
Development of an Indirect ELISA to Detect Equine Antibodies to Theileria haneyi


 ,
,
Abstract
:1. Introduction
2. Results
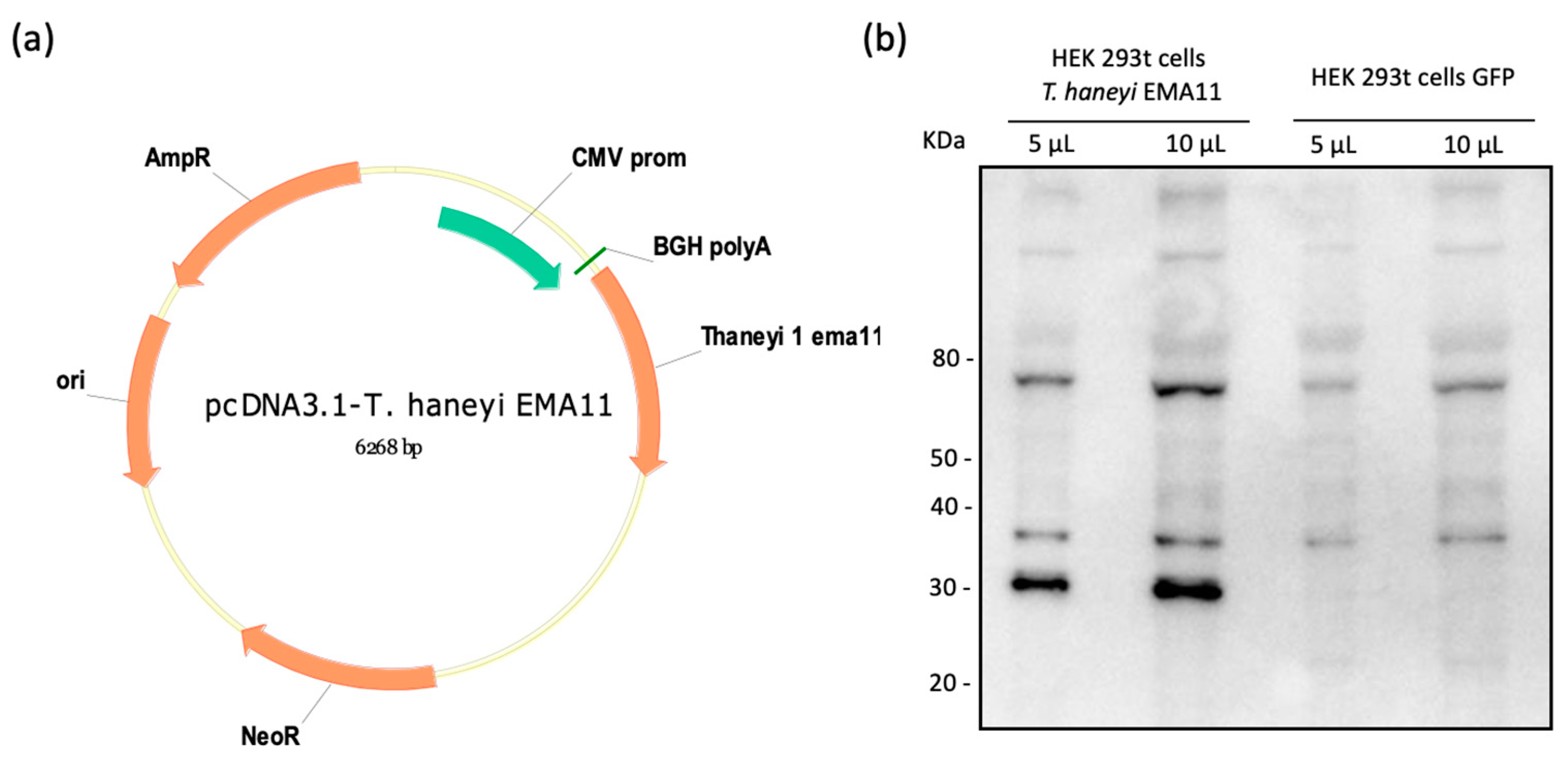
2.1. Cloning and Expression of ThEMA11
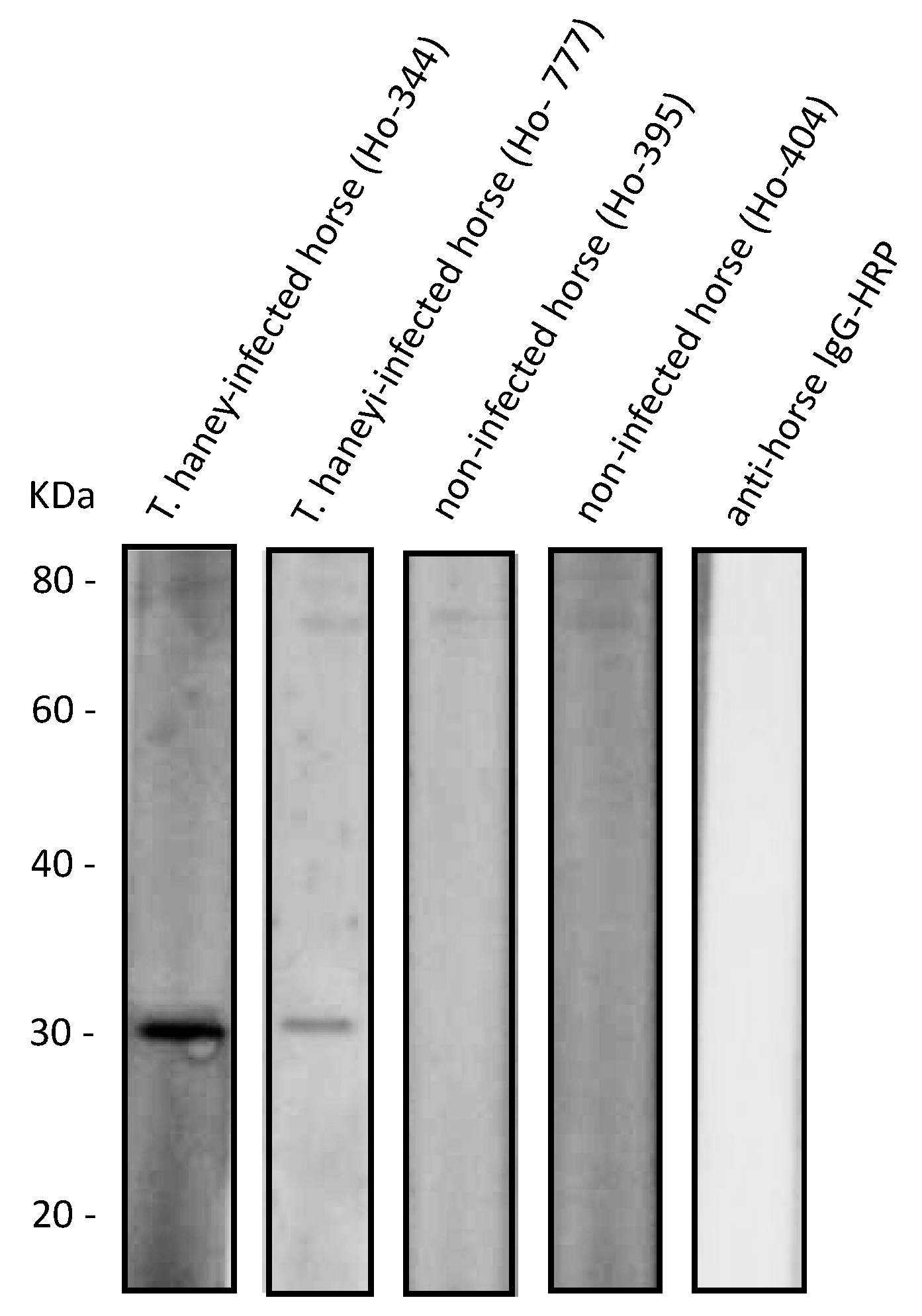
2.2. Recombinant ThEMA11 Is Recognized by Serum from T haneyi-Infected Horses, but Not Serum from T. equi-Infected or Uninfected Horses
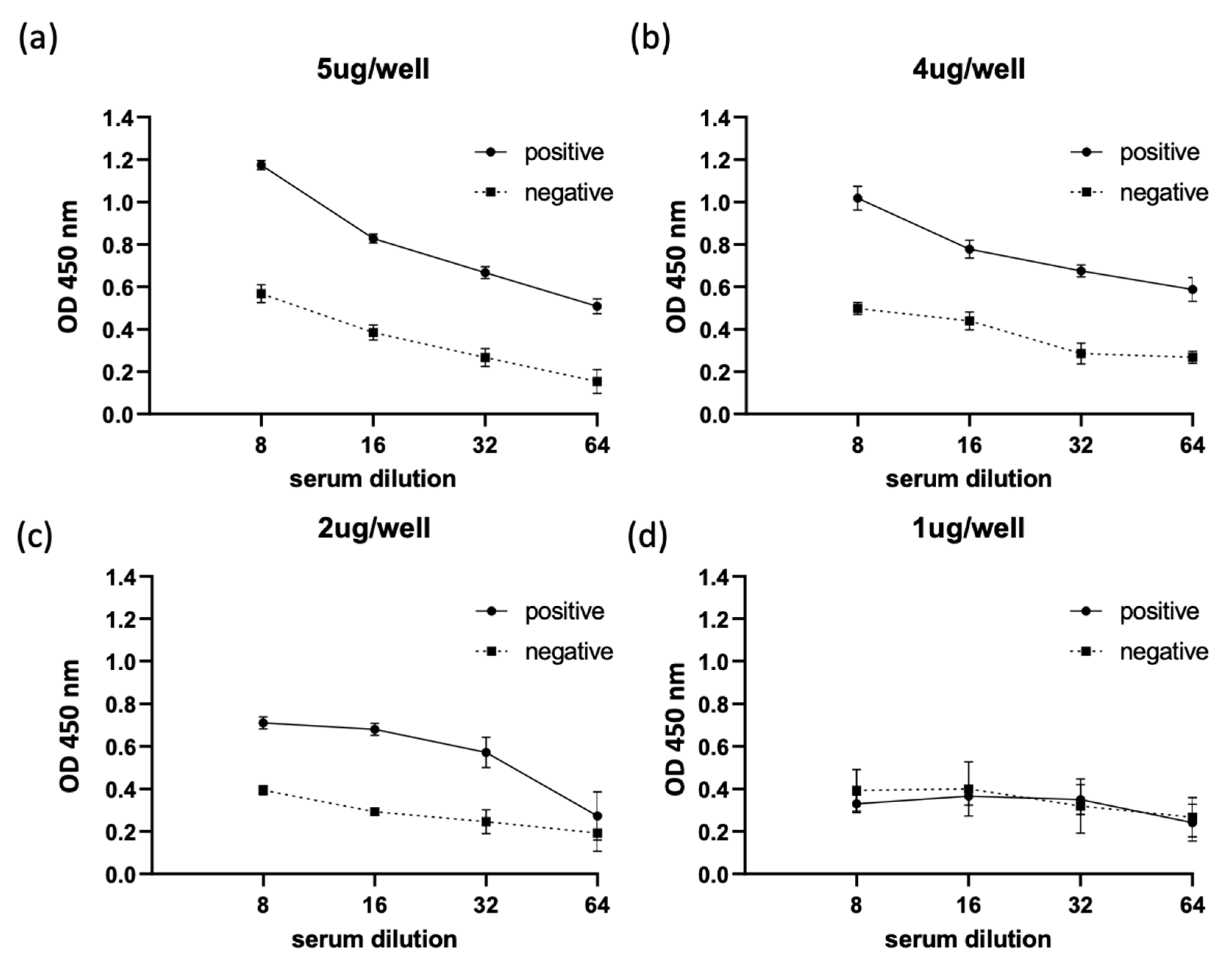
2.3. Optimization of ThEMA11 as an Antigen for Use in the Indirect ELISA Fformat
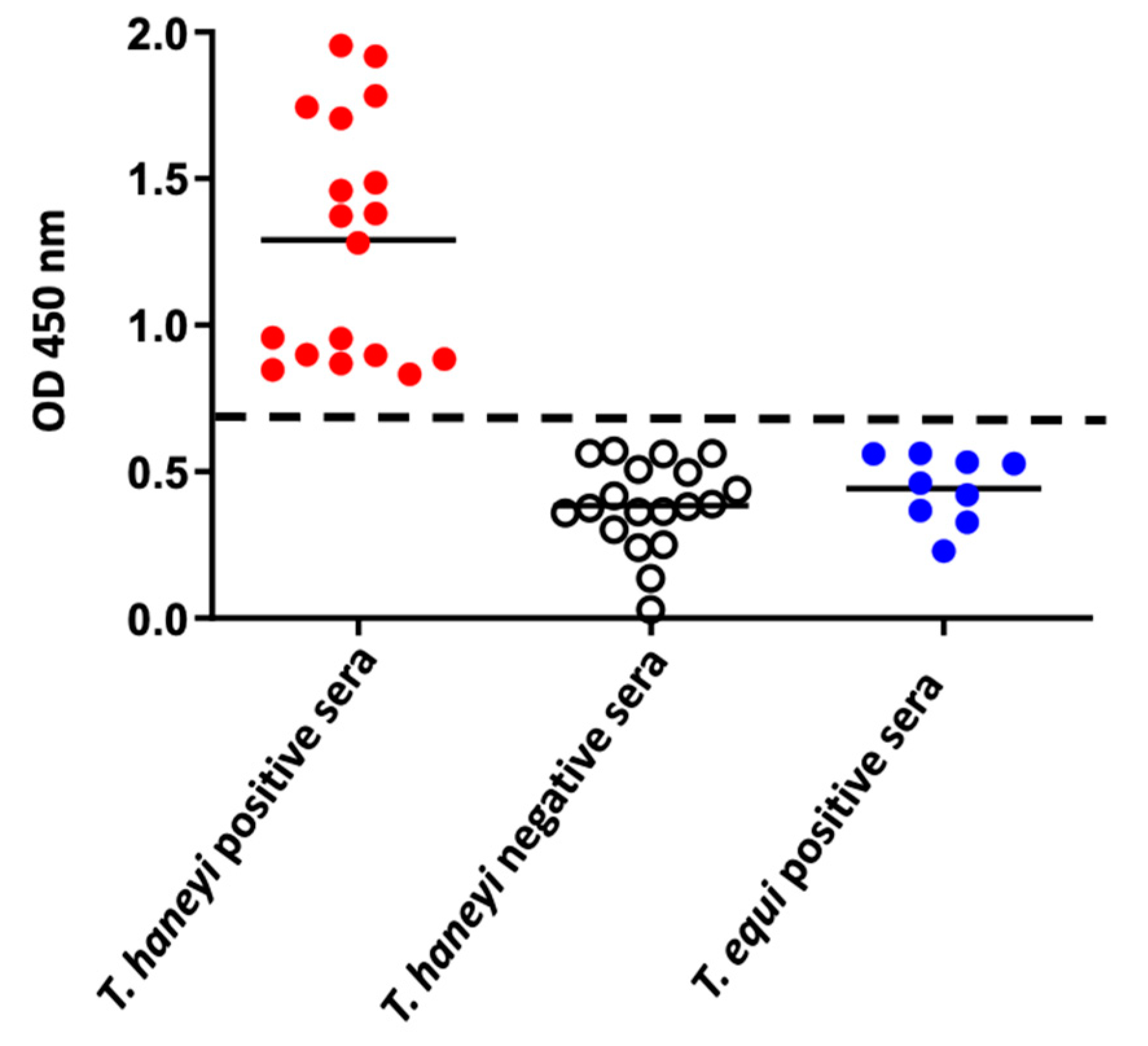
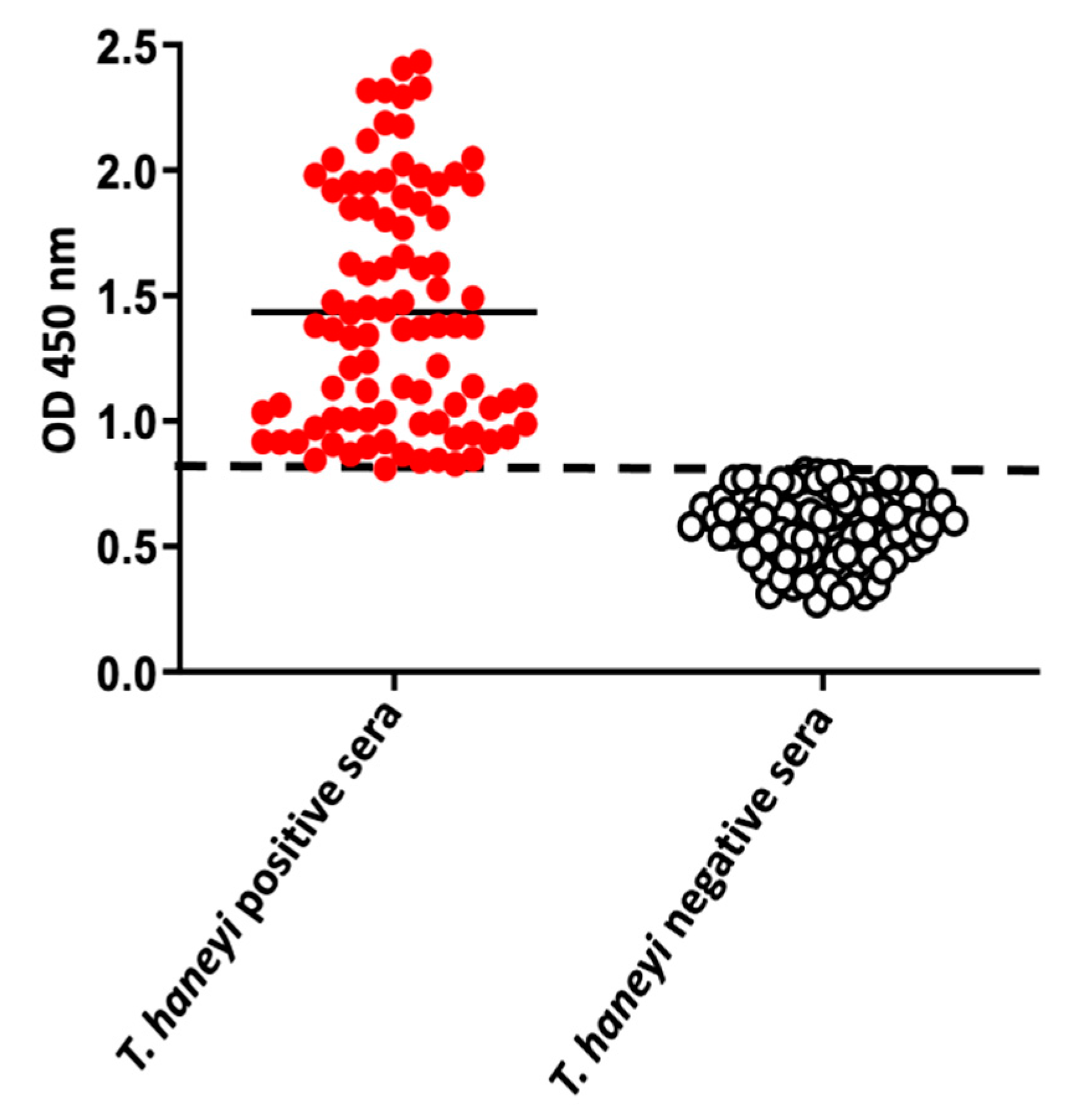
2.4. Use of the ThEMA11-Based ELISA to Detect Antibodies to T. haneyi in Horse Sera from Different Geographic Regions
3. Discussion
4. Materials and Methods
4.1. T. haneyi EMA11 Cloning, Expression, and Purification
4.2. Horse Serum Samples
4.3. Competitive ELISA (cELISA) for T. equi or B. caballi
4.4. Immunoblot Analysis
4.5. Indirect ELISA
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bishop, R.P.; Kappmeyer, L.S.; Onzere, C.K.; Odongo, D.O.; Githaka, N.; Sears, K.P.; Knowles, D.P.; Fry, L.M. Equid infective Theileria cluster in distinct 18S rRNA gene clades comprising multiple taxa with unusually broad mammalian host ranges. Parasit Vectors 2020, 13, 261. [Google Scholar] [CrossRef] [PubMed]
- Bhoora, R.V.; Collins, N.E.; Schnittger, L.; Troskie, C.; Marumo, R.; Labuschagne, K.; Smith, R.M.; Dalton, D.L.; Mbizeni, S. Molecular genotyping and epidemiology of equine piroplasmids in South Africa. Ticks Tick Borne Dis. 2019, 11, 101358. [Google Scholar] [CrossRef] [PubMed]
- Knowles, D.P.; Kappmeyer, L.S.; Haney, D.; Herndon, D.R.; Fry, L.M.; Munro, J.B.; Sears, K.; Ueti, M.W.; Wise, L.N.; Silva, M.; et al. Discovery of a novel species, Theileria haneyi n. sp., infective to equids, highlights exceptional genomic diversity within the genus Theileria: Implications for apicomplexan parasite surveillance. Int. J. Parasitol. 2018, 48, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Vieira, M.I.B.; Costa, M.M.; de Oliveira, M.T.; Goncalves, L.R.; Andre, M.R.; Machado, R.Z. Serological detection and molecular characterization of piroplasmids in equids in Brazil. Acta Trop. 2018, 179, 81–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mshelia, P.W.; Kappmeyer, L.; Johnson, W.C.; Kudi, C.A.; Oluyinka, O.O.; Balogun, E.O.; Richard, E.E.; Onoja, E.; Sears, K.P.; Ueti, M.W. Molecular detection of Theileria species and Babesia caballi from horses in Nigeria. Parasitol. Res. 2020, 119, 2955–2963. [Google Scholar] [CrossRef] [PubMed]
- Sears, K.P.; Kappmeyer, L.S.; Wise, L.N.; Silva, M.; Ueti, M.W.; White, S.; Reif, K.E.; Knowles, D.P. Infection dynamics of Theileria equi and Theileria haneyi, a newly discovered apicomplexan of the horse. Vet. Parasitol. 2019, 271, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Sears, K.; Knowles, D.; Dinkel, K.; Mshelia, P.W.; Onzere, C.; Silva, M.; Fry, L. Imidocarb Dipropionate Lacks Efficacy against Theileria haneyi and Fails to Consistently Clear Theileria equi in Horses Co-Infected with T. haneyi. Pathogens 2020, 9, 1035. [Google Scholar] [CrossRef] [PubMed]
- Wise, L.N.; Kappmeyer, L.S.; Knowles, D.P.; White, S.N. Evolution and diversity of the EMA families of the divergent equid parasites, Theileria equi and T. haneyi. Infect. Genet. Evol. 2019, 68, 153–160. [Google Scholar] [CrossRef]
- Knowles, D.P., Jr.; Perryman, L.E.; Goff, W.L.; Miller, C.D.; Harrington, R.D.; Gorham, J.R. A monoclonal antibody defines a geographically conserved surface protein epitope of Babesia equi merozoites. Infect. Immun. 1991, 59, 2412–2417. [Google Scholar] [CrossRef] [Green Version]
- Cunha, C.W.; Kappmeyer, L.S.; McGuire, T.C.; Dellagostin, O.A.; Knowles, D.P. Conformational Dependence and Conservation of an Immunodominant Epitope within the Babesia equi Erythrocyte-Stage Surface Protein Equi Merozoite Antigen 1. Clin. Vaccine Immunol. 2002, 9, 1301–1306. [Google Scholar] [CrossRef] [Green Version]
- Ueti, M.W.; Palmer, G.H.; Kappmeyer, L.S.; Scoles, G.A.; Knowles, D.P. Expression of Equi Merozoite Antigen 2 during Development of Babesia equi in the Midgut and Salivary Gland of the Vector Tick Boophilus microplus. J. Clin. Microbiol. 2003, 41, 5803–5809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wise, L.N.; Kappmeyer, L.S.; Silva, M.G.; White, S.N.; Grause, J.F.; Knowles, D.P. Verification of post-chemotherapeutic clearance of Theileria equi through concordance of nested PCR and immunoblot. Ticks Tick Borne Dis. 2018, 9, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Wise, L.N.; Kappmeyer, L.S.; Mealey, R.H.; Knowles, D.P. Review of equine piroplasmosis. J. Vet. Intern. Med. 2013, 27, 1334–1346. [Google Scholar] [CrossRef]
- Schrader, C.; Schielke, A.; Ellerbroek, L.; Johne, R. PCR inhibitors—Occurrence, properties and removal. J. Appl. Microbiol. 2012, 113, 1014–1026. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.G.; Villarino, N.F.; Knowles, D.P.; Suarez, C.E. Assessment of Draxxin® (tulathromycin) as an inhibitor of in vitro growth of Babesia bovis, Babesia bigemina and Theileria equi. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Bowden, G.D.; Reis, P.M.; Rogers, M.B.; Bone Relat, R.M.; Brayton, K.A.; Wilson, S.K.; Di Genova, B.M.; Knoll, L.J.; Nepveux, V.F.; Tai, A.K.; et al. A conserved coccidian gene is involved in Toxoplasma sensitivity to the anti-apicomplexan compound, tartrolon E. Int. J. Parasitol. Drugs Drug Resist. 2020, 14, 1–7. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, R.M.; Nepveux, V.F.; Abenoja, J.; Bowden, G.; Reis, P.; Beaushaw, J.; Bone Relat, R.M.; Driskell, I.; Gimenez, F.; Riggs, M.W.; et al. A symbiotic bacterium of shipworms produces a compound with broad spectrum anti-apicomplexan activity. PLoS Pathog. 2020, 16, e1008600. [Google Scholar] [CrossRef] [PubMed]
- Bastos, R.G.; Sears, K.; Dinkel, K.D.; Knowles, D.P.; Fry, L.M. Changes in the Molecular and Functional Phenotype of Bovine Monocytes during Theileria parva Infection. Infect. Immun. 2019, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knowles, D.P.; Kappmeyer, L.S.; Stiller, D.; Hennager, S.G.; Perryman, L.E. Antibody to a Recombinant Merozoite Protein Epitope Identifies Horses Infected with Babesia equi. J. Clin. Microbiol. 1992, 30, 3122–3126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kappmeyer, L.S.; Perryman, L.E.; Hines, S.A.; Baszler, T.V.; Katz, J.B.; Hennager, S.G.; Knowles, D.P. Detection of equine antibodies to Babesia caballi by recombinant B. caballi rhoptry-associated protein 1 in a competitive-inhibition enzyme-linked immunosorbent assay. J. Clin. Microbiol. 1999, 37, 2285–2290. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T. haneyi Positive Samples (n = 87) | T. haneyi Negative Samples (n = 81) | ||
|---|---|---|---|
| T. equi positive | T. equi negative | T. equi positive | T. equi negative |
| 58.62% (n = 51) | 41.37% (n = 36) | 62.96% (n = 51) | 37.07% (n = 30) |
| T. haneyi Positive Samples (n = 87) | T. haneyi Negative Samples (n = 81) | ||
|---|---|---|---|
| B. caballi positive | B. caballi negative | B. caballi positive | B. caballi negative |
| 26.92% (n = 7) | 73.07% (n = 19) | 24.41% (n = 5) | 70.58% (n = 12) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bastos, R.G.; Sears, K.P.; Dinkel, K.D.; Kappmeyer, L.; Ueti, M.W.; Knowles, D.P.; Fry, L.M. Development of an Indirect ELISA to Detect Equine Antibodies to Theileria haneyi . Pathogens 2021, 10, 270. https://doi.org/10.3390/pathogens10030270
Bastos RG, Sears KP, Dinkel KD, Kappmeyer L, Ueti MW, Knowles DP, Fry LM. Development of an Indirect ELISA to Detect Equine Antibodies to Theileria haneyi . Pathogens. 2021; 10(3):270. https://doi.org/10.3390/pathogens10030270
Chicago/Turabian StyleBastos, Reginaldo G., Kelly P. Sears, Kelcey D. Dinkel, Lowell Kappmeyer, Massaro W. Ueti, Donald P. Knowles, and Lindsay M. Fry. 2021. "Development of an Indirect ELISA to Detect Equine Antibodies to Theileria haneyi " Pathogens 10, no. 3: 270. https://doi.org/10.3390/pathogens10030270