
Murine Models of Obesity
by
 , , , and
, , , and
Tânia Martins
1,2,3,†, 
Catarina Castro-Ribeiro
1,2,†,
Sílvia Lemos
1,2,
Tiago Ferreira
1,2,
Elisabete Nascimento-Gonçalves
1,2,
Eduardo Rosa
1,2,
Paula Alexandra Oliveira
1,2,3 and
Luís Miguel Antunes
1,2,3,* 1
Centre for the Research and Technology of Agro-Environmental and Biological Sciences (CITAB), University of Trás-os-Montes and Alto Douro (UTAD), 5000-801 Vila Real, Portugal
2
Inov4Agro—Institute for Innovation, Capacity Building and Sustainability of Agri-Food Production, University of Trás-os-Montes and Alto Douro (UTAD), 5000-801 Vila Real, Portugal
3
Department of Veterinary Sciences, School of Agrarian and Veterinary Sciences, University of Trás-os-Montes and Alto Douro (UTAD), 5000-801 Vila Real, Portugal
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Obesities 2022, 2(2), 127-147; https://doi.org/10.3390/obesities2020012
Submission received: 8 February 2022
/
Revised: 28 March 2022
/
Accepted: 29 March 2022
/
Published: 31 March 2022
Abstract
:Obesity, classified as an epidemic by the WHO, is a disease that continues to grow worldwide. Obesity results from abnormal or excessive accumulation of fat and usually leads to the development of other associated diseases, such as type 2 diabetes, hypertension, cancer, cardiovascular diseases, among others. In vitro and in vivo models have been crucial for studying the underlying mechanisms of obesity, discovering new therapeutic targets, and developing and validating new pharmacological therapies against obesity. Preclinical animal models of obesity comprise a variety of species: invertebrates, fishes, and mammals. However, small rodents are the most widely used due to their cost-effectiveness, physiology, and easy genetic manipulation. The induction of obesity in rats or mice can be achieved by the occurrence of spontaneous single-gene mutations or polygenic mutations, by genetic modifications, by surgical or chemical induction, and by ingestion of hypercaloric diets. In this review, we describe some of the most commonly used murine models in obesity research.
1. Introduction
According to the World Health Organization (WHO), in 2016, nearly 2 billion adults were overweight, of which more than 650 million were obese [1]. Obesity is common in both developed and developing countries, affecting all ages without distinguishing social classes [2,3]. Over the past 50 years, overweight and obesity have risen sharply. In 2016, the United Stated of America and Saudi Arabia registered the highest percentage of obese adults, 36.2% and 35.4%, respectively. Conversely, Vietnam, Japan, Ethiopia, and India had the lowest percentages of obese adults in 2016 [1]. Overweight and obesity are defined as abnormal or excessive fat accumulation typically resulting in negative health impacts. Although there is a genetic predisposition to developing obesity, the causes of this disease are multifactorial. Thus, the complex interactions between epigenetics, lifestyle, cultural and environmental influences play a fundamental role in the development of obesity (Figure 1) [3,4,5]. Additionally, economic growth, social changes, and the global nutritional transition are seen as contributing to this epidemic [6]. Obesity should not be seen only as a problem associated with physical image since people with excessive weight tend to have a higher risk of developing other comorbidities, such as type 2 diabetes, hypertension, and certain types of cancer, namely breast, ovary, liver, colon, and prostate [2]. Obesity is also a risk factor for cardiovascular diseases, chronic respiratory diseases, and osteoarthritis, which contribute to more than half of all deaths. More serious cases of obesity are associated with an increasing incidence of diseases such as asthma, gallstones, steatohepatitis, glomerulosclerosis, dyslipidemia, and endothelial dysfunction [7]. The list of health consequences is extensive, and in addition to affecting quality of life, it also contributes to a higher risk of premature death [8]. In the last two decades, the need to develop new therapies increased significantly. The physiological treatment forces the patient to make changes in diet and behavior, which is not always feasible due to the patient’s lack of motivation. Thus, the pharmacological approach is more attractive. Currently, pharmacological therapy includes the use of synthetic drugs [9], although the demand for natural substances that have fewer side effects is increasing. To determine the effectiveness of new drugs or promising natural compounds, these potential therapies must be first evaluated in preclinical models (in vitro and in vivo) and later clinically [10]. Cell cultures are important for obtaining information on the pharmacodynamics of compounds; however, more complex interactions, which are those that contribute to the pathophysiology of obesity, are not noticeable. Therefore, preclinical models, specifically animal models, are essential for identifying and validating pharmacological targets [11]. Animal models for studying obesity comprise a variety of species, from non-mammals (zebrafish, Caenorhabditis elegans and Drosophila) to mammals (rodents, large animals, and non-human primates) [7]. Within these animal models, small rodents (rats and mice) are the most widely used. Their cost-effectiveness and multiparity, physiology close to humans, and easy genetic manipulation are some of the advantages that make them the model of excellence [7]. In this review, we describe some of the most commonly used rodent models of research on obesity.
2. Rodent Models of Obesity
The use of animals for scientific purposes is a long-standing practice, dating back to ancient Greece [11,12]. Both humans and other mammals are highly complex organisms, where the organs perform different physiological functions in a regulated way. The anatomical and physiological similarities between human and animal, particularly mammal, led to the use of animals before applying new compounds on humans. To select an animal model, certain parameters must be met: pathophysiological similarities with the human disease; phenotypic correspondence with disease status; simplicity; replicability, reproducibility, and cost-efficiency [11]. Rats and mice are the most widely used preclinical animal models to study obesity [13,14,15]. These animal models help us to understand the biopathology and develop possible preventions and treatments against obesity (Figure 2) [14,16]. Nevertheless, each model presents its advantages and disadvantages (Table 1).
2.1. Monogenic Models
Monogenic animal models are animals with a single gene disorder. They are widely used to study obesity, very reliable, and effective. These models present a well-organized molecular map. However, they differ from humans in the way that they undertake their energy partitioning and fat deposition; and are generally not a good representative of human diseases [17]. Although it has been proven that a single manipulation of almost 250 genes can induce obesity in mice, monogenic models are mainly a result from mutation(s) in the leptin pathway [5,17]. Genetic manipulations or environmental/dietary factors will dictate the severity of obesity and comorbid metabolic modifications [5].
2.1.1. ob/ob Mice
The obese mice (ob/ob mice) were discovered in 1949 by investigators from the Jackson Laboratory [6]. However, only in 1994 was the genetic characterization of this mutation as a single base pair deletion performed and the gene product was called leptin [6,14,18]. This mutation prevents the secretion of bioactive leptin by positional cloning, causing its synthesis to be terminated prematurely [6,7]. Leptin is an adipocyte-derived hormone, the major function of which is related to food intake inhibition, metabolism, and reproduction, through interaction with the central nervous system mainly relating to the hypothalamus. The normal function of the leptin pathway is required for body weight and energy homeostasis [19,20,21]. Mutations in the leptin gene lead to animals with an obese phenotype [14].
The ob/ob mice are widely used because they display early-onset obesity, which is the result of hyperphagia and low energy expense [7,22]. This obesity model possesses the capacity to gain weight in a short period of time, eventually reaching three times the normal weight of wild-type controls [6,22]. These mice exhibit impaired glucose tolerance and insulin sensitivity, most likely as a secondary effect of obesity. Furthermore, they have high levels of circulating corticosterone, develop hypothyroidism, dyslipidemia, decreased body temperature, defective thermogenesis, and are infertile due to hypogonadism [23]. The ob/ob mice have been used to evaluate the influence of several natural compounds on the obesity development [24,25,26,27,28] as well as other concomitant diseases such as metabolic syndromes [29] and oxidative stress [30,31].
2.1.2. db/db Mice
The db/db mouse (db stands for diabetes) [6] is another option for studying the molecular basis of obesity but is commonly used for the investigation on type 2 diabetes [17]. These db/db mice were also developed by investigators from the Jackson Laboratories in 1966 and are phenotypically similar to the ob/ob mice model. They exhibit a mutation in the leptin receptor gene, in an autosomal recessive trait that encodes for a G-to-T point mutation, leading to defective leptin signaling [7,17]. The db/db mice are characterized by hyperphagia, a consequence of impaired leptin signing in the hypothalamus, develop early-onset obesity due to low energy expenditure, are insulin resistant, have decreased insulin levels, and are hypothermic. They also have slow growth due to growth hormone deficiency and are infertile [6,7,14,17,32]. Comparatively to the ob/ob mouse, the db/db model has a marked resistance to leptin, which is a consequence of a mutated leptin receptor, but its leptin levels are markedly elevated [14]. The db/db model can be used in the comprehensive study of diabetes and to evaluate the effect of natural compounds on disease development or as chemopreventive agents [33,34,35,36].
2.1.3. s/s Mice
The s/s mice were developed by Bates and collaborators (2003) to study the role of individual leptin signals, by disturbing the STAT3 pathway, even though it is regularly expressed on the cell surface and can mediate other leptin signals. In this model, the long-form signaling pathways by which leptin exerts its functions are compromised [17,18,37]. The s/s mice are hyperphagic and obese, present normal body length, and contrarily to the ob/ob or the db/db models, they are fertile [14]. The db/db mice and s/s mice are often compared, and share some characteristics such as early development of obesity, hyperphagia, and high leptin/insulin levels [23]. The improvement verified in s/s mice regarding glucose homeostasis is due to decreased insulin resistance and glucose intolerance. These facts are usually attributed to leptin mediation by STAT3, via melanocortin signaling, which affects body energy homeostasis. However, fertility, body growth, and glucose homeostasis are controlled through non-STAT3 signaling [14,38]. The transgenic s/s mouse has played its contribution to the research on obesity and diabetes [18].
2.1.4. B6 (cg)-Tubtub/J
The autosomal recessive mutation called tubby (tub) arose spontaneously in a C57BL/6J colony [39]. The expression of tub in the paraventricular, ventromedial, and arcuate nuclei of the hypothalamus suggests a role in body weight regulation or feeding behavior [40]. These homozygous mice develop obesity at the beginning of maturity. The increase in body weight in these animals is due to the greater accumulation of adipose tissue: males show weight gain in 3–4 months and females later, in 4–6 months. Blood glucose levels are considered normal, in contrast to plasma insulin levels, which are increased even before the obvious signs of obesity. In addition to these, the levels of total cholesterol, triglycerides, and high-density lipoprotein cholesterol are also increased in the plasma. However, these homozygous mutant mice do not have atherosclerotic lesions in the blood vessels and fertility is affected [41].
2.1.5. Zucker Fatty Rat
The Zucker fatty rat (ZFR), also known as Zucker (fa/fa), is the result of breeding Sherman and Merck stock M (13M strain) rat strains and were developed in 1961 by L. M. Zucker and T. F. Zucker [42,43]. The ZFR rats hold a homozygous missense mutation (fatty, fa) in the long form of the leptin receptor, being fairly insensitive to leptin and present a similar phenotype to the ob/ob and db/db mice [23,32]. This mutation is correlated with demarked defects in the leptin receptor. Therefore, there is a low number of leptin receptors on the cell surface, leading to diminished leptin binding and signal transduction [14]. This model is characterized by early-onset obesity, at 5 weeks of age animals are obese [6,42] and by the 14th week of age 40% of body composition is lipids [42] due to the development of hyperphagia [14]. These rats also develop hyperglycemia, hypercholesterolemia, hyperinsulinemia, and adipocyte hypertrophy and hyperplasia [42,44]. These animals develop obesity, without diabetes, but a strain derived from the ZFR, called Zucker diabetic fatty fa/fa (ZDF) was developed, and is widely used for the study of type 2 diabetes [32]. The ZFR is the most used model for the study of genetic obesity traits [42,44,45,46].
2.1.6. Otsuka Long-Evans Tokushima Fatty Rat
Otsuka Long-Evans Tokushima fatty (OLETF) rat was the result of selective breeding at the Tokushima Research Institute in 1982 [44,47]. OLETF rats have been used as a model for studying diabetes and obesity [48,49,50,51]. They are hyperphagic and develop mild obesity [44], which is a consequence of increased food intake and meal size, leading to increase in the bodyweight [49]. These animals also develop polyuria, polydipsia, hypertension, and dyslipidemia [49]. OLETF rats have a deficiency in the cholecystokinin (CCK) receptor type A, which is a contributing factor to their phenotype [52]. CCK is a derived peptide hormone which works as a peripheral satiation signal [7]. OLETF rats are a valuable tool for studying the dysregulated control of eating since CCK interferes with satiation [14].
2.2. Polygenic Models
For the study of obesity, polygenic models provide more accurate information about the biology of obesity, when compared to monogenic models, since human obesity is mediated by multiple genes [6]. These models are some of the most used, are cost-effective, used regularly, and are considered more realistic models for studying human obesity. Among the different types of polygenic models, only the most frequently used models are included in this review [17].
2.2.1. New Zealand Obese Mice
The New Zealand obese mice (NZO) strain is a polygenic model that develops obesity and type 2 diabetes only in males [6,7,53]. This mouse strain was obtained in the Hugh Adam Department of Cancer from the selective inbreeding of a stock colony [54]. Some gene variants involved in obesity and diabetes in NZO mice are the Tbc1d1 gene inactivation, causing enhanced transport and oxidation of fatty acids which in turn reduces glucose transport, and the transcription factor Zfp69, which is associated with altered triglyceride distribution and enhanced diabetes susceptibility [55,56]. Leptin receptor (Lepr), phosphatidyl choline transfer protein (Pctp), ATP-binding cassette transporter G1 (Abcg1), and neuromedin U receptor 2 (Nmur2) are other candidates potentially contributing to the polygenic obesity and diabetes of NZO mice [55,56]. These animals show the highest levels of adiposity among the polygenic models. They also share other characteristics that are similar to human type 2 diabetes, such as insulin resistance in brown adipose tissue and skeletal muscle at the age of 4–5 weeks old, that progress into diabetes. They show hypercholesterolemia and hypertension [7,57]. They display moderate hyperphagia and reduced energy expenditure and voluntary activity [7], especially when compared to control or even ob/ob mice [6,17,57]. This strain is similar to the ob/ob mice, except for its ability to maintain body temperature during a 20 h period of cold temperatures, whereas ob/ob mice would not be able to surpass a temperature of 4 °C [57]. NZO mice fed a high-fat diet develop liver steatosis, which is associated with a loss of autophagy efficiency that leads to the disturbance of proteostasis [58].
2.2.2. Tsumura and Suzuki Obesity and Diabetes Mice
The Tsumura and Suzuki obesity and diabetes (TSOD) strain was established in 1992 by Tsumura and Suzuki through selective inbreeding of obese male ddY mice [54,59,60]. Male TSOD mice are characterized by polygenetic obesity, insulin resistance, hyperinsulinemia, hyperglycemia, polydipsia, and polyuria [54,61]. The genome-wide screening of the TSOD strain revealed three quantitative trait loci that are intimately linked with glucose homeostasis and obesity in chromosomes 11, 1, and 2. This strain shares similar symptoms to diabetes mellitus symptoms in humans [54,61]. TSOD mice, being a spontaneous model for metabolic syndrome, are highly utilized for that purpose [62,63,64], as well as for a variety of studies regarding diabetes and/or obesity [65,66,67,68,69,70]. TSOD also present gut dysbiosis, which is implicated in the pathogenesis of metabolic syndrome, and consequent dysregulation of plasma levels of short-chain fatty acids (SCFAs), the main fermentation products of gut microbiota [71].
2.2.3. Kuo Kondo-Ay Mice
The Kuo Kondo (KK) mouse is a polygenic model of moderate obesity and type 2 diabetes mellitus developed in 1975 in Japan [54]. The KK-Ay mouse is a substrain derived from the KK mouse strain, which was the result of the transference of the lethal yellow obese gene (Ay). KK-Ay mice are severely obese and characterized by distinct adiposity, display hyperphagia, hyperglycemia, hyperinsulinemia, and glucose intolerance by the age of eight weeks [54,72]. This model is used to study obesity and the effect of anti-diabetic drugs for type 2 diabetes [6,72,73].
2.2.4. M16 Mice
The M16 mouse is a polygenic model of early-onset obesity and moderate hyperglycemia [59]. This model originates from a long-term selection of ICR outbred mice, with the goal of rapid weight gain between the third and sixth weeks of age [6,59]. M16 mice exhibit a rapid development of body fat percentage and body size [59] maintaining at all times a larger body weight than the ICR strain. This mouse model also develops hyperphagia, hyperinsulinemia, and hyperleptinemia [6,59,74]. These M16 mice closely simulate obesity and diabetes in young adults [72], being used as a model for early-onset polygenic obesity and diabetes [74].
2.3. Genetically Modified Mice
Genetically modified mice are widely used in research to study genetic influence, modulate a disease, or analyze a biological process in vivo. Mice are considered to be the most suitable mammals for this purpose, as they share the same organ and tissue systems as humans [75]. Genetically models of obesity include transgenic and knockout mice.
2.3.1. Transgenic Mice
Transgenic models of obesity were created to promote access to animals with genetic characteristics identical to those described in obese humans [17].
Corticotropin-Releasing Factor Overexpressing Mice
The corticotropin-releasing factor (CRF) is a peptide that acts as a neuroendocrine hormone and neurotransmitter through the hypothalamic–pituitary–adrenal axis and hypothalamic and extrahypothalamic neuronal pathways [76]. CRF induces a rapid decrease in food intake and gastric emptying that leads to stress-related alterations in food intake and the gastric motor function. Hormones related to the pattern of eating, such as the neuropeptide Y and leptin, are also regulated by CRF [76]. The CRF-overexpressing mice exhibit truncal obesity, muscle wasting, thin skin and hair loss, and are commonly used for study conditions associated with hyperadrenocorticism such as Cushing’s syndrome [14]. This mouse model is often used for studies regarding the response to different feeding conditions [76,77] and studies related to chronic stress [78,79].
Melanin-Concentrating Hormone Overexpressing (MCH-OE) Mice
The melanin-concentrating hormone (MCH) is a hypothalamic peptide known for its role in the regulation of energy balance and feeding behavior [80,81]. MCH is also modulated in response to leptin and insulin [82]. The MCH-OE mice become obese, and display hyperphagia, hyperinsulinemia, and are insulin resistant when exposed to a high-fat diet (HFD) [14,83]. MCH-OE mice are commonly used to study the effects of different feeding behaviors on obesity and insulin resistance [80,82,84].
Overexpression of 11β-Hydroxysteroid Dehydrogenase Type 1 (11β HSD1) Mice
11β-hydroxysteroid dehydrogenase type 1 (11β HSD1) is a metabolic reductase and dehydrogenase that is responsible for converting inactive cortisone to active cortisol, leading to an increase in glucocorticoid function. Selective inhibition of 11β HSD1 leads to a decrease in fasting blood glucose, insulin, cholesterol, free fatty acids, and triglycerides [85]. Overexpression of 11β HSD1 in mice leads to the development of obesity, insulin resistance, hyperphagia, high leptin levels [86], and most of the features of metabolic syndrome in mice, especially when these animals eat a HFD [14,85]. This model is an interesting option for studying central obesity [86,87].
Overexpression of Glucose Transporter Subtype 4
The glucose transporter subtype 4 (GLUT4) is an insulin-responsive facilitative glucose transporter that is expressed in adipose, skeletal, muscle, and cardiac muscle cells [83,88,89]. The enhanced expression of GLUT4 in adipocytes suggests that its expression is probably implicated in the pathophysiology of obesity [17]. Transgenic mice with an overexpression of GLUT4 in white and brown fat were first generated by Shepherd and collaborators in 1993 [83,90]. The GLUT4-overexpressing mice are known to develop early onset obesity due to an increase in glucose transportation, stimulated by an improvement in the whole-body insulin action, with the consequence of an augmented nutrient substrate for adipogenesis. Due to the large number of cells, this mouse model is used to study fat cell replication and differentiation during obesity [14].
UCP-DTA Mice
Brown adipose tissue (BAT) is specialized in the dispersion of energy through the production of heat and is different from white adipose tissue due to the presence of uncoupling protein 1 (UCP-1) [91]. To understand the role that BAT plays in balancing systemic energy, Lowell and colleagues [92] created transgenic mice (UCP diphtheria toxin (DTA) A-chain mice) without a BAT function. This was accomplished by using the BAT-specific UCP promoter to drive the expression of a gene for DTA [93]. These transgenic mice (UCP-DTA) are markedly obese, have decreased energy expenditure, and showed an ablation of approximately 70% of the BAT. With obesity development, as body weight increases, animals may develop insulin resistance (possibly developing diabetes mellitus type 2), hyperphagia, and dyslipidemia [14,83,94]. When fed with a high-fat diet, UCP-DTA mice show greater body mass, lipid mass, intramuscular triglyceride content, and increased plasma insulin levels beginning early in the experiment [95].
2.3.2. Knockout Mice
Knockout mice are a valuable tool used to explore potential strategies for the treatment of specific diseases, that includes pharmacological and genetic therapy approaches. Another major purpose of these mice models is to understand the function and significance of specific genes. They are an excellent tool for producing a model of human disease and to evaluate the physiological functions of specific genes [96].
Beta-3 Adrenergic Receptor Knockout Mice
In rodents, the β-3 adrenergic receptors (β3-ARs) are largely expressed in the white and brown adipose tissue [83,97,98]. β3-ARs are linked to the control of thermogenesis in brown adipocytes and stimulation of lipolysis in white adipocytes, through activation of the sympathetic nervous system [97]. The β3-AR knockout mice are moderately obese, a phenotype that is more common in females. Since food intake is similar to the control groups, changes are considered to be due to a decrease in activity of the sympathetic nervous system [14,83]. This mouse model can be helpful for the study of selectivity and function of β3-AR ligands and may present as an important model to distinguish physiology mediated by the sympathetic nervous system [83].
Bombesin Receptor Subtype 3 Knockout Mice
The bombesin-like peptides bind to G protein coupled receptors, such as gastrin releasing peptide receptor (GRP-R), neuromedin B receptor (NMB-R), and bombesin receptor subtype 3 (BRS-3). Bombesin-like peptides are found throughout the entire central nervous system and gastrointestinal tract and its main functions are the modulation of smooth-muscle contraction, exocrine and endocrine processes, metabolism, and behavior [83]. BRS-3 in both melanocortin receptor 4 (MC4R)- and single-minded homolog 1 (SIM1)-expressing neurons contributes to the regulation of food intake, body weight, adiposity, body temperature, and insulin sensitivity [99]. The BRS-3 knockout leads to mild obesity, hypertension, impaired glucose metabolism, reduced metabolic rates, increased feeding efficiency, and hyperphagia [100,101]. This mouse model is a great option for the study of moderate obesity, related with type-2 diabetes [83,101].
Deletion of the Neuronal Insulin Receptor (NIRKO) Mice
Deletion of the neuronal insulin receptor in mice (NIRKO mice) leads to the development of an animal model of diet-induced obesity and insulin resistance, due to increased food intake [83,100]. They become mildly obese with hypertriglyceridemia, and an increase in plasma insulin and leptin levels is detected [83]. They exhibit hyperphagia, have reduced fertility, decreased counterregulatory response to hypoglycemia [102], and reduced phosphorylation of Akt and GSK3 in the brain [103]. This mouse strain model could be an option for the study of the relationship between energy balance regulation and reproductive system control, as well as for the study of insulin-related and insulin-independent feedback loops for postprandial inhibition of food intake [83].
Disruption of the Neuropeptide-Y 1 Receptor (NPY1R) Mice
Neuropeptide Y (NPY) is a neurotransmitter that is highly studied because it is correlated with the modulation of feeding behavior, control of autonomic and endocrine actions, is associated with energy storage and regulation of the cardiovascular and sympathetic nervous systems [100]. Pharmacological studies demonstrated that the NPY 1 receptor (NPY1R) is correlated with food intake stimulation and positive energy balance. NPY1R knockout mice Y1 show a striking diminished feeding response to fasting and decrease in food intake. These animals are moderately obese and display hyperinsulinemia, without hyperphagia. Obesity is more prominent in females than in males and occurs mostly not because of hyperphagia but due to impaired control of insulin secretion or low energy expenditure because of the reduced expression of uncoupling protein type 2 in white adipose tissue. This mouse model is ideal for studying moderate obesity, which is a consequence of a diminished energy expenditure without showing hyperphagia, and to study NPYR-1 agonists [83]. Conditional knockout of the Npy1r gene in the excitatory neurons of the forebrain of adolescent male mice decreases body weight growth and adipose tissue [104]. Conversely, when mice with conditional knockout of the Npy1r gene in limbic areas are fed a high-fat diet, they show an increase in body weight, visceral adipose tissue, blood glucose levels, hyperphagia, and dysregulation of calory intake [105].
Knockout of the Serotonin 5-HT-2C Receptor Gene
In the current therapy, targeting of the 5-hydroxytryptamine (5-HT; serotonin) has shown to be one of the most promising options for the treatment of obesity [106]. 5-HT is a monoaminergic transmitter related to the modulation of sensory, motor, and behavioral processes in the mammalian nervous system through the activation of a large family of receptor subtypes [83]. The selective activation of 5-HT-2CR of the brainstem nucleus of the solitary tract decreases feeding behavior and food intake. Additionally, the precursor polypeptide pro-opiomelanocortin (POMC) of the nucleus of the solitary tract and arcuate nucleus are required for 5-HT2CR agonist lorcaserin to reduce feeding [107]. The serotonin 5-HT-2CR knockout mice were created by Tecott and collaborators (1995) [83,108]. They display hyperphagia since weaning and a normal weight at young ages. Late-onset obesity is a result of increased white adipose tissue, due to abnormal control of feeding behavior. Changes in insulin or glucose levels were non-existent until mice became obese [14,83,100]. In addition to being used to study obesity [107,109], this mouse model is also used in studies related to changes in the locomotor activity, anxiety, and fear memory [110,111].
Neuropeptide Y Receptor Y2 (NPY2R) Knockout Mice
Mice that lack Neuropeptide Y Receptor Y2 (NPY2R), a protein coding gene, are mildly hyperphagic and obese, and show a perturbed regulation of heart rate [83]. The NPY2R signaling in the aguti-related protein (AGRP) neurons, in the arcuate nucleus of the hypothalamus, controls energy expenditure and food foraging behavior [112]. When NPY2R knockout mice are fed a normal chow diet, no changes in serum concentrations of triglyceride, cholesterol, or glucose were observed [14]. The NPY2R knockout mice are ideal for studying selectivity of NPYR-Y2 agonists [83].
2.4. Chemical or Surgical Models of Obesity
The hypothalamus, despite its numerous functions, plays a fundamental role in the transmission of afferent signals from the intestine and brain stem, as well as in the processing of efferent signals that modulate food intake and energy expenditure. It is subdivided into nuclei connected to each other, of which the arcuate nucleus (ARC), paraventricular nucleus (PVN), ventromedial nucleus (VMN), dorsomedial nucleus (DMN), and lateral hypothalamic area (LHA) stand out [113]. The lesions produced in these nuclei can be obtained mechanically, through surgery, radiofrequency or electrolytically, or chemically using neuronal toxins such as monosodium glutamate, gold thioglucose, kainic acid, ibotenic acid, and bipiperidyl mustard [83]. Medial hypothalamic lesions (ARC, VMN, and DMN) are associated with strong hyperphagia and activation of the TLR4/NF-κB-pathway, as well as reduced expression of oxytocin in the hypothalamus [114]. Surgical removal of the ovaries is also used to study the incidence of obesity in women.
2.4.1. Lesions of the Ventromedial Hypothalamus
The ventromedial hypothalamus (VMH) nucleus, located in the hypothalamus, is a very complex and important nucleus, which receives several inputs that project to the brain stem and spinal cord, and is associated with the regulation of eating behavior and satiety. One of the first models developed to induce obesity in rodents used rats with lesions in the ventromedial region of the mediobasal hypothalamus (VMH lesions) [7,14,29]. VMH lesion further exaggerates insulin resistance and dyslipidemia in rats fed a sucrose diet, with males being more prone to this exacerbation [115]. Bilateral lesions of the VMH could be made using electrical current or the neurotoxin monosodium glutamate (MSG), leading to the development of hyperphagia, vagal hyperactivity, sympathetic hypoactivity, enlarged pancreatic islets, and hyperinsulinemia [10,116]. Obesity can develop in these animals even when food intake is restricted, although the mechanism by which hypothalamic injury induces obesity is still unknown [14,83]. Nevertheless, the nuclear receptor steroidogenic factor 1 (SF-1), exclusively expressed in the VMH region of the brain, seems to be a key component in the VMH-mediated regulation of energy homeostasis, playing a protective role against metabolic stressors including aging and high-fat diets [117].
Monosodium Glutamate (MSG)
MSG is a neurotoxin that can be administered subcutaneously or i.p. (2–4 mg/g of body weight) during the neonatal period, for 4–10 times to obtain obesity. It is a substance that can be found in many foods and therefore the effects of this substance when taken orally have been studied [10,118]. The administration of MSG in rats or mice will induce vagal hyperactivity and sympathetic-adrenal hypoactivity, which will result in obesity, due to the lack of control between nutrient absorption and energy expenditure. The administration of this neurotoxin induces changes related to the lack of control of the hypothalamic–pituitary axis with dose-dependent curve, including hypophagia, development of obesity, hypoactivity, late puberty, reduced ovaries weight, and elevated serum levels of corticosteroids [118]. In addition, the levels of glucose, triglycerides, and leptin are also increased, resulting in an increase in white adipose tissue deposition and insulin resistance. Adult rats who received MSG in the neonatal period developed endocrine dysfunction syndromes which is characterized by obesity development, disturbances in the regulation of caloric balance, reduced growth, behavioral changes, and hypogonadism [10,119].
Electrical VMH Lesion
Described by Saito et al. (1985), and subject to some changes, this procedure uses an electrical current from stereotaxic instruments in anesthetized adult rodents [120]. Contrary to what was discovered in the MSG injury, the use of electric current causes obesity due to hyperphagia. More recent studies report weight gain in animals subject to this procedure due to the involvement and interrelationships of leptin, insulin, and NPY in the hypothalamus. In animals with hypothalamic lesions via electrical current, leptin levels increase, NPY decreases, and circadian rhythm fluctuations are maintained. The loss of the feedback mechanism between insulin and leptin results in fat accumulation [10,29,120].
2.4.2. Lesions of the Hypothalamic Paraventricular Nucleus
Lesions of the hypothalamic paraventricular nucleus (PVN), similar to those of VMH, induce obesity, but in this model energy expenditure and activity are not affected, showing that the mechanisms of obesity induction in animals with PVN lesions differ from those with lesions made in VMH. The induction of obesity in these animals is dependent on food intake increase. However, in animals with lesions in the VMH, obesity is independent of food intake increase, as it results in metabolic obesity. Rats with PVN lesions, in addition to being hyperphagic and obese, can develop insulin resistance and hyperinsulinemia [14,83,121,122,123]. Brain-derived neurotrophic factor (BDNF)-positive neurons in the PVN can regulate energy intake and adaptive thermogenesis in interscapular brown adipose tissue [124]. Additionally, it is suggested that the substantia nigra (SN)-PVN pathway may be an upstream neural pathway that can affect BDNF expression in the PVN, and that its regulatory effects may be mediated by dopamine receptors [125].
2.4.3. Lesions of the Arcuate Nucleus
It is possible to induce obesity by damaging the arcuate nucleus (ARC). However, this procedure is difficult to perform due to its location and anatomical shape. Several researchers, in order to make the destruction of neurons more selective, began to administer MSG to rats in the first 10 postnatal days, and later they performed the surgical procedure [10,126]. However, lesions induced by MSG are not restricted to ARC, as they also cause damage to circumventricular neurons, due to their open blood–brain barrier. These animals become hyperphagic, develop obesity, and consequently hyperinsulinemia [83]. Local administration of gold thioglucose also allows ARC neuron destruction, and the induced effects on animals are similar to those described above [10,14].
2.4.4. Ovariectomy
To understand the increased incidence of obesity in women, which occurs during menopause, female rats and mice are subjected to ovariectomy (OVX). Due to the lack of estradiol, these animals show an increase in food consumption both in estrus and in the other days of the cycle. This cyclic effect is hypothetically caused by the lack of estradiol in meal controllers, such as cholecystokinin (CCK), glucagon-like peptide-1 (GLP-1), glucagon, and others [14]. In OVX animals, body and total adipose weights increase, and adipose tissue accumulates as subcutaneous fat [127]. Animal studies have revealed a consistent correlation between bilateral oophorectomy and insulin resistance, total and LDL cholesterol levels, and adiposity [10,14,17]. OVX mice also have differential expression of genes in the hypothalamus involved in thermoregulation, food intake, glucose and lipid metabolism [128]. In addition, the hypothalamic mTOR signaling pathway seems to play an important role in regulating weight gain and adiposity in OVX mice [129].
2.5. Diet-Induced Obesity (DIO)
Animal models of diet-induced obesity (DIO) are useful for studying the polygenic causes of obesity since they allow to reproduce the most common cause of this disease in man: dietary imbalances [7,10,16,41,130]. Animals, usually laboratory rodents (rats and mice), are exposed to specifically tailored diets that induce obesity. These diets should be prepared by companies that are aware of the nutritional specificities of laboratory animals, modifying their formulation in accordance with the objectives of each researcher [11]. Additionally, different diets could be used such as a high-fat diet (HFD), high-sugar diet (HSD), and the combination of the last two, called Western diets [15,130]. The diet, especially in Western countries, is rich in saturated fat and carbohydrates such as fructose and sucrose, and when given to animals it allows them to mimic the signs and symptoms that are characteristic of human metabolic syndrome [131]. Nevertheless, the poor standardization of the diets and feeding protocols usually results in inconsistent and irreproducible data, leading to differences in experimental findings between similar studies [13].
2.5.1. High-Fat Diet (HFD)/Exposure to High-Fat and Palatable Diets
The obese mouse model, induced by a HFD, is one of the most important tools for understanding the relationship between hyperlipidemic diets and the pathophysiology of obesity development [41]. There are several strains of mice susceptible to DIO, although there are also animals that appear to be resistant to increasing body weight when exposed to HFDs, termed ‘diet resistant (DR)’ [16,131]. The mouse C57BL/6 strain is highly susceptible in contrast to the mouse strains SWR/J, A/J and CAST/Ei which tend to be resistant to these diets. The C57BL/6J mouse, an inbred strain, is the most commonly used due to the similarities that are observed between them and humans in relation to the development of the metabolic syndrome when fed a HFD, resulting in an obese phenotype. After eating a HFD these animals develop obesity, high adiposity accumulation, insulin resistance, hyperglycemia, hyperlipidemia, and hypertension. They are also susceptible to non-alcoholic fatty liver disease and endothelium damage, commonly associated with cardiovascular diseases. All the characteristics described above are described in humans with complex metabolic syndrome [6,7,41,132,133].
In addition to mice, some rat strains can also be used for HFD-induced obesity models, such as outbred Sprague Dawley, Wistar, or Long-Evans rats [14]. Weight gain induced by diet changes causes defects in the neuronal response to negative feedback signals from circulating adiposity, such as insulin. Insulin resistance involves cellular inflammatory responses caused by excess lipids. This model of diet-induced obesity (DIO with HFD diets) has become one of the most important tools for understanding the interaction of these diets high in saturated fat and the development of this disease [41]. Outbred Sprague-Dawley rats showed diverse responses, where some animals show DIO while others show DR responses [16].
2.5.2. High-Carbohydrate Diet (HCD)
A high-carbohydrate diet combined with animal or vegetable fat can mimic the human diet. This diet, in rodents, induces metabolic syndrome. The fat source can vary, and fructose and sucrose are the most common carbohydrates present in these diets, also varying their amounts [131]. When fed diets high in fat and sucrose, rodents tend to increase their body weight and deposition of abdominal fat, as well as develop hyperglycemia, hyperinsulinemia, and hyperleptinemia [131,134]. In addition, consumption of these diets can also cause fatty liver and increase liver lipogenic enzymes [135,136]. Fructose ingestion in animals induces an increase in body weight and deposition of abdominal fat, increases plasma levels of triglycerides, cholesterol, free fatty acids and leptin, induces insulin resistance, hyperglycemia, and is associated with fatty liver and inflammation [137]. These animals also show cardiac hypertrophy, ventricular stiffness and dilation, hypertension, cardiac inflammation and fibrosis, decreased cardiac function and endothelial dysfunction together with mild renal damage and increased pancreatic islet mass [131]. Hypercaloric diets induce hyperglycemia and consequently induce glucose tolerance and increase plasma levels of TAG, TNF-α, and MCP-1/JE, as well as MCP-1/JE levels in organs such as the liver and adipose tissue [127]. Animals fed with a HCD show, however, a lower body weight gain when compared to animals fed with a HFD [41].
2.5.3. Cafeteria Diet (CAF)
The cafeteria diet (CAF) is a varied and tasty diet [14]. Here, the animal selects its own food, which is very tasty and easily accessible. It is mainly composed of foods with a high content of sodium, fat, and sugar (cookies, sweets, cheeses, and sausages). A major disadvantage is the lack of defined composition and the fact that animals can make different food choices, making it an inaccurate and unreliable diet for obesity studies [138]. In CAF-fed rats, their body weight increases dramatically and remains significantly elevated, making these animals obese. Hyperphagia, hyperinsulinemia, glucose intolerance, and hyperglycemia are exaggeratedly high when compared to other models of HFD. This model shows increased adiposity and hepatosteatosis, white fat, and more inflammation in the adipose tissue and liver [14,41]. A chronic CAF on a restricted schedule has also been shown to increase BDNF levels and its receptor TrkB in the prefrontal cortex of rats [139].
2.5.4. Maternal Overfeeding and Exposure to High-Fat Diets
When feeding pregnant rats or mice with a HFD, effects will appear on their progeny, where, for example, the offspring will be heavier and more obese when compared to the offspring of DR animals. Moreover, the obesity in the offspring of DIO pregnant females is extended up to adulthood. The same is true in humans, where maternal feeding with a HFD leads to offspring at increased risk of obesity in adulthood. In addition, they develop hyperphagia and a preference for sweet and fatty foods. Maternal obesity is associated with an increase in obesity in genetically predisposed individuals, intensified by the consumption of diets [14]. Maternal CAF diet has also been shown to affect fat content, metabolic profiles, and inflammation parameters in offspring, with female offspring being more susceptible to the diet [140].
2.5.5. Other Models
Other models are the sand rat (Psammomys obesus) and the Acomys species. The Psammomys obesus, also known as the Israeli sand rat, is an herbivorous and diurnal gerbil from the North African and Middle East Regions [141,142]. The poor adaptation of this model to a new nutritional diet and changes in exercise rate makes it a good example for research on early-onset obesity, followed by diabetes [7,143]. This model is also used for studying cardiovascular diseases [142,144], among other subjects. However, these animals are not available for purchase. The Acomys species (called “spiny” mice) inhabit the arid areas of eastern Mediterranean and North African countries. The use of spiny mice as a model for the study of obesity and diabetes was a random discovery. Scientists discovered that they become overweight when fed bird food, rich in pumpkin, sesame, and sunflower seeds. The Acomys species are highly utilized for their tissue-regenerative properties [145,146,147,148]. These animals are also not available for purchase.
2.5.6. Animal Models and Clinical Setting
Translational medicine drives progress of research along the continuum from basic biomedical research findings into clinical practice [149]. The diversity of obesity models has allowed the identification of specific genes and their role in the regulation of energy balance, the understanding of the central control, and the detailed molecular and neural energy-balance mechanisms [17,150]. Furthermore, these animal models allowed to study the combination of genetics and dietary influences in the development of obesity [58,151]. In humans, obesity is also a multifactorial disease with genetic and environmental influences, presenting different etiologies, with diverse clinical signs [152,153]. Thus, the different models of obesity complement each other in terms of knowledge and in the search for effective therapies to solve the public health problem that obesity represents. However, all results must be critically evaluated, since preclinical animal models can never be considered fully valid due to the uncertainties introduced by species differences [154].
3. Potential Markers for Obesity Assessment in Rodents
There are no standard parameters to classify obesity in rodents as there are in humans. However, there are some markers that are routinely used to analyze the degree of obesity and evaluate the effect of anti-obesity compounds. These parameters include body weight, body mass index, total body fat, adiposity index, Lee index, fat pad mass, insulin tolerance test, blood pressure, magnetic resonance imaging, and dual-energy X ray absorptiometry [155,156,157].
Body mass index is calculated by dividing body weight (g) by the square of tailless body length (m2), while the adiposity index is the total body fat (the sum of epididymal, retroperitoneal, and visceral fat depots) divided by the final body weight and multiplied by 100. The adiposity index is used as a marker of obesity because the degree of fat tends to increase with obesity, allowing accurate assessment of body fat percentage [158,159]. The whole-body composition can also be measured using imaging methods such as the magnetic resonance imaging system [157] and the dual energy X-ray absorptiometry [156]. Another parameter that can be determined is the body weight gain, which consists of subtracting the final body weight value by the initial body weight, dividing by the initial body weight, and multiplying by 100 [160]. The food intake can also be calculated as well as the feed efficiency ratio (FER) according to the equation: FER = (body weight gain/food intake) × 100 [161]. The Lee index consists of the division of cubicle root of the weight (g) by the nasoanal length (mm) multiplied by 100 [162], and obesity is defined by a Lee index greater than 310,100 [163].
The glucose tolerance test (GTT) is a simple approach used for assessing glucose homeostasis in rodents. It consists of measuring blood glucose after fasting followed by administration of a glucose dose. The fasting period, glucose dose, and routes of administration are variables to be considered in the experimental design [164,165,166]. The glucose solution is usually administered by oral gavage or intraperitoneal injection [165]. Both routes of administration are accepted; however, the plasma glucose levels are significantly higher in response to intraperitoneal administration compared to the same glucose dose administered orally [165]. The fasting period is variable, and the standard approach comprises a period of 14–18 h fasting (overnight fast) [165,167]. Although the overnight is the most common method, a shorter fast (5–6 h—morning fast) is more physiological for mice [168]. After glucose administration, the blood sample is collected at several 15 min intervals and the glucose levels are measured on a glucometer [168]. The insulin tolerance test (ITT) is a method similar to GTT, in which insulin is administered instead of glucose, usually in animals that have fasted for 6 h. After insulin administration, the blood glucose levels are measured every 15–30 min during 60–90 min [166].
Blood pressure is a parameter used to estimate cardiovascular functions in rodents. Blood pressure can be determinate by invasive, noninvasive, and radiotelemetry methods, with the invasive method being the standard approach [169]. The most common methods used are: tail cuff plethysmography (noninvasive); tip catheter (invasive); and blood pressure telemetry [170].
Additionally, adipose tissue histology (size and frequency of adipocytes), blood biochemistry analysis (for the assessment of total cholesterol, low-density lipoprotein, high-density lipoprotein, serum triglyceride, leptin, adiponectin, glucose and insulin levels), and liver histology (% of hepatocytes containing one or several lipid vacuoles for verified presence of steatosis) may be performed [3,171].
4. Conclusions
The rodent models used to study obesity have been an extremely important tool over the years, allowing not only the understanding of the cellular and molecular mechanisms underlying the development of obesity, but also the evaluation of new therapeutic approaches. The diversity of rodent models allows us to assess the various interactions between genetic and environmental factors involved in this disease, but all models have their advantages and limitations, which should be taken into account when choosing a model for a particular study objective.
Author Contributions
Conceptualization, T.M., C.C.-R., S.L., T.F., E.N.-G., E.R., P.A.O. and L.M.A.; writing—original draft preparation, T.M., C.C.-R., S.L., T.F. and E.N.-G.; writing—review and editing, T.M., C.C.-R., S.L., T.F., E.N.-G., L.M.A. and P.A.O.; supervision, P.A.O. and L.M.A.; funding acquisition, E.R. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Portuguese Foundation for Science and Technology (FCT) and co-financed by the European Regional Development Fund (FEDER) through COMPETE 2020—Operational Competitiveness and Internationalization Programme (POCI), grant PTDC/ASP-HOR/29152/2017, POCI-01-0145-FEDER-029152 (VALORIZEBYPRODUCTS). This work was also supported by National Funds by FCT—Portuguese Foundation for Science and Technology, under the project UIDB/04033/2020. The authors acknowledge the financial support by the Portuguese Foundation for Science and Technology (FCT) through a Doctoral Grant (2020.04789.BD, Tiago Ferreira and BD/136747/2018, Elisabete Nascimento-Gonçalves).
Conflicts of Interest
The authors declare no conflict of interest.
References
- WHO. Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 10 January 2022).
- Lau, D.C.; Douketis, J.D.; Morrison, K.M.; Hramiak, I.M.; Sharma, A.M.; Ur, E.; Obesity Canada Clinical Practice Guidelines Expert Panel. 2006 Canadian clinical practice guidelines on the management and prevention of obesity in adults and children [summary]. CMAJ 2007, 176, S1–S13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novelli, E.L.; Diniz, Y.S.; Galhardi, C.M.; Ebaid, G.M.; Rodrigues, H.G.; Mani, F.; Fernandes, A.A.; Cicogna, A.C.; Novelli Filho, J.L. Anthropometrical parameters and markers of obesity in rats. Lab. Anim. 2007, 41, 111–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaila, B.; Raman, M. Obesity: A review of pathogenesis and management strategies. Can. J. Gastroenterol. 2008, 22, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Mittwede, P.N.; Clemmer, J.S.; Bergin, P.F.; Xiang, L. Obesity and Critical Illness: Insights from Animal Models. Shock 2016, 45, 349–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanasaki, K.; Koya, D. Biology of obesity: Lessons from animal models of obesity. J. Biomed. Biotechnol. 2011, 2011, 197636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleinert, M.; Clemmensen, C.; Hofmann, S.M.; Moore, M.C.; Renner, S.; Woods, S.C.; Huypens, P.; Beckers, J.; de Angelis, M.H.; Schurmann, A.; et al. Animal models of obesity and diabetes mellitus. Nat. Rev. Endocrinol. 2018, 14, 140–162. [Google Scholar] [CrossRef] [Green Version]
- Grotto, D.; Camargo, I.F.; Kodaira, K.; Mazzei, L.G.; Castro, J.; Vieira, R.A.L.; Bergamaschi, C.C.; Lopes, L.C. Effect of mushrooms on obesity in animal models: Study protocol for a systematic review and meta-analysis. Syst. Rev. 2019, 8, 288. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, G.A.; Ibrahim, S.R.M.; Elkhayat, E.S.; El Dine, R.S. Natural anti-obesity agents. Bull. Fac. Pharm. Cairo Univ. 2014, 52, 269–284. [Google Scholar] [CrossRef] [Green Version]
- Parasuraman, S.; Wen, L.E. Animal Model for Obesity-An Overview. Syst. Rev. Pharm. 2016, 6, 9–12. [Google Scholar] [CrossRef]
- Geiger, B.M.; Pothos, E.N. Chapter 1—Translating Animal Models of Obesity and Diabetes to the Clinic. In Handbook of Behavioral Neuroscience; Nomikos, G.G., Feltner, D.E., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; Volume 29, pp. 1–16. [Google Scholar]
- Barré-Sinoussi, F.; Montagutelli, X. Animal models are essential to biological research: Issues and perspectives. Future Sci. OA 2015, 1, FSO63. [Google Scholar] [CrossRef] [Green Version]
- Barrett, P.; Mercer, J.G.; Morgan, P.J. Preclinical models for obesity research. Dis. Models Mech. 2016, 9, 1245–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutz, T.A.; Woods, S.C. Overview of animal models of obesity. Curr. Protoc. Pharm. 2012, 58, 5–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preguica, I.; Alves, A.; Nunes, S.; Fernandes, R.; Gomes, P.; Viana, S.D.; Reis, F. Diet-induced rodent models of obesity-related metabolic disorders-A guide to a translational perspective. Obes. Rev. 2020, 21, e13081. [Google Scholar] [CrossRef] [PubMed]
- Speakman, J.; Hambly, C.; Mitchell, S.; Krol, E. Animal models of obesity. Obes. Rev. 2007, 8 (Suppl. 1), 55–61. [Google Scholar] [CrossRef]
- Suleiman, J.B.; Mohamed, M.; Bakar, A.B.A. A systematic review on different models of inducing obesity in animals: Advantages and limitations. J. Adv. Vet. Anim. Res. 2020, 7, 103–114. [Google Scholar] [CrossRef]
- Speakman, J.; Hambly, C.; Mitchell, S.; Krol, E. The contribution of animal models to the study of obesity. Lab. Anim. 2008, 42, 413–432. [Google Scholar] [CrossRef]
- Crujeiras, A.B.; Carreira, M.C.; Cabia, B.; Andrade, S.; Amil, M.; Casanueva, F.F. Leptin resistance in obesity: An epigenetic landscape. Life Sci. 2015, 140, 57–63. [Google Scholar] [CrossRef]
- Liu, J.; Lee, J.; Salazar Hernandez, M.A.; Mazitschek, R.; Ozcan, U. Treatment of obesity with celastrol. Cell 2015, 161, 999–1011. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.T.; Acohido, B.V.; Clifton, D.K.; Steiner, R.A. KiSS-1 neurones are direct targets for leptin in the ob/ob mouse. J. Neuroendocr. 2006, 18, 298–303. [Google Scholar] [CrossRef]
- Ferguson, D.; Blenden, M.; Hutson, I.; Du, Y.; Harris, C.A. Mouse Embryonic Fibroblasts Protect ob/ob Mice From Obesity and Metabolic Complications. Endocrinology 2018, 159, 3275–3286. [Google Scholar] [CrossRef]
- van der Spek, R.; Kreier, F.; Fliers, E.; Kalsbeek, A. Chapter 11—Circadian rhythms in white adipose tissue. In Progress in Brain Research; Kalsbeek, A., Merrow, M., Roenneberg, T., Foster, R.G., Eds.; Elsevier: Amsterdam, The Netherlands, 2012; Volume 199, pp. 183–201. [Google Scholar]
- Bao, R.; Meng, Y.; Zhang, H.; Yang, C.; Li, W.; Zhang, C.; Zhang, J.; Sun, R.; Li, Z.; Jiang, W.; et al. Elaiophylin reduces body weight and lowers glucose levels in obese mice by activating AMPK. Cell Death Dis. 2021, 12, 972. [Google Scholar] [CrossRef] [PubMed]
- Hua, H.; Zhang, Y.; Zhao, F.; Chen, K.; Wu, T.; Liu, Q.; Huang, S.; Zhang, A.; Jia, Z. Celastrol inhibits intestinal lipid absorption by reprofiling the gut microbiota to attenuate high-fat diet-induced obesity. iScience 2021, 24, 102077. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wang, L.; Li, X.; Wu, Y.; Yin, F.; Liu, J. Trilobatin ameliorates insulin resistance through IRS-AKT-GLUT4 signaling pathway in C2C12 myotubes and ob/ob mice. Chin. Med. 2020, 15, 110. [Google Scholar] [CrossRef] [PubMed]
- Simperova, A.; Al-Nakkash, L.; Faust, J.J.; Sweazea, K.L. Genistein supplementation prevents weight gain but promotes oxidative stress and inflammation in the vasculature of female obese ob/ob mice. Nutr. Res. 2016, 36, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Wargent, E.T.; Zaibi, M.S.; Silvestri, C.; Hislop, D.C.; Stocker, C.J.; Stott, C.G.; Guy, G.W.; Duncan, M.; Di Marzo, V.; Cawthorne, M.A. The cannabinoid Delta(9)-tetrahydrocannabivarin (THCV) ameliorates insulin sensitivity in two mouse models of obesity. Nutr. Diabetes 2013, 3, e68. [Google Scholar] [CrossRef]
- Sun, S.-S.; Wang, K.; Ma, K.; Bao, L.; Liu, H.-W. An insoluble polysaccharide from the sclerotium of Poria cocos improves hyperglycemia, hyperlipidemia and hepatic steatosis in ob/ob mice via modulation of gut microbiota. Chin. J. Nat. Med. 2019, 17, 3–14. [Google Scholar] [CrossRef]
- Chung, M.Y.; Park, H.J.; Manautou, J.E.; Koo, S.I.; Bruno, R.S. Green tea extract protects against nonalcoholic steatohepatitis in ob/ob mice by decreasing oxidative and nitrative stress responses induced by proinflammatory enzymes. J. Nutr. Biochem. 2012, 23, 361–367. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Manabe, Y.; Sugawara, T. Siphonaxanthin, a carotenoid from green algae Codium cylindricum, protects Ob/Ob mice fed on a high-fat diet against lipotoxicity by ameliorating somatic stresses and restoring anti-oxidative capacity. Nutr. Res. 2020, 77, 29–42. [Google Scholar] [CrossRef]
- Kitada, M.; Ogura, Y.; Koya, D. Rodent models of diabetic nephropathy: Their utility and limitations. Int. J. Nephrol. Renov. Dis. 2016, 9, 279–290. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Mazcorro, J.F.; Mills, D.A.; Murphy, K.; Noratto, G. Effect of barley supplementation on the fecal microbiota, caecal biochemistry, and key biomarkers of obesity and inflammation in obese db/db mice. Eur. J. Nutr. 2018, 57, 2513–2528. [Google Scholar] [CrossRef]
- Guo, Y.; Fu, W.; Xin, Y.; Bai, J.; Peng, H.; Fu, L.; Liu, J.; Li, L.; Ma, Y.; Jiang, H. Antidiabetic and Antiobesity Effects of Artemether in db/db Mice. BioMed Res. Int. 2018, 2018, 8639523. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Koh, E.; Kim, D.; Lee, N.; Cho, S.M.; Lee, Y.J.; Cho, I.H.; Yang, H.J. Dendropanax trifidus Sap-Mediated Suppression of Obese Mouse Body Weight and the Metabolic Changes Related with Estrogen Receptor Alpha and AMPK-ACC Pathways in Muscle Cells. Nutrients 2022, 14, 1098. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Fang, K.; Dan, X.; Gu, M. Crocin ameliorates hepatic steatosis through activation of AMPK signaling in db/db mice. Lipids Health Dis. 2019, 18, 11. [Google Scholar] [CrossRef] [Green Version]
- Bates, S.H.; Stearns, W.H.; Dundon, T.A.; Schubert, M.; Tso, A.W.; Wang, Y.; Banks, A.S.; Lavery, H.J.; Haq, A.K.; Maratos-Flier, E.; et al. STAT3 signalling is required for leptin regulation of energy balance but not reproduction. Nature 2003, 421, 856–859. [Google Scholar] [CrossRef] [PubMed]
- Saadat, N.; IglayReger, H.B.; Myers, M.G., Jr.; Bodary, P.; Gupta, S.V. Differences in metabolomic profiles of male db/db and s/s, leptin receptor mutant mice. Physiol. Genom. 2012, 44, 374–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, D.L.; Eicher, E.M. Fat (fat) and Tubby (tub): Two Autosomal Recessive Mutations Causing Obesity Syndromes in the Mouse. J. Hered. 1990, 81, 424–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleyn, P.W.; Fan, W.; Kovats, S.G.; Lee, J.J.; Pulido, J.C.; Wu, Y.; Berkemeier, L.R.; Misumi, D.J.; Holmgren, L.; Charlat, O.; et al. Identification and Characterization of the Mouse Obesity Gene tubby: A Member of a Novel Gene Family. Cell 1996, 85, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Granados, M.J.; Ramírez-Emiliano, J.; Elena Franco-Robles, E. Rodent Models of Obesity and Diabetes. In Experimental Animal Models of Human Diseases—An Effective Therapeutic Strategy; Bartholomew, I., Ed.; IntechOpen: London, UK, 2018. [Google Scholar]
- Yorek, M.A. Alternatives to the Streptozotocin-Diabetic Rodent. Int. Rev. Neurobiol. 2016, 127, 89–112. [Google Scholar] [CrossRef] [Green Version]
- Zucker, L.M.; Zucker, T.F. Fatty, A new mutation in the rat. J. Hered. 1961, 52, 275–278. [Google Scholar] [CrossRef]
- Owens, D.R. Chapter 23—Spontaneous, Surgically and Chemically Induced Models of Disease. In The Laboratory Rat, 2nd ed.; Suckow, M.A., Weisbroth, S.H., Franklin, C.L., Eds.; Academic Press: Burlington, VT, USA, 2006; pp. 711–732. [Google Scholar]
- Agil, A.; Navarro-Alarcon, M.; Ali, F.A.Z.; Albrakati, A.; Salagre, D.; Campoy, C.; Elmahallawy, E.K. Melatonin Enhances the Mitochondrial Functionality of Brown Adipose Tissue in Obese-Diabetic Rats. Antioxidants 2021, 10, 1482. [Google Scholar] [CrossRef]
- Kato, Y.; Sakoh, M.; Nagai, T.; Yoshida, A.; Ishida, H.; Inoue, N.; Yanagita, T.; Nagao, K. Ozonated Olive Oil Alleviates Hepatic Steatosis in Obese Zucker (fa/fa) Rats. J. Oleo Sci. 2022; ahead of print. [Google Scholar] [CrossRef]
- Szabadfi, K.; Pinter, E.; Reglodi, D.; Gabriel, R. Chapter One—Neuropeptides, Trophic Factors, and Other Substances Providing Morphofunctional and Metabolic Protection in Experimental Models of Diabetic Retinopathy. In International Review of Cell and Molecular Biology; Jeon, K.W., Ed.; Academic Press: Cambridge, MA, USA, 2014; Volume 311, pp. 1–121. [Google Scholar]
- Cho, I.J.; Oh, D.H.; Yoo, J.; Hwang, Y.C.; Ahn, K.J.; Chung, H.Y.; Jeong, S.W.; Moon, J.Y.; Lee, S.H.; Lim, S.J.; et al. Allopurinol ameliorates high fructose diet induced hepatic steatosis in diabetic rats through modulation of lipid metabolism, inflammation, and ER stress pathway. Sci. Rep. 2021, 11, 9894. [Google Scholar] [CrossRef] [PubMed]
- Schwarzer, M. Chapter 8—Models to Investigate Cardiac Metabolism. In The Scientist’s Guide to Cardiac Metabolism; Schwarzer, M., Doenst, T., Eds.; Academic Press: Boston, MA, USA, 2016; pp. 103–122. [Google Scholar]
- Shin, Y.; Lee, D.; Ahn, J.; Lee, M.; Shin, S.S.; Yoon, M. The herbal extract ALS-L1023 from Melissa officinalis reduces weight gain, elevated glucose levels and β-cell loss in Otsuka Long-Evans Tokushima fatty rats. J. Ethnopharmacol. 2021, 264, 113360. [Google Scholar] [CrossRef] [PubMed]
- Wanezaki, S.; Saito, S.; Inoue, N.; Tachibana, N.; Shirouchi, B.; Sato, M.; Yanagita, T.; Nagao, K. Soy β-Conglycinin Peptide Attenuates Obesity and Lipid Abnormalities in Obese Model OLETF Rats. J. Oleo Sci. 2020, 69, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Bi, S.; Moran, T.H. Actions of CCK in the controls of food intake and body weight: Lessons from the CCK-A receptor deficient OLETF rat. Neuropeptides 2002, 36, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.-Y.; Lin, C.-H.; Huang, T.-H.; Chuang, S.-Y. In Vivo Rodent Models of Type 2 Diabetes and Their Usefulness for Evaluating Flavonoid Bioactivity. Nutrients 2019, 11, 530. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.W.; Sun, G.D.; Sun, J.; Liu, S.J.; Wang, J.; Xu, X.H.; Miao, L.N. Spontaneous type 2 diabetic rodent models. J. Diabetes Res. 2013, 2013, 401723. [Google Scholar] [CrossRef]
- Joost, H.G. The genetic basis of obesity and type 2 diabetes: Lessons from the new zealand obese mouse, a polygenic model of the metabolic syndrome. Results Probl. Cell Differ. 2010, 52, 1–11. [Google Scholar] [CrossRef]
- Kluge, R.; Scherneck, S.; Schürmann, A.; Joost, H.G. Pathophysiology and genetics of obesity and diabetes in the New Zealand obese mouse: A model of the human metabolic syndrome. Methods Mol. Biol. 2012, 933, 59–73. [Google Scholar] [CrossRef]
- Jurgens, H.S.; Schurmann, A.; Kluge, R.; Ortmann, S.; Klaus, S.; Joost, H.G.; Tschop, M.H. Hyperphagia, lower body temperature, and reduced running wheel activity precede development of morbid obesity in New Zealand obese mice. Physiol. Genom. 2006, 25, 234–241. [Google Scholar] [CrossRef] [Green Version]
- Korovila, I.; Höhn, A.; Jung, T.; Grune, T.; Ott, C. Reduced Liver Autophagy in High-Fat Diet Induced Liver Steatosis in New Zealand Obese Mice. Antioxidants 2021, 10, 501. [Google Scholar] [CrossRef]
- Fajardo, R.J.; Karim, L.; Calley, V.I.; Bouxsein, M.L. A review of rodent models of type 2 diabetic skeletal fragility. J. Bone Min. Res. 2014, 29, 1025–1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noge, I.; Kagawa, Y.; Maeda, T. A new diabetic mouse model derived from the ddY strain. Biol. Pharm. Bull. 2010, 33, 988–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, R. A Comprehensive Review: The Use of Animal Models in Diabetes Research. J. Anal. Pharm. Res. 2016, 3, 00071. [Google Scholar] [CrossRef]
- Hussein, G.M.; Matsuda, H.; Nakamura, S.; Akiyama, T.; Tamura, K.; Yoshikawa, M. Protective and ameliorative effects of mate (Ilex paraguariensis) on metabolic syndrome in TSOD mice. Phytomed. Int. J. Phytother. Phytopharm. 2011, 19, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Nishitsuji, K.; Watanabe, S.; Xiao, J.; Nagatomo, R.; Ogawa, H.; Tsunematsu, T.; Umemoto, H.; Morimoto, Y.; Akatsu, H.; Inoue, K.; et al. Effect of coffee or coffee components on gut microbiome and short-chain fatty acids in a mouse model of metabolic syndrome. Sci. Rep. 2018, 8, 16173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, T.; Nishida, T.; Baba, H.; Hatta, H.; Imura, J.; Sutoh, M.; Toyohara, S.; Hokao, R.; Watanabe, S.; Ogawa, H.; et al. Histopathological characteristics of glutamine synthetase-positive hepatic tumor lesions in a mouse model of spontaneous metabolic syndrome (TSOD mouse). Mol. Clin. Oncol. 2016, 5, 267–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akase, T.; Shimada, T.; Harasawa, Y.; Akase, T.; Ikeya, Y.; Nagai, E.; Iizuka, S.; Nakagami, G.; Iizaka, S.; Sanada, H.; et al. Preventive Effects of Salacia reticulata on Obesity and Metabolic Disorders in TSOD Mice. Evid.-Based Complement. Altern. Med. 2011, 2011, 484590. [Google Scholar] [CrossRef] [Green Version]
- Kudo, T.; Toda, T.; Ushiki, T.; Ohi, K.; Ikarashi, N.; Ochiai, W.; Sugiyama, K. Differences in the pharmacokinetics of Cyp3a substrates in TSOD and streptozotocin-induced diabetic mice. Xenobiotica 2010, 40, 282–290. [Google Scholar] [CrossRef]
- Miki, S.; Inokuma, K.I.; Takashima, M.; Nishida, M.; Sasaki, Y.; Ushijima, M.; Suzuki, J.I.; Morihara, N. Aged garlic extract suppresses the increase of plasma glycated albumin level and enhances the AMP-activated protein kinase in adipose tissue in TSOD mice. Mol. Nutr. Food Res. 2017, 61, 1600797. [Google Scholar] [CrossRef]
- Othman, M.B.; Sakamoto, K. Effect of inactivated Bifidobacterium longum intake on obese diabetes model mice (TSOD). Food Res. Int. 2020, 129, 108792. [Google Scholar] [CrossRef]
- Shida, K.; Ohsawa, M.; Takahashi, S.; Ota, H.; Tamura, T.; Kusama, N.; Nakasone, M.; Yamazaki, H.; Sobue, K. Peripheral neuropathy in the pre-diabetic state of the type 2 diabetes mouse model (TSOD mice) involves TRPV1 expression in dorsal root ganglions. IBRO Neurosci. Rep. 2022, 12, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Tanaka, T.; Tanaka, M. Potential Cancer Chemopreventive Activity of Protocatechuic Acid. J. Exp. Clin. Med. 2011, 3, 27–33. [Google Scholar] [CrossRef]
- Nishitsuji, K.; Xiao, J.; Nagatomo, R.; Umemoto, H.; Morimoto, Y.; Akatsu, H.; Inoue, K.; Tsuneyama, K. Analysis of the gut microbiome and plasma short-chain fatty acid profiles in a spontaneous mouse model of metabolic syndrome. Sci. Rep. 2017, 7, 15876. [Google Scholar] [CrossRef] [PubMed]
- Scroyen, I.; Hemmeryckx, B.; Lijnen, H.R. From mice to men--mouse models in obesity research: What can we learn? Thromb. Haemost. 2013, 110, 634–640. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.-S.; Kim, J. Application of Animal Models in Diabetic Cardiomyopathy. DMJ 2021, 45, 129–145. [Google Scholar] [CrossRef]
- Allan, M.F.; Eisen, E.J.; Pomp, D. The M16 mouse: An outbred animal model of early onset polygenic obesity and diabesity. Obes. Res. 2004, 12, 1397–1407. [Google Scholar] [CrossRef]
- Huijbers, I.J. Generating Genetically Modified Mice: A Decision Guide. In Site-Specific Recombinases, 2017/08/18 ed.; Eroshenko, N., Ed.; Methods in Molecular Biology; Humana Press: New York, NY, USA, 2017; Volume 1642, pp. 1–19. [Google Scholar]
- Stengel, A.; Goebel, M.; Million, M.; Stenzel-Poore, M.P.; Kobelt, P.; Monnikes, H.; Tache, Y.; Wang, L. Corticotropin-releasing factor-overexpressing mice exhibit reduced neuronal activation in the arcuate nucleus and food intake in response to fasting. Endocrinology 2009, 150, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Goebel-Stengel, M.; Yuan, P.Q.; Stengel, A.; Tache, Y. Corticotropin-releasing factor overexpression in mice abrogates sex differences in body weight, visceral fat, and food intake response to a fast and alters levels of feeding regulatory hormones. Biol. Sex Differ. 2017, 8, 2. [Google Scholar] [CrossRef] [Green Version]
- Kang, W.; Tong, T.; Park, T. Corticotropin releasing factor-overexpressing mouse is a model of chronic stress-induced muscle atrophy. PLoS ONE 2020, 15, e0229048. [Google Scholar] [CrossRef] [Green Version]
- Ross, J.A.; Alexis, R.; Reyes, B.A.S.; Risbrough, V.; Van Bockstaele, E.J. Localization of amyloid beta peptides to locus coeruleus and medial prefrontal cortex in corticotropin releasing factor overexpressing male and female mice. Brain Struct. Funct. 2019, 224, 2385–2405. [Google Scholar] [CrossRef]
- Ludwig, D.S.; Tritos, N.A.; Mastaitis, J.W.; Kulkarni, R.; Kokkotou, E.; Elmquist, J.; Lowell, B.; Flier, J.S.; Maratos-Flier, E. Melanin-concentrating hormone overexpression in transgenic mice leads to obesity and insulin resistance. J. Clin. Investig. 2001, 107, 379–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pissios, P.; Ozcan, U.; Kokkotou, E.; Okada, T.; Liew, C.W.; Liu, S.; Peters, J.N.; Dahlgren, G.; Karamchandani, J.; Kudva, Y.C.; et al. Melanin concentrating hormone is a novel regulator of islet function and growth. Diabetes 2007, 56, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Pereira-da-Silva, M.; De Souza, C.T.; Gasparetti, A.L.; Saad, M.J.; Velloso, L.A. Melanin-concentrating hormone induces insulin resistance through a mechanism independent of body weight gain. J. Endocrinol. 2005, 186, 193–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tschop, M.; Heiman, M.L. Overview of rodent models for obesity research. Curr. Protoc. Neurosci. 2002, 17, 9–10. [Google Scholar] [CrossRef] [PubMed]
- Lord, M.N.; Subramanian, K.; Kanoski, S.E.; Noble, E.E. Melanin-concentrating hormone and food intake control: Sites of action, peptide interactions, and appetition. Peptides 2021, 137, 170476. [Google Scholar] [CrossRef]
- Li, C.; Xia, J.; Zhu, W.; Xin, L.; An, C.; Yang, S.; Li, K. Systemic overexpression of the 11betaHSD1 promotes endoplasmic reticulum stress in multiple tissues and the development of metabolic syndrome in mice. Mol. Med. Rep. 2017, 16, 7738–7744. [Google Scholar] [CrossRef]
- Arch, J.R. Lessons in obesity from transgenic animals. J. Endocrinol. Investig. 2002, 25, 867–875. [Google Scholar] [CrossRef]
- Dammann, C.; Stapelfeld, C.; Maser, E. Expression and activity of the cortisol-activating enzyme 11β-hydroxysteroid dehydrogenase type 1 is tissue and species-specific. Chem.-Biol. Interact. 2019, 303, 57–61. [Google Scholar] [CrossRef]
- Atkinson, M.D.; Atkinson, E. Sambucus nigra L. J. Ecol. 2002, 90, 895–923. [Google Scholar] [CrossRef]
- Tsao, T.-S.; Katz, E.B.; Pommer, D.; Charron, M.J. Amelioration of insulin resistance but not hyperinsulinemia in obese mice overexpressing GLUT4 selectively in skeletal muscle. Metab. Clin. Exp. 2000, 49, 340–346. [Google Scholar] [CrossRef]
- Shepherd, P.R.; Gnudi, L.; Tozzo, E.; Yang, H.; Leach, F.; Kahn, B.B. Adipose cell hyperplasia and enhanced glucose disposal in transgenic mice overexpressing GLUT4 selectively in adipose tissue. J. Biol. Chem. 1993, 268, 22243–22246. [Google Scholar] [CrossRef]
- Townsend, K.; Tseng, Y.H. Brown adipose tissue: Recent insights into development, metabolic function and therapeutic potential. Adipocyte 2012, 1, 13–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowell, B.B.; Hamann, A.; Lawitts, J.A.; Himms-Hagen, J.; Boyer, B.B.; Kozak, L.P.; Flier, J.S. Development of obesity in transgenic mice after genetic ablation of brown adipose tissue. Nature 1993, 366, 740–742. [Google Scholar] [CrossRef]
- Klaus, S.; Munzberg, H.; Truloff, C.; Heldmaier, G. Physiology of transgenic mice with brown fat ablation: Obesity is due to lowered body temperature. Am. J. Physiol. 1998, 274, R287–R293. [Google Scholar] [CrossRef] [PubMed]
- Hamann, A.; Flier, J.S.; Lowell, B.B. Decreased brown fat markedly enhances susceptibility to diet-induced obesity, diabetes, and hyperlipidemia. Endocrinology 1996, 137, 21–29. [Google Scholar] [CrossRef]
- Callahan, Z.J.; Oxendine, M.J.; Schaeffer, P.J. Intramuscular triglyceride content precedes impaired glucose metabolism without evidence for mitochondrial dysfunction during early development of a diabetic phenotype. Appl. Physiol. Nutr. Metab. 2017, 42, 963–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hui, D.Y. Utility and importance of gene knockout animals for nutritional and metabolic research. J. Nutr. 1998, 128, 2052–2057. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, Y.; Oniki, K.; Kumagae, N.; Morita, K.; Otake, K.; Ogata, Y.; Saruwatari, J. Beta-3-adrenergic Receptor rs4994 Polymorphism Is a Potential Biomarker for the Development of Nonalcoholic Fatty Liver Disease in Overweight/Obese Individuals. Dis. Markers 2019, 2019, 4065327. [Google Scholar] [CrossRef]
- Schena, G.; Caplan, M.J. Everything You Always Wanted to Know about β3-AR * (* But Were Afraid to Ask). Cells 2019, 8, 357. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.; Liu, N.; Province, H.; Piñol, R.A.; Gavrilova, O.; Reitman, M.L. BRS3 in both MC4R- and SIM1-expressing neurons regulates energy homeostasis in mice. Mol. Metab. 2020, 36, 100969. [Google Scholar] [CrossRef]
- Fruhbeck, G.; Gomez-Ambrosi, J. Control of body weight: A physiologic and transgenic perspective. Diabetologia 2003, 46, 143–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, N.; Moreno, P.; Jensen, R.T. Bombesin receptor subtype 3 as a potential target for obesity and diabetes. Expert Opin. Targets 2015, 19, 1153–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleinridders, A.; Cai, W.; Cappellucci, L.; Ghazarian, A.; Collins, W.R.; Vienberg, S.G.; Pothos, E.N.; Kahn, C.R. Insulin resistance in brain alters dopamine turnover and causes behavioral disorders. Proc. Natl. Acad. Sci. USA 2015, 112, 3463–3468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schubert, M.; Gautam, D.; Surjo, D.; Ueki, K.; Baudler, S.; Schubert, D.; Kondo, T.; Alber, J.; Galldiks, N.; Kustermann, E.; et al. Role for neuronal insulin resistance in neurodegenerative diseases. Proc. Natl. Acad. Sci. USA 2004, 101, 3100–3105. [Google Scholar] [CrossRef] [Green Version]
- Bertocchi, I.; Oberto, A.; Longo, A.; Palanza, P.; Eva, C. Conditional inactivation of Npy1r gene in mice induces sex-related differences of metabolic and behavioral functions. Horm. Behav. 2020, 125, 104824. [Google Scholar] [CrossRef] [PubMed]
- Paterlini, S.; Panelli, R.; Gioiosa, L.; Parmigiani, S.; Franceschini, P.; Bertocchi, I.; Oberto, A.; Bartolomucci, A.; Eva, C.; Palanza, P. Conditional Inactivation of Limbic Neuropeptide Y-1 Receptors Increases Vulnerability to Diet-Induced Obesity in Male Mice. Int. J. Mol. Sci. 2021, 22, 8745. [Google Scholar] [CrossRef]
- Burke, L.K.; Doslikova, B.; D’Agostino, G.; Garfield, A.S.; Farooq, G.; Burdakov, D.; Low, M.J.; Rubinstein, M.; Evans, M.L.; Billups, B.; et al. 5-HT obesity medication efficacy via POMC activation is maintained during aging. Endocrinology 2014, 155, 3732–3738. [Google Scholar] [CrossRef] [Green Version]
- D’Agostino, G.; Lyons, D.; Cristiano, C.; Lettieri, M.; Olarte-Sanchez, C.; Burke, L.K.; Greenwald-Yarnell, M.; Cansell, C.; Doslikova, B.; Georgescu, T.; et al. Nucleus of the Solitary Tract Serotonin 5-HT2C Receptors Modulate Food Intake. Cell Metab. 2018, 28, 619–630.e5. [Google Scholar] [CrossRef] [Green Version]
- Tecott, L.H.; Sun, L.M.; Akana, S.F.; Strack, A.M.; Lowenstein, D.H.; Dallman, M.F.; Julius, D. Eating disorder and epilepsy in mice lacking 5-HT2c serotonin receptors. Nature 1995, 374, 542–546. [Google Scholar] [CrossRef]
- Sargent, B.J.; Henderson, A.J. Targeting 5-HT receptors for the treatment of obesity. Curr. Opin. Pharm. 2011, 11, 52–58. [Google Scholar] [CrossRef]
- Canal, C.E.; Olaghere da Silva, U.B.; Gresch, P.J.; Watt, E.E.; Sanders-Bush, E.; Airey, D.C. The serotonin 2C receptor potently modulates the head-twitch response in mice induced by a phenethylamine hallucinogen. Psychopharmacology 2010, 209, 163–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nebuka, M.; Ohmura, Y.; Izawa, S.; Bouchekioua, Y.; Nishitani, N.; Yoshida, T.; Yoshioka, M. Behavioral characteristics of 5-HT2C receptor knockout mice: Locomotor activity, anxiety-, and fear memory-related behaviors. Behav. Brain Res. 2020, 379, 112394. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Lee, N.J.; Ip, C.K.; Enriquez, R.; Tasan, R.; Zhang, L.; Herzog, H. NPY derived from AGRP neurons controls feeding via Y1 and energy expenditure and food foraging behaviour via Y2 signalling. Mol. Metab. 2022, 59, 101455. [Google Scholar] [CrossRef] [PubMed]
- Simpson, K.A.; Martin, N.M.; Bloom, S.R. Hypothalamic regulation of food intake and clinical therapeutic applications. Arq. Bras. Endocrinol. Metabol. 2009, 53, 120–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roth, C.L.; D’Ambrosio, G.; Elfers, C. Activation of nuclear factor kappa B pathway and reduction of hypothalamic oxytocin following hypothalamic lesions. J. Syst. Integr. Neurosci. 2016, 2, 79–84. [Google Scholar] [CrossRef] [Green Version]
- Gaur, A.; Pal, G.K.; Pal, P. Role of Ventromedial Hypothalamus in Sucrose-Induced Obesity on Metabolic Parameters. Ann. Neurosci. 2021, 28, 39–46. [Google Scholar] [CrossRef]
- Kiba, T.; Tanaka, K.; Numata, K.; Hoshino, M.; Misugi, K.; Inoue, S. Ventromedial hypothalamic lesion-induced vagal hyperactivity stimulates rat pancreatic cell proliferation. Gastroenterology 1996, 110, 885–893. [Google Scholar] [CrossRef]
- Kinyua, A.W.; Yang, D.J.; Chang, I.; Kim, K.W. Steroidogenic Factor 1 in the Ventromedial Nucleus of the Hypothalamus Regulates Age-Dependent Obesity. PLoS ONE 2016, 11, e0162352. [Google Scholar] [CrossRef]
- Von Diemen, V.; Trindade, E.N.; Trindade, M.R. Experimental model to induce obesity in rats. Acta Cir. Bras. 2006, 21, 425–429. [Google Scholar] [CrossRef]
- Miśkowiak, B.; Partyka, M. Effects of neonatal treatment with MSG (monosodium glutamate) on hypothalamo-pituitary-thyroid axis in adult male rats. Histol. Histopathol. 1993, 8, 731–734. [Google Scholar]
- Saito, M.; Minokoshi, Y.; Shimazu, T. Brown adipose tissue after ventromedial hypothalamic lesions in rats. Am. J. Physiol. 1985, 248, E20–E25. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Dallman, M.F. Hypothalamic obesity: Multiple routes mediated by loss of function in medial cell groups. Endocrinology 1999, 140, 4081–4088. [Google Scholar] [CrossRef]
- Fukushima, M.; Tokunaga, K.; Lupien, J.; Kemnitz, J.W.; Bray, G.A. Dynamic and static phases of obesity following lesions in PVN and VMH. Am. J. Physiol. 1987, 253, R523–R529. [Google Scholar] [CrossRef] [PubMed]
- Sims, J.S.; Lorden, J.F. Effect of paraventricular nucleus lesions on body weight, food intake and insulin levels. Behav. Brain Res. 1986, 22, 265–281. [Google Scholar] [CrossRef]
- An, J.J.; Liao, G.Y.; Kinney, C.E.; Sahibzada, N.; Xu, B. Discrete BDNF Neurons in the Paraventricular Hypothalamus Control Feeding and Energy Expenditure. Cell Metab. 2015, 22, 175–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhou, L.; Lian, H.; Zhang, Y.; Tong, S.; Wang, Z. Dopamine receptor 2 downregulation and brain-derived neurotrophic factor upregulation in the paraventricular nucleus are correlated with brown adipose tissue thermogenesis in rats with bilateral substantia nigra lesions. J. Chem. Neuroanat. 2021, 117, 102016. [Google Scholar] [CrossRef]
- Franca, L.M.; Freitas, L.N.; Chagas, V.T.; Coelho, C.F.; Barroso, W.A.; Costa, G.C.; Silva, L.A.; Debbas, V.; Laurindo, F.R.; Paes, A.M. Mechanisms underlying hypertriglyceridemia in rats with monosodium L-glutamate-induced obesity: Evidence of XBP-1/PDI/MTP axis activation. Biochem. Biophys. Res. Commun. 2014, 443, 725–730. [Google Scholar] [CrossRef]
- Gloy, V.; Langhans, W.; Hillebrand, J.J.; Geary, N.; Asarian, L. Ovariectomy and overeating palatable, energy-dense food increase subcutaneous adipose tissue more than intra-abdominal adipose tissue in rats. Biol. Sex Differ. 2011, 2, 6. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Yang, Q.; Zhou, C.; Jiang, H.; Sun, Y.; Wang, H.; Luo, X.; Wang, Z.; Zhang, J.; Wang, K.; et al. Transcriptomic changes in the hypothalamus of ovariectomized mice: Data from RNA-seq analysis. Ann. Anat.-Anat. Anz. 2022, 241, 151886. [Google Scholar] [CrossRef]
- Romero-Picó, A.; Novelle, M.G.; Al-Massadi, O.; Beiroa, D.; Tojo, M.; Heras, V.; Ruiz-Pino, F.; Senra, A.; López, M.; Blouet, C.; et al. Kappa-Opioid Receptor Blockade Ameliorates Obesity Caused by Estrogen Withdrawal via Promotion of Energy Expenditure through mTOR Pathway. Int. J. Mol. Sci. 2022, 23, 3118. [Google Scholar] [CrossRef]
- de Moura e Dias, M.; dos Reis, S.A.; da Conceição, L.L.; Sediyama, C.M.N.O.; Pereira, S.S.; de Oliveira, L.L.; Gouveia Peluzio, M.C.; Martinez, J.A.; Milagro, F.I. Diet-induced obesity in animal models: Points to consider and influence on metabolic markers. Diabetol. Metab. Syndr. 2021, 13, 32. [Google Scholar] [CrossRef] [PubMed]
- Panchal, S.K.; Brown, L. Rodent models for metabolic syndrome research. J. Biomed. Biotechnol. 2011, 2011, 351982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glastras, S.J.; Chen, H.; Teh, R.; McGrath, R.T.; Chen, J.; Pollock, C.A.; Wong, M.G.; Saad, S. Mouse Models of Diabetes, Obesity and Related Kidney Disease. PLoS ONE 2016, 11, e0162131. [Google Scholar] [CrossRef]
- Yang, Y.; Smith, D.L., Jr.; Keating, K.D.; Allison, D.B.; Nagy, T.R. Variations in body weight, food intake and body composition after long-term high-fat diet feeding in C57BL/6J mice. Obesity 2014, 22, 2147–2155. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.; Chandrasekera, P.C.; Barnard, N.D. You are what you eat, or are you? The challenges of translating high-fat-fed rodents to human obesity and diabetes. Nutr. Diabetes 2014, 4, e135. [Google Scholar] [CrossRef] [Green Version]
- Eng, J.M.; Estall, J.L. Diet-Induced Models of Non-Alcoholic Fatty Liver Disease: Food for Thought on Sugar, Fat, and Cholesterol. Cells 2021, 10, 1805. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Chen, D.; Zhang, K.; Yu, B. A high-amylopectin diet caused hepatic steatosis associated with more lipogenic enzymes and increased serum insulin concentration. Br. J. Nutr. 2011, 106, 1470–1475. [Google Scholar] [CrossRef] [Green Version]
- Wong, S.K.; Chin, K.Y.; Suhaimi, F.H.; Fairus, A.; Ima-Nirwana, S. Animal models of metabolic syndrome: A review. Nutr. Metab. 2016, 13, 65. [Google Scholar] [CrossRef] [Green Version]
- Bastias-Perez, M.; Serra, D.; Herrero, L. Dietary Options for Rodents in the Study of Obesity. Nutrients 2020, 12, 3234. [Google Scholar] [CrossRef]
- Virtuoso, A.; Tveden-Nyborg, P.; Schou-Pedersen, A.M.V.; Lykkesfeldt, J.; Müller, H.K.; Elfving, B.; Sørensen, D.B. A Long-Term Energy-Rich Diet Increases Prefrontal BDNF in Sprague-Dawley Rats. Nutrients 2022, 14, 126. [Google Scholar] [CrossRef]
- Matuszewska, J.; Zalewski, T.; Klimaszyk, A.; Ziarniak, K.; Jurga, S.; Chmurzynska, A.; Sliwowska, J.H. Mothers’ cafeteria diet induced sex-specific changes in fat content, metabolic profiles, and inflammation outcomes in rat offspring. Sci. Rep. 2021, 11, 18573. [Google Scholar] [CrossRef] [PubMed]
- Baccouche, B.; Benlarbi, M.; Barber, A.J.; Chaouacha-Chekir, R.B. Short-Term Administration of Astaxanthin Attenuates Retinal Changes in Diet-Induced Diabetic Psammomys obesus. Curr. Eye Res. 2018, 43, 1177–1189. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, R.; Walder, K.R.; Hagemeyer, C.E.; Kanwar, J.R. Psammomys obesus: A Natural Diet-Controlled Model for Diabetes and Cardiovascular Diseases. Curr. Atheroscler. Rep. 2018, 20, 46. [Google Scholar] [CrossRef]
- Shafrir, E.; Ziv, E.; Kalman, R. Nutritionally induced diabetes in desert rodents as models of type 2 diabetes: Acomys cahirinus (spiny mice) and Psammomys obesus (desert gerbil). ILAR J. 2006, 47, 212–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaouad, B.; Moudilou, E.N.; Ghoul, A.; Zerrouk, F.; Moulahoum, A.; Othmani-Mecif, K.; Cherifi, M.E.H.; Exbrayat, J.M.; Benazzoug, Y. Hyperhomocysteinemia and myocardial remodeling in the sand rat, Psammomys obesus. Acta Histochem. 2019, 121, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.X.; Harn, H.I.; Ou, K.L.; Lei, M.; Chuong, C.M. Comparative regenerative biology of spiny (Acomys cahirinus) and laboratory (Mus musculus) mouse skin. Exp. Dermatol. 2019, 28, 442–449. [Google Scholar] [CrossRef]
- Maden, M.; Brant, J.O.; Rubiano, A.; Sandoval, A.G.W.; Simmons, C.; Mitchell, R.; Collin-Hooper, H.; Jacobson, J.; Omairi, S.; Patel, K. Perfect chronic skeletal muscle regeneration in adult spiny mice, Acomys cahirinus. Sci. Rep. 2018, 8, 8920. [Google Scholar] [CrossRef] [Green Version]
- Simkin, J.; Gawriluk, T.R.; Gensel, J.C.; Seifert, A.W. Macrophages are necessary for epimorphic regeneration in African spiny mice. eLife 2017, 6, e24623. [Google Scholar] [CrossRef]
- Wong, W.; Kim, A.; Monaghan, J.R.; Seifert, A.W.; Maden, M.; Crane, J.D. Spiny mice (Acomys) exhibit attenuated hallmarks of aging and rapid cell turnover after UV exposure in the skin epidermis. PLoS ONE 2020, 15, e0241617. [Google Scholar] [CrossRef]
- Choudhary, A.; Ibdah, J.A. Animal models in today’s translational medicine world. Mo. Med. 2013, 110, 220–222. [Google Scholar]
- Loos, R.J.F.; Yeo, G.S.H. The genetics of obesity: From discovery to biology. Nat. Rev. Genet. 2022, 23, 120–133. [Google Scholar] [CrossRef] [PubMed]
- Sinasac, D.S.; Riordan, J.D.; Spiezio, S.H.; Yandell, B.S.; Croniger, C.M.; Nadeau, J.H. Genetic control of obesity, glucose homeostasis, dyslipidemia and fatty liver in a mouse model of diet-induced metabolic syndrome. Int. J. Obes. 2016, 40, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Flores-Dorantes, M.T.; Díaz-López, Y.E.; Gutiérrez-Aguilar, R. Environment and Gene Association With Obesity and Their Impact on Neurodegenerative and Neurodevelopmental Diseases. Front. Neurosci. 2020, 14, 863. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Li, H. Obesity: Epidemiology, Pathophysiology, and Therapeutics. Front. Endocrinol. 2021, 12, 706978. [Google Scholar] [CrossRef]
- Pound, P.; Ritskes-Hoitinga, M. Is it possible to overcome issues of external validity in preclinical animal research? Why most animal models are bound to fail. J. Transl. Med. 2018, 16, 304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardis, L.L. Prediction of carcass fat, water and lean body mass from Lee’s "nutritive ratio" in rats with hypothalamic obesity. Experientia 1970, 26, 789–790. [Google Scholar] [CrossRef]
- Gargiulo, S.; Gramanzini, M.; Megna, R.; Greco, A.; Albanese, S.; Manfredi, C.; Brunetti, A. Evaluation of growth patterns and body composition in C57Bl/6J mice using dual energy X-ray absorptiometry. BioMed Res. Int. 2014, 2014, 253067. [Google Scholar] [CrossRef] [Green Version]
- Nixon, J.P.; Zhang, M.; Wang, C.; Kuskowski, M.A.; Novak, C.M.; Levine, J.A.; Billington, C.J.; Kotz, C.M. Evaluation of a quantitative magnetic resonance imaging system for whole body composition analysis in rodents. Obesity 2010, 18, 1652–1659. [Google Scholar] [CrossRef] [Green Version]
- Francisqueti, F.V.; Nascimento, A.F.; Minatel, I.O.; Dias, M.C.; Luvizotto, R.A.M.; Berchieri-Ronchi, C.; Ferreira, A.L.A.; Correa, C.R. Metabolic syndrome and inflammation in adipose tissue occur at different times in animals submitted to a high-sugar/fat diet. J. Nutr. Sci. 2017, 6, e41. [Google Scholar] [CrossRef] [Green Version]
- Leopoldo, A.S.; Lima-Leopoldo, A.P.; Nascimento, A.F.; Luvizotto, R.A.; Sugizaki, M.M.; Campos, D.H.; da Silva, D.C.; Padovani, C.R.; Cicogna, A.C. Classification of different degrees of adiposity in sedentary rats. Braz. J. Med. Biol. Res. 2016, 49, e5028. [Google Scholar] [CrossRef] [Green Version]
- Arantes-Rodrigues, R.; Henriques, A.; Pires, M.J.; Colaco, B.; Calado, A.M.; Rema, P.; Colaco, A.; Fernandes, T.; De la Cruz, P.L.; Lopes, C.; et al. High doses of olive leaf extract induce liver changes in mice. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2011, 49, 1989–1997. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.R.; Begum, S.; Oh, D.S.; Wee, A.J.; Yun, B.S.; Sung, C.K. Ameliorating Effect of Mycoleptodonoides aitchisonii on High-fat Diet-induced Obese Mice. Prev. Nutr. Food Sci. 2014, 19, 69–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.O. Determination of the surface area of the white rat with its application to the expression of metabolic results. Am. J. Physiol.-Leg. Content 1929, 89, 24–33. [Google Scholar] [CrossRef]
- Simson, E.L.; Gold, R.M. The lee obesity index vindicated? Physiol. Behav. 1982, 29, 371–376. [Google Scholar] [CrossRef]
- Ayala, J.E.; Samuel, V.T.; Morton, G.J.; Obici, S.; Croniger, C.M.; Shulman, G.I.; Wasserman, D.H.; McGuinness, O.P.; Consortium, N.I.H.M.M.P.C. Standard operating procedures for describing and performing metabolic tests of glucose homeostasis in mice. Dis. Models Mech. 2010, 3, 525–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benede-Ubieto, R.; Estevez-Vazquez, O.; Ramadori, P.; Cubero, F.J.; Nevzorova, Y.A. Guidelines and Considerations for Metabolic Tolerance Tests in Mice. Diabetes Metab. Syndr. Obes. 2020, 13, 439–450. [Google Scholar] [CrossRef] [Green Version]
- Bowe, J.E.; Franklin, Z.J.; Hauge-Evans, A.C.; King, A.J.; Persaud, S.J.; Jones, P.M. Metabolic phenotyping guidelines: Assessing glucose homeostasis in rodent models. J. Endocrinol. 2014, 222, G13–G25. [Google Scholar] [CrossRef]
- Jensen, T.L.; Kiersgaard, M.K.; Sorensen, D.B.; Mikkelsen, L.F. Fasting of mice: A review. Lab. Anim. 2013, 47, 225–240. [Google Scholar] [CrossRef]
- Nagy, C.; Einwallner, E. Study of In Vivo Glucose Metabolism in High-fat Diet-fed Mice Using Oral Glucose Tolerance Test (OGTT) and Insulin Tolerance Test (ITT). J. Vis. Exp. 2018, 7, 56672. [Google Scholar] [CrossRef]
- Parasuraman, S.; Raveendran, R. Measurement of invasive blood pressure in rats. J. Pharm. Pharm. 2012, 3, 172–177. [Google Scholar] [CrossRef]
- Plehm, R.; Barbosa, M.E.; Bader, M. Animal models for hypertension/blood pressure recording. In Cardiovascular Disease, 2006/11/07 ed.; Wang, Q., Ed.; Methods in Molecular Medicine; Humana Press: Totowa, NJ, USA, 2006; Volume 129, pp. 115–126. [Google Scholar]
- Sikaris, K.A. The clinical biochemistry of obesity. Clin. Biochem. Rev. 2004, 25, 165–181. [Google Scholar] [PubMed]
Figure 1.
Risk factors of obesity.

Figure 2.
Potential rodent models that can be used to study obesity.


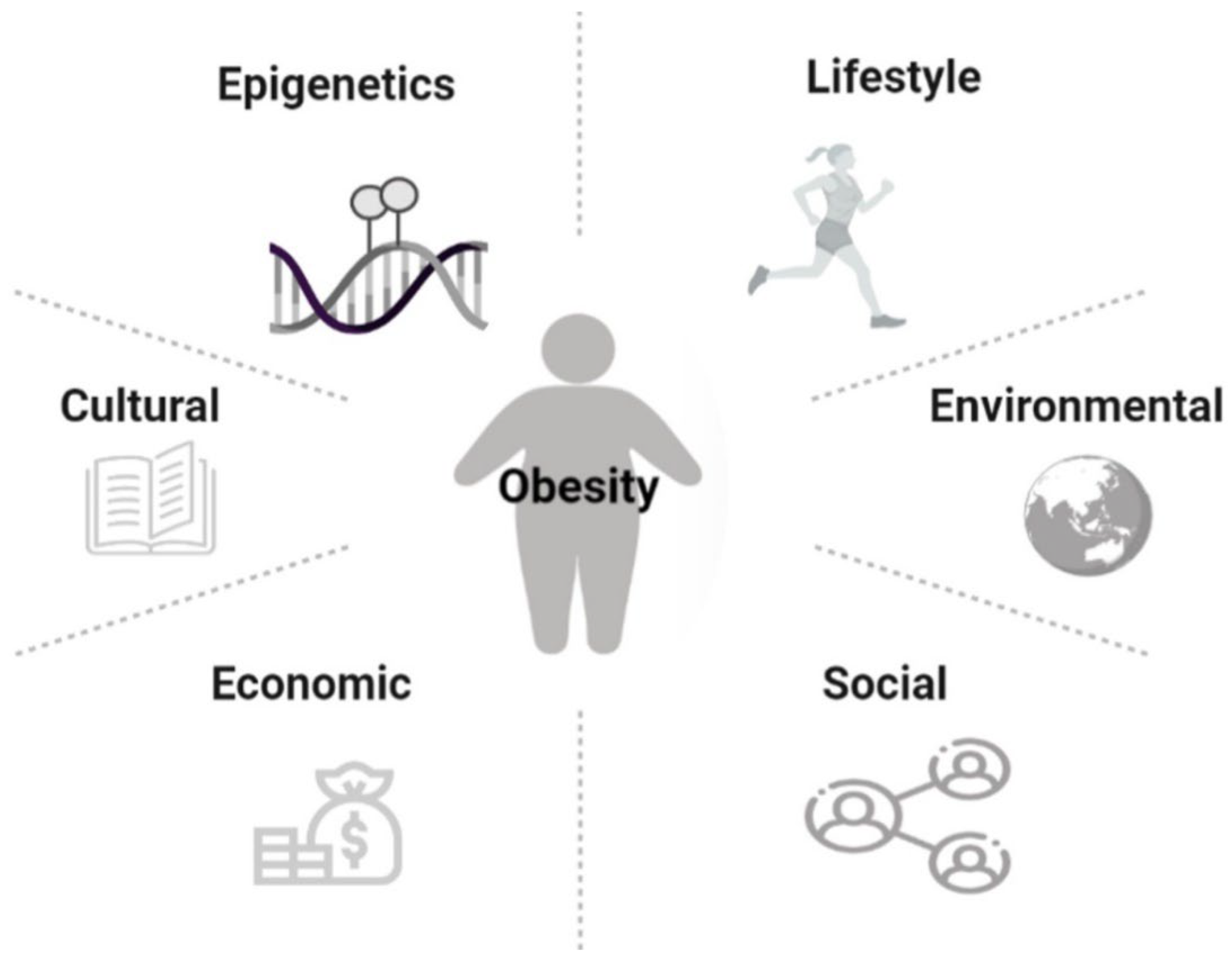{kind=link}
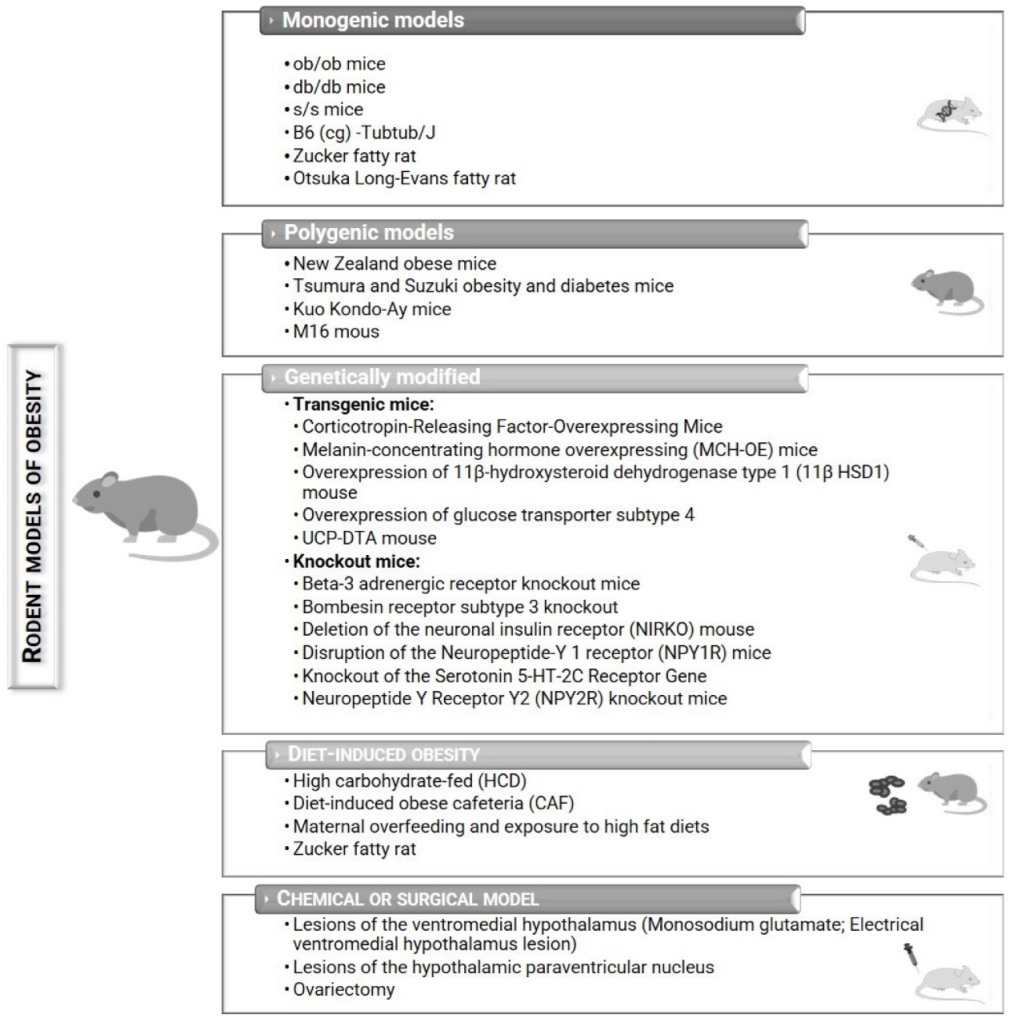{kind=link}
Table 1.
Main advantages and disadvantages of murine monogenic, polygenic, genetically modified, diet-induced, chemically induced, and surgically induced models of obesity.
Table 1.
Main advantages and disadvantages of murine monogenic, polygenic, genetically modified, diet-induced, chemically induced, and surgically induced models of obesity.
| Models | Advantages | Disadvantages |
|---|---|---|
| Monogenic |
|
|
| Polygenic |
|
|
| Genetically modified |
|
|
| Diet-induced |
|
|
| Chemical or surgical |
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Martins, T.; Castro-Ribeiro, C.; Lemos, S.; Ferreira, T.; Nascimento-Gonçalves, E.; Rosa, E.; Oliveira, P.A.; Antunes, L.M. Murine Models of Obesity. Obesities 2022, 2, 127-147. https://doi.org/10.3390/obesities2020012
AMA Style
Martins T, Castro-Ribeiro C, Lemos S, Ferreira T, Nascimento-Gonçalves E, Rosa E, Oliveira PA, Antunes LM. Murine Models of Obesity. Obesities. 2022; 2(2):127-147. https://doi.org/10.3390/obesities2020012
Chicago/Turabian StyleMartins, Tânia, Catarina Castro-Ribeiro, Sílvia Lemos, Tiago Ferreira, Elisabete Nascimento-Gonçalves, Eduardo Rosa, Paula Alexandra Oliveira, and Luís Miguel Antunes. 2022. "Murine Models of Obesity" Obesities 2, no. 2: 127-147. https://doi.org/10.3390/obesities2020012