
SCMTHP: A New Approach for Identifying and Characterizing of Tumor-Homing Peptides Using Estimated Propensity Scores of Amino Acids

 , , , and
, , , and
Abstract
:
1. Introduction
- To the best of our knowledge, SCMTHP is the first propensity score-based predictor that is employed to create and optimize several new propensity scores of 20 amino acids in becoming THPs via the scoring card method (SCM) [15,16,17]. In the meantime, a single feature descriptor (i.e., AAC) and a single threshold value were implemented in the SCMTHP predictor, and it was found that the approach could easily distinguish THPs from non-THPs.
- Extensive benchmarking experiments show that SCMTHP could outperforms almost all ML-based predictors (e.g., decision tree (DT), k-nearest neighbor (KNN), multi-layer perceptron (MLP), naive Bayes (NB) and partial least squares regression (PLS)) as well as state-of-the-art THP predictors in terms of accuracy, cost-effectiveness and simplicity.
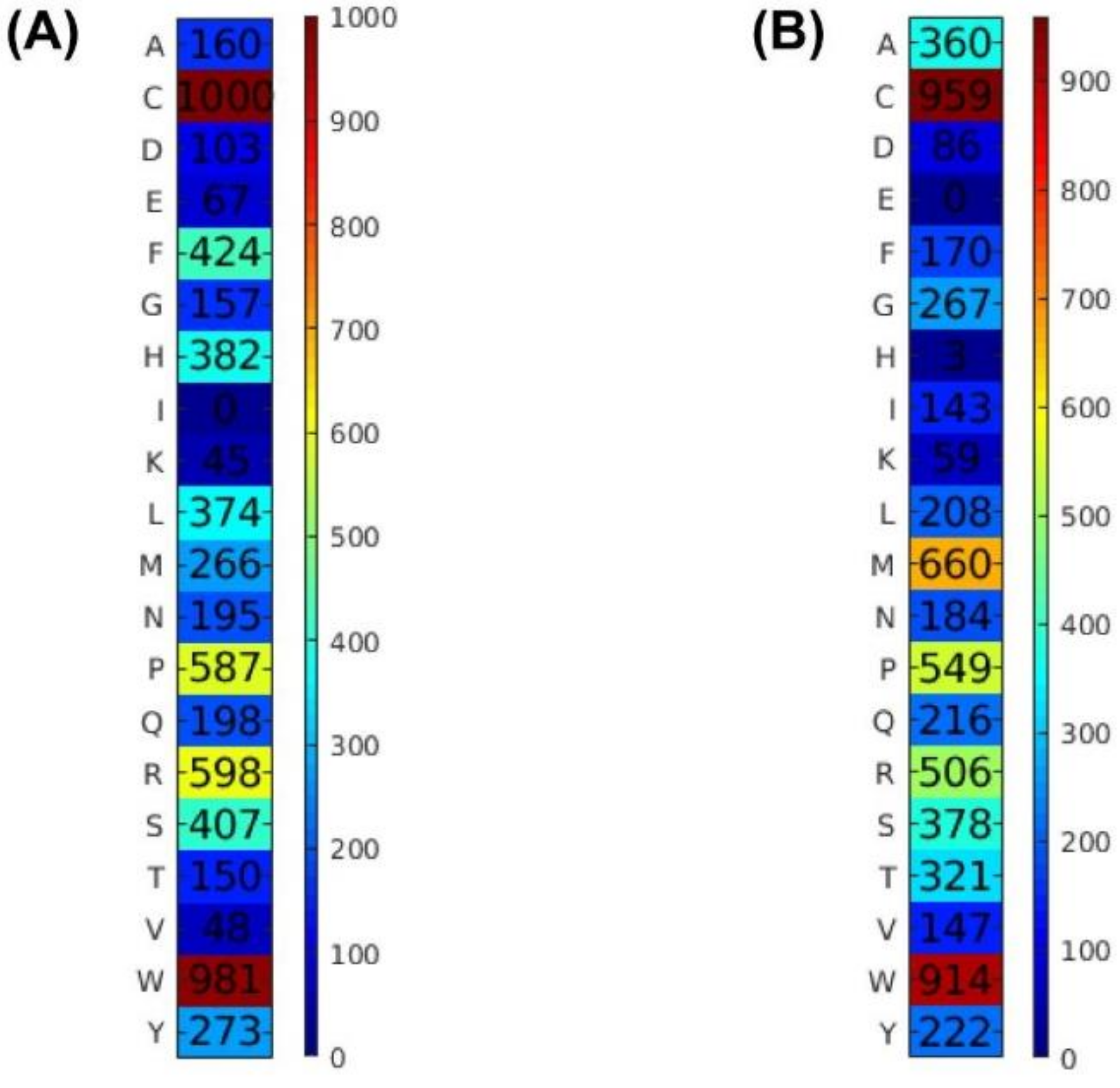
- In order to characterize tumor-homing activities of peptides, SCMTHP-derived propensity scores of 20 amino acids were employed to determine informative physicochemical properties (PCPs) of amino acids as provided in the AAIndex database [18]. The importance of Cys residue in stabilization and the preference for high extinction coefficients are revealed by an analysis of SCMTHP-derived propensity scores.
- A user-friendly online web server was built and deployed publicly at http://pmlabstack.pythonanywhere.com/SCMTHP (accessed on 27 December 2021) in order to facilitate online high-throughput THP identification. We believe that the SCMTHP predictor and SCMTHP-derived propensity scores will be helpful in facilitating THP identification as well as improving our understanding of their biophysical and biochemical properties.
2. Materials and Methods
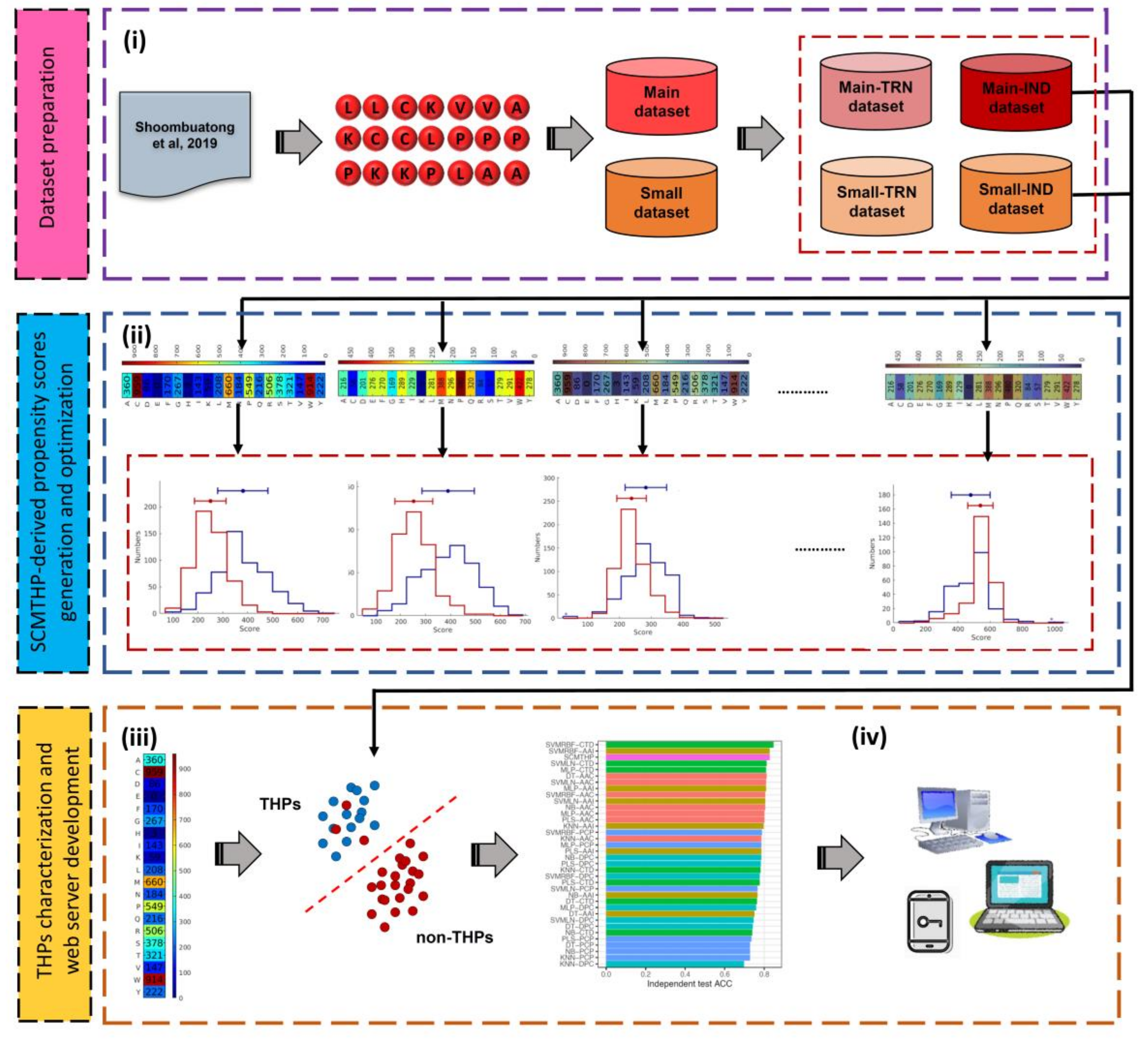
2.1. Dataset Preparation
2.2. Scoring Card Method
2.3. Characterization of THPs Using Informative Physicochemical Properties
2.4. Conventional ML-Based Classifiers
2.5. Performance Evaluation
3. Results and Discussion
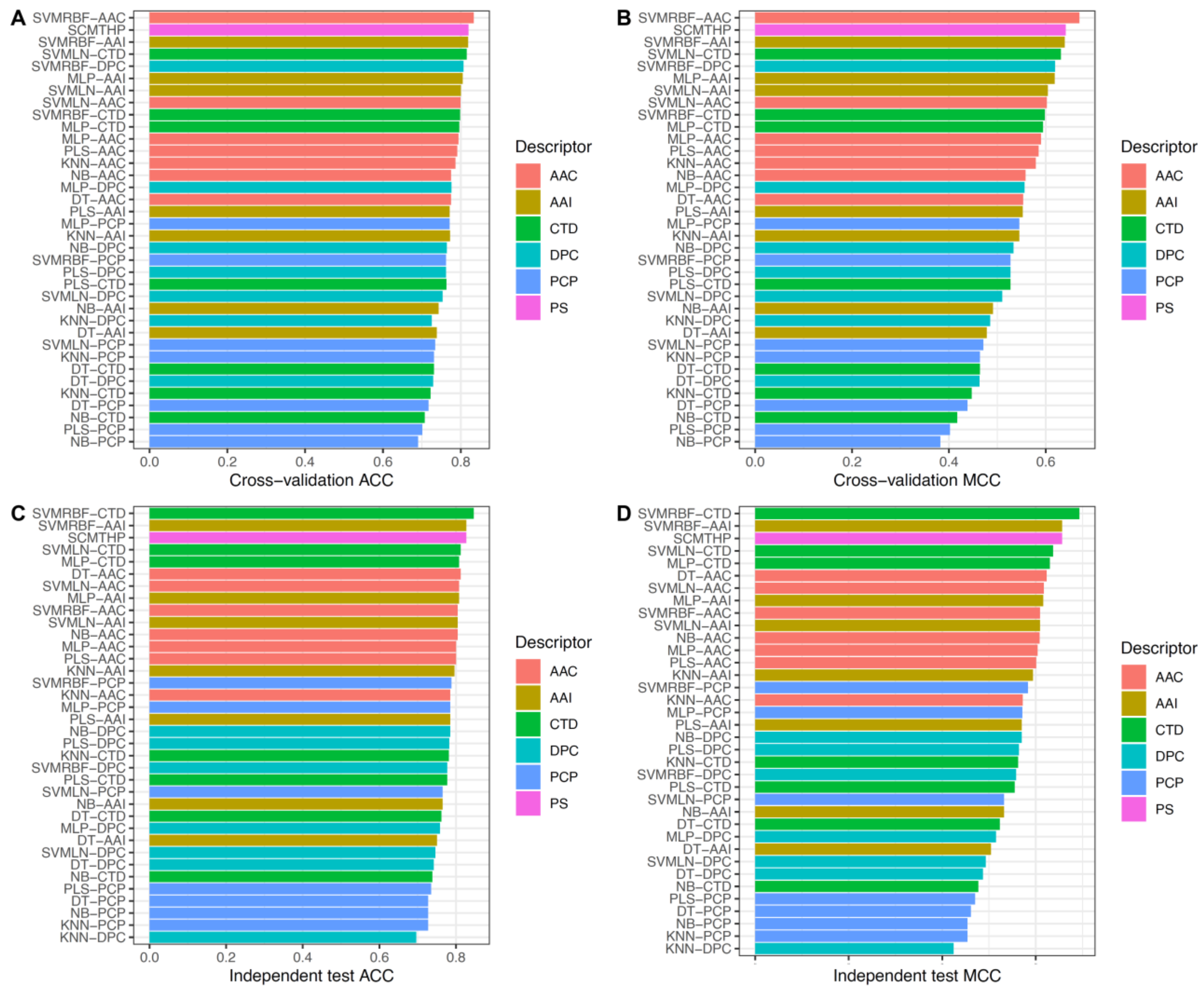
3.1. Performance of Different Propensity Scores
3.2. Comparison of SCMTHP with Well-Known ML Classifiers and Existing Methods
3.3. Contribution of Optimized Propensity Scores
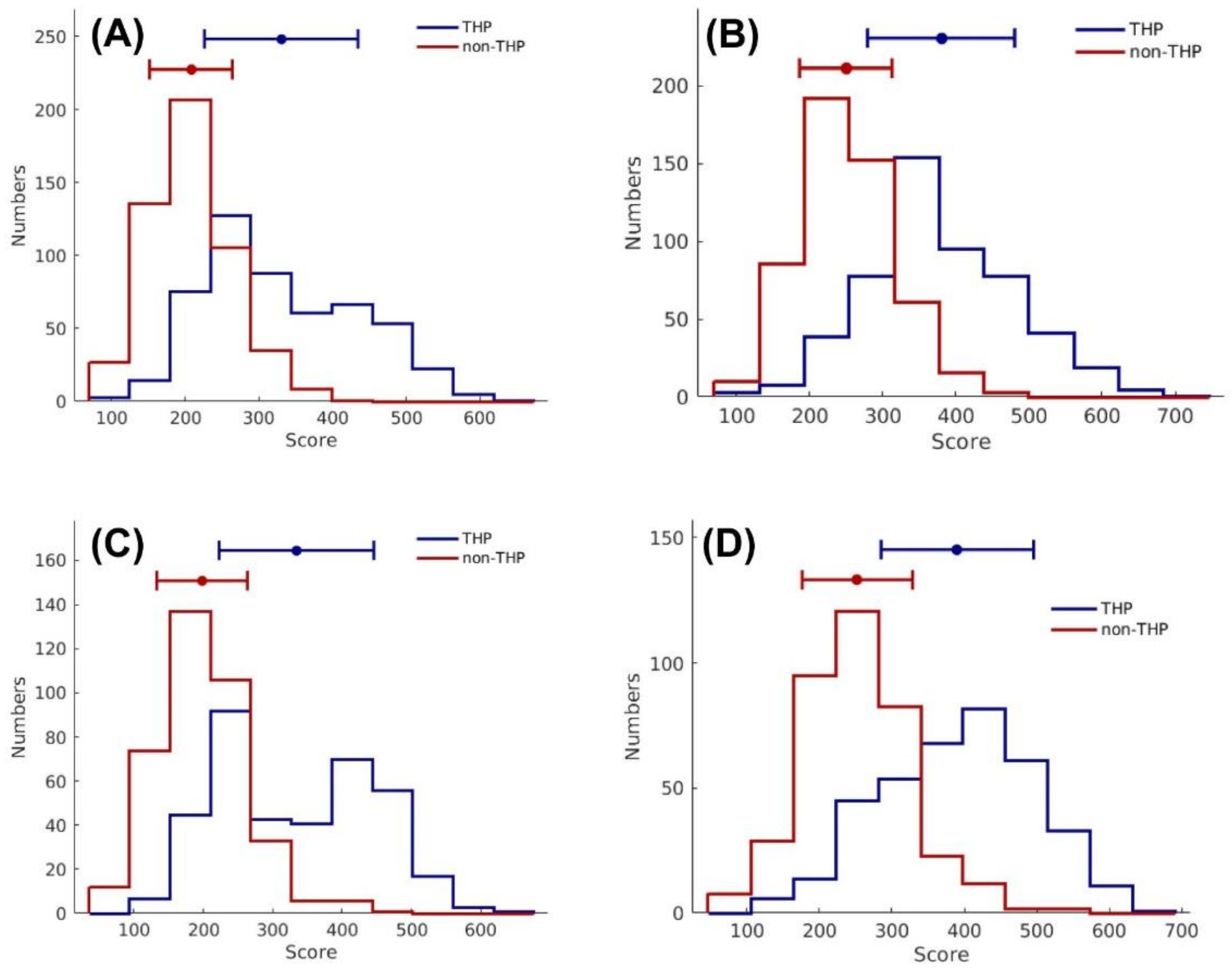
3.4. Identification of Potential THPs Using SCMTHP-Derived Propensity Scores
3.5. Characterization of THPs Using SCMTHP-Derived Propensity Scores
3.6. Characterization of THPs Using Informative PCPs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kondo, E.; Iioka, H.; Saito, K. Tumor-homing peptide and its utility for advanced cancer medicine. Cancer Sci. 2021, 112, 2118–2125. [Google Scholar]
- Lingasamy, P.; Teesalu, T. Homing Peptides for Cancer Therapy. Adv. Exp. Med. Biol. 2021, 1295, 29–48. [Google Scholar]
- Goyal, R.; Jerath, G.; Akhil, R.; Chandrasekharan, A.; Puppala, E.R.; Ponneganti, S.; Sarma, A.; Naidu, V.G.M.; Santhoshkumar, T.R.; Ramakrishnan, V. Geometry encoded functional programming of tumor homing peptides for targeted drug delivery. J. Control. Release 2021, 333, 16–27. [Google Scholar]
- Chen, J.; Yang, H.; Feng, Y.; Shi, Q.; Li, Z.; Tao, Z.; Fan, J.; Jin, Y.; Li, S.; Cheng, J.; et al. A single nucleotide mutation drastically increases the expression of tumor-homing NGR-TNFalpha in the E. coli M15-pQE30 system by improving gene transcription. Appl. Microbiol. Biotechnol. 2021, 105, 1447–1460. [Google Scholar]
- Ottaiano, T.F.; Andrade, S.S.; de Oliveira, C.; Silva, M.C.C.; Buri, M.V.; Juliano, M.A.; Girao, M.; Sampaio, M.U.; Schmaier, A.H.; Wlodawer, A.; et al. Plasma kallikrein enhances platelet aggregation response by subthreshold doses of ADP. Biochimie 2017, 135, 72–81. [Google Scholar]
- Donner, L.; Gremer, L.; Ziehm, T.; Gertzen, C.G.W.; Gohlke, H.; Willbold, D.; Elvers, M. Relevance of N-terminal residues for amyloid-beta binding to platelet integrin alphaIIbbeta3, integrin outside-in signaling and amyloid-beta fibril formation. Cell Signal. 2018, 50, 121–130. [Google Scholar]
- Kodandapani, R.; Veerapandian, B.; Kunicki, T.J.; Ely, K.R. Crystal structure of the OPG2 Fab. An antireceptor antibody that mimics an RGD cell adhesion site. J. Biol. Chem. 1995, 270, 2268–2273. [Google Scholar]
- Kuo, Y.J.; Chung, C.H.; Huang, T.F. From Discovery of Snake Venom Disintegrins to A Safer Therapeutic Antithrombotic Agent. Toxins 2019, 11, 372. [Google Scholar]
- Gautam, A.; Kapoor, P.; Chaudhary, K.; Kumar, R.; Open Source Drug Discovery, C.; Raghava, G.P. Tumor homing peptides as molecular probes for cancer therapeutics, diagnostics and theranostics. Curr. Med. Chem. 2014, 21, 2367–2391. [Google Scholar]
- Schug, C.; Sievert, W.; Urnauer, S.; Muller, A.M.; Schmohl, K.A.; Wechselberger, A.; Schwenk, N.; Lauber, K.; Schwaiger, M.; Multhoff, G.; et al. External Beam Radiation Therapy Enhances Mesenchymal Stem Cell-Mediated Sodium-Iodide Symporter Gene Delivery. Hum. Gene Ther. 2018, 29, 1287–1300. [Google Scholar]
- Dutoit, V.; Migliorini, D.; Ranzanici, G.; Marinari, E.; Widmer, V.; Lobrinus, J.A.; Momjian, S.; Costello, J.; Walker, P.R.; Okada, H.; et al. Antigenic expression and spontaneous immune responses support the use of a selected peptide set from the IMA950 glioblastoma vaccine for immunotherapy of grade II and III glioma. Oncoimmunology 2018, 7, e1391972. [Google Scholar]
- Migliorini, D.; Dutoit, V.; Allard, M.; Grandjean Hallez, N.; Marinari, E.; Widmer, V.; Philippin, G.; Corlazzoli, F.; Gustave, R.; Kreutzfeldt, M.; et al. Phase I/II trial testing safety and immunogenicity of the multipeptide IMA950/poly-ICLC vaccine in newly diagnosed adult malignant astrocytoma patients. Neuro. Oncol. 2019, 21, 923–933. [Google Scholar]
- Sharma, A.; Kapoor, P.; Gautam, A.; Chaudhary, K.; Kumar, R.; Chauhan, J.S.; Tyagi, A.; Raghava, G.P. Computational approach for designing tumor homing peptides. Sci. Rep. 2013, 3, 1607. [Google Scholar]
- Shoombuatong, W.; Schaduangrat, N.; Pratiwi, R.; Nantasenamat, C. THPep: A machine learning-based approach for predicting tumor homing peptides. Comput. Biol. Chem. 2019, 80, 441–451. [Google Scholar]
- Charoenkwan, P.; Chiangjong, W.; Lee, V.S.; Nantasenamat, C.; Hasan, M.M.; Shoombuatong, W. Improved prediction and characterization of anticancer activities of peptides using a novel flexible scoring card method. Sci. Rep. 2021, 11, 3017. [Google Scholar]
- Charoenkwan, P.; Shoombuatong, W.; Lee, H.-C.; Chaijaruwanich, J.; Huang, H.-L.; Ho, S.-Y. SCMCRYS: Predicting protein crystallization using an ensemble scoring card method with estimating propensity scores of P-collocated amino acid pairs. PLoS ONE 2013, 8, e72368. [Google Scholar]
- Huang, H.-L.; Charoenkwan, P.; Kao, T.-F.; Lee, H.-C.; Chang, F.-L.; Huang, W.-L.; Ho, S.-J.; Shu, L.-S.; Chen, W.-L.; Ho, S.-Y. Prediction and analysis of protein solubility using a novel scoring card method with dipeptide composition. BMC Bioinform. 2012, 13, S3. [Google Scholar]
- Kawashima, S.; Kanehisa, M. AAindex: Amino acid index database. Nucleic Acids Res. 2000, 28, 374-374. [Google Scholar]
- Kapoor, P.; Singh, H.; Gautam, A.; Chaudhary, K.; Kumar, R.; Raghava, G.P. TumorHoPe: A database of tumor homing peptides. PLoS ONE 2012, 7, e35187. [Google Scholar]
- Charoenkwan, P.; Kanthawong, S.; Nantasenamat, C.; Hasan, M.M.; Shoombuatong, W. iAMY-SCM: Improved prediction and analysis of amyloid proteins using a scoring card method with propensity scores of dipeptides. Genomics 2021, 113, 689–698. [Google Scholar]
- Charoenkwan, P.; Kanthawong, S.; Nantasenamat, C.; Hasan, M.M.; Shoombuatong, W. iDPPIV-SCM: A sequence-based predictor for identifying and analyzing dipeptidyl peptidase IV (DPP-IV) inhibitory peptides using a scoring card method. J. Proteome Res. 2020, 19, 4125–4136. [Google Scholar]
- Vasylenko, T.; Liou, Y.-F.; Chiou, P.-C.; Chu, H.-W.; Lai, Y.-S.; Chou, Y.-L.; Huang, H.-L.; Ho, S.-Y. SCMBYK: Prediction and characterization of bacterial tyrosine-kinases based on propensity scores of dipeptides. BMC Bioinform. 2016, 17, 203–217. [Google Scholar]
- Liou, Y.-F.; Charoenkwan, P.; Srinivasulu, Y.S.; Vasylenko, T.; Lai, S.-C.; Lee, H.-C.; Chen, Y.-H.; Huang, H.-L.; Ho, S.-Y. SCMHBP: Prediction and analysis of heme binding proteins using propensity scores of dipeptides. BMC Bioinform. 2014, 15, S4. [Google Scholar]
- Charoenkwan, P.; Yana, J.; Nantasenamat, C.; Hasan, M.M.; Shoombuatong, W. iUmami-SCM: A novel sequence-based predictor for prediction and analysis of umami peptides using a scoring card method with propensity scores of dipeptides. J. Chem. Inf. Modeling 2020, 60, 6666–6678. [Google Scholar]
- Charoenkwan, P.; Chotpatiwetchkul, W.; Lee, V.S.; Nantasenamat, C.; Shoombuatong, W. A novel sequence-based predictor for identifying and characterizing thermophilic proteins using estimated propensity scores of dipeptides. Sci. Rep. 2021, 11, 23782. [Google Scholar]
- Liou, Y.-F.; Vasylenko, T.; Yeh, C.-L.; Lin, W.-C.; Chiu, S.-H.; Charoenkwan, P.; Shu, L.-S.; Ho, S.-Y.; Huang, H.-L. SCMMTP: Identifying and characterizing membrane transport proteins using propensity scores of dipeptides. BMC Genom. 2015, 16, S6. [Google Scholar]
- Vasylenko, T.; Liou, Y.-F.; Chen, H.-A.; Charoenkwan, P.; Huang, H.-L.; Ho, S.-Y. SCMPSP: Prediction and characterization of photosynthetic proteins based on a scoring card method. BMC Bioinform. 2015, 16, S8. [Google Scholar]
- Charoenkwan, P.; Schaduangrat, N.; Nantasenamat, C.; Piacham, T.; Shoombuatong, W. iQSP: A Sequence-Based Tool for the Prediction and Analysis of Quorum Sensing Peptides Using Informative Physicochemical Properties. Int. J. Mol. Sci. 2020, 21, 75. [Google Scholar]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V. Scikit-learn: Machine learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Azadpour, M.; McKay, C.M.; Smith, R.L. Estimating confidence intervals for information transfer analysis of confusion matrices. J. Acoust. Soc. Am. 2014, 135, EL140–EL146. [Google Scholar]
- Charoenkwan, P.; Chiangjong, W.; Nantasenamat, C.; Hasan, M.M.; Manavalan, B.; Shoombuatong, W. StackIL6: A stacking ensemble model for improving the prediction of IL-6 inducing peptides. Brief. Bioinform. 2021, 22, bbab172. [Google Scholar]
- Su, R.; Hu, J.; Zou, Q.; Manavalan, B.; Wei, L. Empirical comparison and analysis of web-based cell-penetrating peptide prediction tools. Brief. Bioinform. 2020, 21, 408–420. [Google Scholar]
- Basith, S.; Manavalan, B.; Hwan Shin, T.; Lee, G. Machine intelligence in peptide therapeutics: A next-generation tool for rapid disease screening. Med. Res. Rev. 2020, 40, 1276–1314. [Google Scholar]
- Shoombuatong, W.; Prathipati, P.; Owasirikul, W.; Worachartcheewan, A.; Simeon, S.; Anuwongcharoen, N.; Wikberg, J.E.; Nantasenamat, C. Towards the revival of interpretable QSAR models. In Advances in QSAR Modeling; Springer: Berlin/Heidelberg, Germany, 2017; pp. 3–55. [Google Scholar]
- Zitzmann, S.; Ehemann, V.; Schwab, M. Arginine-glycine-aspartic acid (RGD)-peptide binds to both tumor and tumor-endothelial cells in vivo. Cancer Res. 2002, 62, 5139–5143. [Google Scholar]
- Pasqualini, R.; Koivunen, E.; Kain, R.; Lahdenranta, J.; Sakamoto, M.; Stryhn, A.; Ashmun, R.A.; Shapiro, L.H.; Arap, W.; Ruoslahti, E. Aminopeptidase N is a receptor for tumor-homing peptides and a target for inhibiting angiogenesis. Cancer Res. 2000, 60, 722–727. [Google Scholar]
- Pasqualini, R.; Koivunen, E.; Rouslahti, E. αv integrins as receptors for tumor targeting by circulating ligands. Nat. Biotechnol. 1997, 15, 542–546. [Google Scholar]
- De Groot, F.M.; Broxterman, H.J.; Adams, H.P.; Van Vliet, A.; Tesser, G.I.; Elderkamp, Y.W.; Schraa, A.J.; Kok, R.J.; Molema, G.; Pinedo, H.M. Design, Synthesis, and Biological Evaluation of a Dual Tumor-specific Motive Containing Integrin-targeted Plasmin-cleavable Doxorubicin Prodrug 1 This work was partly supported by the Spinoza Award (to HMP). 1. Mol. Cancer Ther. 2002, 1, 901–911. [Google Scholar]
- Burkhart, D.J.; Kalet, B.T.; Coleman, M.P.; Post, G.C.; Koch, T.H. Doxorubicin-formaldehyde conjugates targeting αvβ3 integrin. Mol. Cancer Ther. 2004, 3, 1593–1604. [Google Scholar]
- Colombo, G.; Curnis, F.; De Mori, G.M.; Gasparri, A.; Longoni, C.; Sacchi, A.; Longhi, R.; Corti, A. Structure-activity relationships of linear and cyclic peptides containing the NGR tumor-homing motif. J. Biol. Chem. 2002, 277, 47891–47897. [Google Scholar]
- Pang, H.-B.; Braun, G.B.; She, Z.-G.; Kotamraju, V.R.; Sugahara, K.N.; Teesalu, T.; Ruoslahti, E. A free cysteine prolongs the half-life of a homing peptide and improves its tumor-penetrating activity. J. Control. Release 2014, 175, 48–53. [Google Scholar]
- Rackovsky, S.; Scheraga, H.A. Hydrophobicity, hydrophilicity, and the radial and orientational distributions of residues in native proteins. Proc. Natl. Acad. Sci. USA 1977, 74, 5248–5251. [Google Scholar]
- Fukuchi, S.; Nishikawa, K. Protein surface amino acid compositions distinctively differ between thermophilic and mesophilic bacteria. J. Mol. Biol. 2001, 309, 835–843. [Google Scholar]
- Qian, N.; Sejnowski, T.J. Predicting the secondary structure of globular proteins using neural network models. J. Mol. Biol. 1988, 202, 865–884. [Google Scholar]
- Nagpal, G.; Usmani, S.S.; Dhanda, S.K.; Kaur, H.; Singh, S.; Sharma, M.; Raghava, G.P. Computer-aided designing of immunosuppressive peptides based on IL-10 inducing potential. Sci. Rep. 2017, 7, 42851. [Google Scholar]
- Kumar, R.; Chaudhary, K.; Chauhan, J.S.; Nagpal, G.; Kumar, R.; Sharma, M.; Raghava, G.P. An in silico platform for predicting, screening and designing of antihypertensive peptides. Sci. Rep. 2015, 5, 12512. [Google Scholar]
- Chaudhary, K.; Kumar, R.; Singh, S.; Tuknait, A.; Gautam, A.; Mathur, D.; Anand, P.; Varshney, G.C.; Raghava, G.P. A web server and mobile app for computing hemolytic potency of peptides. Sci. Rep. 2016, 6, 22843. [Google Scholar]
- McMeekin, T.L.; Groves, M.L.; Hipp, N.J. Refractive indices of amino acids, proteins, and related substances. In Amino Acids and Serum Proteins; Advances in Chemistry Series, No. 44; American Chemical Society: Washington, DC, USA, 1964; Chapter 4; pp. 54–66. [Google Scholar]
- Murata, R.; Inoue, K.-i.; Wang, L.; Ye, S.; Morita, A. Dispersion of Complex Refractive Indices for Intense Vibrational Bands. I. Quantitative Spectra. J. Phys. Chem. B 2021, 125, 9794–9803. [Google Scholar]
- Kuipers, B.J.; Gruppen, H. Prediction of molar extinction coefficients of proteins and peptides using UV absorption of the constituent amino acids at 214 nm to enable quantitative reverse phase high-performance liquid chromatography-mass spectrometry analysis. J. Agric. Food Chem. 2007, 55, 5445–5451. [Google Scholar]
- Liu, Q.; Zhao, H.; Jiang, Y.; Wu, M.; Tian, Y.; Wang, D.; Lao, Y.; Xu, N.; Li, Z. Development of a lytic peptide derived from BH3-only proteins. Cell Death Discov. 2016, 2, 16008. [Google Scholar]
- Xie, J.; Bi, Y.; Zhang, H.; Dong, S.; Teng, L.; Lee, R.J.; Yang, Z. Cell-Penetrating Peptides in Diagnosis and Treatment of Human Diseases: From Preclinical Research to Clinical Application. Front. Pharm. 2020, 11, 697. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dataset | Cross-Validation | Method | ACC | Sn | Sp | MCC | AUC |
|---|---|---|---|---|---|---|---|
| Main | 10-fold CV | THPred | 0.857 | 0.877 | 0.837 | 0.716 | 0.929 |
| SCMTHP | 0.820 | 0.819 | 0.820 | 0.641 | 0.869 | ||
| Independent test | THPred | 0.846 | 0.792 | 0.900 | 0.696 | 0.939 | |
| SCMTHP | 0.827 | 0.869 | 0.785 | 0.656 | 0.869 | ||
| Small | 10-fold CV | THPred | 0.824 | 0.781 | 0.867 | 0.653 | 0.893 |
| SCMTHP | 0.808 | 0.723 | 0.893 | 0.628 | 0.853 | ||
| Independent test | THPred | 0.798 | 0.862 | 0.734 | 0.601 | 0.885 | |
| SCMTHP | 0.798 | 0.766 | 0.830 | 0.597 | 0.853 |
| # | Sequence | THP Score | Length | Molecular Weight | Extinction Coefficient (M−1·cm−1) | pI | Net Charge | Hydrophobicity (Kcal·mol−1) |
|---|---|---|---|---|---|---|---|---|
| 1 | CFWPNRC | 684 | 7 | 925.17 | 5625 | 8.30 | 1 | 6.86 |
| 2 | QWCSRRWCT | 657 | 9 | 1225.52 | 11125 | 8.93 | 2 | 8.78 |
| 3 | WTCRASWCS | 632 | 9 | 1099.35 | 11125 | 8.25 | 1 | 7.16 |
| 4 | SGWCYRC | 631 | 7 | 874.08 | 7115 | 8.21 | 1 | 8.48 |
| 5 | RWCREKSCW | 631 | 9 | 1253.57 | 11125 | 8.85 | 2 | 14.19 |
| 6 | CSDWQHPWC | 627 | 9 | 1161.39 | 11125 | 4.97 | −1 | 11.02 |
| 7 | CPRGSRC | 621 | 7 | 777.99 | 125 | 9.66 | 2 | 13.23 |
| 8 | CWRKFYC | 617 | 7 | 1005.3 | 7115 | 9.24 | 2 | 7.96 |
| 9 | CSDSWHYWC | 615 | 9 | 1186.39 | 12615 | 4.97 | −1 | 9.86 |
| 10 | WRPCES | 607 | 6 | 776.93 | 5500 | 6.16 | 0 | 11.83 |
| 11 | CWLCNGRCGR | 606 | 10 | 1167.52 | 5625 | 8.60 | 2 | 11.27 |
| 12 | RHCFSQWCS | 600 | 9 | 1153.41 | 5625 | 8.19 | 1 | 9.89 |
| 13 | CDCRGDCFC | 598 | 9 | 1021.26 | 250 | 3.91 | −1 | 16.35 |
| 14 | CPHSKPCLC | 598 | 9 | 987.33 | 125 | 8.01 | 1 | 12.46 |
| 15 | CWGCNGRCRM | 595 | 10 | 1185.55 | 5625 | 8.60 | 2 | 11.85 |
| 16 | CSRPRRSEC | 585 | 9 | 1093.34 | 125 | 9.65 | 2 | 17.98 |
| 17 | CSRPRRSVC | 583 | 9 | 1063.36 | 125 | 11.33 | 3 | 13.89 |
| 18 | CVLCNGRCWS | 576 | 10 | 1140.49 | 5625 | 8.00 | 1 | 8.31 |
| 19 | CRGDGWC | 571 | 7 | 795.97 | 5625 | 5.94 | 0 | 13.52 |
| 20 | WREWFL | 571 | 6 | 936.16 | 11000 | 6.70 | 0 | 6.20 |
| 610.25 | 8 | 1041.50 | 6117.25 | 7.82 | 1.00 | 11.05 |
| # | Sequence | THP Score | Length | Molecular Weight | Extinction Coefficient (M−1·cm−1) | pI | Net Charge | Hydrophobicity (Kcal·mol−1) |
|---|---|---|---|---|---|---|---|---|
| 1 | IKIQD | 69 | 5 | 615.80 | 0 | 6.72 | 0 | 12.87 |
| 2 | KKEKDIMKKTI | 74 | 11 | 1361.87 | 0 | 10.39 | 3 | 26.51 |
| 3 | INGKVT | 99 | 6 | 630.83 | 0 | 10.15 | 1 | 11.37 |
| 4 | VKNNVEVN | 105 | 8 | 915.13 | 0 | 6.81 | 0 | 15.50 |
| 5 | IGIGAG | 105 | 6 | 486.66 | 0 | 5.60 | 0 | 9.61 |
| 6 | AVKKAYDIAIQ | 108 | 11 | 1219.60 | 1490 | 9.73 | 1 | 16.00 |
| 7 | DVGTTE | 113 | 6 | 620.69 | 0 | 2.87 | −2 | 16.36 |
| 8 | IGDAT | 114 | 5 | 475.56 | 0 | 3.00 | −1 | 12.32 |
| 9 | VAIDM | 115 | 5 | 547.73 | 0 | 3.02 | −1 | 9.79 |
| 10 | DVKGVFVNI | 119 | 9 | 990.30 | 0 | 6.77 | 0 | 12.13 |
| 11 | DLAVVEVDQVMVVD | 119 | 14 | 1530.96 | 0 | 2.63 | −4 | 19.04 |
| 12 | TDIDDKIINRAI | 121 | 12 | 1386.74 | 0 | 4.21 | −1 | 20.55 |
| 13 | GDVVANT | 123 | 7 | 674.80 | 0 | 3.00 | −1 | 13.37 |
| 14 | IDKQLE | 131 | 6 | 744.93 | 0 | 7.00 | −1 | 16.37 |
| 15 | FGKKKKYKD | 131 | 9 | 1141.50 | 1490 | 10.49 | 4 | 24.27 |
| 16 | KENILNE | 135 | 7 | 859.05 | 0 | 4.08 | −1 | 17.29 |
| 17 | HEAVGI | 136 | 6 | 624.78 | 0 | 5.06 | −1 | 13.93 |
| 18 | HKNKGKKN | 139 | 8 | 953.23 | 0 | 11.03 | 4 | 24.28 |
| 19 | ENAKAAVAEMKDGDVVLLE | 139 | 19 | 2002.54 | 0 | 3.84 | −3 | 31.12 |
| 20 | ITDMAA | 140 | 6 | 620.79 | 0 | 3.13 | −1 | 11.00 |
| 116.75 | 8 | 920.17 | 149.00 | 5.98 | −0.20 | 16.68 |
| Amino Acid | PS-THP | THP (%) | non-THP (%) | Difference | p-Value |
|---|---|---|---|---|---|
| C-Cys | 1000(1) | 9.635 | 1.082 | 8.552(1) | <0.01 * |
| W-Trp | 981(2) | 3.459 | 1.088 | 2.371(3) | <0.01 * |
| R-Arg | 598(3) | 8.947 | 5.062 | 3.885(2) | <0.01 * |
| P-Pro | 587(4) | 6.831 | 4.940 | 1.891(4) | <0.01 * |
| F-Phe | 424(5) | 3.018 | 3.846 | −0.828(13) | 0.017 |
| S-Ser | 407(6) | 8.525 | 6.860 | 1.666(5) | <0.01 * |
| H-His | 382(7) | 3.084 | 2.699 | 0.385(6) | 0.287 |
| L-Leu | 374(8) | 8.157 | 9.394 | −1.237(14) | 0.020 |
| Y-Tyr | 273(9) | 3.023 | 2.912 | 0.111(8) | 0.741 |
| M-Met | 266(10) | 2.629 | 2.604 | 0.025(9) | 0.940 |
| Q-Gln | 198(11) | 3.284 | 4.052 | −0.769(11) | 0.046 |
| N-Asn | 195(12) | 3.365 | 4.169 | −0.804(12) | 0.033 |
| A-Ala | 160(13) | 5.717 | 8.099 | −2.382(16) | <0.01 * |
| G-Gly | 157(14) | 7.552 | 7.203 | 0.349(7) | 0.516 |
| T-Thr | 150(15) | 4.744 | 5.364 | −0.620(10) | 0.186 |
| D-Asp | 103(16) | 3.798 | 5.664 | −1.866(15) | <0.01 * |
| E-Glu | 67(17) | 3.544 | 6.153 | −2.609(19) | <0.01 * |
| V-Val | 48(18) | 4.392 | 6.906 | −2.514(17) | <0.01 * |
| K-Lys | 45(19) | 3.469 | 6.008 | −2.540(18) | <0.01 * |
| I-Ile | 0(20) | 2.828 | 5.894 | −3.066(20) | <0.01 * |
| R | 1.000 | 0.462 | −0.589 | 0.876 | - |
| Amino Acid | PS-THP | MCMT640101 a | Molar Extinction Coefficients |
|---|---|---|---|
| C-Cys | 1000(1) | 35.77(2) | 225(6) |
| W-Trp | 981(2) | 42.53(1) | 29,050(1) |
| R-Arg | 598(3) | 26.66(5) | 102(9) |
| P-Pro | 587(4) | 10.93(17) | 30(19) |
| F-Phe | 424(5) | 29.4(4) | 5200(3) |
| S-Ser | 407(6) | 6.35(18) | 34(17) |
| H-His | 382(7) | 21.81(6) | 5125(4) |
| L-Leu | 374(8) | 18.78(10) | 45(13) |
| Y-Tyr | 273(9) | 31.53(3) | 5375(2) |
| M-Met | 266(10) | 21.64(7) | 980(5) |
| Q-Gln | 198(11) | 17.56(11) | 142(7) |
| N-Asn | 195(12) | 13.28(14) | 136(8) |
| A-Ala | 160(13) | 4.34(19) | 32(18) |
| G-Gly | 157(14) | 0(20) | 21(20) |
| T-Thr | 150(15) | 11.01(16) | 41(16) |
| D-Asp | 103(16) | 12(15) | 58(11) |
| E-Glu | 67(17) | 17.26(12) | 78(10) |
| V-Val | 48(18) | 13.92(13) | 43(14) |
| K-Lys | 45(19) | 21.29(8) | 41(15) |
| I-Ile | 0(20) | 19.06(9) | 45(12) |
| R | 1.000 | 0.635 | 0.556 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Charoenkwan, P.; Chiangjong, W.; Nantasenamat, C.; Moni, M.A.; Lio’, P.; Manavalan, B.; Shoombuatong, W. SCMTHP: A New Approach for Identifying and Characterizing of Tumor-Homing Peptides Using Estimated Propensity Scores of Amino Acids. Pharmaceutics 2022, 14, 122. https://doi.org/10.3390/pharmaceutics14010122
Charoenkwan P, Chiangjong W, Nantasenamat C, Moni MA, Lio’ P, Manavalan B, Shoombuatong W. SCMTHP: A New Approach for Identifying and Characterizing of Tumor-Homing Peptides Using Estimated Propensity Scores of Amino Acids. Pharmaceutics. 2022; 14(1):122. https://doi.org/10.3390/pharmaceutics14010122
Chicago/Turabian StyleCharoenkwan, Phasit, Wararat Chiangjong, Chanin Nantasenamat, Mohammad Ali Moni, Pietro Lio’, Balachandran Manavalan, and Watshara Shoombuatong. 2022. "SCMTHP: A New Approach for Identifying and Characterizing of Tumor-Homing Peptides Using Estimated Propensity Scores of Amino Acids" Pharmaceutics 14, no. 1: 122. https://doi.org/10.3390/pharmaceutics14010122