1. Introduction
Rickettsiae are obligate intracellular Gram-negative bacteria that are transmitted to vertebrate hosts by arthropod vectors such as ticks, fleas, lice, and mites. The genus
Rickettsia (family Rickettsiaceae; order Rickettsiales) comprises 31 species that cause diseases in vertebrate hosts, including humans and domestic and wild animals [
1]. Ticks can transmit these bacteria to humans and animals by feeding. Members of the genus
Rickettsia can be classified into four phylogenetic groups: (1) the spotted-fever group (SFG), (2) the typhus group (TG), which includes the agents of epidemic typhus and murine typhus, (3) the ancestral group (AG), consisting of
Ri. bellii and
Ri. canadensis, and (4) the transitional group, the
Ri. akari group (TRG) [
2]. The SFG and AG Rickettsiae are mainly associated with ticks, while TG and TRG Rickettsiae are associated with other arthropods such as lice, fleas, and mites [
3].
Rickettsia spp. and their associated human clinical diseases vary depending on geographical location [
4]. In Europe, rickettsioses are caused mainly by tick-borne SFG Rickettsiae [
4]. In the Mediterranean area, Rickettsiae species have been detected in several tick species, but human rickettsioses are usually considered to be caused by
Ri. conorii, the causative agent of Mediterranean spotted fever (MSF) [
1,
5].
Rickettsia aeschlimanii,
Ri. massiliae, and
Ri. monacensis can also cause a similar clinical presentation. Other species that have been associated with human disease and detected in Europe include
Ri. slovaca and
Ri. raoultii, both causes of tick-borne lymphadenopathy and
Dermacentor-borne necrosis lymphadenopathy (TIBOLA/DEBONEL) [
6,
7].
Information about the circulation and distribution of Rickettsia-infected tick species in Corsica, a French Mediterranean island, is scarce and fragmented.
Rickettsia aeschlimanii and
Candidatus Ri. barbariae were identified in
Hyalomma marginatum and
Rhipicephalus bursa collected from domestic animals in 2016 [
8].
Rickettsia slovaca has been detected in ticks collected from vegetation.
Rickettsia massiliae was reported from an
R. turanicus collected from a dog in Corsica in 2005, and
Ri. africae was detected in one
Amblyomma variegatum collected from bovine in August 2018.
To obtain a better understanding of the current magnitude of tick-borne rickettsioses in Corsica, we aimed to use molecular methods to describe the presence of tick-borne Rickettsiae in tick species collected from domestic and wild animals. From a public health perspective, a better understanding of local tick species and of Rickettsia species carried by them is essential to improve local surveillance systems.
3. Discussion
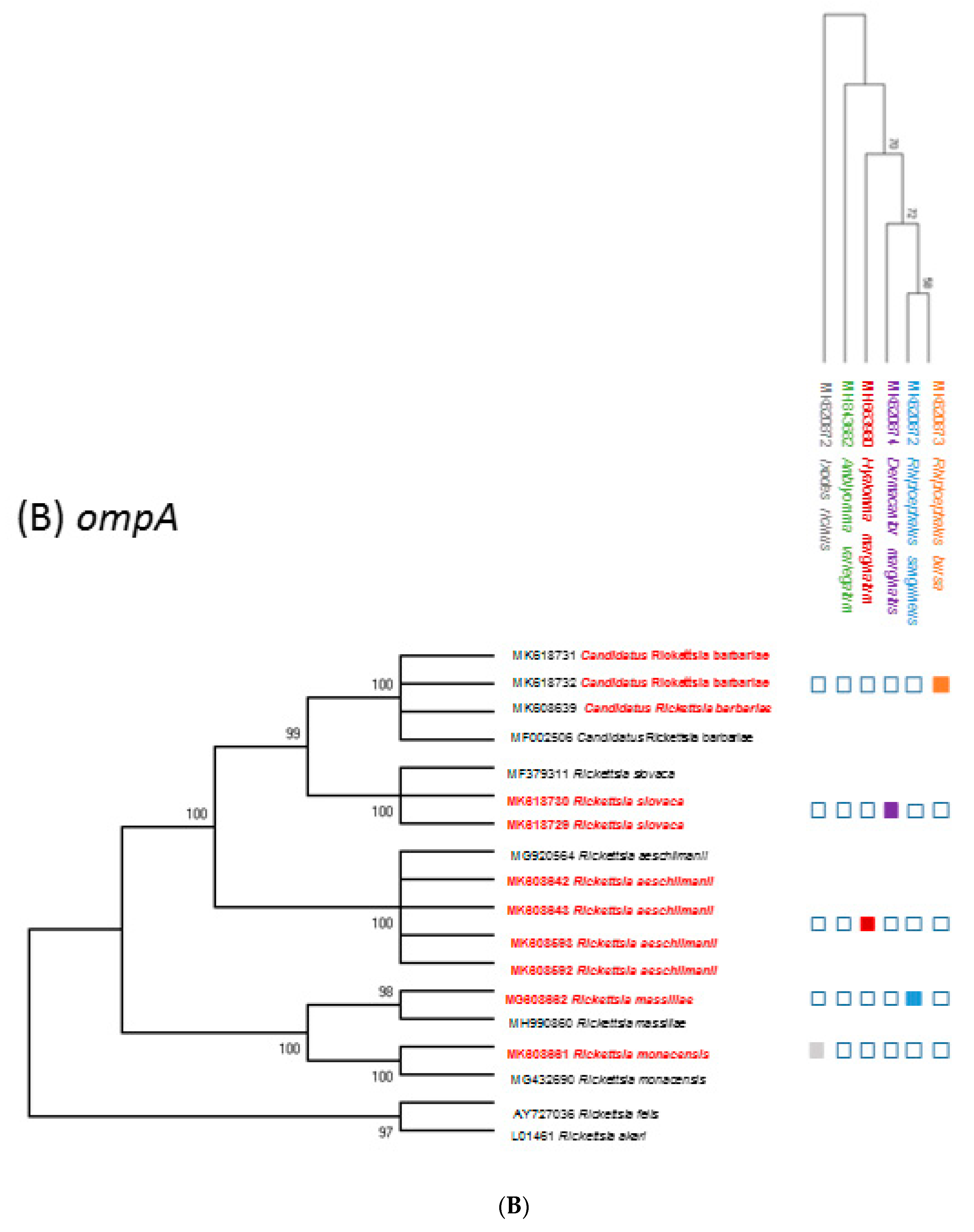
To our knowledge, this is the first reported evidence of the circulation of five bacteria from the genus of Rickettsia in four genera of ticks (Rhipicephalus, Hyalomma, Dermacentor, and Ixodes), collected from wild and domestic animals mainly reared in Northern Corsica.
In the present study, five tick species were collected, namely,
R. bursa,
H. marginatum,
I. ricinus,
D. marginatus, and
R. sanguineus s.l.
Rhipicephalus ticks were the most abundant species collected in this study. The diversity and seasonal pattern of ticks observed in this study, though collected not over 12 months and on selected animals kept under various conditions, is consistent with that reported previously for Corsica [
8,
9] and other Mediterranean countries [
10,
11,
12]. In this study, overall, the low infestation rate observed in sheep with respect to cattle could be due to the short collection period and to a low density of tick populations usually reported in pastures where livestock had grazed with respect to undisturbed fallow lands [
13].
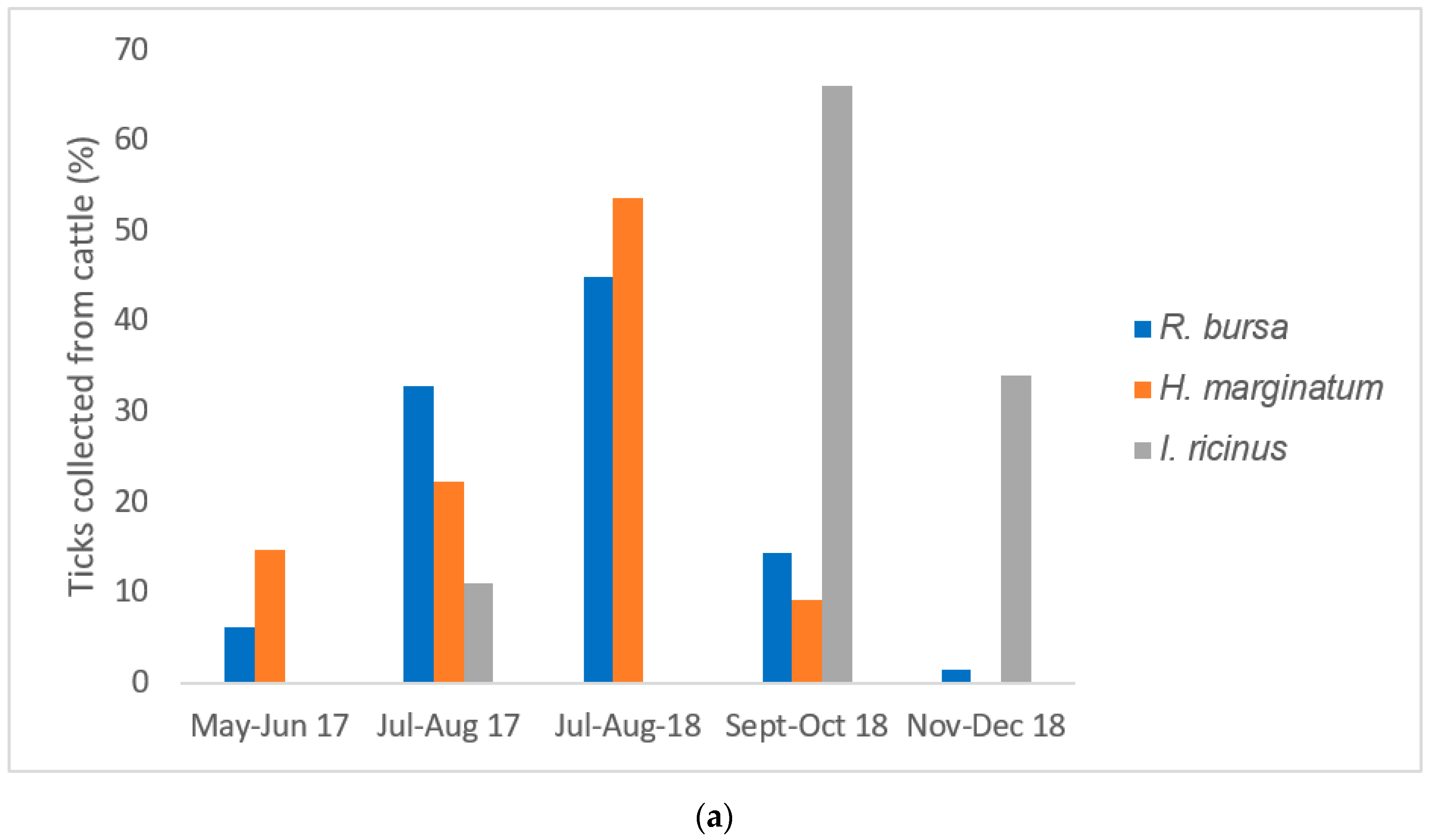
In the present study,
R. bursa was the tick species most frequently collected in cattle and sheep, with a peak of activity observed in the summer. This typical Mediterranean species is well established in the whole of Corsica [
8,
9]. In this study overall, almost 50% of
R. bursa tick pools, mainly collected from cattle and sheep, were positive for
Rickettsia spp. Interestingly,
Candidatus Ri. barbariae DNA was exclusively detected in tick pools of
R. bursa removed from sheep (100% of sequenced tick pools) and mostly in cattle (76%). Our detection of
Candidatus Ri. barbariae in
R. bursa collected from domestic animals reinforces its presence in Corsica ([
8], pp. 606–613) and highlights the possible role played by
R. bursa ticks in the natural maintenance of this bacterium. To understand better the role of
R. bursa in the maintaining of
Candidatus Ri. barbariae, ticks should be collected from nature.
Candidatus Ri. barbariae is a presumed new species of the genus
Rickettsia genetically characterized for the first time in
R. turanicus from Sardinia [
14], a Mediterranean island separated from Corsica by 10 km of sea. The presence of this bacterium has also been reported in several
Rhipicephalus spp. in Portugal, Italy, France, Algeria, Cyprus, Israel, Cameroon, Lebanon [
12,
15,
16,
17,
18,
19], and in
Hyalomma ticks in the West Bank [
20]. The pathogenic role of
Candidatus R. barbariae remains unknown for humans and animals, although it has been identified in an
R. bursa tick removed from a woman in Greece [
21].
Hyalomma marginatum was the second main tick species collected from cattle in the northern region of Corsica. This tick species was almost exclusively collected from cattle (99%). One
H. marginatum was collected from a wild boar.
Rickettsia spp. DNA was detected in more than 80% of
H. marginatum pools collected from cattle sampling in Northern Corsica.
Hyalomma marginatum ticks were exclusively infected with the human pathogen
Ri. aeschlimanii, regardless
of the host species (cattle or wild boars). In Corsica, cattle are reared outside all year round and acaricide treatments are rarely performed, increasing the opportunity of a wide circulation of this pathogen in Corsica. Such a high infection rate is consistent with the previously reported rate of infection of
Hyalomma by
Ri. aeschlimanii (>50% and >70%) in Corsica [
8,
22], in Croatia (64%) [
23], and in Germany (50%) [
24].
Rickettsia aeschlimanii may be spread through migratory birds from Africa, and it was detected in
Hyalomma ticks collected from barn swallows (
Hirundo rustica) in Corsica [
25]. The first human case of infection was described in a patient who developed symptoms after returning from Morocco [
26].
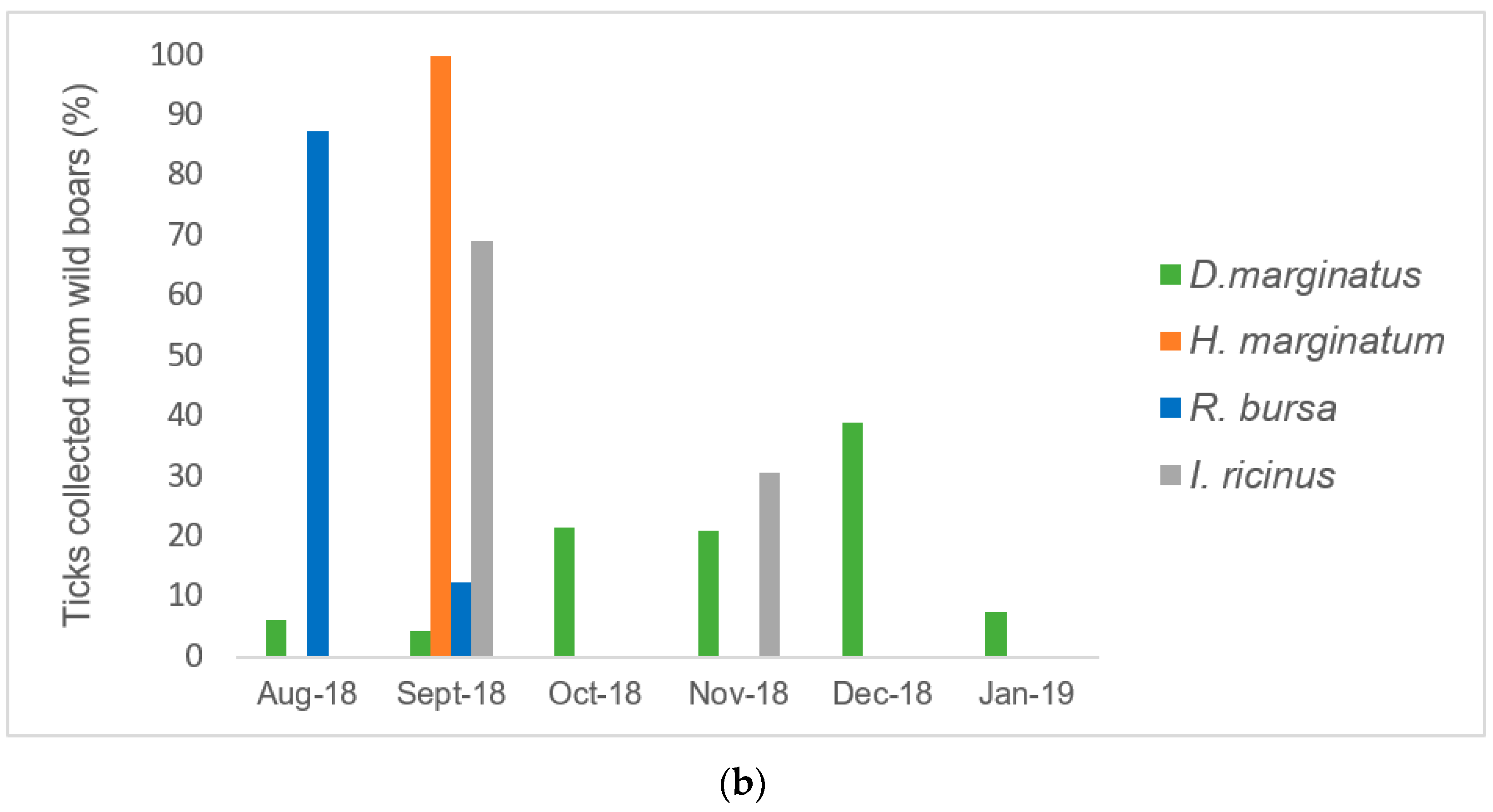
Even if wild boars and cattle share the same environment in Corsica [
9], dominant tick species collected in the present study from these animals differed. In the present study, we confirmed that
D. marginatus was the main tick species collected from wild boars, confirming the host preference previously reported. This tick species was not observed in cattle investigated in this study.
In the present study, most specimens of
D. marginatus were collected during winter because the adults are active within a temperature range of 4–16 °C [
27]. More than half of the pools of
D. marginatus collected were positive for
Rickettsia spp. Sequence analyses revealed for the first time the presence of
Ri. slovaca DNA in
D. marginatus ticks collected from wild boars in Corsica. This bacterium was previously detected in
D. marginatus collected by flagging in Corsica and in other regions of the northern Mediterranean area.
Rickettsia slovaca was identified in
D. marginatum ticks collected on swine and wild boars in Sardinia [
28]. Overall, these results suggested that the role of
D. marginatus is important for the maintenance of
Ri. slovaca. In Corsica, recreational hunting of wild boars could provide an ideal environment for the transmission of
Ri. slovaca between wild boars, humans, and companion animals (
e.g., hunting dogs). At present, the
D. marginatus tick is recognized as the main vector and reservoir for
Ri. slovaca in Mediterranean areas, including southern Europe and North Africa [
1]. This human pathogen has been detected in almost half of
D. marginatus ticks collected from rodents in the northern Apennines (Italy) and by flagging in Kazakhstan [
29,
30].
Rickettsia slovaca is associated with a syndrome characterized by neck lymphadenopathy following tick bites [
6]. Human infection with
Ri. slovaca has been described in several European countries, including France [
31]. We also reported for the first time the detection of
Ri. slovaca DNA in one
D. marginatus collected from a cat.
In this study,
R. sanguineus s.l. ticks were exclusively collected from dogs. We reported the detection of
Ri. massiliae DNA in one pool of
R. sanguineus s.l. collected from a dog.
Rhipicephalus sanguineus s.l. is the likely reservoir of
Ri. massiliae, with transovarian passage rates up to 100%.
Rickettsia massiliae was reported from a specimen of
R. turanicus collected from a dog in Corsica [
32] and has been putatively linked to mild to moderately severe illnesses in dogs in California [
33].
Rickettsia massiliae is recognized as a pathogenic Rickettsia causing spotted fever in humans [
1].
In the present study,
I. ricinus, mainly collected during autumn, was sporadic and represented almost 2% of all ticks sampled. The DNA of
Ri. slovaca was detected in pools of
I. ricinus ticks collected from wild boars. We report for the first time the detection in Corsica of
Ri. monacensis DNA in one pool of
I. ricinus ticks collected from one dog.
Rickettsia monacensis, an emerging human pathogen of the SFG Rickettsiae family, has been previously detected in
I. ricinus collected from dogs in Spain [
34] and in dog blood samples from the Maio Islands [
35].
This study has some limitations. Finding the DNA of human and veterinary pathogens in feeding ticks is only a marker of the community of tick-transmitted pathogens circulating in the target territory [
36]. Ticks were mostly analyzed in pooled samples and not individually. Working with pools can lead to underestimation of the infection rate, which must be interpreted with caution.
The ticks were mostly collected in a municipal slaughterhouse in Northern Corsica. We have no data about the southern part of the island. Moreover, ticks from cattle, wild boars, and dogs were collected from several locations while the sheep were only from one farm.

 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}