
Effect of a Fiber D-Limonene-Enriched Food Supplement on Intestinal Microbiota and Metabolic Parameters of Mice on a High-Fat Diet

 ,
,  , , , ,
, , , ,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. D-Limonene and FLS Pharmacokinetic Analysis
2.3. Animal Treatment
2.4. Blood Lipidic Profile and Glucose Measurement
2.5. Determination of Plasma Chemokine Levels and Hormonal Profile
2.6. Histological Evaluation of Hepatic Steatosis and Lipid Profile
2.7. Hepatic Tissue Lipid Profile (Lipidomic Analysis)
2.8. Characterization of the Fecal Microbiota
2.9. Statistical Analysis
3. Results
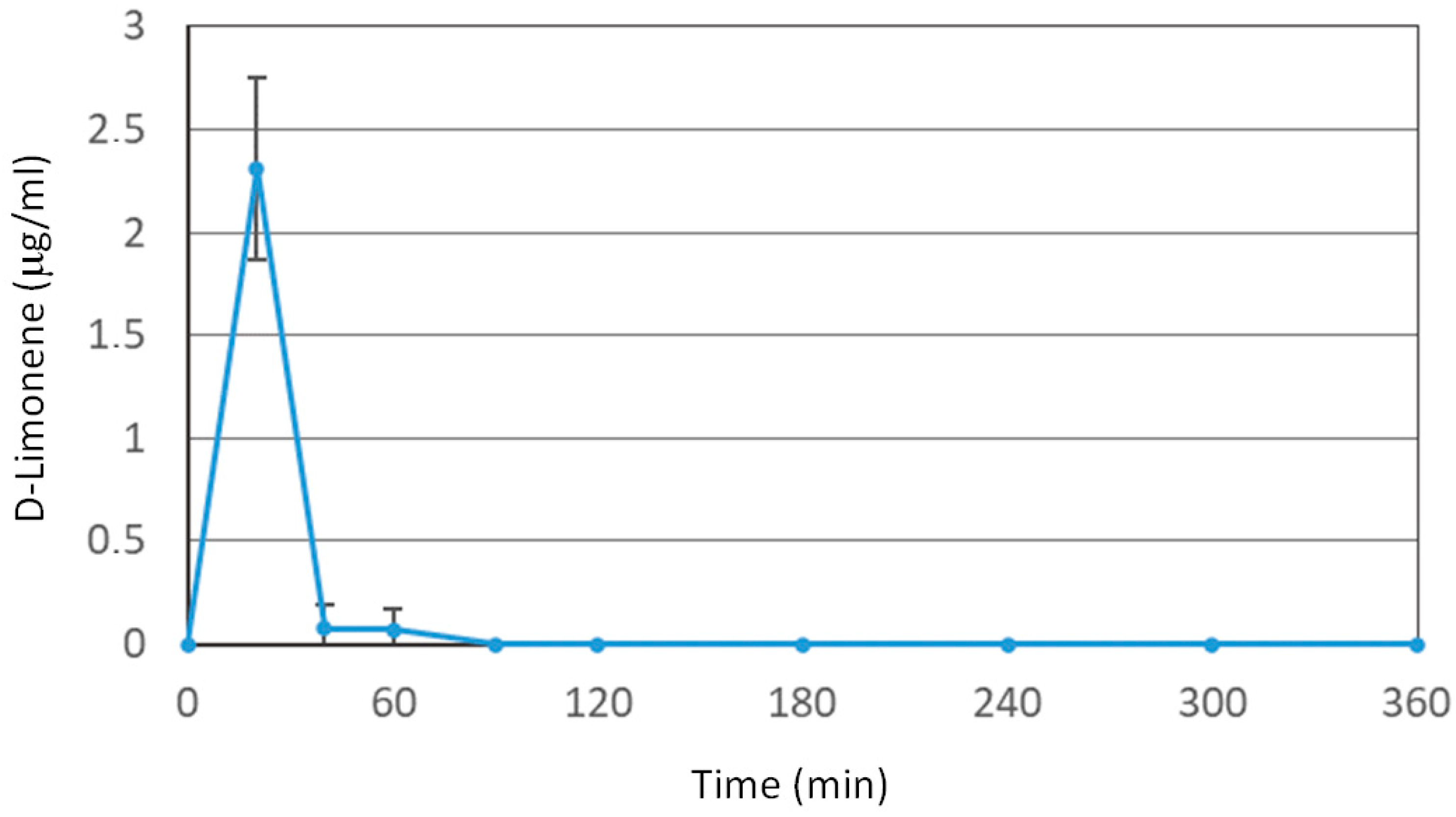
3.1. D-Limonene and FLS Pharmacokinetic Analysis
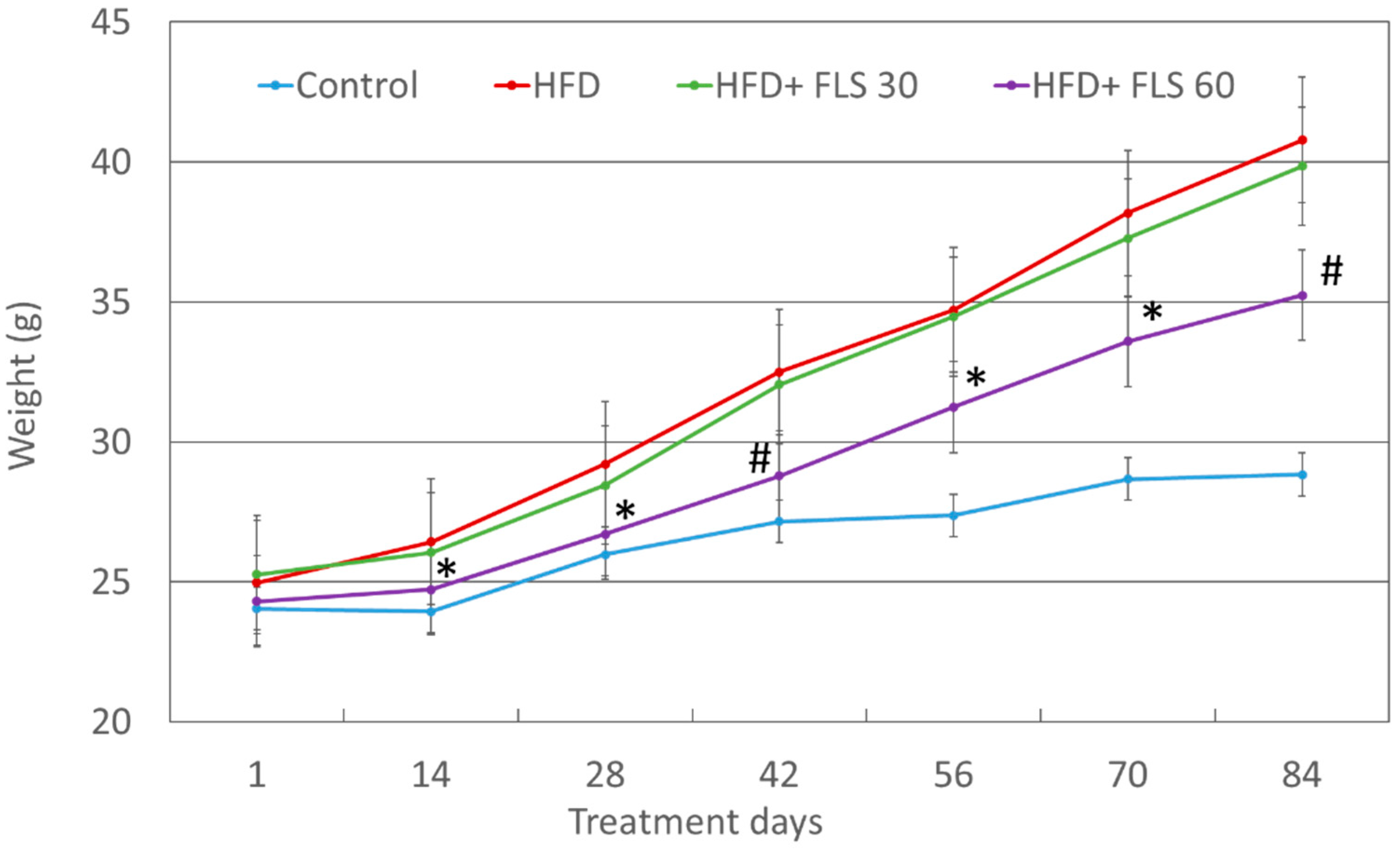
3.2. Effect of FLS on Weight Gain
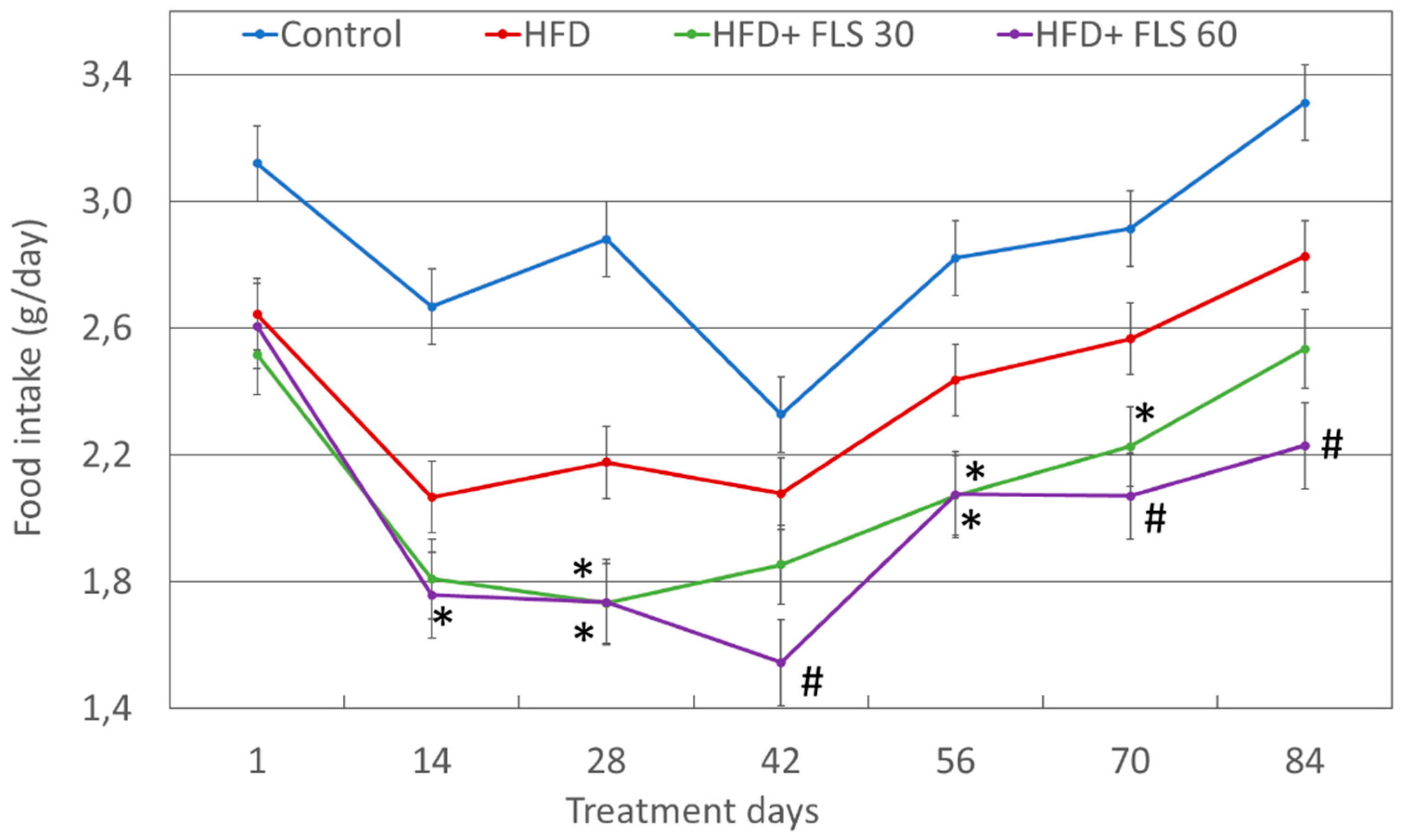
3.3. Effect of FLS on Food Intake
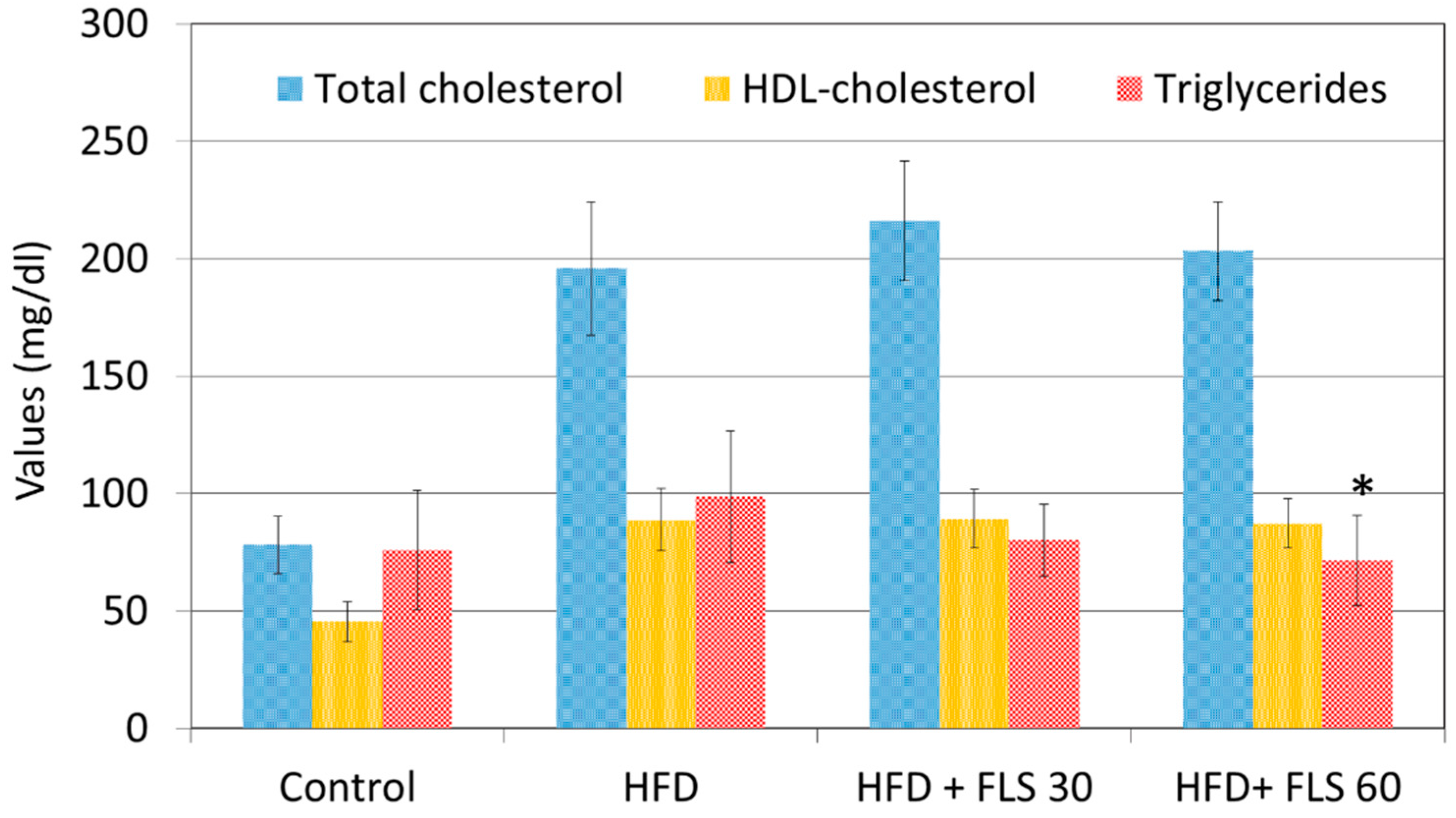
3.4. Effect of FLS on Lipid Profile
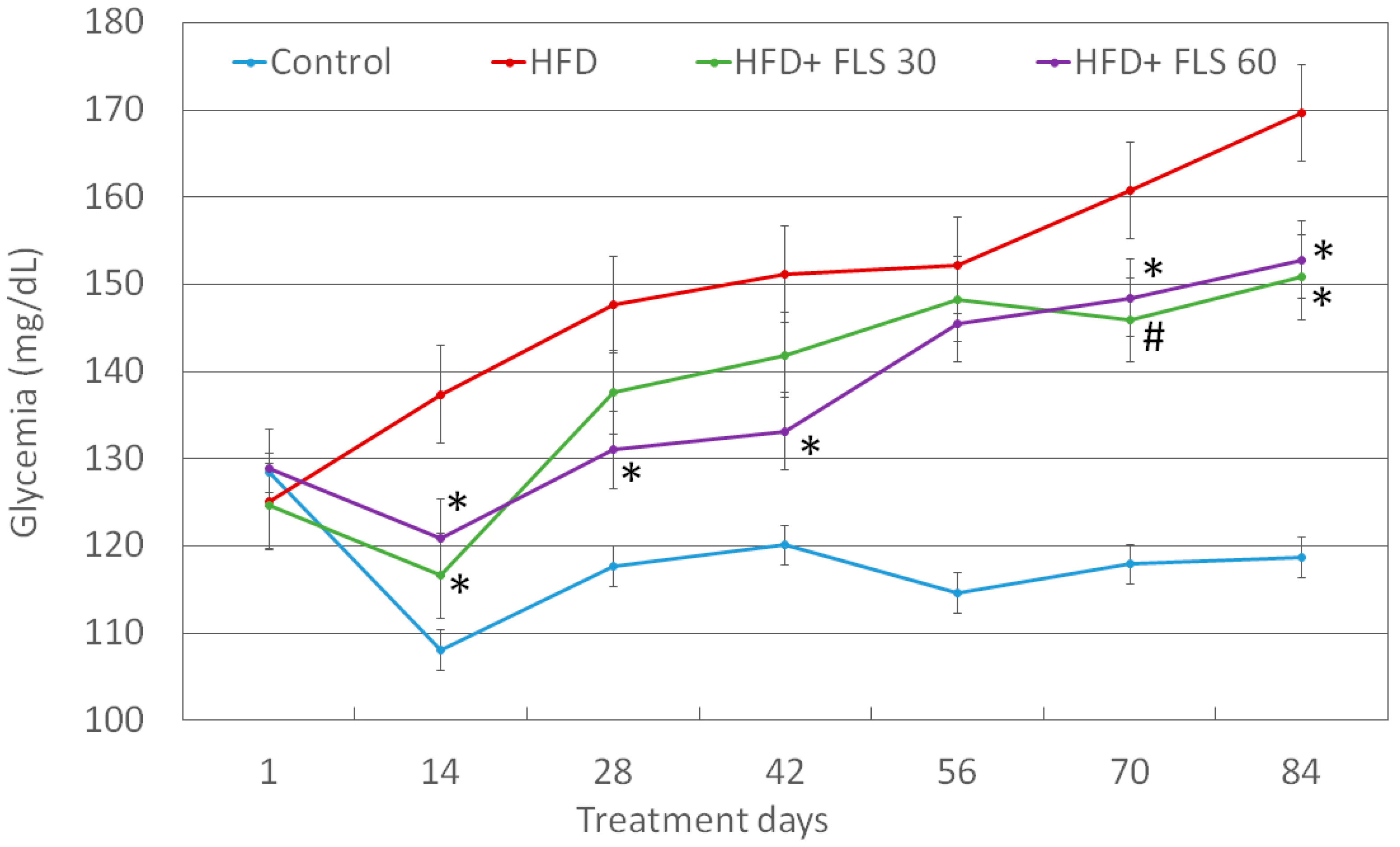
3.5. Effect of FLS on Fasting Glycemia
3.6. Effect of FLS on Circulating Cytokines, Adipokines and Hormones
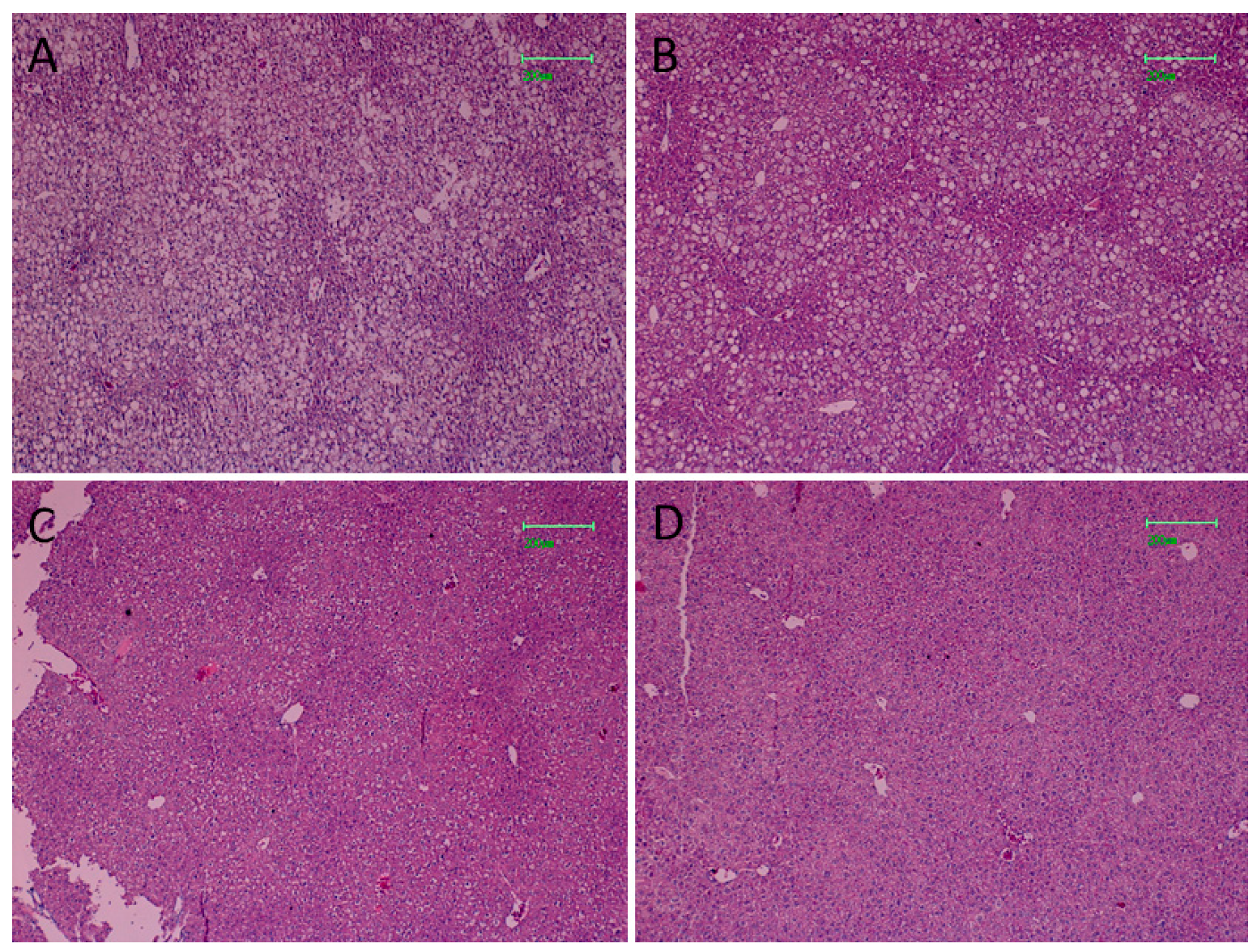
3.7. Effect of FLS on Hepatic Steatosis of HFD Mice
3.8. Effect of FLS on Hepatic Lipids of HFD Mice
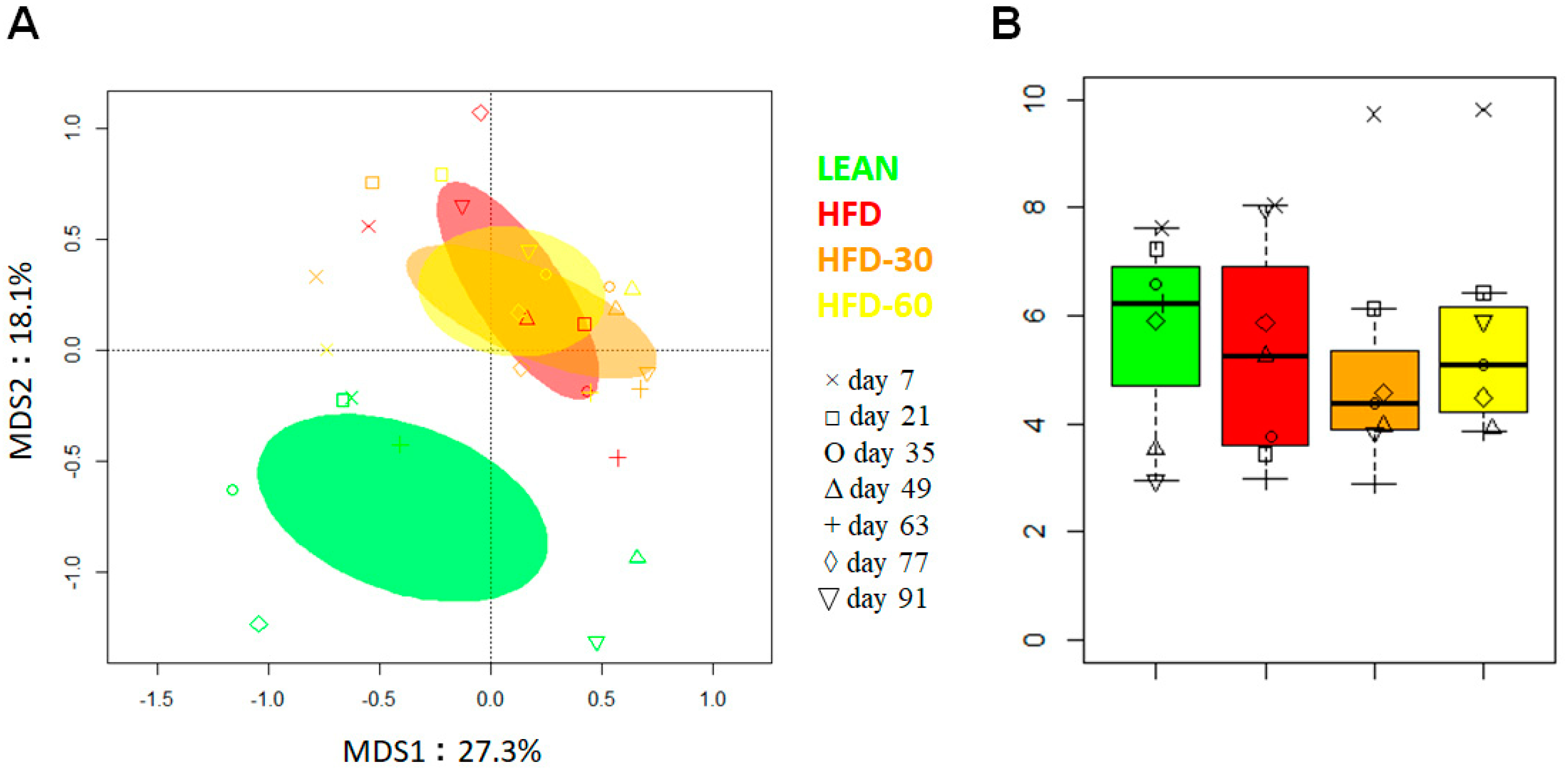
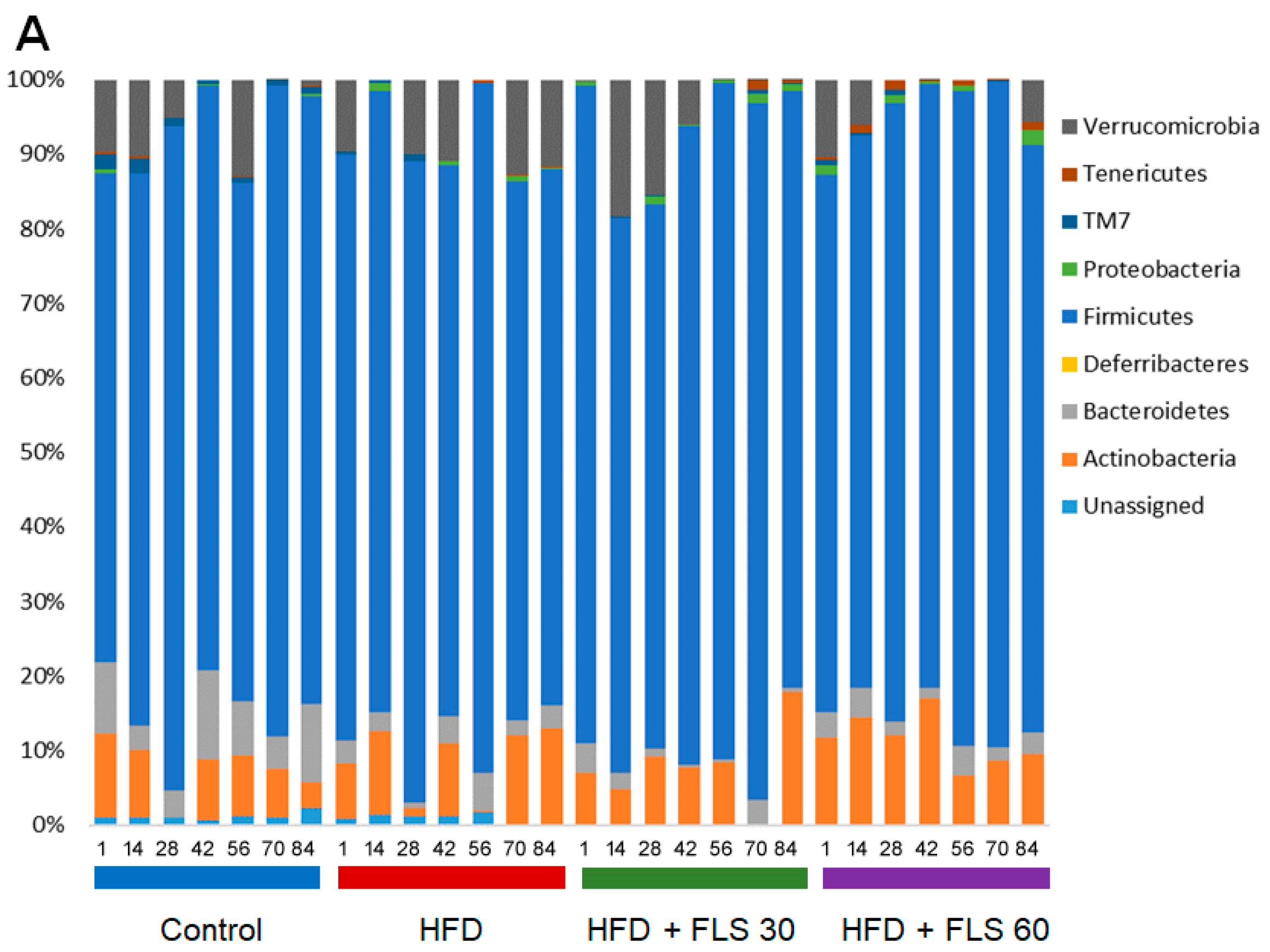
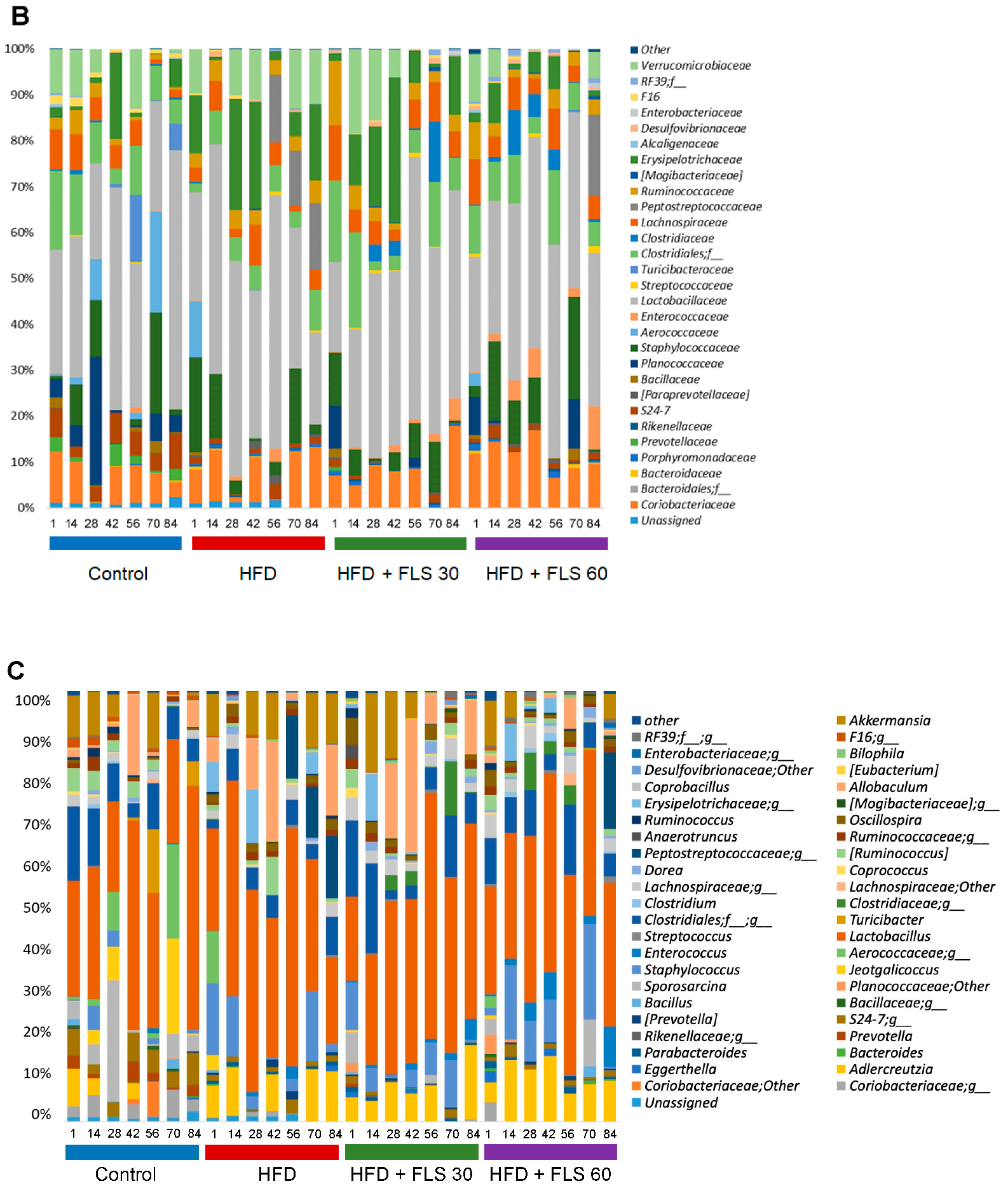
3.9. Effect of FLS on Fecal Microbiota Composition in HFD Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Blüher, M. Obesity: Global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Berthoud, H.R.; Klein, S. Advances in obesity: Causes, consequences, and therapy. Gastroenterology 2017, 152, 1635–1637. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.D.; Ryan, D.H.; Apovian, C.M.; Ard, J.D.; Comuzzie, A.G.; Donato, K.A.; Hu, F.B.; Hubbard, V.S.; Jakicic, J.M.; Kushner, R.F.; et al. 2013 AHA/ACC/TOS guideline for the management of overweight and obesity in adults: A report of the American College of Cardiology/American Heart Association Task Force on Practice Guidelines and The Obesity Society. J. Am. Coll. Cardiol. 2014, 63, 2985–3023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef] [Green Version]
- Cani, P.D.; Van Hul, M. Mediterranean diet, gut microbiota and health: When age and calories do not add up! Gut 2020, 69, 1167–1168. [Google Scholar] [CrossRef] [Green Version]
- Bäckhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The gut microbiota as an environmental factor that regulates fat storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Festi, D.; Schiumerini, R.; Eusebi, L.H.; Marasco, G.; Taddia, M.; Colecchia, A. Gut microbiota and metabolic syndrome. World J. Gastroenterol. 2014, 20, 16079–16094. [Google Scholar] [CrossRef]
- Santos-Paulo, S.; Costello, S.; Forster, S.; Travis, S.; Bryant, R. The gut microbiota as a therapeutic target for obesity: A scoping review. Nutr. Res. Rev. 2021, 8, 1–33. [Google Scholar] [CrossRef]
- Waltenberger, B.; Mocan, A.; Šmejkal, K.; Heiss, E.H.; Atanasov, A.G. Natural products to counteract the epidemic of cardiovascular and metabolic disorders. Molecules 2016, 21, 807. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, Y.; Fan, G.; Ren, J.N.; Zhang, L.L.; Pan, S.Y. Effects of orange essential oil on intestinal microflora in mice. J. Sci. Food Agric. 2019, 99, 4019–4028. [Google Scholar] [CrossRef]
- Li, D.; Wu, H.; Dou, H. Weight loss effect of sweet orange essential oil microcapsules on obese SD rats induced by high-fat diet. Biosci. Biotechnol. Biochem. 2019, 83, 923–932. [Google Scholar] [CrossRef]
- Victor Antony Santiago, J.; Jayachitra, J.; Shenbagam, M.; Nalini, N. Dietary d-limonene alleviates insulin resistance and oxidative stress-induced liver injury in high-fat diet and L-NAME-treated rats. Eur. J. Nutr. 2012, 51, 57–68. [Google Scholar] [CrossRef] [PubMed]
- EFSA FEEDAP Panel (EFSA Panel on Additives and Products or Substances used in Animal Feed). Scientific Opinion on the safety and efficacy of aliphatic and aromatic hydrocarbons (chemical group 31) when used as flavourings for all animal species. EFSA J. 2015, 13, 4053. [Google Scholar] [CrossRef]
- Sun, J. D-Limonene: Safety and clinical applications. Altern. Med. Rev. 2007, 12, 259–264. [Google Scholar] [PubMed]
- Kim, Y.W.; Kim, M.J.; Chung, B.Y.; Bang du, Y.; Lim, S.K.; Choi, S.M.; Lim, D.S.; Cho, M.C.; Yoon, K.; Kim, H.S.; et al. Safety evaluation and risk assessment of d-Limonene. J. Toxicol. Environ. Health B Crit. Rev. 2013, 16, 17–38. [Google Scholar] [CrossRef]
- Nissen, L.; Valerii, M.C.; Spisni, E.; Casciano, F.; Gianotti, A. Multiunit in vitro colon model for the evaluation of prebiotic potential of a fiber D-Limonene enriched food supplement. Foods 2021, 10, 2371. [Google Scholar] [CrossRef]
- De Oliveira Junior, E.R.; Truzzi, E.; Ferraro, L.; Fogagnolo, M.; Pavan, B.; Beggiato, S.; Rustichelli, C.; Maretti, E.; Lima, E.M.; Leo, E.; et al. Nasal administration of nanoencapsulated geraniol/ursodeoxycholic acid conjugate: Towards a new approach for the management of Parkinson’s disease. J. Control. Release 2020, 321, 540–552. [Google Scholar] [CrossRef]
- Vignali, D.A. Multiplexed particle-based flow cytometric assays. J. Immunol. Methods 2000, 243, 243–255. [Google Scholar] [CrossRef]
- Jauregibeitia, I.; Portune, K.; Rica, I.; Tueros, I.; Velasco, O.; Grau, G.; Castaño, L.; Di Nolfo, F.; Ferreri, C.; Arranz, S. Potential of erythrocyte membrane lipid profile as a novel inflammatory biomarker to distinguish metabolically healthy obesity in children. J. Pers. Med. 2021, 11, 337. [Google Scholar] [CrossRef]
- Nichols, P.D.; Volkman, J.K.; Everitt, D.A. Occurrence of Cis-6-Hexadecenoic Acid and Other Unusual Monounsaturated Fatty Acids in the Lipids of Oceanic Particulate Matter. Oceanol. Acta 1989, 12, 393–403. Available online: https://archimer.ifremer.fr/doc/00106/21735/ (accessed on 25 May 2021).
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-end assembler for illumina sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-7. Available online: https://cran.r-project.org/package=vegan (accessed on 25 May 2021).
- Terzo, S.; Mulè, F.; Caldara, G.F.; Baldassano, S.; Puleio, R.; Vitale, M.; Cassata, G.; Ferrantelli, V.; Amato, A. Pistachio consumption alleviates inflammation and improves gut microbiota composition in mice fed a high-fat diet. Int. J. Mol. Sci. 2020, 21, 365. [Google Scholar] [CrossRef] [Green Version]
- Denisenko, Y.K.; Kytikova, O.Y.; Novgorodtseva, T.P.; Antonyuk, M.V.; Gvozdenko, T.A.; Kantur, T.A. Lipid-induced mechanisms of metabolic syndrome. J. Obes. 2020, 2020, 5762395. [Google Scholar] [CrossRef]
- Oteng, A.B.; Kersten, S. Mechanisms of action of trans fatty acids. Adv. Nutr. 2020, 11, 697–708. [Google Scholar] [CrossRef]
- Obara, N.; Fukushima, K.; Ueno, Y.; Wakui, Y.; Kimura, O.; Tamai, K.; Kakazu, E.; Inoue, J.; Kondo, Y.; Ogawa, N.; et al. Possible involvement and the mechanisms of excess trans-fatty acid consumption in severe NAFLD in mice. J. Hepatol. 2010, 53, 326–334. [Google Scholar] [CrossRef]
- Guirro, M.; Costa, A.; Gual-Grau, A.; Herrero, P.; Torrell, H.; Canela, N.; Arola, L. Effects from diet-induced gut microbiota dysbiosis and obesity can be ameliorated by fecal microbiota transplantation: A multiomics approach. PLoS ONE 2019, 14, e0218143. [Google Scholar] [CrossRef] [Green Version]
- Sonnenburg, J.L.; Erica, D.; Sonnenburg, E.D. Vulnerability of the industrialized microbiota. Science 2019, 366, 6464. [Google Scholar] [CrossRef] [Green Version]
- Kriss, M.; Hazleton, K.Z.; Nusbacher, N.M.; Martin, C.G.; Lozupone, C.A. Low diversity gut microbiota dysbiosis: Drivers, functional implications and recovery. Curr. Opin. Microbiol. 2018, 44, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.L.; Vieira-Silva, S.; Liston, A.; Raes, J. How informative is the mouse for human gut microbiota research? Dis. Model. Mech. 2015, 8, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safari, Z.; Bruneau, A.; Monnoye, M.; Mariadassou, M.; Philippe, C.; Zatloukal, K.; Gérard, P. Murine genetic background overcomes gut microbiota changes to explain metabolic response to high-fat diet. Nutrients 2020, 12, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Kong, Q.; Li, X.; Zhao, J.; Zhang, H.; Chen, W.; Wang, G. A high-fat diet increases gut microbiota biodiversity and energy expenditure due to nutrient difference. Nutrients 2020, 12, 3197. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Roberts, J.D.; Grimm, S.A.; Lih, F.B.; Deterding, L.J.; Li, R.; Chrysovergis, K.; Wade, P.A. An obesity-associated gut microbiome reprograms the intestinal epigenome and leads to altered colonic gene expression. Genome Biol. 2018, 9, 7. [Google Scholar] [CrossRef] [PubMed]
- Jing, L.; Zhang, Y.; Fan, S.; Gu, M.; Guan, Y.; Lu, X.; Huang, C.; Zhou, Z. Preventive and ameliorating effects of citrus D-limonene on dyslipidemia and hyperglycemia in mice with high-fat diet-induced obesity. Eur. J. Pharmacol. 2013, 715, 46–55. [Google Scholar] [CrossRef]
- Ramos, C.A.F.; Sá, R.C.D.S.; Alves, M.F.; Benedito, R.B.; de Sousa, D.P.M.; Diniz, M.F.F.M.; Araújo, M.S.T.; de Almeida, R.N. Histopathological and biochemical assessment of d-limonene-induced liver injury in rats. Toxicol. Rep. 2015, 2, 482–488. [Google Scholar] [CrossRef] [Green Version]
- Turroni, S.; Petracci, E.; Edefonti, V.; Giudetti, A.M.; D’Amico, F.; Paganelli, L.; Giovannetti, G.; Del Coco, L.; Fanizzi, F.P.; Rampelli, S.; et al. Effects of a diet based on foods from symbiotic agriculture on the gut microbiota of subjects at risk for metabolic syndrome. Nutrients 2021, 13, 2081. [Google Scholar] [CrossRef]
- Wang, D.; Dong, Y.; Wang, Q.; Wang, X.; Fan, W. Limonene, the compound in essential oil of nutmeg displayed antioxidant effect in sunflower oil during the deep-frying of chinese maye. Food Sci. Nutr. 2019, 8, 511–520. [Google Scholar] [CrossRef] [Green Version]
- Ibáñez, M.D.; Sanchez-Ballester, N.M.; Blázquez, M.A. Encapsulated limonene: A pleasant lemon-like aroma with promising application in the agri-food industry. A review. Molecules 2020, 25, 2598. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | STD | HFD |
|---|---|---|
| Total Energy, Kcal/g | 3.5 | 6 |
| Protein, % | 20 | 20 |
| Carbohydrate, % | 70 | 20 |
| Fat, % | 10 | 60 |
| Fatty Acids 1 | Control | HFD | HFD + FLS 30 | HFD + FLS 60 |
|---|---|---|---|---|
| 14:0 | 0.37 ± 0.05 | 0.47 ± 0.07 | 0.45 ± 0.03 | 0.49 ± 0.13 |
| 16:0 | 24.33 ± 0.90 | 24.25 ± 0.59 | 20.46 ± 0.35 ### | 22.07 ± 0.72 §, £ |
| 16:1 6t | 0.02 ± 0.00 | 0.03 ± 0.01 | 0.02 ± 0.01 | 0.02 ± 0.01 |
| 16:1 9t | nd | nd | nd | nd |
| 16:1 (6 + 7 c) | 0.50 ± 0.13 | 1.73 ± 0.23 ** | 1.63 ± 0.21 | 1.31 ± 0.56 |
| 16:1 9 c | 3.76 ± 0.43 | 2.96 ± 0.36 | 2.35 ± 0.30 | 2.20 ± 0.15 § |
| 18:0 | 8.61 ± 0.14 | 4.13 ± 0.40 *** | 6.90 ± 1.31 # | 8.10 ± 2.31 § |
| 18:1 9 t | 0.02 ± 0.01 | 0.04 ± 0.02 | 0.03 ± 0.01 | 0.04 ± 0.02 |
| 18:18c | 0.08 ± 0.04 | 0.13 ± 0.08 | 0.09 ± 0.01 | 0.09 ± 0.02 |
| 18:1 9 c | 18.50 ± 2.07 | 41.55 ± 0.81 *** | 36.18 ± 3.36 | 33.78 ± 3.67 § |
| 18:1 11 c | 4.12 ± 0.48 | 3.57 ± 0.20 | 3.50 ± 0.37 | 3.31 ± 0.29 |
| 18:2 5c.8 cis | 0.05 ± 0.01 | 0.04 ± 0.00 ** | 0.05 ± 0.02 | 0.04 ± 0.02 |
| mt 18:2 | 0.06 ± 0.02 | 0.14 ± 0.04 * | 0.09 ± 0.01 | 0.10 ± 0.01 |
| 18:2 w6 | 15.75 ± 0.32 | 10.41 ± 0.46 *** | 10.39 ± 0.19 | 10.93 ± 1.68 |
| 18:3 w 6 | 0.32 ± 0.01 | 0.28 ± 0.07 | 0.18 ± 0.00 | 0.19 ± 0.08 |
| 18:3 w 3 | 0.45 ± 0.03 | 0.30 ± 0.07 * | 0.27 ± 0.08 | 0.27 ± 0.07 |
| 20:0 | 0.17 ± 0.03 | 0.13 ± 0.09 | 0.18 ± 0.04 | 0.18 ± 0.04 |
| 20:1 9c | 0.42 ± 0.10 | 0.79 ± 0.05 ** | 0.92 ± 0.03 # | 0.75 ± 0.22 |
| 20:3 w 6 DGLA | 3.20 ± 0.30 | 1.57 ± 0.14 ** | 2.65 ± 0.51 # | 2.66 ± 0.37 §§ |
| 20:4 w 6 ARA | 8.49 ± 1.93 | 4.31 ± 0.40 * | 8.66 ± 1.86 # | 8.14 ± 1.21 §§ |
| monotrans 20:4 | 0.05 ± 0.03 | 0.02 ± 0.01 | 0.01 ± 0.00 | 0.06 ± 0.04 |
| 20:3 w3 EPA | 0.66 ± 0.08 | 0.17 ± 0.08 ** | 0.09 ± 0.01 | 0.08 ± 0.03 |
| 22:5 w 3 DPA | 0.50 ± 0.05 | 0.33 ± 0.09 * | 0.23 ± 0.08 | 0.23 ± 0.08 |
| 22:6 w 3 DHA | 9.49 ± 0.78 | 2.67 ± 0.19 *** | 4.60 ± 0.81 # | 3.97 ± 0.76 § |
| SFA s | 33.48 ± 0.91 | 28.98 ± 0.12 ** | 27.99 ± 0.99 ** | 30.84 ± 2.97 |
| MUFAs | 27.37 ± 2.32 | 50.73 ± 1.01 *** | 44.67 ± 4.21 | 41.44 ± 4.86 § |
| PUFAs | 38.92 ± 3.09 | 20.07 ± 0.98 *** | 27.12 ± 3.27 # | 26.50 ± 1.84 §§ |
| w6 | 27.77 ± 2.45 | 16.57 ± 0.71 ** | 21.88 ± 2.54 # | 21.91 ± 1.26 §§ |
| w3 | 11.10 ± 0.66 | 3.46 ± 0.32 *** | 5.19 ± 0.80 # | 4.55 ± 0.81 |
| SFAs/MUFAs | 1.23 ± 0.08 | 0.57 ± 0.01 *** | 0.63 ± 0.08 | 0.76 ± 0.16 |
| w6/w3 | 2.50 ± 0.08 | 4.80 ± 0.32 *** | 4.24 ± 0.29 | 4.90 ± 0.76 |
| Tot trans FAs | 0.15 ± 0.04 | 0.23 ± 0.02 * | 0.16 ± 0.01 ## | 0.22 ± 0.03 £ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valerii, M.C.; Turroni, S.; Ferreri, C.; Zaro, M.; Sansone, A.; Dalpiaz, A.; Botti, G.; Ferraro, L.; Spigarelli, R.; Bellocchio, I.; et al. Effect of a Fiber D-Limonene-Enriched Food Supplement on Intestinal Microbiota and Metabolic Parameters of Mice on a High-Fat Diet. Pharmaceutics 2021, 13, 1753. https://doi.org/10.3390/pharmaceutics13111753
Valerii MC, Turroni S, Ferreri C, Zaro M, Sansone A, Dalpiaz A, Botti G, Ferraro L, Spigarelli R, Bellocchio I, et al. Effect of a Fiber D-Limonene-Enriched Food Supplement on Intestinal Microbiota and Metabolic Parameters of Mice on a High-Fat Diet. Pharmaceutics. 2021; 13(11):1753. https://doi.org/10.3390/pharmaceutics13111753
Chicago/Turabian StyleValerii, Maria Chiara, Silvia Turroni, Carla Ferreri, Michela Zaro, Anna Sansone, Alessandro Dalpiaz, Giada Botti, Luca Ferraro, Renato Spigarelli, Irene Bellocchio, and et al. 2021. "Effect of a Fiber D-Limonene-Enriched Food Supplement on Intestinal Microbiota and Metabolic Parameters of Mice on a High-Fat Diet" Pharmaceutics 13, no. 11: 1753. https://doi.org/10.3390/pharmaceutics13111753