
Molecular Diversity of Giardia duodenalis, Cryptosporidium spp., and Blastocystis sp. in Symptomatic and Asymptomatic Schoolchildren in Zambézia Province (Mozambique)


Abstract
:1. Introduction
2. Results
2.1. Occurrence of Enteric Parasites
2.2. Prevalence and Molecular Characterization of G. duodenalis
2.3. Prevalence and Molecular Characterization of Cryptosporidium spp.
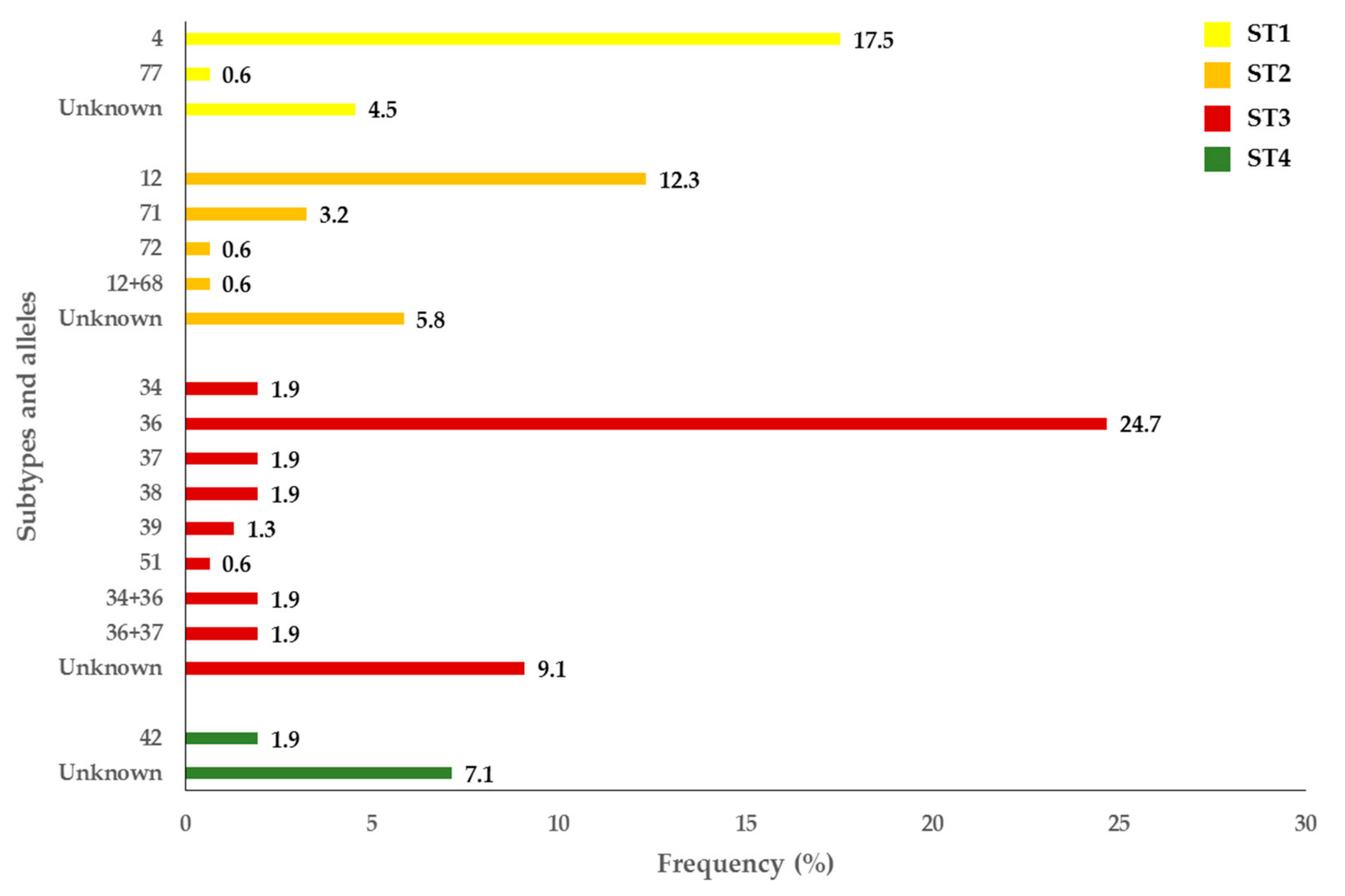
2.4. Prevalence and Molecular Characterization of Blastocystis sp.
3. Discussion
4. Materials and Methods
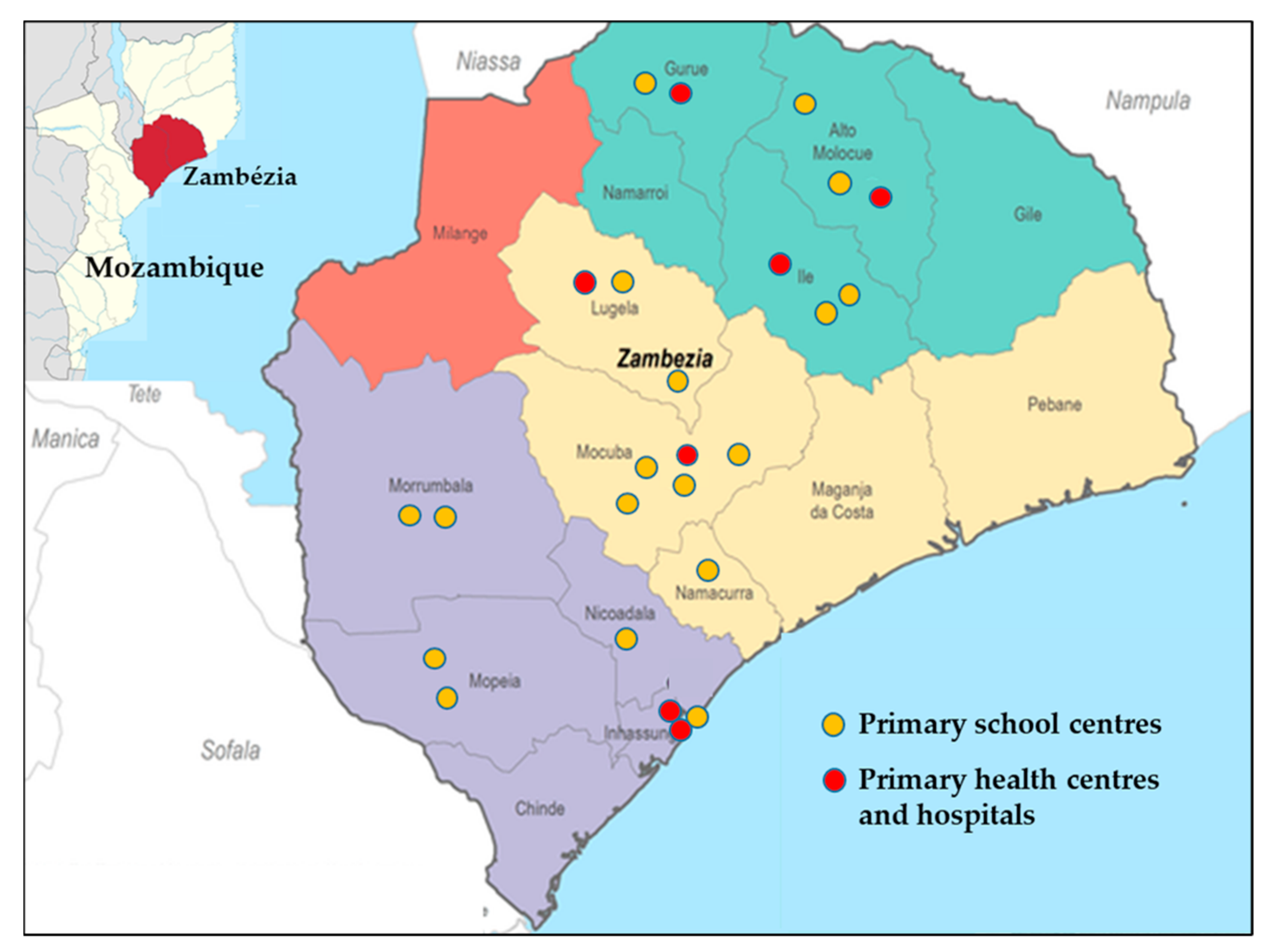
4.1. Study Area and Stool Sample Collection
4.2. DNA Extraction and Purification
4.3. Molecular Detection and Characterization of Giardia duodenalis
4.4. Molecular Detection and Characterization of Cryptosporidium spp.
4.5. Molecular Detection and Characterization of Blastocystis sp.
4.6. Sequence and Phylogenetic Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Snyder, J.D.; Merson, M.H. The magnitude of the global problem of acute diarrhoeal disease: A review of active surveillance data. Bull. World Health Org. 1982, 60, 605–613. [Google Scholar] [PubMed]
- GBD 2016 Diarrhoeal Disease Collaborators. Estimates of the global, regional, and national morbidity, mortality, and aetiologies of diarrhoea in 195 countries: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Infect. Dis. 2018, 18, 1211–1228. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Johnson, H.L.; Cousens, S.; Perin, J.; Scott, S.; Lawn, J.E.; Rudan, I.; Campbell, H.; Cibulskis, R.; Li, M.; et al. Global, regional, and national causes of child mortality: An updated systematic analysis for 2010 with time trends since 2000. Lancet 2012, 379, 2151–2161. [Google Scholar] [CrossRef]
- Black, R.E.; Morris, S.S.; Bryce, J. Where and why are 10 million children dying every year? Lancet 2003, 361, 2226–2234. [Google Scholar] [CrossRef]
- GBD Diarrhoeal Diseases Collaborators. Estimates of global, regional, and national morbidity, mortality, and aetiologies of diarrhoeal diseases: A systematic analysis for the Global Burden of Disease Study 2015. Lancet Infect. Dis. 2017, 17, 909–948. [Google Scholar] [CrossRef] [Green Version]
- Torgerson, P.R.; Devleesschauwer, B.; Praet, N.; Speybroeck, N.; Willingham, A.L.; Kasuga, F.; Rokni, M.B.; Zhou, X.N.; Fèvre, E.M.; Sripa, B.; et al. World Health Organization estimates of the global and regional disease burden of 11 foodborne parasitic diseases, 2010: A data synthesis. PLoS Med. 2015, 12, e1001920. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. The World Health Report—Fighting Disease Fostering Development; World Health Organization: Geneva, Switzerland, 1996; Available online: http://www.who.int/whr/1996/en/ (accessed on 26 January 2021).
- Kotloff, K.L.; Nataro, J.P.; Blackwelder, W.C.; Nasrin, D.; Farag, T.H.; Panchalingam, S.; Wu, Y.; Sow, S.O.; Sur, D.; Breiman, R.F.; et al. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): A prospective, case-control study. Lancet 2013, 382, 209–222. [Google Scholar] [CrossRef]
- Tan, K.S.; Mirza, H.; Teo, J.D.; Wu, B.; Macary, P.A. Current views on the clinical relevance of Blastocystis spp. Curr. Infect. Dis. Rep. 2010, 12, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Halliez, M.C.; Buret, A.G. Extra-intestinal and long term consequences of Giardia duodenalis infections. World J. Gastroenterol. 2013, 19, 8974–8985. [Google Scholar] [CrossRef] [PubMed]
- Korpe, P.S.; Valencia, C.; Haque, R.; Mahfuz, M.; McGrath, M.; Houpt, E.; Kosek, M.; McCormick, B.J.J.; Penataro Yori, P.; Babji, S.; et al. Epidemiology and risk factors for cryptosporidiosis in children from 8 low-income sites: Results from the MAL-ED Study. Clin. Infect. Dis. 2018, 67, 1660–1669. [Google Scholar] [CrossRef] [Green Version]
- Efstratiou, A.; Ongerth, J.E.; Karanis, P. Waterborne transmission of protozoan parasites: Review of worldwide outbreaks—An update 2011–2016. Water Res. 2017, 114, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Ryan, U.; Hijjawi, N.; Xiao, L. Foodborne cryptosporidiosis. Int. J. Parasitol. 2018, 48, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, U.; Cacciò, S.M. Zoonotic potential of Giardia. Int. J. Parasitol. 2013, 43, 943–956. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Ryan, U.; Xiao, L. Genetic diversity and population structure of Cryptosporidium. Trends Parasitol. 2018, 34, 997–1011. [Google Scholar] [CrossRef]
- Stensvold, C.R.; Clark, C.G. Pre-empting Pandora’s box: Blastocystis subtypes revisited. Trends Parasitol. 2020, 36, 229–232. [Google Scholar] [CrossRef]
- Mor, S.M.; Tzipori, S. Cryptosporidiosis in children in Sub-Saharan Africa: A lingering challenge. Clin. Infect. Dis. 2008, 47, 915–921. [Google Scholar] [CrossRef] [Green Version]
- Aldeyarbi, H.M.; El-Ezz, N.M.A.; Karanis, P. Cryptosporidium and cryptosporidiosis: The African perspective. Environ. Sci. Pollut. Res. Int. 2016, 23, 13811–13821. [Google Scholar] [CrossRef]
- Squire, S.A.; Ryan, U. Cryptosporidium and Giardia in Africa: Current and future challenges. Parasit. Vectors 2017, 10, 195. [Google Scholar] [CrossRef] [Green Version]
- Khorshidvand, Z.; Khazaei, S.; Amiri, M.; Taherkhani, H.; Mirzaei, A. Worldwide prevalence of emerging parasite Blastocystis in immunocompromised patients: A systematic review and meta-analysis. Microb. Pathog. 2021, in press. [Google Scholar] [CrossRef]
- Meurs, L.; Polderman, A.M.; Vinkeles Melchers, N.V.; Brienen, E.A.; Verweij, J.J.; Groosjohan, B.; Mendes, F.; Mechendura, M.; Hepp, D.H.; Langenberg, M.C.; et al. Diagnosing polyparasitism in a high-prevalence setting in Beira, Mozambique: Detection of intestinal parasites in fecal samples by microscopy and real-time PCR. PLoS Negl. Trop. Dis. 2017, 11, e0005310. [Google Scholar] [CrossRef]
- Platts-Mills, J.A.; Babji, S.; Bodhidatta, L.; Gratz, J.; Haque, R.; Havt, A.; McCormick, B.J.; McGrath, M.; Olortegui, M.P.; Samie, A.; et al. MAL-ED Network Investigators. Pathogen-specific burdens of community diarrhoea in developing countries: A multisite birth cohort study (MAL-ED). Lancet Glob. Health 2015, 3, e564–e575. [Google Scholar] [CrossRef] [Green Version]
- Messa, A., Jr.; Köster, P.C.; Garrine, M.; Gilchrist, C.; Bartelt, L.A.; Nhampossa, T.; Massora, S.; Kotloff, K.; Levine, M.M.; Alonso, P.L.; et al. Molecular diversity of Giardia duodenalis in children under 5 years from the Manhiça district, Southern Mozambique enrolled in a matched case-control study on the aetiology of diarrhoea. PLoS Negl. Trop. Dis. 2021, 15, e0008987. [Google Scholar] [CrossRef]
- Messa, A., Jr.; Köster, P.C.; Garrine, M.; Nhampossa, T.; Cossa, A.; Kotloff, K.; Levine, M.M.; Alonso, P.L.; Carmena, D.; Mandomando, I. Molecular characterization of Cryptosporidium spp. in children younger than 5 years enrolled in a matched case-control study on the aetiology of diarrhoea in the Manhiça district, Southern Mozambique. Pathogens 2021. under review. [Google Scholar]
- Irisarri-Gutiérrez, M.J.; Mingo, M.H.; de Lucio, A.; Gil, H.; Morales, L.; Seguí, R.; Nacarapa, E.; Muñoz-Antolí, C.; Bornay-Llinares, F.J.; Esteban, J.G.; et al. Association between enteric protozoan parasites and gastrointestinal illness among HIV- and tuberculosis-infected individuals in the Chowke district, southern Mozambique. Acta Trop. 2017, 170, 197–203. [Google Scholar] [CrossRef]
- Casmo, V.; Lebbad, M.; Maungate, S.; Lindh, J. Occurrence of Cryptosporidium spp. and Cystoisospora belli among adult patients with diarrhoea in Maputo, Mozambique. Heliyon 2018, 4, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Muadica, A.S.; Balasegaram, S.; Beebeejaun, K.; Köster, P.C.; Bailo, B.; Hernández-de-Mingo, M.; Dashti, A.; Dacal, E.; Saugar, J.M.; Fuentes, I.; et al. Risk associations for intestinal parasites in symptomatic and asymptomatic schoolchildren in central Mozambique. Clin. Microbiol. Infect. 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- De Lucio, A.; Amor-Aramendía, A.; Bailo, B.; Saugar, J.M.; Anegagrie, M.; Arroyo, A.; López-Quintana, B.; Zewdie, D.; Ayehubizu, Z.; Yizengaw, E.; et al. Prevalence and genetic diversity of Giardia duodenalis and Cryptosporidium spp. among school children in a rural area of the Amhara Region, North-West Ethiopia. PLoS ONE 2016, 11, e0159992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dacal, E.; Saugar, J.M.; de Lucio, A.; Hernández-de-Mingo, M.; Robinson, E.; Köster, P.C.; Aznar-Ruiz-de-Alegría, M.L.; Espasa, M.; Ninda, A.; Gandasegui, J.; et al. Prevalence and molecular characterization of Strongyloides stercoralis, Giardia duodenalis, Cryptosporidium spp., and Blastocystis spp. isolates in school children in Cubal, Western Angola. Parasit. Vectors 2018, 11, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muadica, A.S.; Messa, A.E., Jr.; Dashti, A.; Balasegaram, S.; Santin, M.; Manjate, F.; Chirinda, P.; Garrine, M.; Vubil, D.; Acácio, S.; et al. First identification of genotypes of Enterocytozoon bieneusi (Microsporidia) among symptomatic and asymptomatic children in Mozambique. PLoS Negl. Trop. Dis. 2020, 14, e0008419. [Google Scholar] [CrossRef] [PubMed]
- Mandomando, I.M.; Macete, E.V.; Ruiz, J.; Sanz, S.; Abacassamo, F.; Vallès, X.; Sacarlal, J.; Navia, M.M.; Vila, J.; Alonso, P.L.; et al. Etiology of diarrhea in children younger than 5 years of age admitted in a rural hospital of southern Mozambique. Am. J. Trop. Med. Hyg. 2007, 76, 522–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca, A.M.; Fernandes, N.; Ferreira, F.S.; Gomes, J.; Centeno-Lima, S. Intestinal parasites in children hospitalized at the Central Hospital in Maputo, Mozambique. J. Infect. Dev. Ctries. 2014, 8, 786–789. [Google Scholar] [CrossRef]
- Cerveja, B.Z.; Tucuzo, R.M.; Madureira, A.C.; Nhacupe, N.; Langa, I.A.; Buene, T.; Banze, L.; Funzamo, C.; Noormahomed, E.V. Prevalence of intestinal parasites among HIV Infected and HIV uninfected patients treated at the 1° De Maio Health Centre in Maputo, Mozambique. EC Microbiol. 2017, 9, 231–240. [Google Scholar]
- Nhampossa, T.; Mandomando, I.; Acacio, S.; Quintó, L.; Vubil, D.; Ruiz, J.; Nhalungo, D.; Sacoor, C.; Nhabanga, A.; Nhacolo, A.; et al. Diarrheal disease in rural Mozambique: Burden, risk factors and etiology of diarrheal disease among children aged 0-59 months seeking care at health facilities. PLoS ONE 2015, 10, e0119824. [Google Scholar] [CrossRef]
- Acácio, S.; Mandomando, I.; Nhampossa, T.; Quintó, L.; Vubil, D.; Sacoor, C.; Kotloff, K.; Farag, T.; Nasrin, D.; Macete, E.; et al. Risk factors for death among children 0–59 months of age with moderate-to-severe diarrhea in Manhiça district, southern Mozambique. BMC Infect. Dis. 2019, 19, 322. [Google Scholar] [CrossRef] [PubMed]
- Bauhofer, A.F.L.; Cossa-Moiane, I.; Marques, S.; Guimarães, E.L.; Munlela, B.; Anapakala, E.; Chilaúle, J.J.; Cassocera, M.; Langa, J.S.; Chissaque, A.; et al. Intestinal protozoan infections among children 0–168 months with diarrhea in Mozambique: June 2014–January 2018. PLoS Negl. Trop. Dis. 2020, 14, e0008195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sow, S.O.; Muhsen, K.; Nasrin, D.; Blackwelder, W.C.; Wu, Y.; Farag, T.H.; Panchalingam, S.; Sur, D.; Zaidi, A.K.; Faruque, A.S.; et al. The burden of Cryptosporidium diarrheal disease among children <24 months of age in moderate/high mortality regions of sub-Saharan Africa and South Asia, utilizing data from the Global Enteric Multicenter Study (GEMS). PLoS Negl. Trop. Dis. 2016, 10, e0004729. [Google Scholar]
- Slapeta, J. Cryptosporidium species found in cattle: A proposal for a new species. Trends Parasitol. 2006, 22, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Adamu, H.; Petros, B.; Zhang, G.; Kassa, H.; Amer, S.; Ye, J.; Feng, Y.; Xiao, L. Distribution and clinical manifestations of Cryptosporidium species and subtypes in HIV/AIDS patients in Ethiopia. PLoS Negl. Trop. Dis. 2014, 8, e2831. [Google Scholar] [CrossRef] [Green Version]
- Akinbo, F.O.; Okaka, C.E.; Omoregie, R.; Dearen, T.; Leon, E.T.; Xiao, L. Molecular characterization of Cryptosporidium spp. in HIV-infected persons in Benin City, Edo State, Nigeria. Fooyin J. Health Sci. 2010, 2, 85–89. [Google Scholar] [CrossRef] [Green Version]
- Gatei, W.; Wamae, C.N.; Mbae, C.; Waruru, A.; Mulinge, E.; Waithera, T.; Gatika, S.M.; Kamwati, S.K.; Revathi, G.; Hart, C.A. Cryptosporidiosis: Prevalence, genotype analysis, and symptoms associated with infections in children in Kenya. Am. J. Trop. Med. Hyg. 2006, 75, 78–82. [Google Scholar] [CrossRef] [Green Version]
- Elwin, K.; Hadfield, S.J.; Robinson, G.; Crouch, N.D.; Chalmers, R.M. Cryptosporidium viatorum n. sp. (Apicomplexa: Cryptosporidiidae) among travellers returning to Great Britain from the Indian subcontinent, 2007–2011. Int. J. Parasitol. 2012, 42, 675–682. [Google Scholar] [CrossRef]
- Koehler, A.V.; Wang, T.; Haydon, S.R.; Gasser, R.B. Cryptosporidium viatorum from the native Australian swamp rat Rattus lutreolus—An emerging zoonotic pathogen? Int. J. Parasitol. Parasites Wildl. 2018, 7, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Zhou, H.; Huang, Y.; Xu, L.; Rao, L.; Wang, S.; Wang, W.; Yi, Y.; Zhou, X.; Wu, Y.; et al. Cryptosporidium spp. in wild rats (Rattus spp.) from the Hainan Province, China: Molecular detection, species/genotype identification and implications for public health. Int. J. Parasitol. Parasites Wildl. 2019, 9, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Stensvold, C.R.; Elwin, K.; Winiecka-Krusnell, J.; Chalmers, R.M.; Xiao, L.; Lebbad, M. Development and application of a gp60-based typing assay for Cryptosporidium viatorum. J. Clin. Microbiol. 2015, 53, 1891–1897. [Google Scholar] [CrossRef] [Green Version]
- D’Alfonso, R.; Santoro, M.; Essi, D.; Monsia, A.; Kaboré, Y.; Glé, C.; Di Cave, D.; Sorge, R.P.; Di Cristanziano, V.; Berrilli, F. Blastocystis in Côte d’Ivoire: Molecular identification and epidemiological data. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2243–2250. [Google Scholar] [CrossRef]
- Greigert, V.; Abou-Bacar, A.; Brunet, J.; Nourrisson, C.; Pfaff, A.W.; Benarbia, L.; Pereira, B.; Randrianarivelojosia, M.; Razafindrakoto, J.L.; Rakotomalala, R.S.; et al. Human intestinal parasites in Mahajanga, Madagascar: The kingdom of the protozoa. PLoS ONE 2018, 13, e0204576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfellani, M.A.; Stensvold, C.R.; Vidal-Lapiedra, A.; Onuoha, E.S.; Fagbenro-Beyioku, A.F.; Clark, C.G. Variable geographic distribution of Blastocystis subtypes and its potential implications. Acta Trop. 2013, 126, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Stensvold, C.R.; Alfellani, M.; Clark, C.G. Levels of genetic diversity vary dramatically between Blastocystis subtypes. Infect. Genet. Evol. 2012, 12, 263–273. [Google Scholar] [CrossRef] [Green Version]
- El Safadi, D.; Gaayeb, L.; Meloni, D.; Cian, A.; Poirier, P.; Wawrzyniak, I.; Delbac, F.; Dabboussi, F.; Delhaes, L.; Seck, M.; et al. Children of Senegal River Basin show the highest prevalence of Blastocystis sp. ever observed worldwide. BMC Infect. Dis. 2014, 14, 164. [Google Scholar] [CrossRef]
- Abda, I.B.; Maatoug, N.; Romdhane, R.B.; Bouhelmi, N.; Zallegua, N.; Aoun, K.; Viscogliosi, E.; Bouratbine, A. Prevalence and subtype identification of Blastocystis sp. in healthy individuals in the Tunis area, Tunisia. Am. J. Trop. Med. Hyg. 2017, 96, 202–204. [Google Scholar] [CrossRef] [Green Version]
- Stensvold, C.R.; Christiansen, D.B.; Olsen, K.E.; Nielsen, H.V. Blastocystis sp. subtype 4 is common in Danish Blastocystis-positive patients presenting with acute diarrhea. Am. J. Trop. Med. Hyg. 2011, 84, 883–885. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Márquez, M.V.; Guna, R.; Muñoz, C.; Gómez-Muñoz, M.T.; Borrás, R. High prevalence of subtype 4 among isolates of Blastocystis hominis from symptomatic patients of a health district of Valencia (Spain). Parasitol. Res. 2009, 105, 949–955. [Google Scholar] [CrossRef] [PubMed]
- Mattiucci, S.; Crisafi, B.; Gabrielli, S.; Paoletti, M.; Cancrini, G. Molecular epidemiology and genetic diversity of Blastocystis infection in humans in Italy. Epidemiol. Infect. 2016, 144, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Verweij, J.J.; Schinkel, J.; Laeijendecker, D.; van Rooyen, M.A.; van Lieshout, L.; Polderman, A.M. Real-time PCR for the detection of Giardia lamblia. Mol. Cell. Probes 2003, 17, 223–225. [Google Scholar] [CrossRef]
- Read, C.M.; Monis, P.T.; Thompson, R.C. Discrimination of all genotypes of Giardia duodenalis at the glutamate dehydrogenase locus using PCR-RFLP. Infect. Genet. Evol. 2004, 4, 125–130. [Google Scholar] [CrossRef]
- Lalle, M.; Pozio, E.; Capelli, G.; Bruschi, F.; Crotti, D.; Cacciò, S.M. Genetic heterogeneity at the beta-giardin locus among human and animal isolates of Giardia duodenalis and identification of potentially zoonotic subgenotypes. Int. J. Parasitol. 2005, 35, 207–213. [Google Scholar] [CrossRef]
- Sulaiman, I.M.; Fayer, R.; Bern, C.; Gilman, R.H.; Trout, J.M.; Schantz, P.M.; Das, P.; Lal, A.A.; Xiao, L. Triosephosphate isomerase gene characterization and potential zoonotic transmission of Giardia duodenalis. Emerg. Infect. Dis. 2003, 9, 1444–1452. [Google Scholar] [CrossRef] [PubMed]
- Tiangtip, R.; Jongwutiwes, S. Molecular analysis of Cryptosporidium species isolated from HIV-infected patients in Thailand. Trop. Med. Int. Health 2002, 7, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Feltus, D.C.; Giddings, C.W.; Schneck, B.L.; Monson, T.; Warshauer, D.; McEvoy, J.M. Evidence supporting zoonotic transmission of Cryptosporidium spp. in Wisconsin. J. Clin. Microbiol. 2006, 44, 4303–4308. [Google Scholar] [CrossRef] [Green Version]
- Scicluna, S.M.; Tawari, B.; Clark, C.G. DNA barcoding of Blastocystis. Protist 2006, 157, 77–85. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Centre | Children (n) | Giardia duodenalis | Cryptosporidium spp. | Blastocystis sp. | |||
|---|---|---|---|---|---|---|---|
| Positive (n) | Positive (%) | Positive (n) | Positive (%) | Positive (n) | Positive (%) | ||
| School | |||||||
| 1 | 44 | 23 | 52.3 | 1 | 2.3 | 1 | 2.3 |
| 2 | 50 | 13 | 26.0 | 0 | 0.0 | 23 | 46.0 |
| 3 | 22 | 9 | 40.9 | 0 | 0.0 | 0 | 0.0 |
| 4 | 22 | 12 | 54.5 | 0 | 0.0 | 0 | 0.0 |
| 5 | 24 | 11 | 45.8 | 0 | 0.0 | 0 | 0.0 |
| 6 | 31 | 13 | 41.9 | 0 | 0.0 | 1 | 3.2 |
| 7 | 88 | 36 | 40.9 | 4 | 4.5 | 9 | 10.2 |
| 8 | 60 | 8 | 13.3 | 2 | 3.3 | 0 | 0.0 |
| 9 | 47 | 23 | 48.9 | 0 | 0.0 | 0 | 0.0 |
| 10 | 49 | 20 | 40.8 | 0 | 0.0 | 1 | 2.0 |
| 11 | 47 | 13 | 27.7 | 0 | 0.0 | 4 | 8.5 |
| 12 | 50 | 12 | 24.0 | 0 | 0.0 | 4 | 8.0 |
| 13 | 50 | 25 | 50.0 | 1 | 2.0 | 19 | 38.0 |
| 14 | 30 | 9 | 30.0 | 0 | 0.0 | 0 | 0.0 |
| 15 | 30 | 5 | 16.7 | 0 | 0.0 | 0 | 0.0 |
| 16 | 40 | 16 | 40.0 | 0 | 0.0 | 0 | 0.0 |
| 17 | 75 | 46 | 61.3 | 1 | 1.3 | 47 | 62.7 |
| 18 | 48 | 42 | 87.5 | 2 | 4.2 | 34 | 70.8 |
| Sub-total | 807 | 336 | 41.6 | 11 | 1.4 | 143 | 17.7 |
| Clinic | |||||||
| 1 | 50 | 32 | 64.0 | 3 | 6.0 | 2 | 4.0 |
| 2 | 25 | 11 | 44.0 | 0 | 0.0 | 0 | 0.0 |
| 3 | 15 | 4 | 26.7 | 0 | 0.0 | 2 | 13.3 |
| 4 | 52 | 28 | 53.8 | 0 | 0.0 | 4 | 7.7 |
| 5 | 42 | 10 | 23.8 | 2 | 4.8 | 2 | 4.8 |
| 6 | 29 | 4 | 13.8 | 1 | 3.4 | 1 | 3.4 |
| 7 | 73 | 31 | 42.5 | 0 | 0.0 | 0 | 0.0 |
| Sub-total | 286 | 120 | 42.0 | 6 | 2.1 | 11 | 3.8 |
| Total | 1093 | 456 | 41.7 | 17 | 1.6 | 154 | 14.1 |
| Sample ID | Ct Value in qPCR | gdh | bg | tpi | Assigned Genotype |
| 4 | 24.49 | BIV | Negative | Negative | BIV |
| 17 | 25.8 | BIV | Negative | Negative | BIV |
| 24 1 | 31.6 | BIV | Negative | Negative | BIV |
| 57 | 25.4 | BIV | Negative | Negative | BIV |
| 62 | 20.9 | AII | Negative | AII | AII |
| 67 | 23.9 | BIII/BIV | Negative | Negative | BIII/BIV |
| 69 | 21.9 | BIV | B | BIII | BIII/BIV |
| 70 | 24.3 | BIII/BIV | B | Negative | BIII/BIV |
| 74 | 22.7 | BIII | B | Negative | BIII |
| 79 | 26.1 | AII | Negative | Negative | AII |
| 88 | 20.9 | BIII/BIV | Negative | BIII | BIII/BIV |
| 97 | 26.1 | BIV | Negative | Negative | BIV |
| 100 | 25.9 | BIII/BIV | B | Negative | BIII/BIV |
| 103 | 27.5 | BIII/BIV | Negative | Negative | BIII/BIV |
| 104 | 21.7 | BIII/BIV | B | BIII | BIII/BIV |
| 105 | 24.0 | BIII/BIV | B | Negative | BIII/BIV |
| 118 | 26.8 | BIV | B | Negative | BIV |
| 122 | 20.0 | BIII/BIV | B | BIII/BIV | BIII/BIV |
| 124 | 30.8 | Negative | B | Negative | B |
| 128 | 31.8 | Negative | B | Negative | B |
| 140 | 30.5 | Negative | B | Negative | B |
| 141 | 19.9 | BIII | B | BIII/BIV | BIII/BIV |
| 143 | 25.3 | BIII | Negative | Negative | BIII |
| 164 | 20.9 | AII | AII | AII | AII |
| 165 | 24.1 | BIII | B | Negative | BIII |
| 170 | 28.7 | BIII/BIV | Negative | Negative | BIII/BIV |
| 172 | 25.6 | AII | B | Negative | AII+B |
| 173 | 29.5 | BIV | Negative | Negative | BIV |
| 175 | 23.7 | BIII/BIV | B | Negative | BIII/BIV |
| 176 | 28.5 | BIII/BIV | B | Negative | BIII/BIV |
| 178 | 27.1 | BIV | Negative | Negative | BIV |
| 179 | 25.9 | BIII/BIV | Negative | Negative | BIII/BIV |
| 180 | 22.7 | BIII/BIV | B | AII | AII+BIII/BIV |
| 183 | 26.5 | Negative | B | Negative | B |
| 186 | 26.6 | BIII/BIV | Negative | BIII | BIII/BIV |
| 187 | 25.2 | BIII | Negative | BIII | BIII |
| 190 | 19.8 | BIII/BIV | B | BIII | BIII/BIV |
| 191 | 30.7 | Negative | B | Negative | B |
| 194 | 20.0 | BIII/BIV | B | BIII | BIII/BIV |
| 195 | 23.8 | BIII/BIV | B | BIII | BIII/BIV |
| 196 | 26.4 | Negative | Negative | BIII | BIII |
| 203 | 27.1 | BIII/BIV | B | Negative | BIII/BIV |
| 206 | 22.9 | BIII/BIV | B | BIII | BIII/BIV |
| Species | No. of Isolates | Reference Sequence | Stretch | Single Nucleotide Polymorphisms | GenBank ID |
|---|---|---|---|---|---|
| C. hominis | 2 | AF108865 | 608–979 | None | MW563962 |
| 1 | AF108865 | 540–952 | C607T, 697Del_T 1 | MW563963 | |
| 1 | AF108865 | 622–956 | A808R, G905R | MW563964 | |
| C. parvum | 1 | AF112571 | 573–997 | A646G, T649G, 686_689DelTAAT 1, A691T, C795Y, A892R | MW563965 |
| 1 | AF112571 | 630–997 | A646G, T649G, 686_689DelTAAT 1, A691T, C795Y, T837W, A892R | MW563966 2 | |
| 1 | AF112571 | 539–1,025 | A646G, T649G, 686_690DelTAATT 1, A691T, A892G | MW563967 | |
| 1 | AF112571 | 655–985 | Unknown 3 | – | |
| C. felis | 3 | AF108862 | 631–980 | 661InsT 4, T670A, 700DelT 1 | MW563968 |
| 1 | 648–987 | 661InsT, T670A,700DelT 1, 824InsT 4 | MW563969 | ||
| C. viatorum | 1 | KX174309 | 290–762 | None | MW563970 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muadica, A.S.; Köster, P.C.; Dashti, A.; Bailo, B.; Hernández-de-Mingo, M.; Balasegaram, S.; Carmena, D. Molecular Diversity of Giardia duodenalis, Cryptosporidium spp., and Blastocystis sp. in Symptomatic and Asymptomatic Schoolchildren in Zambézia Province (Mozambique). Pathogens 2021, 10, 255. https://doi.org/10.3390/pathogens10030255
Muadica AS, Köster PC, Dashti A, Bailo B, Hernández-de-Mingo M, Balasegaram S, Carmena D. Molecular Diversity of Giardia duodenalis, Cryptosporidium spp., and Blastocystis sp. in Symptomatic and Asymptomatic Schoolchildren in Zambézia Province (Mozambique). Pathogens. 2021; 10(3):255. https://doi.org/10.3390/pathogens10030255
Chicago/Turabian StyleMuadica, Aly S., Pamela C. Köster, Alejandro Dashti, Begoña Bailo, Marta Hernández-de-Mingo, Sooria Balasegaram, and David Carmena. 2021. "Molecular Diversity of Giardia duodenalis, Cryptosporidium spp., and Blastocystis sp. in Symptomatic and Asymptomatic Schoolchildren in Zambézia Province (Mozambique)" Pathogens 10, no. 3: 255. https://doi.org/10.3390/pathogens10030255