
No Evidence for the Involvement of Leiomodin-1 Antibodies in the Pathogenesis of Onchocerciasis-Associated Epilepsy

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants and Sample Collection
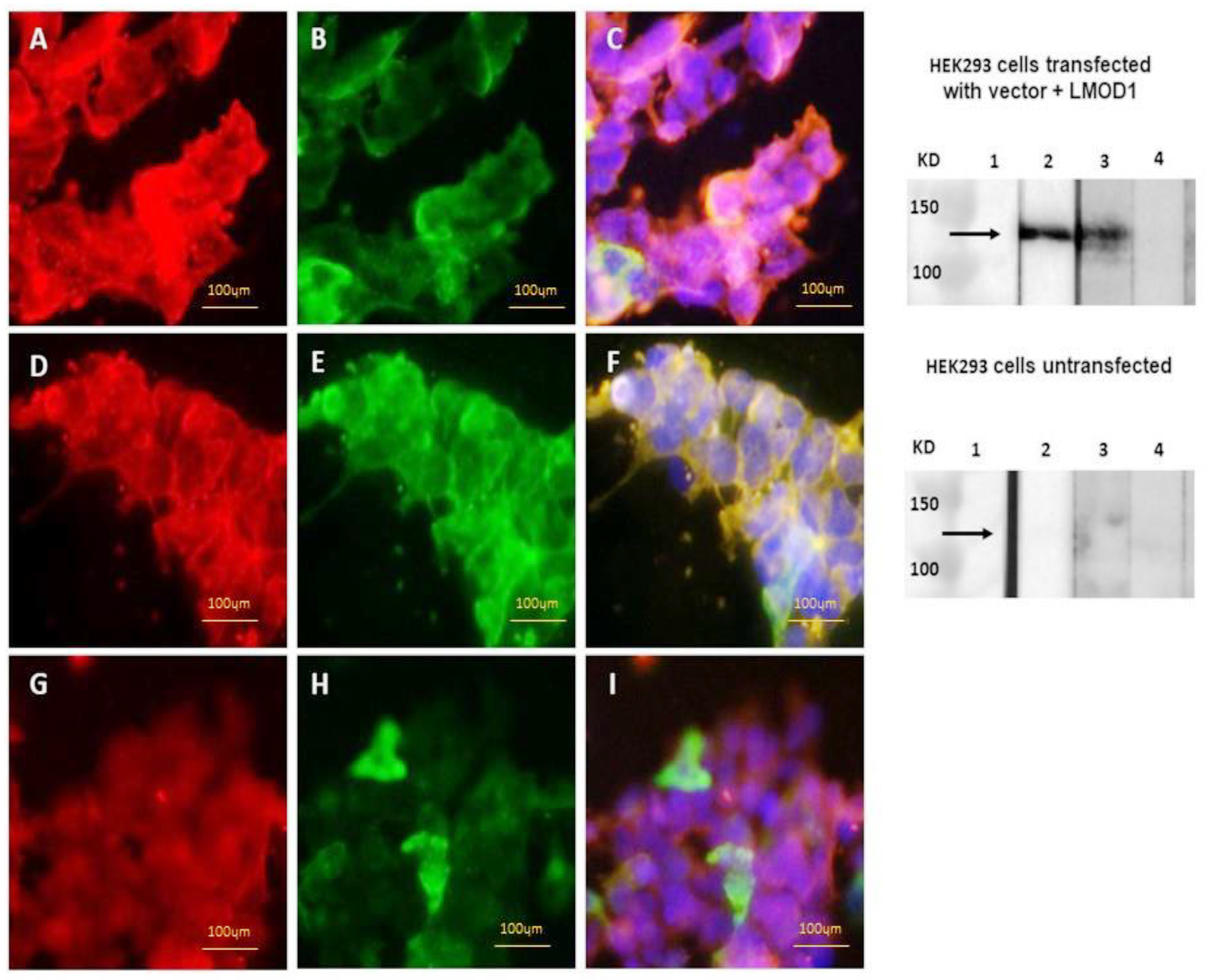
2.2. Detection of Anti-Leiomodin-1 Antibodies in a Cell-Based Assay
2.3. Detection of Anti-Leiomodin-1 Antibodies by Western Blot
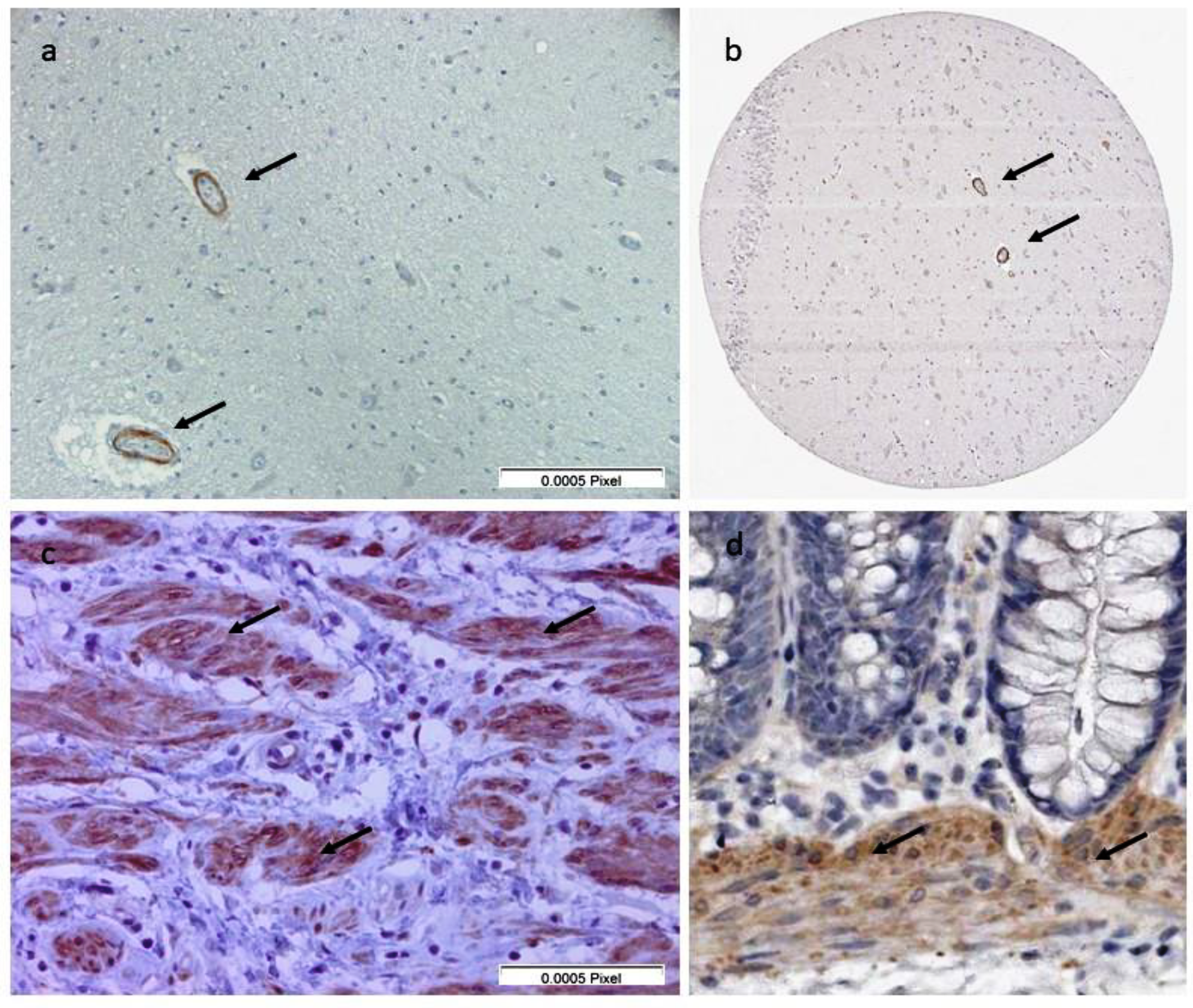
2.4. Immunohistochemistry to Screen for Auto-Antibodies in Serum Samples
2.5. Immunohistochemistry to Detect the Presence of Leiomodin-1 in Post-Mortem Brains of People with OAE Who Died
2.6. Statistics
3. Results
3.1. Leiomodin-1 Antibodies in Serum and CSF
3.2. Immunohistochemistry to Screen for Auto-Antibodies and of Post-Mortem Brains of People with OAE Who Died
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Colebunders, R.; Nelson Siewe, F.J.; Hotterbeekx, A. Onchocerciasis-Associated Epilepsy, an Additional Reason for Strengthening Onchocerciasis Elimination Programs. Trends Parasitol. 2018, 34, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Hotterbeekx, A.; Lammens, M.; Idro, R.; Akun, P.R.; Lukande, R.; Akena, G.; Nath, A.; Taylor, J.; Olwa, F.; Kumar-Singh, S.; et al. Neuroinflammation and Not Tauopathy Is a Predominant Pathological Signature of Nodding Syndrome. J. Neuropathol. Exp. Neurol. 2019. [Google Scholar] [CrossRef]
- Hotterbeekx, A.; Raimon, S.; Abd-Elfarag, G.; Carter, J.Y.; Sebit, W.; Suliman, A.; Fodjo, J.N.S.; De Witte, P.; Logora, M.Y.; Colebunders, R.; et al. Onchocerca volvulus is not detected in the cerebrospinal fluid of persons with onchocerciasis-associated epilepsy. Int. J. Infect. Dis. 2019. [Google Scholar] [CrossRef] [Green Version]
- Pollanen, M.S.; Onzivua, S.; Robertson, J.; McKeever, P.M.; Olawa, F.; Kitara, D.L.; Fong, A. Nodding syndrome in Uganda is a tauopathy. Acta Neuropathol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.P.; Tyagi, R.; Lee, P.R.; Lee, M.H.; Johnson, K.R.; Kowalak, J.; Elkahloun, A.; Medynets, M.; Hategan, A.; Kubofcik, J.; et al. Nodding syndrome may be an autoimmune reaction to the parasitic worm Onchocerca volvulus. Sci. Transl. Med. 2017, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nauen, D.W.; Haffner, M.C.; Kim, J.; Zheng, Q.; Yin, H.; DeMarzo, A.M.; Mahairaki, V.; Colantuoni, C.; Pickering, J.G.; Johnson, T.P. Putative Autoantigen Leiomodin-1 Is Expressed in the Human Brain and in the Membrane Fraction of Newly Formed Neurons. Pathogens 2020, 9, 1036. [Google Scholar] [CrossRef]
- Nanda, V.; Miano, J.M. Leiomodin 1, a new serum response factor-dependent target gene expressed preferentially in differentiated smooth muscle cells. J. Biol. Chem. 2012, 287, 2459–2467. [Google Scholar] [CrossRef] [Green Version]
- Halim, D.; Wilson, M.P.; Oliver, D.; Brosens, E.; Verheij, J.B.G.M.; Han, Y.; Nanda, V.; Lyu, Q.; Doukas, M.; Stoop, H.; et al. Loss of LMOD1 impairs smooth muscle cytocontractility and causes megacystis microcolon intestinal hypoperistalsis syndrome in humans and mice. Proc. Natl. Acad. Sci. USA 2017, 114, E2739–E2747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Human Protein Atlas. Available online: http://www.proteinatlas.org (accessed on 3 July 2021).
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Colebunders, R.; Titulaer, M.J. Nodding syndrome: Preventable and treatable. Sci. Transl. Med. 2017, 9. [Google Scholar] [CrossRef]
- Mandro, M.; Suykerbuyk, P.; Tepage, F.; Rossy, D.; Ngave, F.; Hasan, M.N.; Hotterbeekx, A.; Mambandu, G.; Kashama, J.M.; Laudisoit, A.; et al. Onchocerca volvulus as a risk factor for developing epilepsy in onchocerciasis endemic regions in the Democratic Republic of Congo: A case control study. Infect. Dis. Poverty 2018, 7, 79. [Google Scholar] [CrossRef]
- Colebunders, R.; Mandro, M.; Mokili, J.L.; Mucinya, G.; Mambandu, G.; Pfarr, K.; Reiter-Owona, I.; Hoerauf, A.; Tepage, F.; Levick, B.; et al. Risk factors for epilepsy in Bas-Uele Province, Democratic Republic of the Congo: A case-control study. Int. J. Infect. Dis. 2016, 49, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandro, M.; Siewe Fodjo, J.N.; Dusabimana, A.; Mukendi, D.; Haesendonckx, S.; Lokonda, R.; Nakato, S.; Nyisi, F.; Abhafule, G.; Wonya’rossi, D.; et al. Single versus Multiple Dose Ivermectin Regimen in Onchocerciasis-Infected Persons with Epilepsy Treated with Phenobarbital: A Randomized Clinical Trial in the Democratic Republic of Congo. Pathogens 2020, 9, 205. [Google Scholar] [CrossRef] [Green Version]
- Hotterbeekx, A.; Perneel, J.; Mandro, M.; Abhafule, G.; Siewe Fodjo, J.N.; Dusabimana, A.; Abrams, S.; Kumar-Singh, S.; Colebunders, R. Comparison of Diagnostic Tests for Onchocerca volvulus in the Democratic Republic of Congo. Pathogens 2020, 9, 435. [Google Scholar] [CrossRef] [PubMed]
- Mukendi, D.; Lilo Kalo, J.R.; Mpanya, A.; Minikulu, L.; Kayembe, T.; Lutumba, P.; Barbe, B.; Gillet, P.; Jacobs, J.; Van Loen, H.; et al. Clinical Spectrum, Etiology, and Outcome of Neurological Disorders in the Rural Hospital of Mosango, the Democratic Republic of Congo. Am. J. Trop. Med. Hyg. 2017, 97, 1454–1460. [Google Scholar] [CrossRef] [Green Version]
- Colebunders, R.; Siewe Fodjo, J.N.; Hopkins, A.; Hotterbeekx, A.; Lakwo, T.L.; Kalinga, A.; Logora, M.Y.; Basanez, M.G. From river blindness to river epilepsy: Implications for onchocerciasis elimination programmes. PLoS Negl. Trop. Dis. 2019, 13, e0007407. [Google Scholar] [CrossRef] [Green Version]
- de Bruijn, M.; Bastiaansen, A.E.M.; Mojzisova, H.; van Sonderen, A.; Thijs, R.D.; Majoie, M.J.M.; Rouhl, R.P.W.; van Coevorden-Hameete, M.H.; de Vries, J.M.; Munoz Lopetegi, A.; et al. Antibodies Contributing to Focal Epilepsy Signs and Symptoms Score. Ann. Neurol. 2021. [Google Scholar] [CrossRef]
- van Coevorden-Hameete, M.H.; Titulaer, M.J.; Schreurs, M.W.; de Graaff, E.; Sillevis Smitt, P.A.; Hoogenraad, C.C. Detection and Characterization of Autoantibodies to Neuronal Cell-Surface Antigens in the Central Nervous System. Front. Mol. Neurosci. 2016, 9, 37. [Google Scholar] [CrossRef] [Green Version]
- Colebunders, R.; Abd-Elfarag, G.; Carter, J.Y.; Olore, P.C.; Puok, K.; Menon, S.; Fodjo Siewe, J.N.; Bhattacharyya, S.; Ojok, M.; Lako, R.; et al. Clinical characteristics of onchocerciasis-associated epilepsy in villages in Maridi County, Republic of South Sudan. Seizure 2018, 62, 108–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siewe Fodjo, J.N.; Mandro, M.; Mukendi, D.; Tepage, F.; Menon, S.; Nakato, S.; Nyisi, F.; Abhafule, G.; Wonya’rossi, D.; Anyolito, A.; et al. Onchocerciasis-associated epilepsy in the Democratic Republic of Congo: Clinical description and relationship with microfilarial density. PLoS Negl. Trop. Dis. 2019, 13, e0007300. [Google Scholar] [CrossRef] [Green Version]
- Nockler, K.; Reckinger, S.; Broglia, A.; Mayer-Scholl, A.; Bahn, P. Evaluation of a Western Blot and ELISA for the detection of anti-Trichinella-IgG in pig sera. Vet. Parasitol. 2009, 163, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Ances, B.M.; Vitaliani, R.; Taylor, R.A.; Liebeskind, D.S.; Voloschin, A.; Houghton, D.J.; Galetta, S.L.; Dichter, M.; Alavi, A.; Rosenfeld, M.R.; et al. Treatment-responsive limbic encephalitis identified by neuropil antibodies: MRI and PET correlates. Brain 2005, 128, 1764–1777. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Region and Type of Study | Study Participants | Number |
|---|---|---|
| Ituri Province, Logo health zone, clinical trial [14], case–control study [12,13], REMO study [15] | OAE, skin snip and/or OV16 RDT positive | 54 |
| No epilepsy, O. volvulus infection | 42 | |
| No epilepsy and no O. volvulus infection 1 | 19 | |
| Epilepsy centers and academic hospitals, the Netherlands [18] | Focal epilepsy without antibodies associated with encephalitis | 12 |
| Healthy volunteers | 4 |
| Region | Study Participants | Number |
|---|---|---|
| Maridi, South Sudan [3] | OAE | 12 |
| Mosango hospital, Kwilu Province, DRC [16] | Acute-onset neurological conditions | 16 |
| Case 1 | Case 2 | Case 3 | Case 4 | Case 5 | Control 1 | Control 2 |
|---|---|---|---|---|---|---|
| Hippocampus (dentate gyrus) | Hippocampus (dentate gyrus) | Hippocampus (dentate gyrus) | Hippocampus (dentate gyrus) | Hippocampus | Hippocampus | Hippocampus |
| Hippocampus + amygdala | Cerebellum cortex | Hippocampus anterior part 1 | Hippocampus | Cerebellum cortex | Cerebellum cortex | Cerebellum cortex |
| Cerebellum vermis | Hippocampus anterior part 2 | Cerebellum cortex | Cerebellum vermis | Frontal cortex | Frontal cortex | |
| Frontal cortex | Hippocampus anterior part 3 | Cerebellum vermis | Thalamus + corpus mamillaria | Sulcus calcarinus | ||
| Caudate nucleus | Cerebellum cortex | Hypothalamus anterior + corpus mamillaria | ||||
| Primary motor cortex | Cerebellum vermis | Hypothalamus posterior + corpus mamillaria | ||||
| Parietal cortex | Superior gyrus of the temporal lobe |
| Parameter | Persons with OAE (N = 54) * | Controls without Epilepsy (N = 61) | p-Value |
|---|---|---|---|
| Age (median, IQR) | 19 (14–23) | 31 (15.5–46.5) | <0.001 |
| Male (N, %) | 30 (55%) | 49 (80%) | 0.004 |
| Nodding seizures (N, %) | 16 | NA | NA |
| Age of seizure onset (median, IQR) | 10 (7–13) | NA | NA |
| Ever treated with ivermectin (N, %) | 4 (7%) | 11 (19%) | 0.091 |
| Leiomodin-1 antibodies by CBA (N, %) | 6/52 (11%) | 14 (23%) | 0.113 |
| Leiomodin-1 antibodies by WB (N, %) | 23/54 (43%) | 30 (49%) | 0.479 |
| Leiomodin-1 antibodies positive, CBA and WB (N, %) | 5/52 (10%) | 12 (20%) | 0.136 |
| Leiomodin-1 antibodies positive, CBA or WB (N, %) | 24/54 (44%) | 32 (52%) | 0.391 |
| Ov + (N = 96) | Ov − (N = 19) | p-Value | |
| Leiomodin-1 CBA+ (N, %) | 17 (18%) | 3 (16%) | 0.840 |
| Ov + (N = 96) | Ov − (N = 19) | ||
| Leiomodin-1 WB + (N, %) | 44 (46%) | 10 (53%) | 0.587 |
| Leiomodin-1 antibodies positive, CBA and WB (N, %) | 17 (18%) | 2 (11%) | 0.397 |
| Cell-Based Assay | Western Blot | |||||||
|---|---|---|---|---|---|---|---|---|
| Parameter | Estimate | 95% CI | p-Value | Estimate | 95%CI | p-Value | ||
| Gender (Male vs. Female) | 3.599 | 1.271 | 24.836 | 0.007 | 1.352 | 0.821 | 2.270 | 0.278 |
| Age (years) | 0.977 | 0.940 | 1.014 | 0.230 | 1.033 | 1.003 | 1.065 | 0.031 |
| Persons with (OAE vs. control without epilepsy) | 0.528 | 0.237 | 1.106 | 0.100 | 1.142 | 0.680 | 1.962 | 0.757 |
| O. volvulus-infected vs. no infection | 1.777 | 0.794 | 4.672 | 0.208 | 1.029 | 0.525 | 2.020 | 0.928 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hotterbeekx, A.; Vieri, M.K.; Ramberger, M.; Jozefzoon-Aghai, A.; Mandro, M.; Tepage, F.; Dusabimana, A.; Kumar-Singh, S.; Titulaer, M.J.; Colebunders, R. No Evidence for the Involvement of Leiomodin-1 Antibodies in the Pathogenesis of Onchocerciasis-Associated Epilepsy. Pathogens 2021, 10, 845. https://doi.org/10.3390/pathogens10070845
Hotterbeekx A, Vieri MK, Ramberger M, Jozefzoon-Aghai A, Mandro M, Tepage F, Dusabimana A, Kumar-Singh S, Titulaer MJ, Colebunders R. No Evidence for the Involvement of Leiomodin-1 Antibodies in the Pathogenesis of Onchocerciasis-Associated Epilepsy. Pathogens. 2021; 10(7):845. https://doi.org/10.3390/pathogens10070845
Chicago/Turabian StyleHotterbeekx, An, Melissa Krizia Vieri, Melanie Ramberger, Ashraf Jozefzoon-Aghai, Michel Mandro, Floribert Tepage, Alfred Dusabimana, Samir Kumar-Singh, Maarten J. Titulaer, and Robert Colebunders. 2021. "No Evidence for the Involvement of Leiomodin-1 Antibodies in the Pathogenesis of Onchocerciasis-Associated Epilepsy" Pathogens 10, no. 7: 845. https://doi.org/10.3390/pathogens10070845