
Serosurvey in Two Dengue Hyperendemic Areas of Costa Rica Evidence Active Circulation of WNV and SLEV in Peri-Domestic and Domestic Animals and in Humans
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
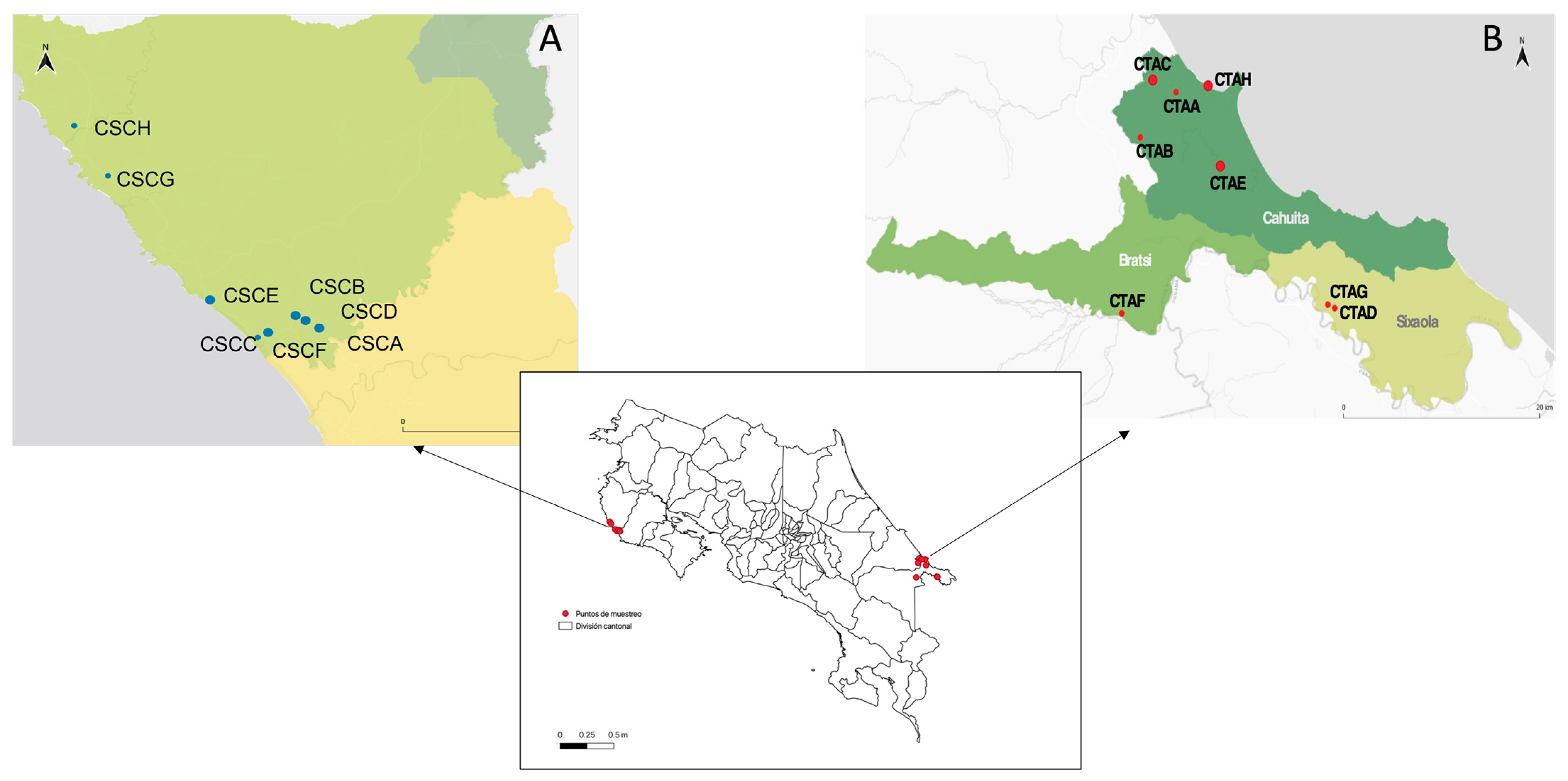
2.1. Study Area
2.2. Sampling and Classification of Wild Birds
2.3. Sampling and Classification of Mosquitoes
2.4. Sampling of Equines, Humans, and Chickens
2.5. Virus Strains
2.6. Serological Screening by Plaque Reduction Neutralization Tests (PRNT)
2.7. Flavivirus RT-PCR in Wild Bird and Mosquito Samples
2.8. Mosquito Blood Meal Identification
2.9. Ethical Statement
3. Results
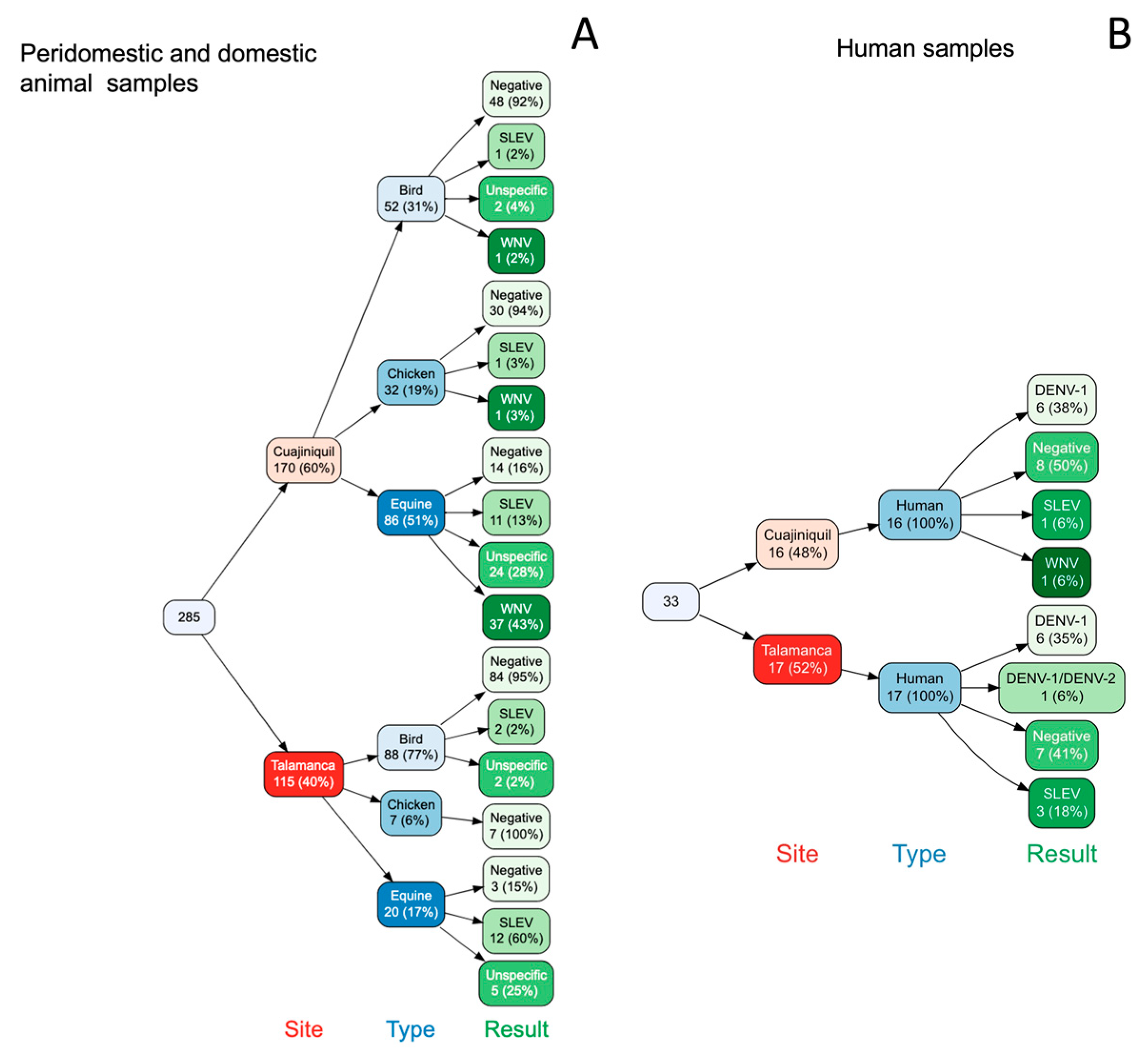
3.1. Several Flaviviruses Cocirculate in Each Sampled Region
3.2. There Was No Evidence of Active Infection in Birds and Mosquitoes
3.3. The Analyses of Mosquito Blood Meals Show a Species Diversity of Feeding Sources
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barrows, N.J.; Campos, R.K.; Liao, K.C.; Prasanth, K.R.; Soto-Acosta, R.; Yeh, S.C.; Schott-Lerner, G.; Pompon, J.; Sessions, O.M.; Bradrick, S.S.; et al. Biochemistry and Molecular Biology of Flaviviruses. Chem. Rev. 2018, 118, 4448–4482. [Google Scholar] [CrossRef]
- Simmonds, P.; Becher, P.; Bukh, J.; Gould, E.A.; Meyers, G.; Monath, T.; Muerhoff, S.; Pletnev, A.; Rico-Hesse, R.; Smith, D.B.; et al. ICTV Virus Taxonomy Profile: Flaviviridae. J. Gen. Virol. 2017, 98, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Holbrook, M.R. Historical Perspectives on Flavivirus Research. Viruses 2017, 9, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierson, T.C.; Diamond, M.S. The continued threat of emerging flaviviruses. Nat. Microbiol. 2020, 5, 796–812. [Google Scholar] [CrossRef]
- Tabachnick, W.J. Climate Change and the Arboviruses: Lessons from the Evolution of the Dengue and Yellow Fever Viruses. Annu. Rev. Virol. 2016, 3, 125–145. [Google Scholar] [CrossRef]
- Weaver, S.C. Urbanization and geographic expansion of zoonotic arboviral diseases: Mechanisms and potential strategies for prevention. Trends Microbiol. 2013, 21, 360–363. [Google Scholar] [CrossRef] [Green Version]
- Gould, E.A.; Higgs, S. Impact of climate change and other factors on emerging arbovirus diseases. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Pandit, P.S.; Doyle, M.M.; Smart, K.M.; Young, C.C.W.; Drape, G.W.; Johnson, C.K. Predicting wildlife reservoirs and global vulnerability to zoonotic Flaviviruses. Nat. Commun. 2018, 9, 5425. [Google Scholar] [CrossRef] [Green Version]
- Daep, C.A.; Muñoz-Jordán, J.L.; Eugenin, E.A. Flaviviruses, an expanding threat in public health: Focus on dengue, West Nile, and Japanese encephalitis virus. J. Neurovirol. 2014, 20, 539–560. [Google Scholar] [CrossRef] [Green Version]
- Rosa, R.; Costa, E.A.; Marques, R.E.; Oliveira, T.S.; Furtini, R.; Bomfim, M.R.; Teixeira, M.M.; Paixão, T.A.; Santos, R.L. Isolation of saint louis encephalitis virus from a horse with neurological disease in Brazil. PLoS Negl. Trop. Dis. 2013, 7, e2537. [Google Scholar] [CrossRef]
- Platt, D.J.; Miner, J.J. Consequences of congenital Zika virus infection. Curr. Opin. Virol. 2017, 27, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kuno, G.; Chang, G.J.; Tsuchiya, K.R.; Karabatsos, N.; Cropp, C.B. Phylogeny of the genus Flavivirus. J. Virol. 1998, 72, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Castillo-Olivares, J.; Wood, J. West Nile virus infection of horses. Vet. Res. 2004, 35, 467–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, E.B.; Sejvar, J.J.; Zaki, S.R.; Lanciotti, R.S.; Bode, A.V.; Campbell, G.L. Virology, pathology, and clinical manifestations of West Nile virus disease. Emerg. Infect Dis. 2005, 11, 1174–1179. [Google Scholar] [CrossRef]
- Diaz, A.; Coffey, L.L.; Burkett-Cadena, N.; Day, J.F. Reemergence of St. Louis Encephalitis Virus in the Americas. Emerg. Infect Dis. 2018, 24, 2150–2157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colpitts, T.M.; Conway, M.J.; Montgomery, R.R.; Fikrig, E. West Nile Virus: Biology, transmission, and human infection. Clin. Microbiol. Rev. 2012, 25, 635–648. [Google Scholar] [CrossRef] [Green Version]
- Ciota, A.T. West Nile virus and its vectors. Curr. Opin. Insect. Sci. 2017, 22, 28–36. [Google Scholar] [CrossRef]
- Kramer, L.; Ciota, A.; Kilpatrick, M. Introduction, Spread, and Establishment of West Nile Virus in the Americas. J. Med. Entomol. 2019, 56, 1448–1455. [Google Scholar] [CrossRef] [PubMed]
- Viana, D.S.; Santamaría, L.; Figuerola, J. Migratory Birds as Global Dispersal Vectors. Trends Ecol. Evol. 2016, 31, 763–775. [Google Scholar] [CrossRef] [Green Version]
- Day, J.F. Predicting St. Louis encephalitis virus epidemics: Lessons from recent, and not so recent, outbreaks. Annu. Rev. Entomol. 2001, 46, 111–138. [Google Scholar] [CrossRef]
- Cerdas-Quesada, C.A.; Retana-Salazar, A.P. Characterization and phylogenetic relationships of strains of Dengue type 1 virus from Costa Rica. Rev. Biol. Trop. 2007, 55, 365–372. [Google Scholar] [CrossRef] [PubMed]
- INCIENSA. Informe de Vigilancia de Arbovirus Basada en Laboratorio. Available online: https://www.inciensa.sa.cr/actualidad/Informes%20de%20vigilancia.aspx#HERMES_TABS_1_3. (accessed on 29 July 2019).
- Vicente-Santos, A.; Moreira-Soto, A.; Soto-Garita, C.; Chaverri, L.G.; Chaves, A.; Drexler, J.F.; Morales, J.A.; Alfaro-Alarcón, A.; Rodríguez-Herrera, B.; Corrales-Aguilar, E. Neotropical bats that co-habit with humans function as dead-end hosts for dengue virus. PLoS Negl. Trop. Dis. 2017, 11, e0005537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto-Garita, C.; Somogyi, T.; Vicente-Santos, A.; Corrales-Aguilar, E. Molecular Characterization of Two Major Dengue Outbreaks in Costa Rica. Am. J. Trop. Med. Hyg. 2016, 95, 201–205. [Google Scholar] [CrossRef] [Green Version]
- Lustig, Y.; Sofer, D.; Bucris, E.D.; Mendelson, E. Surveillance and Diagnosis of West Nile Virus in the Face of Flavivirus Cross-Reactivity. Front. Microbiol. 2018, 9, 2421. [Google Scholar] [CrossRef]
- Hobson-Peters, J.; Arévalo, C.; Cheah, W.Y.; Blitvich, B.J.; Tan, C.S.; Sandis, A.; Araya, L.N.; Hernández, J.L.; Toye, P.; Hall, R.A. Detection of antibodies to West Nile virus in horses, Costa Rica, 2004. Vector Borne. Zoonotic. Dis. 2011, 11, 1081–1084. [Google Scholar] [CrossRef]
- Medlin, S.; Deardorff, E.R.; Hanley, C.S.; Vergneau-Grosset, C.; Siudak-Campfield, A.; Dallwig, R.; da Rosa, A.T.; Tesh, R.B.; Martin, M.P.; Weaver, S.C.; et al. Serosurvey of selected arboviral pathogens in free-ranging, two-toed sloths (Choloepus hoffmanni) and three-toed sloths (Bradypus variegatus) in Costa Rica, 2005–2007. J. Wildl. Dis. 2016, 52, 883–892. [Google Scholar] [CrossRef] [Green Version]
- Jiménez, C.; Romero, M.; Piche, M.; Baldi, M.; Alfaro, A.; Chaves, A.; Morales, J.; León, B.; Hutter, S.; Corrales-Aguilar, E. Arboviral encephalitis in Costa Rican horses: 2009–2016. Int. J. Infect. Dis. 2016, 53, 153. [Google Scholar] [CrossRef] [Green Version]
- Dente, M.G.; Riccardo, F.; Bolici, F.; Colella, N.A.; Jovanovic, V.; Drakulovic, M.; Vasic, M.; Mamlouk, H.; Maazaoui, L.; Bejaoui, M.; et al. Implementation of the One Health approach to fight arbovirus infections in the Mediterranean and Black Sea Region: Assessing integrated surveillance in Serbia, Tunisia and Georgia. Zoonoses Public Health 2019, 66, 276–287. [Google Scholar] [CrossRef] [Green Version]
- Salud, M.d. Análisis de Situación de Salud. Available online: https://www.ministeriodesalud.go.cr/index.php/vigilancia-de-la-salud/analisis-de-situacion-de-salud (accessed on 29 July 2019).
- Animal, S.d.N.d.S. Boletín Epidemiológico Febrero 2016. Available online: https://www.senasa.go.cr/informacion/centro-de-informacion/informacion/estado-sanitario/boletines-epidemiologicos (accessed on 29 July 2019).
- Garrigues, R. The Birds of Costa Rica: A Field Guide; Cornell University Press: Ithaca, NY, USA, 2014. [Google Scholar]
- Association, A.V.M. AVMA Guidelines for Euthanasia of Animals; American Veterinary Medical Association: Schaumburg, IL, USA, 2013; p. 102. [Google Scholar]
- Barrantes Murillo, D.F.; Piche-Ovares, M.; Gamboa-Solano, J.C.; Romero, L.M.; Soto-Garita, C.; Alfaro-Alarcón, A.; Corrales-Aguilar, E. Serological Positivity against Selected. Viruses 2022, 14, 93. [Google Scholar] [CrossRef]
- Vargas, M. El Mosquito un Enemigo Peligroso (Diptera: Culicidae); Universidad de Costa Rica: San José, Costa Rica, 1998. [Google Scholar]
- Darsie, R.F. Key to Mosquitoes of Costa Rica (Diptera: Culicidae); International Center for Disease Control, University of South Carolina: Columbia, SC, USA, 1993. [Google Scholar]
- Guy, B.; Guirakhoo, F.; Barban, V.; Higgs, S.; Monath, T.P.; Lang, J. Preclinical and clinical development of YFV 17D-based chimeric vaccines against dengue, West Nile and Japanese encephalitis viruses. Vaccine 2010, 28, 632–649. [Google Scholar] [CrossRef]
- Pugachev, K.V.; Guirakhoo, F.; Trent, D.W.; Monath, T.P. Chimeric vaccines against Japanese encephalitis, dengue, and West Nile. In New Generation Vaccines; Levine, M.M., Ed.; Informa Healthcare: New York, NY, USA, 2010; pp. 557–569. [Google Scholar]
- Johnson, A.J.; Langevin, S.; Wolff, K.L.; Komar, N. Detection of anti-West Nile virus immunoglobulin M in chicken serum by an enzyme-linked immunosorbent assay. J. Clin. Microbiol. 2003, 41, 2002–2007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, J.Y.; Lee, J.H.; Park, J.Y.; Seo, H.J.; Moon, J.S.; Cho, I.S.; Kim, H.P.; Yang, Y.J.; Ahn, K.M.; Kyung, S.G.; et al. A diagnostic algorithm to serologically differentiate West Nile virus from Japanese encephalitis virus infections and its validation in field surveillance of poultry and horses. Vector Borne. Zoonotic. Dis. 2012, 12, 372–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komar, N.; Langevin, S.; Monath, T.P. Use of a surrogate chimeric virus to detect West Nile virus-neutralizing antibodies in avian and equine sera. Clin. Vaccine. Immunol. 2009, 16, 134–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simões, M.; Camacho, L.A.; Yamamura, A.M.; Miranda, E.H.; Cajaraville, A.C.; da Silva Freire, M. Evaluation of accuracy and reliability of the plaque reduction neutralization test (micro-PRNT) in detection of yellow fever virus antibodies. Biologicals 2012, 40, 399–404. [Google Scholar] [CrossRef] [Green Version]
- Scaramozzino, N.; Crance, J.M.; Jouan, A.; DeBriel, D.A.; Stoll, F.; Garin, D. Comparison of flavivirus universal primer pairs and development of a rapid, highly sensitive heminested reverse transcription-PCR assay for detection of flaviviruses targeted to a conserved region of the NS5 gene sequences. J. Clin. Microbiol. 2001, 39, 1922–1927. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Townzen, J.S.; Brower, A.V.; Judd, D.D. Identification of mosquito bloodmeals using mitochondrial cytochrome oxidase subunit I and cytochrome b gene sequences. Med. Vet. Entomol. 2008, 22, 386–393. [Google Scholar] [CrossRef]
- La Sorte, F.A.; Fink, D.; Hochachka, W.M.; Kelling, S. Convergence of broad-scale migration strategies in terrestrial birds. Proc. Biol. Sci. 2016, 283, 20152588. [Google Scholar] [CrossRef] [Green Version]
- Dolz, G.; Chaves, A.; Gutiérrez-Espeleta, G.A.; Ortiz-Malavasi, E.; Bernal-Valle, S.; Herrero, M.V. Detection of antibodies against flavivirus over time in wild non-human primates from the lowlands of Costa Rica. PLoS ONE 2019, 14, e0219271. [Google Scholar] [CrossRef]
- Calderón-Arguedas, O.; Troyo, A.; Solano, M.E.; Avendaño, A. Culicidofauna Asociada con Contenedores Artificiales en la Comunidad “La Carpio”, Costa Rica. Rev. Costarric. De Salud Pública 2009, 18, 30–36. [Google Scholar]
- Heinen, L.B.; Zuchi, N.; Serra, O.P.; Cardoso, B.F.; Gondim, B.H.; Dos Santos, M.A.; Souto, F.J.; Paula, D.A.; Dutra, V.; Dezengrini-Slhessarenko, R. Saint louis encephalitis virus in mato grosso, central-western brazil. Rev. Inst. Med. Trop. Sao Paulo 2015, 57, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Mondini, A.; Bronzoni, R.V.; Cardeal, I.L.; dos Santos, T.M.; Lázaro, E.; Nunes, S.H.; Silva, G.C.; Madrid, M.C.; Rahal, P.; Figueiredo, L.T.; et al. Simultaneous infection by DENV-3 and SLEV in Brazil. J. Clin. Virol. 2007, 40, 84–86. [Google Scholar] [CrossRef] [PubMed]
- Lopez, H.; Neira, J.; Morales, M.A.; Fabbri, C.; D’Agostino, M.L.; Zitto, T. Encefalitis por virus de San Luis en la ciudad de Buenos Aires durante el brote de Dengue 2009. Medicina 2010, 70, 247–250. [Google Scholar]
- Rappole, J.H.; Derrickson, S.R.; Hubálek, Z. Migratory birds and spread of West Nile virus in the Western Hemisphere. Emerg. Infect Dis. 2000, 6, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Reed, K.D.; Meece, J.K.; Henkel, J.S.; Shukla, S.K. Birds, migration and emerging zoonoses: West nile virus, lyme disease, influenza A and enteropathogens. Clin. Med. Res. 2003, 1, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Mancuso, E.; Toma, L.; Pascucci, I.; d’Alessio, S.G.; Marini, V.; Quaglia, M.; Riello, S.; Ferri, A.; Spina, F.; Serra, L.; et al. Direct and Indirect Role of Migratory Birds in Spreading CCHFV and WNV: A Multidisciplinary Study on Three Stop-Over Islands in Italy. Pathogens 2022, 11, 1056. [Google Scholar] [CrossRef]
- Ain-Najwa, M.Y.; Yasmin, A.R.; Omar, A.R.; Arshad, S.S.; Abu, J.; Mohammed, H.O.; Kumar, K.; Loong, S.K.; Rovie-Ryan, J.J.; Mohd-Kharip-Shah, A.K. Evidence of West Nile virus infection in migratory and resident wild birds in west coast of peninsular Malaysia. One Health 2020, 10, 100134. [Google Scholar] [CrossRef]
- Hannon, E.R.; Jackson, K.C.; Biggerstaff, B.J.; Raman, V.; Komar, N. Bloodmeal Host Selection of Culex quinquefasciatus (Diptera: Culicidae) in Las Vegas, Nevada, United States. J. Med. Entomol. 2019, 56, 603–608. [Google Scholar] [CrossRef]
- Richards, S.L.; Anderson, S.L.; Lord, C.C.; Smartt, C.T.; Tabachnick, W.J. Relationships between infection, dissemination, and transmission of West Nile virus RNA in Culex pipiens quinquefasciatus (Diptera: Culicidae). J. Med. Entomol. 2012, 49, 132–142. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, D.; Piche-Ovares, M.; Romero-Vega, L.M.; Wagman, J.; Troyo, A. The Impact of Deforestation, Urbanization, and Changing Land Use Patterns on the Ecology of Mosquito and Tick-Borne Diseases in Central America. Insects 2022, 13, 20. [Google Scholar] [CrossRef]
- Katzelnick, L.C.; Narvaez, C.; Arguello, S.; Lopez Mercado, B.; Collado, D.; Ampie, O.; Elizondo, D.; Miranda, T.; Bustos Carillo, F.; Mercado, J.C.; et al. Zika virus infection enhances future risk of severe dengue disease. Science 2020, 369, 1123–1128. [Google Scholar] [CrossRef] [PubMed]
- Hadfield, J.; Brito, A.F.; Swetnam, D.M.; Vogels, C.B.F.; Tokarz, R.E.; Andersen, K.G.; Smith, R.C.; Bedford, T.; Grubaugh, N.D. Twenty years of West Nile virus spread and evolution in the Americas visualized by Nextstrain. PLoS Pathog. 2019, 15, e1008042. [Google Scholar] [CrossRef] [PubMed]
- Diaz, L.A.; Goñi, S.E.; Iserte, J.A.; Quaglia, A.I.; Singh, A.; Logue, C.H.; Powers, A.M.; Contigiani, M.S. Exploring Genomic, Geographic and Virulence Interactions among Epidemic and Non-Epidemic St. Louis Encephalitis Virus (Flavivirus) Strains. PLoS ONE 2015, 10, e0136316. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Species | Sample Type | Rainy Season (n) | Dry Season (n) |
|---|---|---|---|
| Equine | Serum | 86 | 57 |
| Wild birds | Tissue/serum | 16 | 36 |
| Humans | Serum | 16 | 12 |
| Mosquitoes | Pools | 119 (377 individuals) | 44 (177 individuals) |
| Blood-engorged | 9 | 3 | |
| Chickens | Serum | 29 | 3 |
| Species | Sample Type | Rainy Season (n) | Dry Season (n) |
|---|---|---|---|
| Equine | Serum | 20 | 16 |
| Wild birds | Tissue/serum | 60 | 28 |
| Humans | Serum | 17 | 15 |
| Mosquitoes | Pools | 133 (573 individuals) | 66 (246 individuals) |
| Blood-engorged | 17 | 5 | |
| Chickens | Serum | 7 | 0 |
| Pool Identification | Household | Species | Homology | GenBank |
|---|---|---|---|---|
| TCC28 | CTAC | Culex (Melanoconion) sp. | Mosquito flavivirus | MN856866.1 |
| TDC19 | CTAD | Aedes aegypti | Aedes flavivirus | MK241496.1 |
| Pool Identification | Household | Species | Homology | GenBank |
|---|---|---|---|---|
| OEC7 | CSCE | Culex restrictor | Equus ferus caballus | MH605334.1 |
| OFC4 | CSCF | Anopheles albimanus | Homo sapiens sapiens | MF588853.1 |
| OFC22 | CSCF | Culex quinquefasciatus | Equus ferus caballus | MG761997 |
| OMC4 | CSCM | Deinocerites pseudes | Canis lupus familiaris | KU290927 |
| TBC1 | CTAB | Culex quinquefasciatus | Canis lupus familiaris | MH105046.1 |
| TCC1 | CTAC | Culex coronator | Canis lupus familiaris | KM061528.1 |
| TCC2 | CTAC | Culex coronator | Homo sapiens sapiens | K792836.1 |
| TCC23 | CTAC | Culex (Melanoconion) sp. | Equus ferus caballus | MG761996.1 |
| TCC24 | CTAC | Culex coronator | Equus ferus caballus | MG761996.1 |
| TDC8 | CTAD | Psorophora ferox | Homo sapiens sapiens | K792836.1 |
| TFC1 | CTAF | Culex quinquefasciatus | Ovis orientalis aries | MG489885.1 |
| TFC2 | CTAF | Culex quinquefasciatus | Homo sapiens sapiens | MK792836.1 |
| TFC3 | CTAF | Culex quinquefasciatus | Homo sapiens sapiens | MK792836.1 |
| TFC4 | CTAF | Culex quinquefasciatus | Ovis orientalis aries | MG489885.1 |
| TFC5 | CTAF | Culex quinquefasciatus | Ovis orientalis aries | MG489885.1 |
| TGC3 | CTAG | Culex pseudostigmatosoma | Canis lupus familiaris | KU290927.1 |
| TGC4 | CTAG | Culex pseudostigmatosoma | Homo sapiens sapiens | MK103007.1 |
| THC3 | CTAH | Culex quinquefasciatus | Columbina paserina | DQ433535.1 |
| THC6 | CTAH | Culex (Melanoconion) sp. | Ovis orientalis aries | MG489885.1 |
| TMC9 | CTAM | Culex quinquefasciatus | Canis lupus familiaris | KM061528.1 |
| TMC7 | CTAM | Culex quinquefasciatus | Canis lupus familiaris | KM061528.1 |
| TMC8 | CTAM | Culex quinquefasciatus | Canis lupus familiaris/ Homo sapiens sapiens | KU696393.2/MK103007.1 |
| TNC9 | CTAN | Mansonia titillans | Equus ferus caballus | MG761996.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piche-Ovares, M.; Romero-Vega, M.; Vargas-González, D.; Murillo, D.F.B.; Soto-Garita, C.; Francisco-Llamas, J.; Alfaro-Alarcón, A.; Jiménez, C.; Corrales-Aguilar, E. Serosurvey in Two Dengue Hyperendemic Areas of Costa Rica Evidence Active Circulation of WNV and SLEV in Peri-Domestic and Domestic Animals and in Humans. Pathogens 2023, 12, 7. https://doi.org/10.3390/pathogens12010007
Piche-Ovares M, Romero-Vega M, Vargas-González D, Murillo DFB, Soto-Garita C, Francisco-Llamas J, Alfaro-Alarcón A, Jiménez C, Corrales-Aguilar E. Serosurvey in Two Dengue Hyperendemic Areas of Costa Rica Evidence Active Circulation of WNV and SLEV in Peri-Domestic and Domestic Animals and in Humans. Pathogens. 2023; 12(1):7. https://doi.org/10.3390/pathogens12010007
Chicago/Turabian StylePiche-Ovares, Marta, Mario Romero-Vega, Diana Vargas-González, Daniel Felipe Barrantes Murillo, Claudio Soto-Garita, Jennifer Francisco-Llamas, Alejandro Alfaro-Alarcón, Carlos Jiménez, and Eugenia Corrales-Aguilar. 2023. "Serosurvey in Two Dengue Hyperendemic Areas of Costa Rica Evidence Active Circulation of WNV and SLEV in Peri-Domestic and Domestic Animals and in Humans" Pathogens 12, no. 1: 7. https://doi.org/10.3390/pathogens12010007