-clinical | [61] | ||
| Clinical | [62] | ||
| Clinical | [63] | ||
| Myeloma | Pre-clinical | [64] | |
| Pre-clinical | [65] | ||
| Colon | Pre-clinical | [45] | |
| Pre-clinical | [66] | ||
| Pre-clinical | [67] | ||
| Breast | Pre-clinical | [68] | |
| Pre-clinical | [69] | ||
| Pre-clinical | [70] | ||
| Pre-clinical | [71] | ||
| Pre-clinical | [72] | ||
| Pre-clinical | [73] | ||
| Pre-clinical | [74] | ||
| Pre-clinical | [75] | ||
| Clinical | [29] | ||
| Ovary | Pre-clinical | [45] | |
| Pre-clinical | [76] | ||
| Pancreas | Pre-clinical | [70] | |
| Pre-clinical | [77] | ||
| Clinical | [78] | ||
| Gastrointestinal | Pre-clinical | [79] | |
| Pre-clinical | [80] | ||
| Pre-clinical | [81] | ||
| Pre-clinical | [82] | ||
| Pre-clinical | [83] | ||
| Pre-clinical | [84] | ||
| Pre-clinical | [85] | ||
| Clinical | [30] | ||
| Prostate | Pre-clinical | [86] | |
| Pre-clinical | [87] | ||
| Liver | Pre-clinical | [88] | |
| Pre-clinical | [89] | ||
| Pre-clinical | [90] | ||
| Pre-clinical | [91] | ||
| Clinical | [92] | ||
| Lung | Clinical | [62] | |
| Clinical | [63] | ||
| Osteosarcoma | Clinical | [28] | |
| Sarcomas | Pre-clinical | [93] | |
| Clinical | [62] | ||
| Clinical | [63] | ||
| Nervous system | Pre-clinical | [94] | |
| Pre-clinical | [95] | ||
| Sensitivity to radiation therapy | Pre-clinical | [96] | |
| Pre-clinical | [97] | ||
| Cardiovascular diseases | Pre-clinical | [98] | |
| Clinical | [99] | ||
| Clinical | [100] | ||
| Clinical | [101] | ||
| Metabolic diseases | Pre-clinical | [102] | |
| Pre-clinical | [103] | ||
| Pre-clinical | [104] | ||
| Pre-clinical | [105] | ||
| Pre-clinical | [106] | ||
| Pre-clinical | [107] | ||
| Clinical | [108] | ||
| [101] | |||
| Nervous system | Pre-clinical | [109] | |
| Pre-clinical | [110] | ||
| Pre-clinical | [111] | ||
| Pre-clinical | [112] | ||
| Pre-clinical | [113] | ||
| Pre-clinical | [114] | ||
| Inflammation and Immune system | Pre-clinical | [115] | |
| Pre-clinical | [116] | ||
| Pre-clinical | [117] | ||
| Pre-clinical | [118] | ||
| Pre-clinical | [119] | ||
| Pre-clinical | [120] | ||
| Renal function | Clinical | [121] | |
| Pain | Clinical | [122] | |
| Clinical | [123] | ||
| Clinical | [124] | ||
| Infectious agents | Viruses | Preclinical | [125] |
| Preclinical | [126] | ||
| Preclinical | [127] | ||
| Preclinical | [128] | ||
| Translational | [129] | ||
| Candida and other yeasts | Preclinical | [130] | |
| Preclinical | [131] | ||
| Preclinical | [130] | ||
| Preclinical | [132] | ||
| Bacteria | Preclinical | [133] | |
| Preclinical | [134] | ||
| Preclinical | [135] | ||
| Preclinical | [136] | ||
| Preclinical | [137] | ||
| Preclinical | [138] | ||
| Preclinical | [139] | ||
| Parasites | Preclinical | [140] | |
| Preclinical | [141] | ||
| Preclinical | [142] | ||
| Preclinical | [143] | ||
| Other conditions | Pre-eclampsia | Translational | [144] |
| Translational | [145] | ||
| Genetic diseases | Preclinical | [146] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Giuliani, A.; Fais, S. Proposal to Consider Chemical/Physical Microenvironment as a New Therapeutic Off-Target Approach. Pharmaceutics 2022, 14, 2084. https://doi.org/10.3390/pharmaceutics14102084
AMA Style
Giuliani A, Fais S. Proposal to Consider Chemical/Physical Microenvironment as a New Therapeutic Off-Target Approach. Pharmaceutics. 2022; 14(10):2084. https://doi.org/10.3390/pharmaceutics14102084
Chicago/Turabian StyleGiuliani, Alessandro, and Stefano Fais. 2022. "Proposal to Consider Chemical/Physical Microenvironment as a New Therapeutic Off-Target Approach" Pharmaceutics 14, no. 10: 2084. https://doi.org/10.3390/pharmaceutics14102084
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.