
Hidradenitis Suppurativa and Comorbid Disorder Biomarkers, Druggable Genes, New Drugs and Drug Repurposing—A Molecular Meta-Analysis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Literature Search
2.2. Study Selection
2.3. Data Extraction
2.4. Data Analysis
2.5. Drug Repurposing Sources
2.6. Statistics
3. Results
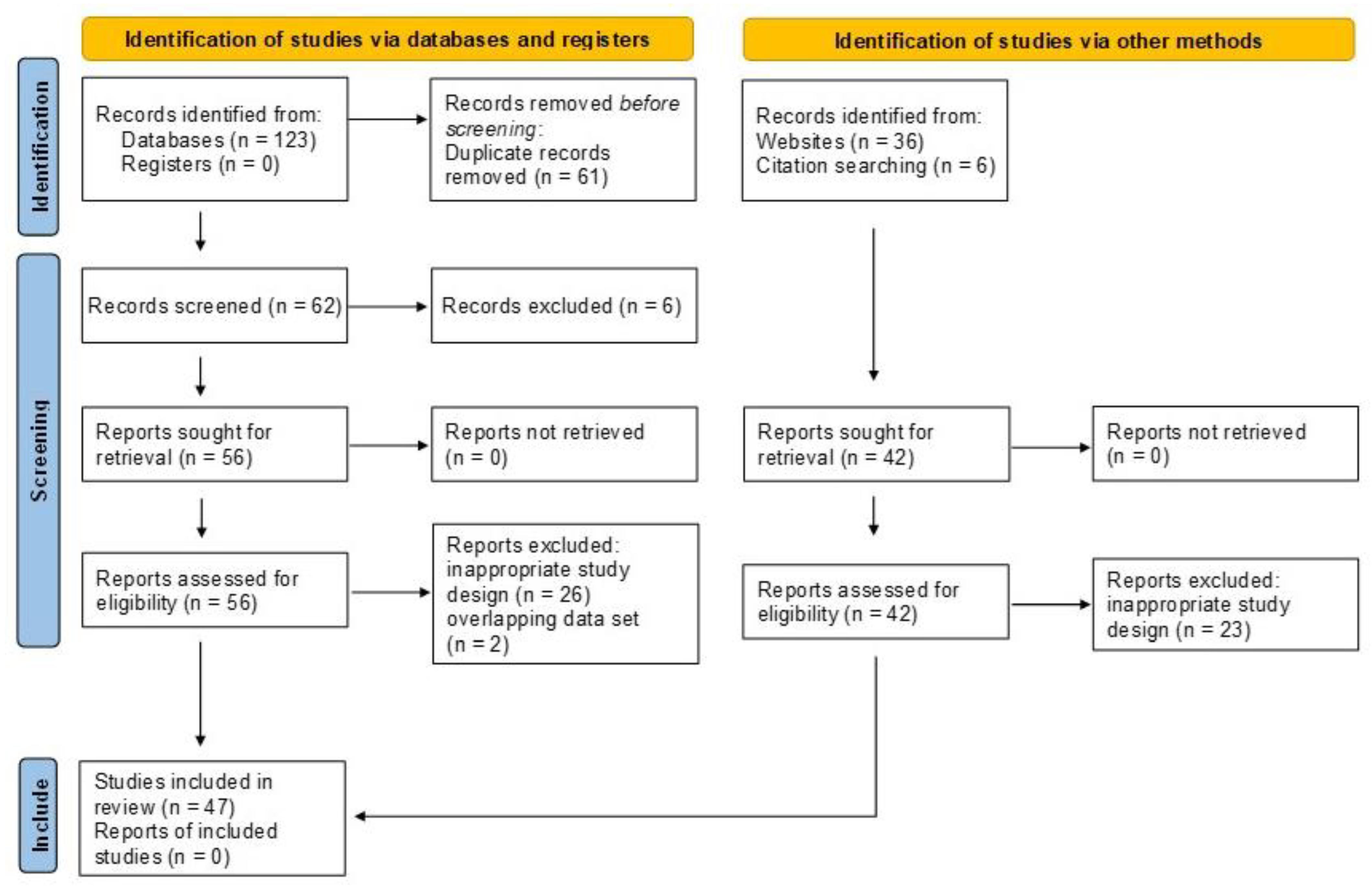
3.1. Study Selection Process
3.2. Differentially Expressed Genes and Proteins in HS
3.3. HS Biomarkers
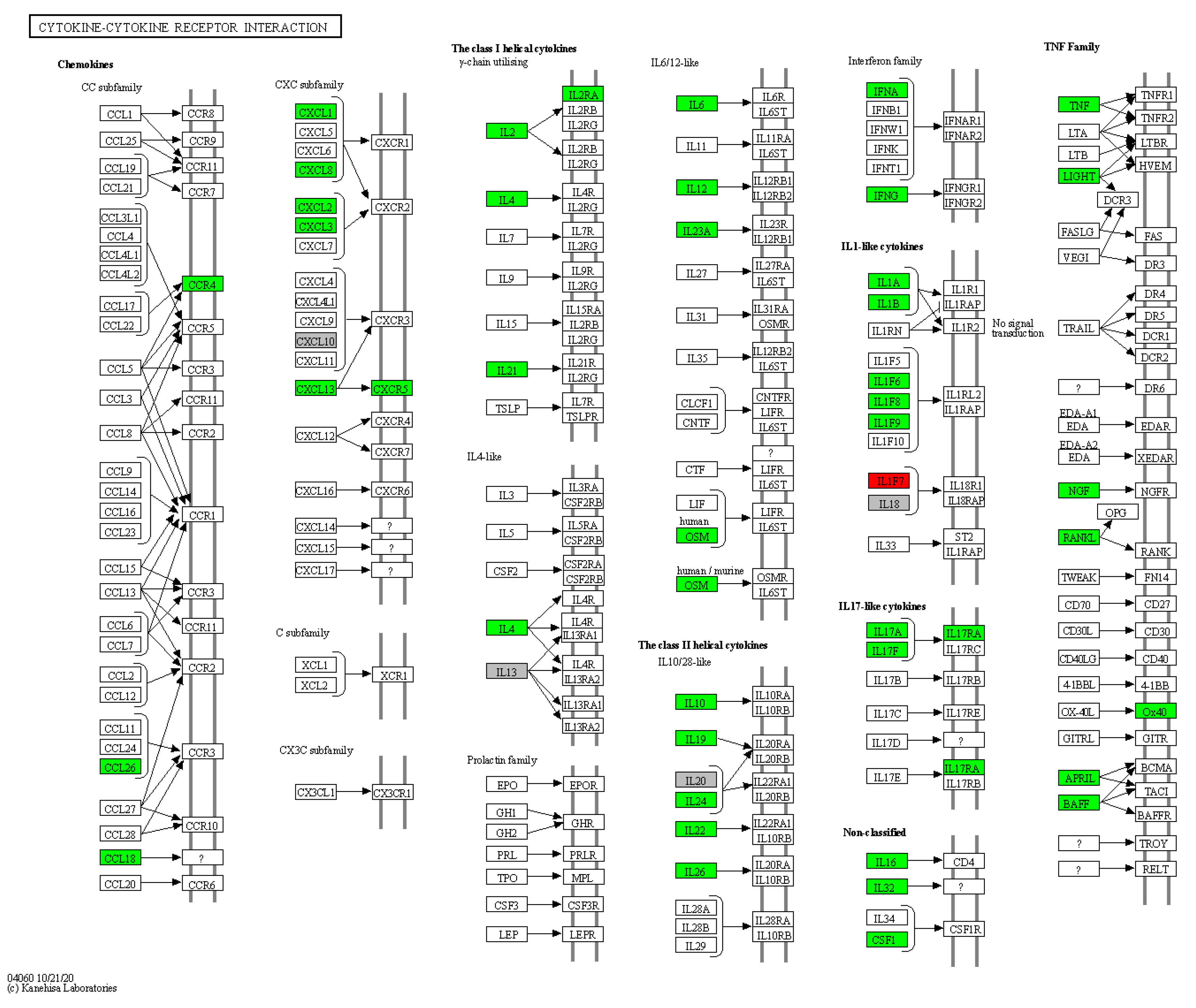
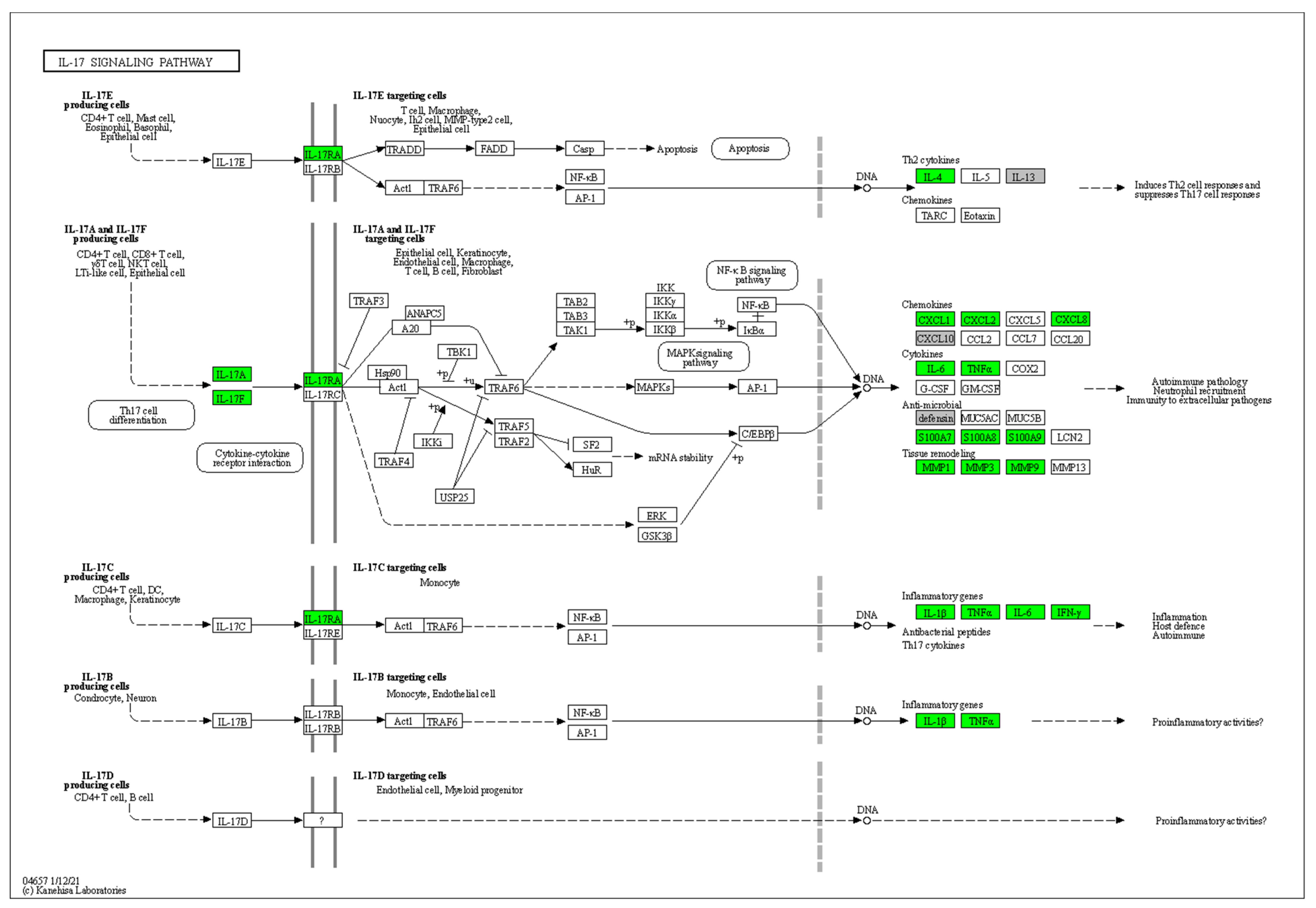
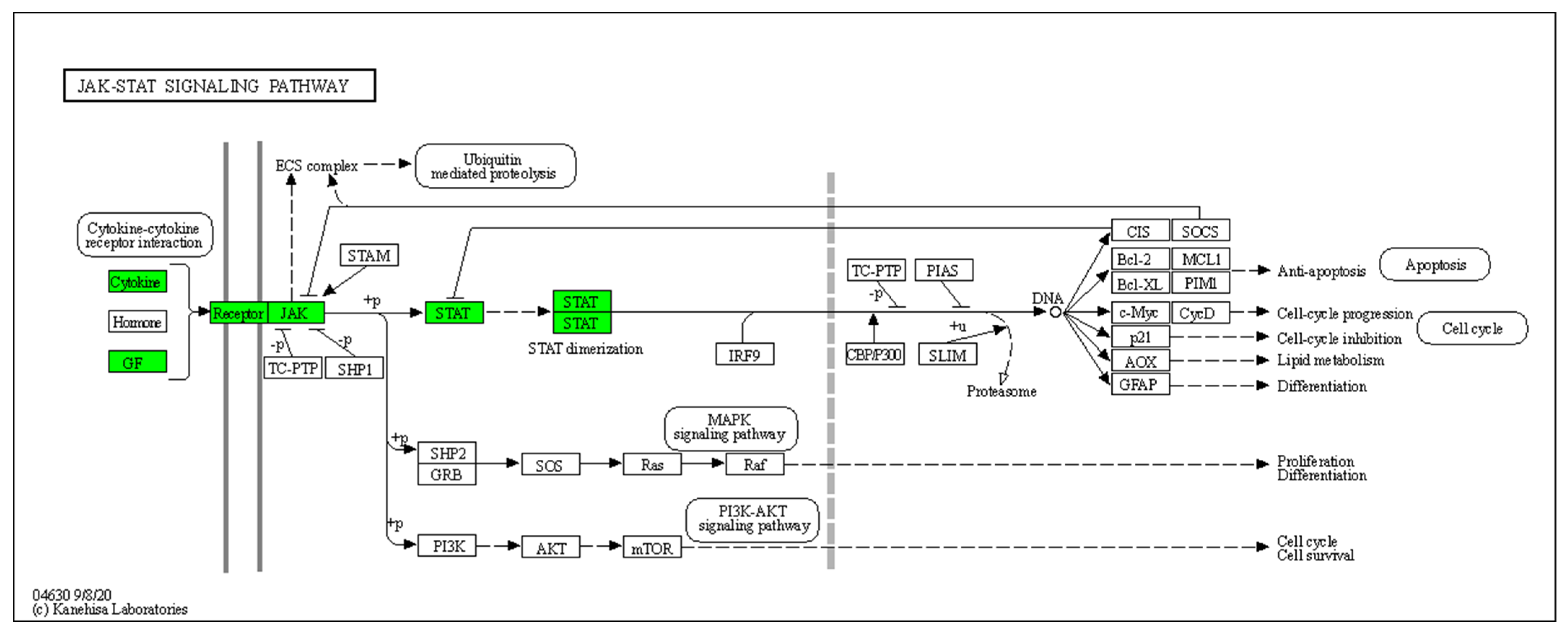
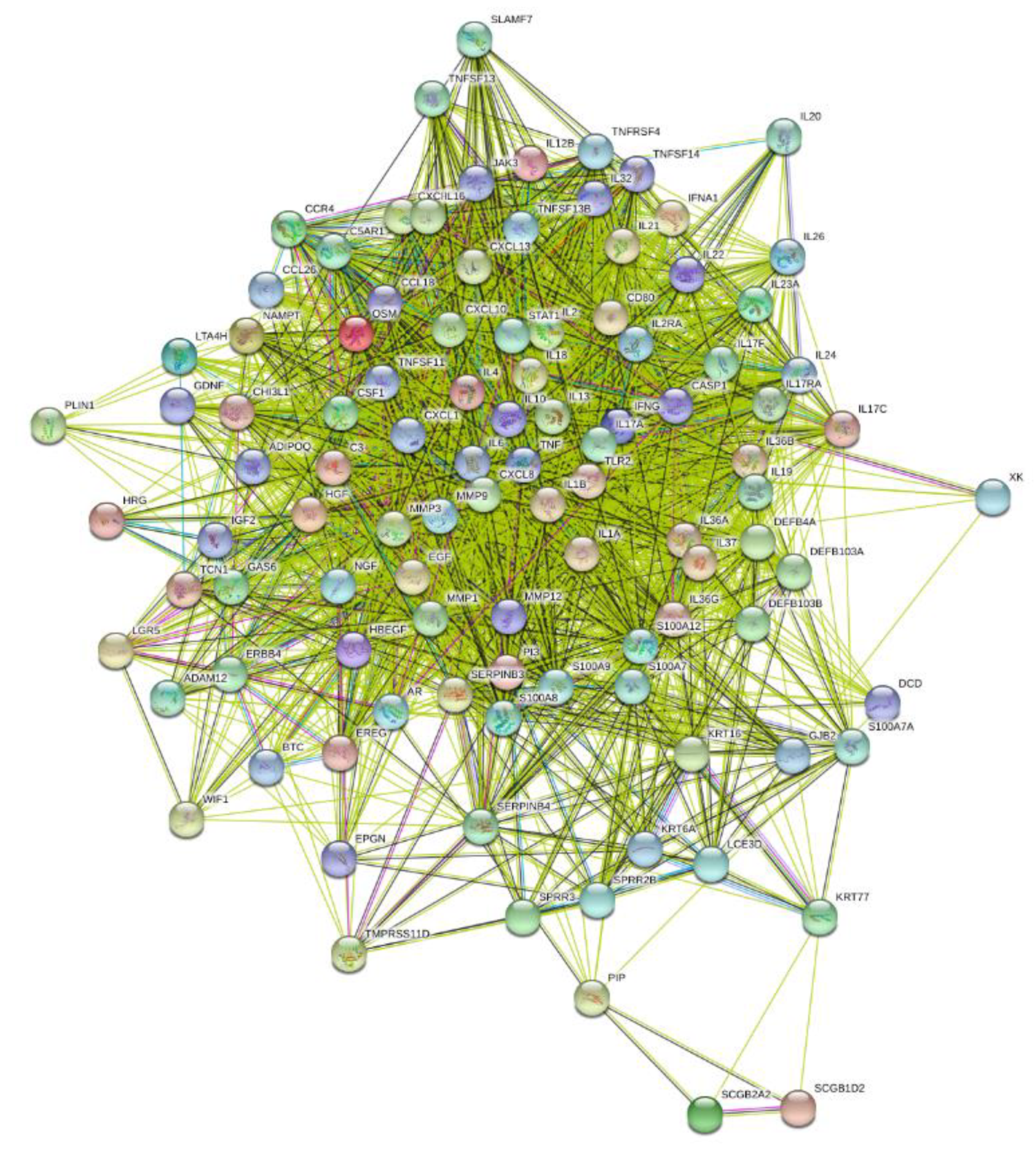
3.4. Enrichment Analysis of HS-Associated Genes
3.5. Enrichment Analysis of HS Druggable Genes
3.6. Study Drugs and Drug Repurposing for HS
4. Discussion
4.1. HS Pathogenesis
4.2. HS Comorbid Disorders
4.3. Study Drugs and Drug Repurposing for HS
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zouboulis, C.C.; Del Marmol, V.; Mrowietz, U.; Prens, E.P.; Tzellos, T.; Jemec, G.B.E. Hidradenitis suppurativa/acne inversa: Criteria for diagnosis, severity assessment, classification and disease evaluation. Dermatology 2015, 231, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Del Duca, E.; Morelli, P.; Bennardo, L.; Di Raimondo, C.; Nisticò, S.P. Cytokine pathways and investigational target therapies in hidradenitis suppurativa. Int. J. Mol. Sci. 2020, 21, 8436. [Google Scholar] [CrossRef]
- Zouboulis, C.C.; Nogueira da Costa, A.; Makrantonaki, E.; Hou, X.X.; Almansouri, D.; Dudley, J.T.; Edwards, H.; Readhead, B.; Balthasar, O.; Jemec, G.B.E.; et al. Alterations in innate immunity and epithelial cell differentiation are the molecular pillars of hidradenitis suppurativa. J. Eur. Acad. Dermatol. Venereol. 2020, 34, 846–861. [Google Scholar] [CrossRef] [PubMed]
- Kaleta, K.P.; Nikolakis, G.; Hossini, A.M.; Balthasar, O.; Almansouri, D.; Vaiopoulos, A.; Knolle, J.; Boguslawska, A.; Wojas-Pelc, A.; Zouboulis, C.C. Metabolic disorders/obesity is a primary risk factor in hidradenitis suppurativa: An immunohistochemical real-world approach. Dermatology 2021. [Google Scholar] [CrossRef]
- Zouboulis, C.C.; Desai, N.; Emtestam, L.; Hunger, R.E.; Ioannides, D.; Juhász, I.; Lapins, J.; Matusiak, L.; Prens, E.P.; Revuz, J.; et al. European S1 guideline for the treatment of hidradenitis suppurativa/acne inversa. J Eur. Acad. Dermatol. Venereol. 2015, 29, 619–644. [Google Scholar] [CrossRef]
- Zouboulis, C.C.; Bechara, F.G.; Dickinson-Blok, J.L.; Gulliver, W.; Horváth, B.; Hughes, R.; Kimball, A.B.; Kirby, B.; Martorell, A.; Podda, M.; et al. Hidradenitis suppurativa/acne inversa: A practical framework for treatment optimization—Systematic review and recommendations from the HS ALLIANCE working group. J. Eur. Acad. Dermatol. Venereol. 2019, 33, 19–31. [Google Scholar] [CrossRef]
- Zouboulis, C.C.; Benhadou, F.; Byrd, A.; Chandran, N.; Giamarellos-Bourboulis, E.; Fabbrocini, G.; Frew, J.; Fujita, H.; González-López, M.A.; Guillem, P.; et al. What causes hidradenitis suppurativa? 15 years after. Exp. Dermatol. 2020, 29, 1154–1170. [Google Scholar] [CrossRef]
- Zouboulis, C.C.; Frew, J.W.; Giamarellos-Bourboulis, E.J.; Jemec, G.B.E.; del Marmol, V.; Marzano, A.V.; Nikolakis, G.; Sayed, C.J.; Tzellos, T.; Wolk, K.; et al. Target molecules for future hidradenitis suppurativa treatment. Exp. Dermatol. 2021, 30 (Suppl. 1), 8–17. [Google Scholar] [CrossRef]
- Kimball, A.B.; Okun, M.M.; Williams, D.A.; Gottlieb, A.B.; Papp, K.A.; Zouboulis, C.C.; Armstrong, A.W.; Kerdel, F.; Gold, M.H.; Forman, S.B.; et al. Two phase 3 trials of adalimumab treatment of hidradenitis suppurativa. N. Engl. J. Med. 2016, 375, 422–434. [Google Scholar] [CrossRef]
- Glatt, S.; Jemec, G.B.; Forman, S.; Sayed, C.; Schmieder, G.; Weisman, J.; Rolleri, R.; Seegobin, S.; Baeten, D.; Ionescu, L.; et al. Bimekizumab in moderate-to-severe hidradenitis suppurativa: A phase 2, double-blind, placebo-controlled randomized clinical trial. JAMA Dermatol. 2021, 157, 1279–1288. [Google Scholar] [CrossRef] [PubMed]
- Zouboulis, C.C.; Nogueira da Costa, A. Drug repurposing through drug-gene interaction profiles for hidradenitis suppurativa/acne inversa treatment. J Eur Acad Dermatol Venereol. 2021, 35, e251–e254. [Google Scholar] [CrossRef]
- Zouboulis, C.C.; Readhead, B.; Dudley, J.T. An additional drug repurposing study for hidradenitis suppurativa/acne inversa. Br. J. Dermatol. 2021, 184, 748–750. [Google Scholar] [CrossRef]
- Giordano, A.; Forte, G.; Massimo, L.; Riccio, R.; Bifulco, G.; Di Micco, S. Discovery of new erbB4 inhibitors: Repositioning an orphan chemical library by inverse virtual screening. Eur. J. Med. Chem. 2018, 152, 253–263. [Google Scholar] [CrossRef]
- Yu, J.L.; Dai, Q.Q.; Li, G.B. Deep learning in target prediction and drug repositioning: Recent advances and challenges. Drug Discov. Today 2021. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Bmj 2021, 372, n71. [Google Scholar] [CrossRef]
- Gene Expression Omnibus. Available online: https://www.ncbi.nlm.nih.gov/geo/ (accessed on 15 November 2021).
- HUGO Gene Nomenclature Committee. Available online: https://www.genenames.org/tools/multi-symbol-checker/ (accessed on 15 November 2021).
- Biological DataBase network. Available online: https://biodbnet-abcc.ncifcrf.gov/db/db2db.php (accessed on 15 November 2021).
- g:Profiler. Available online: https://biit.cs.ut.ee/gprofiler/gost (accessed on 15 November 2021).
- Kyoto Encyclopedia of Genes and Genomes. Available online: https://www.genome.jp/kegg/ (accessed on 15 November 2021).
- Reactome. Available online: https://reactome.org/ (accessed on 15 November 2021).
- WikiPathways. Available online: https://www.wikipathways.org/ (accessed on 15 November 2021).
- Human Phenotype Ontology. Available online: https://hpo.jax.org/app/ (accessed on 15 November 2021).
- The Drug Repurposing Hub. Available online: https://s3.amazonaws.com/data.clue.io/repurposing/downloads/repurposing_drugs_20200324.txt (accessed on 15 November 2021).
- Gene Cards, The Human Gene Database. Available online: https://www.genecards.org/ (accessed on 15 November 2021).
- Lowe, M.M.; Naik, H.B.; Clancy, S.; Pauli, M.; Smith, K.M.; Bi, Y.; Dunstan, R.; Gudjonsson, J.E.; Paul, M.; Harris, H.; et al. Immunopathogenesis of hidradenitis suppurativa and response to anti-TNF-alpha therapy. JCI Insight 2020, 5, e139932. [Google Scholar] [CrossRef]
- Hoffman, L.K.; Tomalin, L.E.; Schultz, G.; Howell, M.D.; Anandasabapathy, D.; Alavi, A.; Suárez-Fariñas, M.; Lowes, M.A. Integrating the skin and blood transcriptomes and serum proteome in hidradenitis suppurativa reveals complement dysregulation and a plasma cell signature. PLoS ONE 2018, 13, e0203672. [Google Scholar] [CrossRef] [Green Version]
- González-López, M.A.; Vilanova, I.; Ocejo-Viñals, G.; Arlegui, R.; Navarro, I.; Guiral, S.; Mata, C.; Pérez-Paredes, M.G.; Portilla, V.; Corrales, A.; et al. Circulating levels of adiponectin, leptin, resistin and visfatin in non-diabetics patients with hidradenitis suppurativa. Arch. Dermatol. Res. 2020, 312, 595–600. [Google Scholar] [CrossRef]
- Hessam, S.; Sand, M.; Skrygan, M.; Bechara, F.G. The microRNA effector RNA-induced silencing complex in hidradenitis suppurativa: A significant dysregulation within active inflammatory lesions. Arch. Dermatol. Res. 2017, 309, 557–565. [Google Scholar] [CrossRef]
- Rumberger, B.E.; Boarder, E.L.; Owens, S.L.; Howell, M.D. Transcriptomic analysis of hidradenitis suppurativa skin suggests roles for multiple inflammatory pathways in disease pathogenesis. Inflamm. Res. 2020, 69, 967–973. [Google Scholar] [CrossRef]
- Penno, C.A.; Jäger, P.; Laguerre, C.; Hasler, F.; Hofmann, A.; Gass, S.K.; Wettstein-Ling, B.; Schaefer, D.J.; Avrameas, A.; Raulf, F.; et al. Lipidomics profiling of hidradenitis suppurativa skin lesions reveals lipoxygenase pathway dysregulation and accumulation of proinflammatory leukotriene B4. J. Investig. Dermatol. 2020, 140, 2421–2432. [Google Scholar] [CrossRef]
- Coates, M.; Mariottoni, P.; Corcoran, D.L.; Kirshner, H.F.; Jaleel, T.; Brown, D.A.; Brooks, S.R.; Murray, J.; Morasso, M.I.; MacLeod, A.S. The skin transcriptome in hidradenitis suppurativa uncovers an antimicrobial and sweat gland gene signature which has distinct overlap with wounded skin. PLoS ONE 2019, 14, e0216249. [Google Scholar] [CrossRef] [Green Version]
- Shanmugam, V.K.; Jones, D.; McNish, S.; Bendall, M.L.; Crandall, K.A. Transcriptome patterns in hidradenitis suppurativa: Support for the role of antimicrobial peptides and interferon pathways in disease pathogenesis. Clin. Exp. Dermatol. 2019, 44, 882–892. [Google Scholar] [CrossRef]
- Blok, J.L.; Li, K.; Brodmerkel, C.; Jonkman, M.F.; Horváth, B. Gene expression profiling of skin and blood in hidradenitis suppurativa. Br. J. Dermatol. 2016, 174, 1392–1394. [Google Scholar] [CrossRef]
- Buimer, M.G.; Wobbes, T.; Klinkenbijl, J.H.; Reijnen, M.M.; Blokx, W.A. Immunohistochemical analysis of steroid hormone receptors in hidradenitis suppurativa. Am. J. Dermatopathol. 2015, 37, 129–132. [Google Scholar] [CrossRef]
- Vossen, A.; van der Zee, H.H.; Davelaar, N.; Mus, A.M.C.; van Doorn, M.B.A.; Prens, E.P. Apremilast for moderate hidradenitis suppurativa: No significant change in lesional skin inflammatory biomarkers. J. Eur. Acad. Dermatol. Venereol. 2019, 33, 761–765. [Google Scholar] [CrossRef]
- Kanni, T.; Zenker, O.; Habel, M.; Riedemann, N.; Giamarellos-Bourboulis, E.J. Complement activation in hidradenitis suppurativa: A new pathway of pathogenesis? Br. J. Dermatol. 2018, 179, 413–419. [Google Scholar] [CrossRef] [Green Version]
- Kelly, G.; Hughes, R.; McGarry, T.; van den Born, M.; Adamzik, K.; Fitzgerald, R.; Lawlor, C.; Tobin, A.M.; Sweeney, C.M.; Kirby, B. Dysregulated cytokine expression in lesional and nonlesional skin in hidradenitis suppurativa. Br. J. Dermatol. 2015, 173, 1431–1439. [Google Scholar] [CrossRef]
- Vossen, A.; Stubbs, A.; van Doorn, M.; van Straalen, K.; van der Zee, H.; Prens, E. Profiling of the transcriptome in hidradenitis suppurativa: A case-control sample. J. Investig. Dermatol. 2016, 136, S193. [Google Scholar] [CrossRef]
- Witte-Händel, E.; Wolk, K.; Tsaousi, A.; Irmer, M.L.; Mößner, R.; Shomroni, O.; Lingner, T.; Witte, K.; Kunkel, D.; Salinas, G.; et al. The IL-1 pathway is hyperactive in hidradenitis suppurativa and contributes to skin infiltration and destruction. J. Investig. Dermatol. 2019, 139, 1294–1305. [Google Scholar] [CrossRef]
- Vossen, A.; van der Zee, H.H.; Tsoi, L.C.; Xing, X.; Devalaraja, M.; Gudjonsson, J.E.; Prens, E.P. Novel cytokine and chemokine markers of hidradenitis suppurativa reflect chronic inflammation and itch. Allergy 2019, 74, 631–634. [Google Scholar] [CrossRef]
- Garcet, S.; Frew, J.W.; Navrazhina, K.; Krueger, J. Hidradenitis suppurativa RNA-seq skin transcriptome overlaps with psoriasis vulgaris and reveals a marked upregulation of multiple targetable cytokines. J. Investig. Dermatol. 2020, 140, S7. [Google Scholar] [CrossRef]
- Byrd, A.S.; Kerns, M.L.; Williams, D.W.; Zarif, J.C.; Rosenberg, A.Z.; Delsante, M.; Liu, H.; Dillen, C.A.; Maynard, J.P.; Caffrey, J.A.; et al. Collagen deposition in chronic hidradenitis suppurativa: Potential role for CD163(+) macrophages. Br. J. Dermatol. 2018, 179, 792–794. [Google Scholar] [CrossRef]
- Hotz, C.; Boniotto, M.; Guguin, A.; Surenaud, M.; Jean-Louis, F.; Tisserand, P.; Ortonne, N.; Hersant, B.; Bosc, R.; Poli, F.; et al. Intrinsic defect in keratinocyte function leads to inflammation in hidradenitis suppurativa. J. Investig. Dermatol. 2016, 136, 1768–1780. [Google Scholar] [CrossRef] [Green Version]
- Gudjonsson, J.E.; Tsoi, L.C.; Ma, F.; Billi, A.C.; van Straalen, K.R.; Vossen, A.R.J.V.; van der Zee, H.H.; Harms, P.W.; Wasikowski, R.; Yee, C.M.; et al. Contribution of plasma cells and B cells to hidradenitis suppurativa pathogenesis. JCI Insight 2020, 5, e139930. [Google Scholar] [CrossRef] [PubMed]
- Wolk, K.; Warszawska, K.; Hoeflich, C.; Witte, E.; Schneider-Burrus, S.; Witte, K.; Kunz, S.; Buss, A.; Roewert, H.J.; Krause, M.; et al. Deficiency of IL-22 contributes to a chronic inflammatory disease: Pathogenetic mechanisms in acne inversa. J. Immunol. 2011, 186, 1228–1239. [Google Scholar] [CrossRef] [Green Version]
- Nelson, A.M.; Cong, Z.; Gettle, S.L.; Longenecker, A.L.; Kidacki, M.; Kirby, J.S.; Adams, D.R.; Stairs, D.B.; Danby, F.W. E-cadherin and p120ctn protein expression are lost in hidradenitis suppurativa lesions. Exp. Dermatol. 2019, 28, 867–871. [Google Scholar] [CrossRef] [PubMed]
- Dany, M.; Elston, D. Gene expression of sphingolipid metabolism pathways is altered in hidradenitis suppurativa. J. Am. Acad. Dermatol. 2017, 77, 268–273. [Google Scholar] [CrossRef]
- Matusiak, L.; Salomon, J.; Nowicka-Suszko, D.; Bieniek, A.; Szepietowski, J.C. Chitinase-3-like protein 1 (YKL-40): Novel biomarker of hidradenitis suppurativa disease activity? Acta Dermatovenereol. 2015, 95, 736–737. [Google Scholar] [CrossRef]
- Salomon, J.; Piotrowska, A.; Matusiak, L.; Dziegiel, P.; Szepietowski, J.C. Chitinase-3-like protein 1 (YKL-40) Is expressed in lesional skin in hidradenitis suppurativa. Vivo 2019, 33, 141–143. [Google Scholar] [CrossRef] [Green Version]
- Wolk, K.; Brembach, T.C.; Simaite, D.; Bartnik, E.; Cucinotta, S.; Pokrywka, A.; Irmer, M.L.; Triebus, J.; Witte-Handel, E.; Salinas, G.; et al. Activity and components of the granulocyte colony-stimulating factor pathway in hidradenitis suppurativa. Br. J. Dermatol. 2021, 185, 164–176. [Google Scholar] [CrossRef]
- Hofmann, S.C.; Saborowski, V.; Lange, S.; Kern, W.V.; Bruckner-Tuderman, L.; Rieg, S. Expression of innate defense antimicrobial peptides in hidradenitis suppurativa. J. Am. Acad. Dermatol. 2012, 66, 966–974. [Google Scholar] [CrossRef]
- Dréno, B.; Khammari, A.; Brocard, A.; Moyse, D.; Blouin, E.; Guillet, G.; Leonard, F.; Knol, A.C. Hidradenitis suppurativa: The role of deficient cutaneous innate immunity. Arch. Dermatol. 2012, 148, 182–186. [Google Scholar] [CrossRef] [Green Version]
- Argyropoulou, M.; Grundhuber, M.; Kanni, T.; Tzanetakou, V.; Micha, S.; Stergianou, D.; Swiniarski, S.; Giamarellos-Bourboulis, E.J. A composite biomarker score for the diagnosis of hidradenitis suppurativa. Exp. Dermatol. 2019, 28, 18. [Google Scholar] [CrossRef] [Green Version]
- Hessam, S.; Sand, M.; Skrygan, M.; Gambichler, T.; Bechara, F.G. Inflammation induced changes in the expression levels of components of the microRNA maturation machinery Drosha, Dicer, Drosha co-factor DGRC8 and Exportin-5 in inflammatory lesions of hidradenitis suppurativa patients. J. Dermatol. Sci. 2016, 82, 166–174. [Google Scholar] [CrossRef]
- Van der Zee, H.H.; de Ruiter, L.; van den Broecke, D.G.; Dik, W.A.; Laman, J.D.; Prens, E.P. Elevated levels of tumour necrosis factor (TNF)-α, interleukin (IL)-1β and IL-10 in hidradenitis suppurativa skin: A rationale for targeting TNF-α and IL-1β. Br. J. Dermatol. 2011, 164, 1292–1298. [Google Scholar] [CrossRef]
- Wieland, C.W.; Vogl, T.; Ordelman, A.; Vloedgraven, H.G.; Verwoolde, L.H.; Rensen, J.M.; Roth, J.; Boer, J.; Hessels, J. Myeloid marker S100A8/A9 and lymphocyte marker, soluble interleukin 2 receptor: Biomarkers of hidradenitis suppurativa disease activity? Br. J. Dermatol. 2013, 168, 1252–1258. [Google Scholar] [CrossRef]
- Montaudié, H.; Seitz-Polski, B.; Cornille, A.; Benzaken, S.; Lacour, J.-P.; Passeron, T. Interleukin 6 and high-sensitivity C-reactive protein are potential predictive markers of response to infliximab in hidradenitis suppurativa. J. Am. Acad. Dermatol. 2017, 76, 156–158. [Google Scholar] [CrossRef] [Green Version]
- Matusiak, L.; Szczech, J.; Bieniek, A.; Nowicka-Suszko, D.; Szepietowski, J.C. Increased interleukin (IL)-17 serum levels in patients with hidradenitis suppurativa: Implications for treatment with anti-IL-17 agents. J. Am. Acad. Dermatol. 2017, 76, 670–675. [Google Scholar] [CrossRef]
- Schlapbach, C.; Hanni, T.; Yawalkar, N.; Hunger, R.E. Expression of the IL-23/Th17 pathway in lesions of hidradenitis suppurativa. J. Am. Acad. Dermatol. 2011, 65, 790–798. [Google Scholar] [CrossRef]
- Thomi, R.; Kakeda, M.; Yawalkar, N.; Schlapbach, C.; Hunger, R.E. Increased expression of the interleukin-36 cytokines in lesions of hidradenitis suppurativa. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 2091–2096. [Google Scholar] [CrossRef]
- Hayran, Y.; Allı, N.; Yücel, Ç.; Akdoğan, N.; Turhan, T. Serum IL-36alpha, IL-36beta, and IL-36gamma levels in patients with hidradenitis suppurativa: Association with disease characteristics, smoking, obesity, and metabolic syndrome. Arch. Dermatol. Res. 2020, 312, 187–196. [Google Scholar] [CrossRef]
- Akdogan, N.; Alli, N.; Uysal, P.I.; Topcuoglu, C.; Candar, T.; Turhan, T. Visfatin and insulin levels and cigarette smoking are independent risk factors for hidradenitis suppurativa: A case-control study. Arch. Dermatol. Res. 2018, 310, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Wolk, K.; Witte, E.; Tsaousi, A.; Witte, K.; Volk, H.; Sterry, W.; Wenzel, J.; Schneider-Burrus, S.; Sabat, R. Lipocalin-2 as a novel biomarker in acne inversa. J. Investig. Dermatol. 2016, 136, S235. [Google Scholar] [CrossRef]
- Blok, J.L.; Li, K.; Brodmerkel, C.; Horvátovich, P.; Jonkman, M.F.; Horváth, B. Ustekinumab in hidradenitis suppurativa: Clin-ical results and a search for potential biomarkers in serum. Br. J. Dermatol. 2016, 174, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Tsaousi, A.; Witte, E.; Witte, K.; Rowert-Huber, H.J.; Volk, H.D.; Sterry, W.; Wolk, K.; Schneider-Burrus, S.; Sabat, R. MMP8 is increased in lesions and blood of acne inversa patients: A potential link to skin destruction and metabolic alterations. Mediators Inflamm. 2016, 2016, 4097574. [Google Scholar] [CrossRef]
- Wang, B.; Yang, W.; Wen, W.; Sun, J.; Su, B.; Liu, B.; Ma, D.; Lv, D.; Wen, Y.; Qu, T.; et al. Gamma-secretase gene mutations in familial acne inversa. Science 2010, 330, 1065. [Google Scholar] [CrossRef]
- Hunger, R.E.; Surovy, A.M.; Hassan, A.S.; Braathen, L.R.; Yawalkar, N. Toll-like receptor 2 is highly expressed in lesions of acne inversa and colocalizes with C-type lectin receptor. Br. J. Dermatol. 2008, 158, 691–697. [Google Scholar] [CrossRef]
- Sartorius, K.; Emtestam, L.; Lapins, J.; Johansson, O. Cutaneous PGP 9.5 distribution patterns in hidradenitis suppurativa. Arch. Dermatol. Res. 2010, 302, 461–468. [Google Scholar] [CrossRef]
- Fabregat, A.; Sidiropoulos, K.; Viteri, G.; Forner, O.; Marin-Garcia, P.; Arnau, V.; D’Eustachio, P.; Stein, L.; Hermjakob, H. Reactome pathway analysis: A high-performance in-memory approach. BMC Bioinform. 2017, 18, 142. [Google Scholar] [CrossRef] [Green Version]
- Frew, J.W.; Hawkes, J.E.; Krueger, J.G. A systematic review and critical evaluation of immunohistochemical associations in hidradenitis suppurativa. F1000Research 2018, 7, 1923. [Google Scholar] [CrossRef] [PubMed]
- Vossen, A.R.J.V.; van der Zee, H.H.; Prens, E.P. Hidradenitis suppurativa: A systematic review integrating inflammatory pathways into a cohesive pathogenic model. Front. Immunol. 2018, 9, 2965. [Google Scholar] [CrossRef] [Green Version]
- Jenei, A.; Dajnoki, Z.; Medgyesi, B.; Gáspár, K.; Béke, G.; Kinyó, Á.; Méhes, G.; Hendrik, Z.; Dinya, T.; Törőcsik, D.; et al. Apocrine gland-rich skin has a non-inflammatory IL-17 related immune milieu, which turns to inflammatory IL-17 mediated disease in hidradenitis suppurativa. J. Investig. Dermatol. 2019, 139, 964–968. [Google Scholar] [CrossRef] [Green Version]
- Bartlett, H.S.; Million, R.P. Targeting the IL-17-T(H)17 pathway. Nat. Rev. Drug Discov. 2015, 14, 11–12. [Google Scholar] [CrossRef]
- Archer, N.K.; Adappa, N.D.; Palmer, J.N.; Cohen, N.A.; Harro, J.M.; Lee, S.K.; Miller, L.S.; Shirtliff, M.E. Interleukin-17A (IL-17A) and IL-17F are critical for antimicrobial peptide production and clearance of Staphylococcus aureus nasal colonization. Infect. Immun. 2016, 84, 3575–3583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzanetakou, V.; Kanni, T.; Giatrakou, S.; Katoulis, A.; Papadavid, E.; Netea, M.G.; Dinarello, C.A.; van der Meer, J.W.M.; Rigopoulos, D.; Giamarellos-Bourboulis, E.J. Safety and efficacy of anakinra in severe hidradenitis suppurativa: A randomized clinical trial. JAMA Dermatol. 2016, 152, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Mintoff, D.; Benhadou, F.; Pace, N.P.; Frew, J.W. Metabolic syndrome and hidradenitis suppurativa: Epidemiological, molecular, and therapeutic aspects. Int. J. Dermatol. 2021. [Google Scholar] [CrossRef]
- Salvator, H.; Grassin-Delyle, S.; Brollo, M.; Couderc, L.J.; Abrial, C.; Victoni, T.; Naline, E.; Devillier, P. Adiponectin inhibits the production of TNF-alpha, IL-6 and chemokines by human lung macrophages. Front. Pharmacol. 2021, 12, 718929. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, Q.; Yan, Y.; Sun, B.; Wang, Y.; Tang, L.; Wang, E.; Yu, J.; Corpuz Nogoy, K.M.; Li, X.; et al. Effect of ciglitazone on adipogenic transdifferentiation of bovine skeletal muscle satellite cells. J. Anim. Sci. Technol. 2021, 63, 934–953. [Google Scholar] [CrossRef]
- Fimmel, S.; Zouboulis, C.C. Comorbidities of hidradenitis suppurativa (acne inversa). Derm.-Endocrinol. 2010, 2, 9–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzellos, T.; Zouboulis, C.C. Review of comorbidities of hidradenitis suppurativa: Implications for daily clinical practice. Dermatol. Ther. 2020, 10, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Zouboulis, C.C.; Okun, M.M.; Prens, E.P.; Gniadecki, R.; Foley, P.A.; Lynde, C.; Weisman, J.; Gu, Y.; Williams, D.A.; Jemec, G.B.E. Long-term adalimumab efficacy in patients with moderate-to-severe hidradenitis suppurativa/acne inversa: 3-year results of a phase 3 open-label extention study. J. Am. Acad. Dermatol. 2019, 80, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Zouboulis, C.C. Adalimumab for the treatment of hidradenitis suppurativa/acne inversa. Expert Rev. Clin. Immunol. 2016, 12, 1015–1026. [Google Scholar] [CrossRef]
- Matusiak, L.; Bieniek, A.; Szepietowski, J.C. Acitretin treatment for hidradenitis suppurativa: A prospective series of 17 patients. Br. J. Dermatol. 2014, 171, 170–174. [Google Scholar] [CrossRef]
- Frew, J.W.; Navrazhina, K.; Grand, D.; Sullivan-Whalen, M.; Gilleaudeau, P.; Garcet, S.; Ungar, J.; Krueger, J.G. The effect of subcutaneous brodalumab on clinical disease activity in hidradenitis suppurativa: An open-label cohort study. J. Am. Acad. Dermatol. 2020, 83, 1341–1348. [Google Scholar] [CrossRef]
- Mortimer, P.S.; Dawber, R.P.; Gales, M.A.; Moore, R.A. A double-blind controlled cross-over trial of cyproterone acetate in females with hidradenitis suppurativa. Br. J. Dermatol. 1986, 115, 263–268. [Google Scholar] [CrossRef]
- Nikolakis, G.; Kyrgidis, A.; Zouboulis, C.C. Is there a role for antiandrogen therapy for hidradenitis suppurativa? A systematic review of published data. Am. J. Clin. Dermatol. 2019, 20, 503–513. [Google Scholar] [CrossRef]
- Van Straalen, K.R.; Tzellos, T.; Guillem, P.; Benhadou, F.; Cuenca-Barrales, C.; Daxhelet, M.; Daoud, M.; Efthymiou, O.; Giamarellos-Bourboulis, E.J.; Jemec, G.B.E.; et al. The efficacy and tolerability of tetracyclines and clindamycin plus rifampicin for the treatment of hidradenitis suppurativa: Results of a prospective European cohort study. J. Am. Acad. Dermatol. 2021, 85, 369–378. [Google Scholar] [CrossRef]
- Lee, R.A.; Dommasch, E.; Treat, J.; Sciacca-Kirby, J.; Chachkin, S.; Williams, J.; Shin, D.B.; Leyden, J.J.; Vittorio, C.; Gelfand, J.M. A prospective clinical trial of open-label etanercept for the treatment of hidradenitis suppurativa. J. Am. Acad. Dermatol. 2009, 60, 565–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, D.R.; Yankura, J.A.; Fogelberg, A.C.; Anderson, B.E. Treatment of hidradenitis suppurativa with etanercept injection. Arch. Dermatol. 2010, 146, 501–504. [Google Scholar] [CrossRef] [PubMed]
- Melendez-Gonzalez, M.D.M.; Hamad, J.; Sayed, C. Golimumab for the treatment of hidradenitis suppurativa in patients with previous TNF-alpha treatment failure. J. Investig. Dermatol. 2021, 141, 2975–2979. [Google Scholar] [CrossRef]
- Grant, A.; Gonzalez, T.; Montgomery, M.O.; Cardenas, V.; Kerdel, F.A. Infliximab therapy for patients with moderate to severe hidradenitis suppurativa: A randomized, double-blind, placebo-controlled crossover trial. J. Am. Acad. Dermatol. 2010, 62, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Verdolini, R.; Clayton, N.; Smith, A.; Alwash, N.; Mannello, B. Metformin for the treatment of hidradenitis suppurativa: A little help along the way. J. Eur. Acad. Dermatol. Venereol. 2013, 27, 1101–1108. [Google Scholar] [CrossRef]
- Prussick, L.; Rothstein, B.; Joshipura, D.; Saraiya, A.; Turkowski, Y.; Abdat, R.; Alomran, A.; Zancanaro, P.; Kachuk, C.; Dumont, N.; et al. Open-label, investigator-initiated, single-site exploratory trial evaluating secukinumab, an anti-interleukin-17A monoclonal antibody, for patients with moderate-to-severe hidradenitis suppurativa. Br. J. Dermatol. 2019, 181, 609–611. [Google Scholar] [CrossRef]
- Giamarellos-Bourboulis, E.J.; Argyropoulou, M.; Kanni, T.; Spyridopoulos, T.; Otto, I.; Zenker, O.; Guo, R.; Riedemann, N.C. Clinical efficacy of complement C5a inhibition by IFX-1 in hidradenitis suppurativa: An open-label single-arm trial in patients not eligible for adalimumab. Br. J. Dermatol. 2020, 183, 176–178. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Blood | Skin | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Gene | +/− | mRNA | Protein | +/− | mRNA | Protein | Name | Other skin disorders | HS comorbid disorders | Drugs |
| ADAM12 | + | [3,27] | ADAM Metallopeptidase Domain 12 | Down syndrome | ||||||
| ADIPOQ | − | [28] | − | [27] | Adiponectin | Glucose intolerance, metabolic syndrome | Piogitazone | |||
| AR | + | [3,33,34] | [35] | Androgen receptor | Polycystic ovary syndrome, alopecia | Androgen insensitivity syndrome | Cyproterone acetate, Flutamide, Nilutamide, Bicalutamide, 17α-Propionate, AZD3514 | |||
| BTK | +/(−) | [3,27,33,34] | Betacellulin | Squamous cell carcinoma | Cetuximab | |||||
| C3 | − | [27] | + | [30] | Complement C3 | Zinc, Zinc acetate | ||||
| C5AR1 | + | [3,30] | Complement C5a Receptor 1 | Hypersensitivity reaction type III disease | Compstatin, PMX 205, PMX 53, W 54011 | |||||
| CASP1 | + | [38,39] | Caspase 1 | Schnitzler syndrome | Familial Mediterranean fever | Minocyclin | ||||
| CCL18 | + | [27,30] | [43] | C-C Motif Chemokine Ligand 18 | Eczema | |||||
| CCL26 | + | [41] | + | [30] | C-C Motif Chemokine Ligand 26 | |||||
| CCR4 | + | [45] | + | [30,45] | C-C Motif Chemokine Receptor 4 | Mycosis fungoides, cutaneous T cell lymphoma, allergic contact dermatitis | ||||
| CD80 | + | [30,38] | CD80 Molecule | Abatacept, Belatacept | ||||||
| CHI3L1 | + | [49] | + | [50] | Chitinase 3-Like 1 | Erysipelas | ||||
| CSF1 | + | [3,33,34,40] | Colony-Stimulating Factor 1 | Rheumatoid arthritis | ||||||
| CXCL1 | + | [27,40,42,44,45] | [40] | C-X-C Motif Chemokine Ligand 1 | Kaposi sarcoma | Formic acid | ||||
| CXCL8 | + | [30,42,44] | [41] | C-X-C Motif Chemokine Ligand 8 | Melanoma | Simvastatin | ||||
| CXCL10 | − | [41] | + | [30] | [41] | C-X-C Motif Chemokine Ligand 10 | Eldelumab | |||
| CXCL13 | + | [30,42,45] | [26] | C-X-C Motif Chemokine Ligand 13 | T cell lymphoma | |||||
| CXCR5 | + | [30] | [26] | C-X-C Motif Chemokine Receptor 5 | T cell lymphoma | |||||
| DCD | − | [27,32,33] | [32] | Dermcidin | Netherton syndrome, tinea pedis | Basiliximab, Zinc sulfate | ||||
| DEFB4A | +/(−) | [3,27,30,32,39,44,45,46] | [3,53] | Defensin β 4A | Tinea corporis, oral candidiasis | |||||
| DEFB103B | + | [46,52] | Defensin β 103B | |||||||
| EGF | + | [3,33,34] | Epidermal Growth Factor | Cetuximab, AG 490, CGP 52411, Genistein, Zanubrutinib (receptor antagonist) | ||||||
| EPGN | + | [3,33,34] | Epithelial Mitogen | Seborrheic dermatitis | ||||||
| ERBB4 | − | [27,32] | Erb-B2 Receptor Tyrosine Kinase 4 | Gefitinib, Afatinib, Fostamatinib, AG 490, CGP 52411, Genistein | ||||||
| EREG | + | [3,33,34] | Epiregulin | |||||||
| GAS6 | +/(−) | [3,33,34] | Growth Arrest Specific 6 | Lupus erythematosus | ||||||
| GDNF | + | [3,33,34] | [36] | Glial Cell Derived Neurotrophic Factor | Chondroitin sulphate | |||||
| GJB2 | + | [3] | [3] | Gap Junction Protein β2 | Keratitis-Ichthyosis-Deafness Syndrome | Carbenoxolone disodium | ||||
| HBEGF | + | [3,33,34] | Heparin Binding EGF-Like Growth Factor | |||||||
| HGF | + | [3,33,34] | Hepatocyte Growth Factor | Dexamethasone, Neratinib, Erlotinib | ||||||
| HRG | + | [3,33,34] | Histidine-Rich Glycoprotein | Zinc sulfate | ||||||
| IFNA1 | + | [3,26,30,33,34] | Interferon α1 | Cryoblobulinemia | ||||||
| IFNG | + | [3,26,30,33,34,40,44,45,46] | Interferon γ | Oksalazine, Emapalumab, Glucosamine | ||||||
| IGF2 | + | [3,33,34] | Insulin-Like Growth Factor 2 | |||||||
| IGHD | + | [27,30] | Immunoglobulin Heavy Constant δ | |||||||
| IGHG3 | + | [27,30] | Immunoglobulin Heavy Constant γ3 (G3m Marker) | |||||||
| IGKV1D-13 | + | [27,30] | Immunoglobulin κ Variable 1D-13 | |||||||
| IGLV | + | [27,30] | Immunoglobulin λ Variable Cluster | |||||||
| IL1A | + | [3,26,30,33,34,40] | [39] | Interleukin 1α | Acne, Irritant dermatitis | Arthritis | Anakinra, Rinolacept, Olanzapine, Pirfenidone, Thalidomide, AMG-108 | |||
| IL1B | + | [26,30,38,40,42,46] | [38,56] | Interleukin 1β | Gingivitis, Muckle–Wells syndrome, Toxic shock syndrome | Canakizumab, Anakinra (receptor antagonist), Rinolacept (receptor antagonist), Minocycline | ||||
| IL2 | + | [26,30] | Interleukin 2 | Graft-versus-host disease, Leprosy | Suplatast tosylate, Daclizumab (receptor antagonist), Basiliximab (receptor antagonist), Rituxomab, Thalidomide, Cafazolin | |||||
| IL2RA | + | [49,56] | + | [30] | Interleukin 2 Receptor Subunit α | Type 1 diabetes mellitus, Juvenile arthritis | Daclizumab, Basiliximab, Pirfenidone, Thalidomide | |||
| IL4 | + | [3,30,33,34,40] | Interleukin 4 | Atopy, Allergic rhinitis, Food allergy | Dupilumab (receptor antagonist), Calcitriol | |||||
| IL6 | + | [40] | + | [3,26,30,33,34,40,42] | [40,58] | Interleukin 6 | Siltuximab, Tocilizumab (receptor antagonist), Sarilumab (receptor antagonist), Satralizumab (receptor antagonist), Vitamin C, Vitamin E | |||
| IL10 | + | [30,38,44,46] | [52,56] | Interleukin 10 | Nicotinamide, Niacin, Cyclosporine A, Methotrexate, Mycofenolate mofetil | |||||
| IL12A | + | [59] | [41] | Interleukin 12A | Adamantiades–Behçet’s disease | Primary biliary cholangiitis | Mycophenolate mofetil, Ustekinumab (IL-12/23), Briakinumab (IL-12/23) | |||
| IL12B | + | [30] | [36] | Interleukin 12B | Psoriasis | Ustekinumab (IL-12/23), Briakinumab (IL-12/23) | ||||
| IL13 | +/(−) | [3,30,45] | Interleukin 13 | Allergic rhinitis, Penicillin allergy | Suplatast tosylate, Montelukast, Omalizumab | |||||
| IL16 | + | [30] | [41] | Interleukin 16 | Allergic asthma | |||||
| IL17A | + | [59] | + | [3,30,33,34,38,39,40,42,44,46,60] | [4,36,38,39,41] | Interleukin 17A | Allergic contact dermatitis | Arthritis | Secukizumab, Ixekizumab, Bimekizumab (IL-17A/F), Brodalumab (receptor antagonist), Vidofludimus | |
| IL17F | + | [30,39,40,42,45] | Interleukin 17F | Candidiasis, Acute generalized exanthematous pustulosis, Mail diseases | Bimekizumab (IL-17A/F), Brodaluman (receptor antagonist) | |||||
| IL17R | + | [3] | [4] | Interleukin 17 Receptor | Candidiasis | Arthritis | Brodalumab | |||
| IL18 | +/− | [26,30] | [38] | Interleukin 18 | IAP antagonist, Iboctadekin + Doxil | |||||
| IL19 | + | [3,30,40] | Interleukin 19 | Psoriasis | Inflammatory bowel disease, Arthritis | |||||
| IL20 | +/− | [30,46] | [46] | Interleukin 20 | Psoriasis | |||||
| IL21 | + | [30,39] | Interleukin 21 | Dacryoadenitis, Inflammatory boel disease | ||||||
| IL22 | +/(−) | [3,30,40,42,46] | [46] | Interleukin 22 | Candidiasis | Inflammatory bowel disease | ||||
| IL22RA1 | − | [30] | [46] | Interleukin 22 Receptor Subunit α1 | Spondyloarthropathy, rheumatoid arthritis, autoimmune uveitis | |||||
| IL23A | + | [30,40,61] | Interleukin 23 Subunit α | Autoimmune disease | Inflammatory bowel disease, Arthritis | Guselkumab, Risankinumab, Tildrakizumab, Ustekinumab (IL-12/23), Briakinumab (IL-12/23) | ||||
| IL24 | + | [30,42,46] | Interleukin 24 | Melanoma, chronic spontaneous urticaria, psoriasis | Spondylarthropathy | |||||
| IL26 | + | [42,46] | Interleukin 26 | Psoriasis | Inflammatory bowel disease, Crohn’s disease | |||||
| IL32 | + | [30,40,61] | Interleukin 32 | Cutaneous diphtheria | ||||||
| IL36A | + | [62] | + | [30,40,42,45,61] | [39,61] | Interleukin 36α | Psoriasis | Spesolimab (receptor antagonist) | ||
| IL36B | + | [62] | + | [61] | Interleukin 36β | Periostitis | Spesolimab (receptor antagonist) | |||
| IL36G | + | [62] | + | [30,40,42,45] | [61] | Interleukin 36γ | Acute generalized exanthematous pustulosis, Psoriasis | Spesolimab (receptor antagonist) | ||
| IL37 | − | [32,33,42] | Interleukin 37 | Still’s disease | Inflammatory bowel disease | Ustekinumab (IL-12/23) | ||||
| JAK3 | + | [3,30] | Janus Kinase 3 | NK cell enteropathy | Decernatinib, Tofacitinib (JAK1/3), Ruxolitinib (JAK1/3), PF-06651600, AT-501, ATI-502, Cerdulatinib (JAK1/2/3, SYK), Delgocitinib (JAK1/2/3), Peficitinib (JAK1/2/3), Zanubrutinib (JAK3/ITR/EGFR), Cercosporamide JAK3/Mnk2) | |||||
| KRT6A | + | [3,32] | [3] | Keratin 6A | Pachyonychia congenita, Lingua plicata, Cheilitis | Zinc, Zinc acetate | ||||
| KRT16 | + | [3,27,30,32] | [3] | Keratin 16 | Pachyonychia congenita, palmoplantar keratoderma | |||||
| KRT77 | − | [27,32,33] | [32] | Keratin 77 | Epidermolytic palmoplantar keratoderma, Buschke-Ollendorff syndrome | |||||
| LCE3D | + | [32] | [32] | Late Cornified Envelope 3D | Psoriasis | |||||
| LGR5 | − | [27,32] | Leucine Rich Repeat Containing G Protein-Coupled Receptor 5 | Type II diabetes mellitus | ||||||
| LTA4H | − | [27,65] | + | [31] | Leukotriene A4 Hydrolase | Captopril, Dexamethasone, Montelukast | ||||
| MMP1 | + | [3,30] | [3] | Matrix Metallopeptidase 1 | Epidermolysis bullosa atrophica, Scleroderma | Zinc, Collagenase | ||||
| MMP3 | + | [40] | [40] | Matrix Metallopeptidase 3 | Coronary heart disease, Arthritis | Pravastatin, Simvastatin, Prothalidone, Lisinopril | ||||
| MMP9 | + | [3,30,40] | [3] | Matrix Metallopeptidase 9 | Minocycline, Capropril, Simvastatin, Zinc, Zinc acetate | |||||
| MMP12 | + | [27,30] | Matrix Metallopeptidase 12 | Dermatitis herpetiformis, Middermal elastolysis | Arthritis | Acetohydroxamic acid, Batimastat | ||||
| NAMPT | + | [28,63] | Nicotinamide Phosphoribosyl transferase | Skin aging, pellagra, diabetes mellitus type 2, polycystic ovary syndrome | Nicotinamide, Niacin | |||||
| NGF | + | [3,33,34] | [36] | Nerve Growth Factor | Clenbuterol | |||||
| OSM | + | [3,26] | [36] | Oncostatin M | Kaposi sarcoma | |||||
| PI3 | + | [3,27,32,33] | [3] | Peptidase Inhibitor 3 | Pustular psoriasis, impetigo herpetiformis, erysipelas | |||||
| PIP | − | [27,32] | Prolactin Induced Protein | |||||||
| PLIN1 | +/− | [27,48] | Perilipin 1 | Rosiglitazone | ||||||
| S100A7 | + | [3,30,33,39,42,44,46] | [32] | S100 Calcium-Binding Protein A7 | Psoriasis, Squamous cell carcinoma | Anal fistula | Ibuprofen, Dexibuprofen, Zinc, Zinc acetate, Zinc chloride | |||
| S100A7A | + | [3,27,32] | [3,32] | S100 Calcium-Binding Protein A7A | Psoriasis | |||||
| S100A8 | + | [57] | + | [3,33,34,44] | [3,32] | S100 Calcium-Binding Protein A8 | Zinc, Zinc acetate, Zinc chloride, Copper | |||
| S100A9 | + | [57] | + | [3,27,32,33,42,44,46] | [3,32] | S100 Calcium-Binding Protein A9 | Crohn’s disease, Rheumatoid arthritis | Zinc, Zinc acetate, Zinc chloride, Calcium | ||
| S100A12 | + | [3,30,32,42] | [3,41] | S100 Calcium-Binding Protein A12 | Kawasaki disease | Psoriatic arthritis | Amlexanox, Olopatadine | |||
| SCGB1D2 | − | [27,32] | Secretoglobin Family 1D Member 2 | |||||||
| SCGB2A2 | − | [27,32,33] | Secretoglobin Family 2A Member 2 | |||||||
| SERPINB3 | + | [3,27,30] | [3] | Serpin Family B Member 3 | Squamous cell caecinoma | Phosphoserine | ||||
| SERPINB4 | + | [3,27,30] | [3] | Serpin Family B Member 4 | Squamous cell carcinoma | |||||
| SLAMF7 | + | [3,27] | SLAM Family Member 7 | IgG4-related disease | Elotuzumab | |||||
| SPRR2B | + | [32] | [32] | Small Proline Rich Protein 2B | Photosensitive trichothio-dystrophy 1, Autosomal reces-sive congenital ichthyosis | |||||
| SPRR2C (pseudogene) | + | [32] | [32] | Small Proline Rich Protein 2C (Pseudogene) | ||||||
| SPRR3 | + | [3] | [3] | Small Proline Rich Protein 3 | Genodermatoses | |||||
| STAT1 | + | [3,26,30,44] | [36] | Signal Transducer and Activator of Transcription 1 | Methimazole, Niclosamide, Nifuroxazide, Sulforaphane | |||||
| TCN1 | + | [3,27,45] | [3] | Transcobalamin 1 | Hydroxycobalamin, Cyanocobalamin, Cobalt | |||||
| TLR2 | + | [3,68] | Toll-Like Receptor 2 | Leprosy, Borreliosis | Colorectal cancer | Adapalene, Cyproterone acetate | ||||
| TLR4 | +/− | [26] | [53] | Toll-like Receptor 4 | Paclitaxel, Tacrolimus, Cyclobenzaprine | |||||
| TMPRSS1D | + | [3] | [3] | Transmembrane Serine Protease 11D | ||||||
| TNF | + | [3,26,30,32,33,38,40] | [56] | Tumor Necrosis Factor | Psoriasis, Toxic shock syndrome | Inflammatory bowel diseases, Arthritis | Adalimumab, Infliximab, Golimumab, Etanercept (receptor antagonist), Certolizumab pegol, Thalidomide, Lenalidomide, Pomalidomide, Calcitriol, Bay 11-7821, (R)-DOI, Cannabidiol | |||
| TNFRSF4 | + | [45] | + | [45] | TNF Receptor Superfamily Member 4 | Kaposi sarcoma, Graft-versus-host disease, Drug reaction with eosinophilia | OX-40 ligand | |||
| TNFSF11 | + | [30] | [36] | TNF Superfamily Member 11 | Letrozole, Thiocolchicoside | |||||
| TNFSF13 (APRIL) | + | [30] | [26] | TNF Superfamily Member 13 | Autoimmune diseases | Rheumatoid arthritis | Pomalidomide, TACI-IG | |||
| TNFSF13B (BAFF) | + | [30] | [26] | TNF Superfamily Member 13b | Autoimmune diseases, Sialadenitis, Sjogren syndrome | Belimumab, Blisibimod, LY2127399, TACI-IG | ||||
| TNFSF14 | + | [30] | [36] | TNF Superfamily Member 14 | Herpes simplex | Rheumatoid arthritis | ||||
| TNIP1 | +/− | [26,30] | TNFAIP3 Interacting Protein 1 | Systemic lupus erythematosus, Psoriatic arthritis | Rheumatoid arthritis, Arthritis | |||||
| WIF1 | − | [27,32] | WNT Inhibitory Factor 1 | |||||||
| Compound | Function | Gene Regulation | Development Phase |
|---|---|---|---|
| Probable repurposing HS drugs | |||
| 3,3’-Diindolylmethane | CHK inhibitor, cytochrome P450 activator, indoleamine 2,3-dioxygenase inhibitor | AR, HIF1A, IFNG, PI3 | 3 |
| AG-490 | EGFR inhibitor, JAK inhibitor | EGFR, JAK2, JAK3 | preclinical |
| Andrographolide | tumor necrosis factor production inhibitor | IL1B, IL6, NFKB1, NFKB2, TNF | 2 |
| Apratastat | matrix metalloprotease inhibitor, tumor necrosis factor production inhibitor | ADAM17, MMP1, MMP13, MMP9 | 2 |
| Atractylenolide-I | JAK inhibitor | JAK1, JAK2, JAK3 | preclinical |
| AZD1480 | JAK inhibitor | JAK1, JAK2, JAK3 | 1 |
| Balsalazide | cyclooxygenase inhibitor | ALOX5, PPARG, PTGS1, PTGS2 | launched |
| BMS-911543 | JAK inhibitor | JAK1, JAK2, JAK3 | 1/2 |
| Ciglitazone | PPARγ agonist | GPD1, PPARG, TBXA2R | 2 |
| Curcumol | JAK inhibitor | JAK1, JAK2, JAK3 | 1 |
| Cyt387 | JAK inhibitor | JAK1, JAK2, JAK3 | 3 |
| Delgocitinib | JAK inhibitor | JAK1, JAK2, JAK3 | 2 |
| Fedratinib | FLT3 inhibitor, JAK inhibitor | BRD4, JAK1, JAK2, JAK3, TYK2 | launched |
| Filgotinib | JAK inhibitor | JAK1, JAK2, JAK3, TYK2 | 3 |
| Ganoderic-acid-a | JAK inhibitor | JAK1, JAK2, JAK3 | preclinical |
| JTE-607 | cytokine production inhibitor | IL10, IL1B, IL6, TNF | 2 |
| Latamoxef | Cephalosporine | DACB, MRCA, MRCB, PBPC | launched |
| LXR-623 | Liver X receptor agonist | AR, NR1H2, NR1H3, NR1I2, NR3C1 | 1 |
| NS-018 | JAK inhibitor | JAK1, JAK2, JAK3, TYK2 | 1/2 |
| Pacritinib | FLT3 inhibitor, JAK inhibitor | FLT3, JAK1, JAK2, JAK3 | 3 |
| Paracetamol | cyclooxygenase inhibitor | FAAH, PTGS1, PTGS2, TRPV1 | launched |
| Peficitinib | JAK inhibitor | JAK1, JAK2, JAK3 | launched |
| PF-06651600 | JAK inhibitor | JAK1, JAK2, JAK3 | 2/3 |
| Plerixafor | CC chemokine receptor antagonist | ACKR3, CCR4, CXCR4, MMP1, PI3 | launched |
| Ruxolitinib | JAK inhibitor | JAK1, JAK2, JAK3, TYK2 | launched |
| Sirolimus | mTOR inhibitor | CFD1, FKBP1A, GPD1, MMP1, MTOR, PI3, RPL38 | launched |
| Tofacitinib | JAK inhibitor | JAK1, JAK2, JAK3 | launched |
| Trofinetide | cytokine production inhibitor | IFNG, IL6, TNFA | 2 |
| Upadacitinib | JAK inhibitor | JAK1, JAK2, JAK3 | launched |
| WHI-P154 | JAK inhibitor | EGFR, JAK1, JAK2, JAK3 | preclinical |
| XL019 | JAK inhibitor | JAK1, JAK2, JAK3 | 1 |
| Drugs with known molecular profile registered ** or off-label administered in HS | |||
| Acitretin | retinoid receptor agonist | KRT16, PI3, RARA, RARB, RARG, RBP1, RXRA, RXRB, RXRG, STAT3 | launched |
| Adalimumab ** | TNF-α inhibitor | TNF | launched |
| Anakinra | IL-1 receptor antagonist | IL1R1 | launched |
| Avacopan | C5α receptor antagonist | C5AR1 | 2 |
| Bimekizumab | IL-17A/F inhibitor | IL17A, IL17F | 3 |
| Brodalumab | IL-17 receptor inhibitor | IL17R, KRT6A, S100A7A, S100A8, S100A9 | launched |
| Clindamycin | Protein synthesis inhibitor | launched | |
| Cyproterone acetate | AR antagonist | ADORA1, AR | launched |
| Doxycycline | bacterial 30S ribosomal subunit inhibitor, metalloproteinase inhibitor | MMP1, MMP8, PI3 | launched |
| Etanercept | TNF-α receptor antagonist | TNFRSF1A | launched |
| Golimumab | TNF inhibitor | TNF | launched |
| INCB 54707 | JAK1 inhibitor | JAK1 | 2 |
| Infliximab | TNF inhibitor | IL6, TNF | launched |
| Metformin | insulin sensitizer | ACACB, PRKAB1 | launched |
| Rifampicin | RNA polymerase inhibitor | NR1I2, SLCO1A2, SLCO1B1, SLCO1B3 | launched |
| Secukinumab | IL-17A inhibitor | IL17A | 3 |
| Spesolimab | IL-36R antagonist | IL36RN | 2 |
| Ustekinumab | IL12/IL23 inhibitor | FSH, HCG, LH, LTA4H | Launched |
| Vilobelimab | C5α inhibitor | C5 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zouboulis, V.A.; Zouboulis, K.C.; Zouboulis, C.C. Hidradenitis Suppurativa and Comorbid Disorder Biomarkers, Druggable Genes, New Drugs and Drug Repurposing—A Molecular Meta-Analysis. Pharmaceutics 2022, 14, 44. https://doi.org/10.3390/pharmaceutics14010044
Zouboulis VA, Zouboulis KC, Zouboulis CC. Hidradenitis Suppurativa and Comorbid Disorder Biomarkers, Druggable Genes, New Drugs and Drug Repurposing—A Molecular Meta-Analysis. Pharmaceutics. 2022; 14(1):44. https://doi.org/10.3390/pharmaceutics14010044
Chicago/Turabian StyleZouboulis, Viktor A., Konstantin C. Zouboulis, and Christos C. Zouboulis. 2022. "Hidradenitis Suppurativa and Comorbid Disorder Biomarkers, Druggable Genes, New Drugs and Drug Repurposing—A Molecular Meta-Analysis" Pharmaceutics 14, no. 1: 44. https://doi.org/10.3390/pharmaceutics14010044