
In Vitro and In Vivo Antihypertensive Effect of Milk Fermented with Different Strains of Common Starter Lactic Acid Bacteria

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Cultivation Conditions
2.2. Proteolytic, Antioxidant, and Angiotensin-I-Converting Enzyme Inhibitory Activities
2.3. Fatty Acid Profile of Fermented Milk
2.4. Spontaneously Hypertensive Rat (SHR) Animal Model
2.5. Measurements of Blood Pressure
2.6. Measurments of Biochemical Parameters
2.7. Statistical Analysis
3. Results
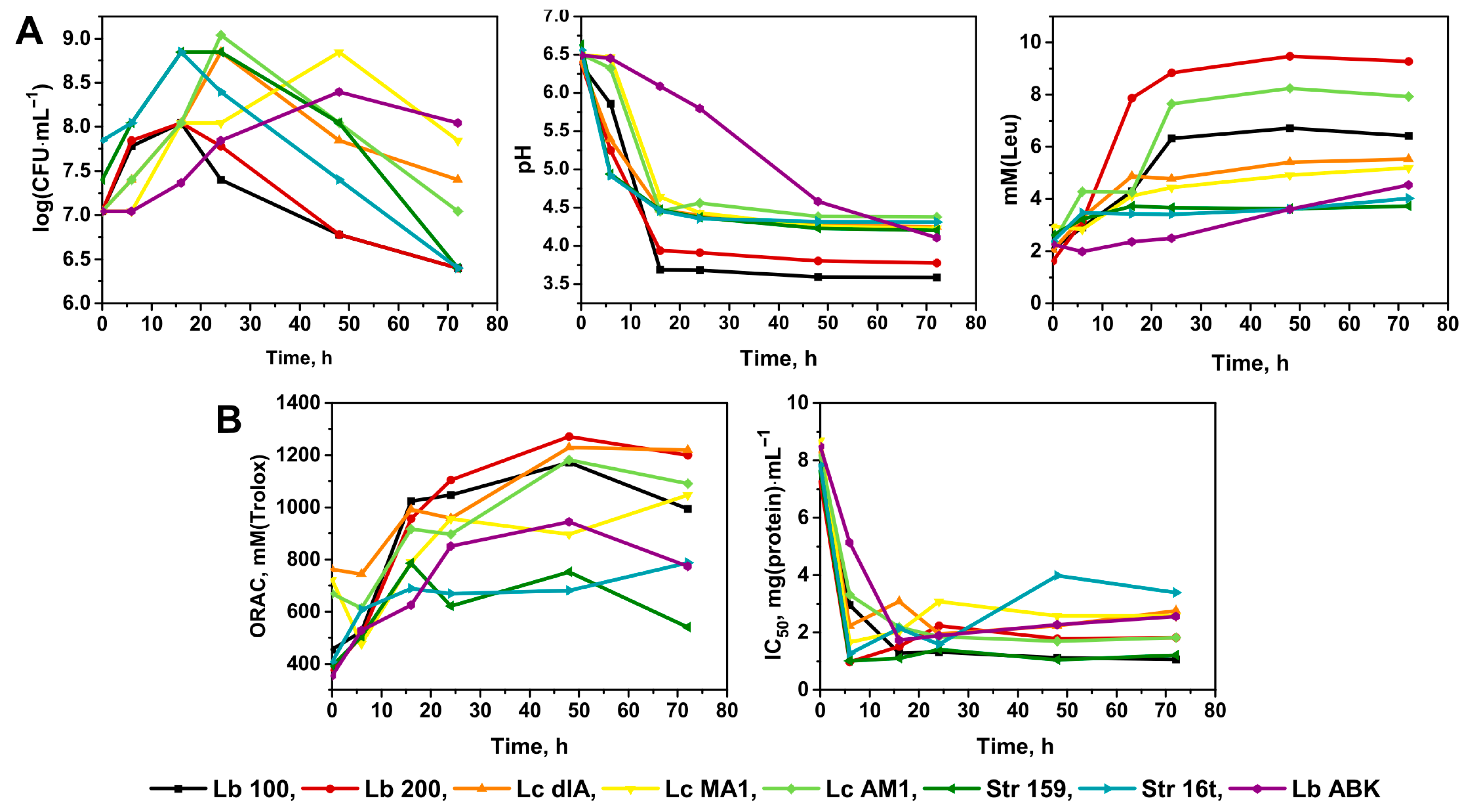
3.1. The Fermentation Perfomance of Different Strains of Lactic Acid Bacteria (LAB): Selection of the Most Promissing, in Terms of the In Vitro Antioxidant and Antihypertensive Properties of the Fermented Milk, LAB Strains
3.2. Profile of Fatty Acids (FA) and FA Nutritional Indices for the Milk Fermented by the Selected LAB Strains
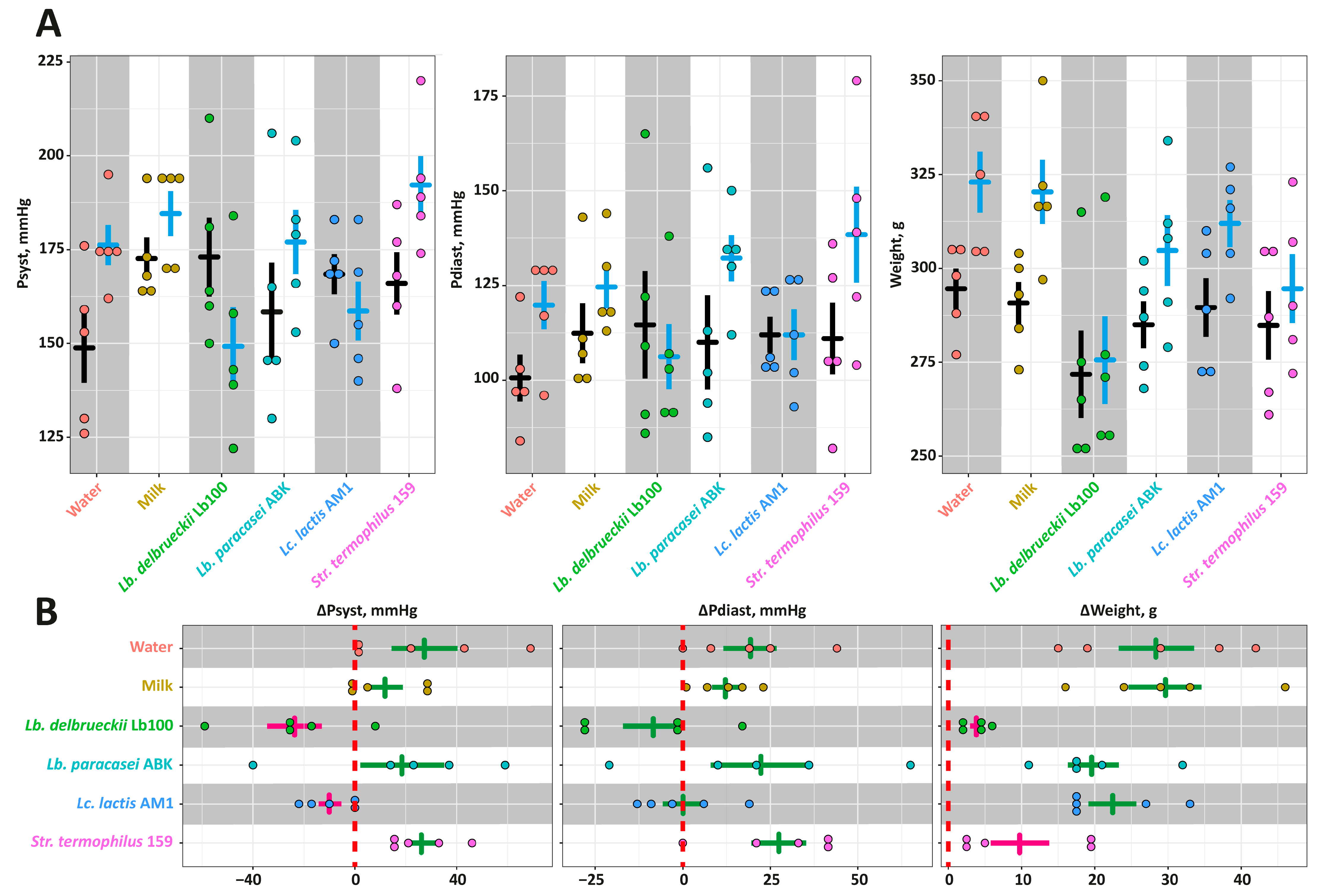
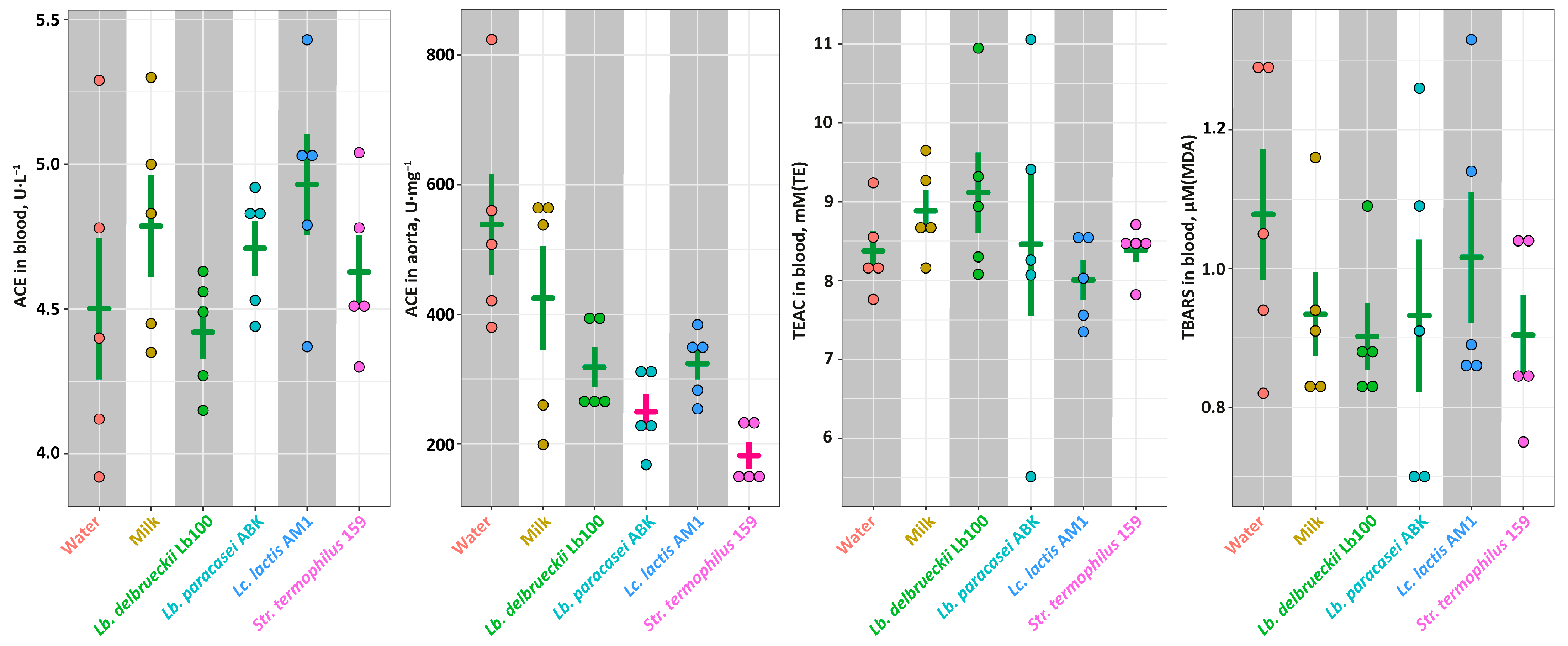
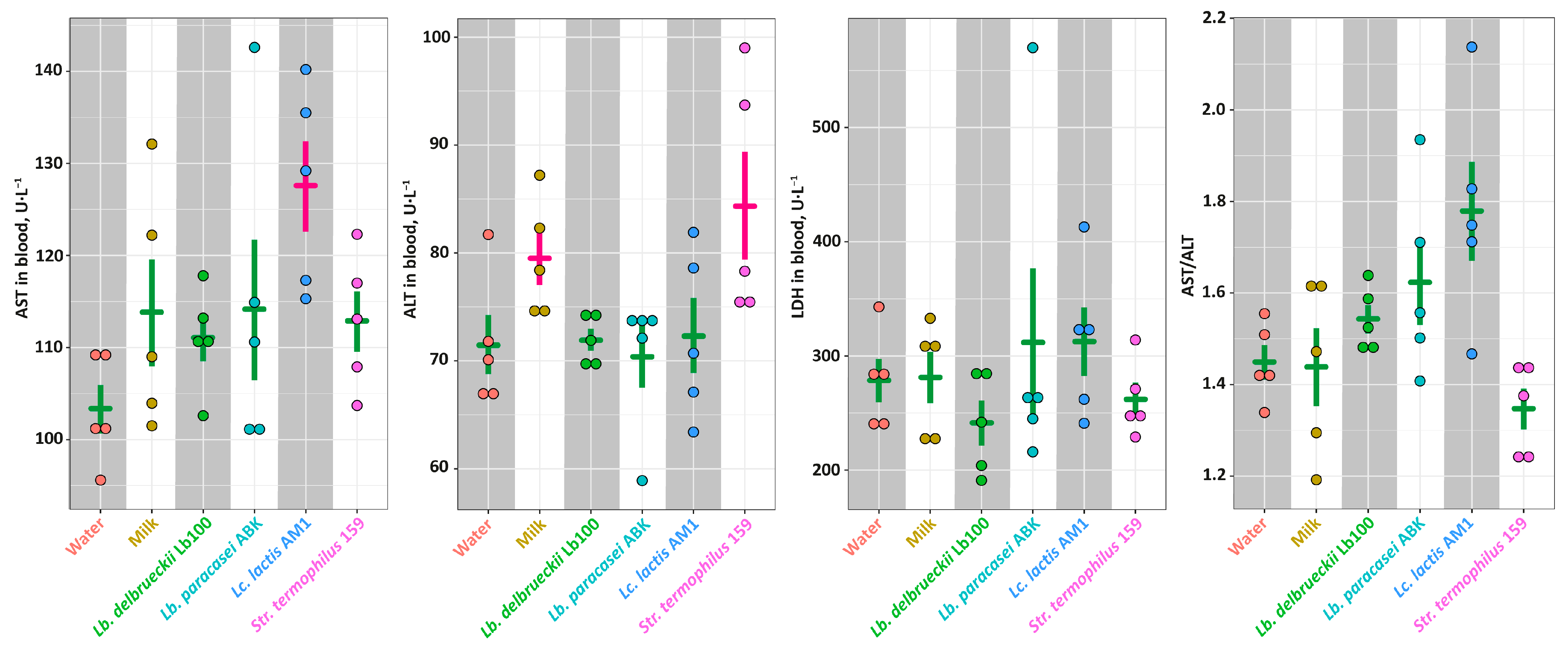
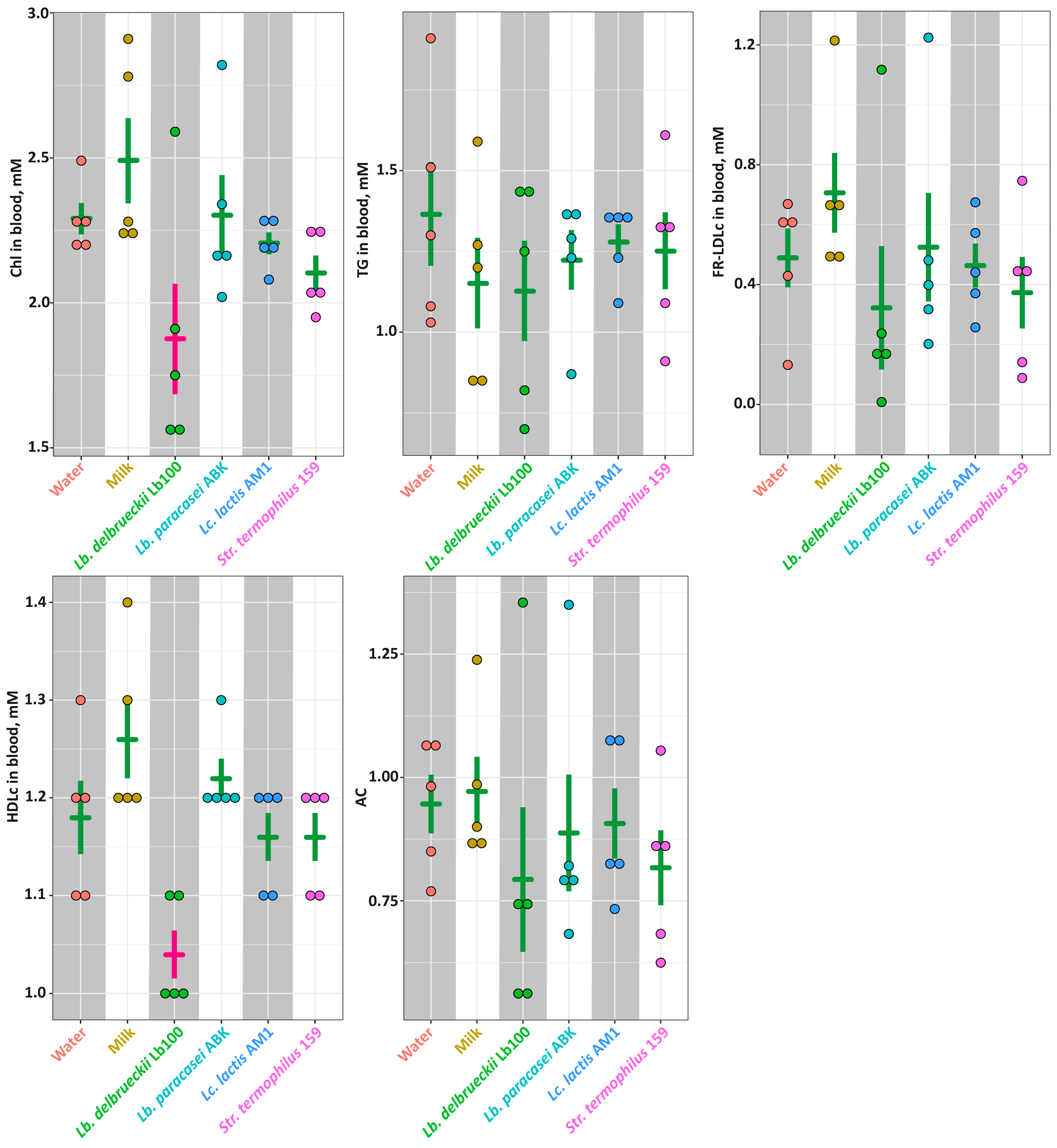
3.3. The In Vivo Assessment of the Antihypertensive Properties for the Milk Fermented by the Selected LAB Strains in the SHR Animal Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhou, B.; Carrillo-Larco, R.M.; Danaei, G.; Riley, L.M.; Paciorek, C.J.; Stevens, G.A.; Gregg, E.W.; Bennett, J.E.; Solomon, B.; Singleton, R.K.; et al. Worldwide trends in hypertension prevalence and progress in treatment and control from 1990 to 2019: A pooled analysis of 1201 population-representative studies with 104 million participants. Lancet 2021, 398, 957–980. [Google Scholar] [CrossRef] [PubMed]
- Brouwers, S.; Sudano, I.; Kokubo, Y.; Sulaica, E.M. Arterial hypertension. Lancet 2021, 398, 249–261. [Google Scholar] [CrossRef]
- Riccio, E.; Capuano, I.; Buonanno, P.; Andreucci, M.; Provenzano, M.; Amicone, M.; Rizzo, M.; Pisani, A. RAAS Inhibitor Prescription and Hyperkalemia Event in Patients with Chronic Kidney Disease: A Single-Center Retrospective Study. Front. Cardiovasc. Med. 2022, 9, 74. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, R. RAAS inhibition and mortality in hypertension. Glob. Cardiol. Sci. Pract. 2013, 2013, 34. [Google Scholar] [CrossRef] [PubMed]
- Ghazi, L.; Drawz, P. Advances in understanding the renin-angiotensin-aldosterone system (RAAS) in blood pressure control and recent pivotal trials of RAAS blockade in heart failure and diabetic nephropathy. F1000Research 2017, 6, 297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steckelings, U.M.; Rompe, F.; Kaschina, E.; Unger, T. The evolving story of the RAAS in hypertension, diabetes and CV disease—moving from macrovascular to microvascular targets. Fundam. Clin. Pharmacol. 2009, 23, 693–703. [Google Scholar] [CrossRef]
- te Riet, L.; van Esch, J.H.M.; Roks, A.J.M.; van den Meiracker, A.H.; Danser, A.H.J. Hypertension. Circ. Res. 2015, 116, 960–975. [Google Scholar] [CrossRef] [Green Version]
- Aprotosoaie, A.C.; Costache, A.-D.; Costache, I.-I. Therapeutic Strategies and Chemoprevention of Atherosclerosis: What Do We Know and Where Do We Go? Pharmaceutics 2022, 14, 722. [Google Scholar] [CrossRef]
- Xia, Y.; Yu, J.; Xu, W. Purification and characterization of angiotensin-I-converting enzyme inhibitory peptides isolated from whey proteins of milk fermented with Lactobacillus plantarum QS670. J. Dairy Sci. 2020, 103, 4919–4928. [Google Scholar] [CrossRef]
- Jogi, N.; Yathisha, U.G.; Bhat, I.; Mamatha, B.S. Antihypertensive activity of orally consumed ACE-I inhibitory peptides. Crit. Rev. Food Sci. Nutr. 2022, 62, 8986–8999. [Google Scholar] [CrossRef]
- Das, S.; Hati, S. Food derived ACE inhibitory peptides. In Nutrition and Functional Foods in Boosting Digestion, Metabolism and Immune Health; Elsevier: Amsterdam, The Netherlands, 2022; pp. 39–54. [Google Scholar]
- García-Burgos, M.; Moreno-Fernández, J.; Alférez, M.J.M.; Díaz-Castro, J.; López-Aliaga, I. New perspectives in fermented dairy products and their health relevance. J. Funct. Foods 2020, 72, 104059. [Google Scholar] [CrossRef]
- Ghavami, A.; Ziaei, R.; Moradi, S.; Sharifi, S.; Reza Moravejolahkami, A.; Ghaffari, S.; Irandoost, P.; Khorvash, F.; Mokari_yamchi, A.; Nattagh-Eshtivani, E.; et al. Potential of favorable effects of probiotics fermented milk supplementation on blood pressure: A systematic review and meta-analysis. Int. J. Food Prop. 2020, 23, 1925–1940. [Google Scholar] [CrossRef]
- Martin, M.; Deussen, A. Effects of natural peptides from food proteins on angiotensin converting enzyme activity and hypertension. Crit. Rev. Food Sci. Nutr. 2019, 59, 1264–1283. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, A.; Shah, N.P. Cell growth and proteolytic activity of Lactobacillus acidophilus, Lactobacillus helveticus, Lactobacillus delbrueckii ssp. bulgaricus, and Streptococcus thermophilus in milk as affected by supplementation with peptide fractions. Int. J. Food Sci. Nutr. 2014, 65, 937–941. [Google Scholar] [CrossRef] [PubMed]
- Ji, D.; Ma, J.; Xu, M.; Agyei, D. Cell-envelope proteinases from lactic acid bacteria: Biochemical features and biotechnological applications. Compr. Rev. Food Sci. Food Saf. 2021, 20, 369–400. [Google Scholar] [CrossRef]
- Szliszka, E.; Czuba, Z.P.; Domino, M.; Mazur, B.; Zydowicz, G.; Krol, W. Ethanolic Extract of Propolis (EEP) Enhances the Apoptosis- Inducing Potential of TRAIL in Cancer Cells. Molecules 2009, 14, 738–754. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.K.; Sanjukta, S.; Jeyaram, K. Production of angiotensin I converting enzyme inhibitory (ACE-I) peptides during milk fermentation and their role in reducing hypertension. Crit. Rev. Food Sci. Nutr. 2017, 57, 2789–2800. [Google Scholar] [CrossRef] [PubMed]
- Beltrán-Barrientos, L.M.; Hernández-Mendoza, A.; Torres-Llanez, M.J.; González-Córdova, A.F.; Vallejo-Córdoba, B. Invited review: Fermented milk as antihypertensive functional food. J. Dairy Sci. 2016, 99, 4099–4110. [Google Scholar] [CrossRef] [Green Version]
- Udenigwe, C.C.; Mohan, A. Mechanisms of food protein-derived antihypertensive peptides other than ACE inhibition. J. Funct. Foods 2014, 8, 45–52. [Google Scholar] [CrossRef]
- Mell, B.; Jala, V.R.; Mathew, A.V.; Byun, J.; Waghulde, H.; Zhang, Y.; Haribabu, B.; Vijay-Kumar, M.; Pennathur, S.; Joe, B. Evidence for a link between gut microbiota and hypertension in the Dahl rat. Physiol. Genom. 2015, 47, 187–197. [Google Scholar] [CrossRef]
- Yang, T.; Santisteban, M.M.; Rodriguez, V.; Li, E.; Ahmari, N.; Carvajal, J.M.; Zadeh, M.; Gong, M.; Qi, Y.; Zubcevic, J.; et al. Gut Dysbiosis Is Linked to Hypertension. Hypertension 2015, 65, 1331–1340. [Google Scholar] [CrossRef] [Green Version]
- Adnan, S.; Nelson, J.W.; Ajami, N.J.; Venna, V.R.; Petrosino, J.F.; Bryan, R.M.; Durgan, D.J. Alterations in the gut microbiota can elicit hypertension in rats. Physiol. Genom. 2017, 49, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Toral, M.; Robles-Vera, I.; Visitación, N.; Romero, M.; Sánchez, M.; Gómez-Guzmán, M.; Rodriguez-Nogales, A.; Yang, T.; Jiménez, R.; Algieri, F.; et al. Role of the immune system in vascular function and blood pressure control induced by faecal microbiota transplantation in rats. Acta Physiol. 2019, 227, e13285. [Google Scholar] [CrossRef] [PubMed]
- Beltrán-Barrientos, L.M.; García, H.S.; Hernández-Mendoza, A.; González-Córdova, A.F.; Vallejo-Cordoba, B. Invited review: Effect of antihypertensive fermented milks on gut microbiota. J. Dairy Sci. 2021, 104, 3779–3788. [Google Scholar] [CrossRef]
- Paszczyk, B.; Tońska, E. Fatty Acid Content, Lipid Quality Indices, and Mineral Composition of Cow Milk and Yogurts Produced with Different Starter Cultures Enriched with Bifidobacterium bifidum. Appl. Sci. 2022, 12, 6558. [Google Scholar] [CrossRef]
- Adler-Nissen, J. Determination of the degree of hydrolysis of food protein hydrolysates by trinitrobenzenesulfonic acid. J. Agric. Food Chem. 1979, 27, 1256–1262. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and Validation of an Improved Oxygen Radical Absorbance Capacity Assay Using Fluorescein as the Fluorescent Probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef]
- Nikolaev, I.V.; Sforza, S.; Lambertini, F.; Ismailova, D.Y.; Khotchenkov, V.P.; Volik, V.G.; Dossena, A.; Popov, V.O.; Koroleva, O.V. Biocatalytic conversion of poultry processing leftovers: Optimization of hydrolytic conditions and peptide hydrolysate characterization. Food Chem. 2016, 197, 611–621. [Google Scholar] [CrossRef]
- Torkova, A.A.; Ryazantseva, K.A.; Agarkova, E.Y.; Kruchinin, A.G.; Tsentalovich, M.Y.; Fedorova, T.V. Rational design of enzyme compositions for the production of functional hydrolysates of cow milk whey proteins. Appl. Biochem. Microbiol. 2017, 53, 669–679. [Google Scholar] [CrossRef]
- Moiseenko, K.V.; Glazunova, O.A.; Savinova, O.S.; Ajibade, B.O.; Ijabadeniyi, O.A.; Fedorova, T.V. Analytical Characterization of the Widely Consumed Commercialized Fermented Beverages from Russia (Kefir and Ryazhenka) and South Africa (Amasi and Mahewu): Potential Functional Properties and Profiles of Volatile Organic Compounds. Foods 2021, 10, 3082. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liu, H. Nutritional Indices for Assessing Fatty Acids: A Mini-Review. Int. J. Mol. Sci. 2020, 21, 5695. [Google Scholar] [CrossRef] [PubMed]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Kruchinin, A.G.; Savinova, O.S.; Glazunova, O.A.; Moiseenko, K.V.; Agarkova, E.Y.; Fedorova, T.V. Hypotensive and Hepatoprotective Properties of the Polysaccharide-Stabilized Foaming Composition Containing Hydrolysate of Whey Proteins. Nutrients 2021, 13, 1031. [Google Scholar] [CrossRef]
- Jentzsch, A.M.; Bachmann, H.; Fürst, P.; Biesalski, H.K. Improved analysis of malondialdehyde in human body fluids. Free Radic. Biol. Med. 1996, 20, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Begunova, A.V.; Savinova, O.S.; Glazunova, O.A.; Moiseenko, K.V.; Rozhkova, I.V.; Fedorova, T.V. Development of Antioxidant and Antihypertensive Properties during Growth of Lactobacillus helveticus, Lactobacillus rhamnosus and Lactobacillus reuteri on Cow’s Milk: Fermentation and Peptidomics Study. Foods 2020, 10, 17. [Google Scholar] [CrossRef]
- Sarmadi, B.H.; Ismail, A. Antioxidative peptides from food proteins: A review. Peptides 2010, 31, 1949–1956. [Google Scholar] [CrossRef]
- Pihlanto, A. Antioxidative peptides derived from milk proteins. Int. Dairy J. 2006, 16, 1306–1314. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Aluko, R.E. Food Protein-Derived Bioactive Peptides: Production, Processing, and Potential Health Benefits. J. Food Sci. 2012, 77, R11–R24. [Google Scholar] [CrossRef]
- Gómez-Cortés, P.; Juárez, M.; de la Fuente, M.A. Milk fatty acids and potential health benefits: An updated vision. Trends Food Sci. Technol. 2018, 81, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Calder, P.C. Functional Roles of Fatty Acids and Their Effects on Human Health. J. Parenter. Enter. Nutr. 2015, 39, 18S–32S. [Google Scholar] [CrossRef]
- Williams, C.M. Dietary fatty acids and human health. Ann. Zootech. 2000, 49, 165–180. [Google Scholar] [CrossRef] [Green Version]
- Lerman, L.O.; Kurtz, T.W.; Touyz, R.M.; Ellison, D.H.; Chade, A.R.; Crowley, S.D.; Mattson, D.L.; Mullins, J.J.; Osborn, J.; Eirin, A.; et al. Animal Models of Hypertension: A Scientific Statement From the American Heart Association. Hypertension 2019, 73, e87–e120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerman, L.O.; Chade, A.R.; Sica, V.; Napoli, C. Animal models of hypertension: An overview. J. Lab. Clin. Med. 2005, 146, 160–173. [Google Scholar] [CrossRef]
- Pinto, Y. Lessons from rat models of hypertension from Goldblatt to genetic engineering. Cardiovasc. Res. 1998, 39, 77–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvathova, M.; Zitnanova, I.; Kralovicova, Z.; Balis, P.; Puzserova, A.; Muchova, J.; Kluknavsky, M.; Durackova, Z.; Bernatova, I. Sex differences in the blood antioxidant defense system in juvenile rats with various genetic predispositions to hypertension. Hypertens. Res. 2016, 39, 64–69. [Google Scholar] [CrossRef]
- Nimmanapalli, H.; Kasi, A.; Devapatla, P.; Nuttakki, V. Lipid ratios, atherogenic coefficient and atherogenic index of plasma as parameters in assessing cardiovascular risk in type 2 diabetes mellitus. Int. J. Res. Med. Sci. 2016, 4, 2863–2869. [Google Scholar] [CrossRef] [Green Version]
- Mills, K.T.; Stefanescu, A.; He, J. The global epidemiology of hypertension. Nat. Rev. Nephrol. 2020, 16, 223–237. [Google Scholar] [CrossRef]
- González, S.; Fernández-Navarro, T.; Arboleya, S.; de los Reyes-Gavilán, C.G.; Salazar, N.; Gueimonde, M. Fermented Dairy Foods: Impact on Intestinal Microbiota and Health-Linked Biomarkers. Front. Microbiol. 2019, 10, 1046. [Google Scholar] [CrossRef]
- Ramchandran, L.; Shah, N.P. Proteolytic Profiles and Angiotensin-I Converting Enzyme and α-Glucosidase Inhibitory Activities of Selected Lactic Acid Bacteria. J. Food Sci. 2008, 73, M75–M81. [Google Scholar] [CrossRef]
- Nejati, F.; Rizzello, C.G.; Di Cagno, R.; Sheikh-Zeinoddin, M.; Diviccaro, A.; Minervini, F.; Gobbetti, M. Manufacture of a functional fermented milk enriched of Angiotensin-I Converting Enzyme (ACE)-inhibitory peptides and γ-amino butyric acid (GABA). LWT-Food Sci. Technol. 2013, 51, 183–189. [Google Scholar] [CrossRef]
- Loghman, S.; Moayedi, A.; Mahmoudi, M.; Khomeiri, M.; Gómez-Mascaraque, L.G.; Garavand, F. Single and Co-Cultures of Proteolytic Lactic Acid Bacteria in the Manufacture of Fermented Milk with High ACE Inhibitory and Antioxidant Activities. Fermentation 2022, 8, 448. [Google Scholar] [CrossRef]
- Gobbetti, M.; Ferranti, P.; Smacchi, E.; Goffredi, F.; Addeo, F. Production of Angiotensin-I-Converting-Enzyme-Inhibitory Peptides in Fermented Milks Started by Lactobacillus delbrueckii subsp. bulgaricus SS1 and Lactococcus lactis subsp. cremoris FT4. Appl. Environ. Microbiol. 2000, 66, 3898–3904. [Google Scholar] [CrossRef] [Green Version]
- Pihlanto, A.; Virtanen, T.; Korhonen, H. Angiotensin I converting enzyme (ACE) inhibitory activity and antihypertensive effect of fermented milk. Int. Dairy J. 2010, 20, 3–10. [Google Scholar] [CrossRef]
- Hagi, T.; Kobayashi, M.; Nomura, M. Metabolome analysis of milk fermented by γ-aminobutyric acid–producing Lactococcus lactis. J. Dairy Sci. 2016, 99, 994–1001. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Masuda, O.; Takano, T. Decrease of Tissue Angiotensin I-Converting Enzyme Activity upon Feeding Sour Milk in Spontaneously Hypertensive Rats. Biosci. Biotechnol. Biochem. 1996, 60, 488–489. [Google Scholar] [CrossRef]
- Kim, S.M.; Park, S.; Choue, R. Effects of fermented milk peptides supplement on blood pressure and vascular function in spontaneously hypertensive rats. Food Sci. Biotechnol. 2010, 19, 1409–1413. [Google Scholar] [CrossRef]
- Ramchandran, L.; Shah, N.P. Yogurt Can Beneficially Affect Blood Contributors of Cardiovascular Health Status in Hypertensive Rats. J. Food Sci. 2011, 76, H131–H136. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.-Y.; Li, Z.-M.; Mao, Y.-Q.; Chen, H.-L.; Hu, W.; Han, B.; Wang, L.-S. Probiotic yogurt blunts the increase of blood pressure in spontaneously hypertensive rats via remodeling of the gut microbiota. Food Funct. 2021, 12, 9773–9783. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, W.; Xue, J.; Yang, J.; Chen, X.; Shao, Y.; Kwok, L.; Bilige, M.; Mang, L.; Zhang, H. Angiotensin-converting enzyme inhibitory activity of Lactobacillus helveticus strains from traditional fermented dairy foods and antihypertensive effect of fermented milk of strain H9. J. Dairy Sci. 2014, 97, 6680–6692. [Google Scholar] [CrossRef] [Green Version]
- Sipola, M.; Finckenberg, P.; Korpela, R.; Vapaatalo, H.; Nurminen, M.L. Effect of long-term intake of milk products on blood pressure in hypertensive rats. J. Dairy Res. 2002, 69, 103–111. [Google Scholar] [CrossRef]
- Jauhiainen, T.; Collin, M.; Narva, M.; Cheng, Z.J.; Poussa, T.; Vapaatalo, H.; Korpela, R. Effect of long-term intake of milk peptides and minerals on blood pressure and arterial function in spontaneously hypertensive rats. Milchwissenschaft 2005, 60, 358–363. [Google Scholar]
- Yamamoto, N.; Akino, A.; Takano, T. Antihypertensive Effects of Different Kinds of Fermented Milk in Spontaneously Hypertensive Rats. Biosci. Biotechnol. Biochem. 1994, 58, 776–778. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Figueroa, J.C.; González-Córdova, A.F.; Astiazaran-García, H.; Hernández-Mendoza, A.; Vallejo-Cordoba, B. Antihypertensive and hypolipidemic effect of milk fermented by specific Lactococcus lactis strains. J. Dairy Sci. 2013, 96, 4094–4099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehlers, P.I.; Kivimäki, A.S.; Turpeinen, A.M.; Korpela, R.; Vapaatalo, H. High blood pressure-lowering and vasoprotective effects of milk products in experimental hypertension. Br. J. Nutr. 2011, 106, 1353–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halperin, R.O.; Sesso, H.D.; Ma, J.; Buring, J.E.; Stampfer, M.J.; Gaziano, J.M. Dyslipidemia and the Risk of Incident Hypertension in Men. Hypertension 2006, 47, 45–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuwabara, M.; Kuwabara, R.; Niwa, K.; Hisatome, I.; Smits, G.; Roncal-Jimenez, C.; MacLean, P.; Yracheta, J.; Ohno, M.; Lanaspa, M.; et al. Different Risk for Hypertension, Diabetes, Dyslipidemia, and Hyperuricemia According to Level of Body Mass Index in Japanese and American Subjects. Nutrients 2018, 10, 1011. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Zhang, Y.; Guo, Z.; Yang, H.; Ren, M.; Xing, X.; Cong, H. The association of triglyceride and glucose index, and triglyceride to high-density lipoprotein cholesterol ratio with prehypertension and hypertension in normoglycemic subjects: A large cross-sectional population study. J. Clin. Hypertens. 2021, 23, 1405–1412. [Google Scholar] [CrossRef]
- Oparil, S.; Zaman, M.A.; Calhoun, D.A. Pathogenesis of Hypertension. Ann. Intern. Med. 2003, 139, 761. [Google Scholar] [CrossRef]
- Lye, H.; Kuan, C.; Ewe, J.; Fung, W.; Liong, M. The Improvement of Hypertension by Probiotics: Effects on Cholesterol, Diabetes, Renin, and Phytoestrogens. Int. J. Mol. Sci. 2009, 10, 3755–3775. [Google Scholar] [CrossRef]
- Silva Figueiredo, P.; Carla Inada, A.; Marcelino, G.; Maiara Lopes Cardozo, C.; de Cássia Freitas, K.; de Cássia Avellaneda Guimarães, R.; Pereira de Castro, A.; Aragão do Nascimento, V.; Aiko Hiane, P. Fatty Acids Consumption: The Role Metabolic Aspects Involved in Obesity and Its Associated Disorders. Nutrients 2017, 9, 1158. [Google Scholar] [CrossRef] [Green Version]
- Baum, S.J.; Kris-Etherton, P.M.; Willett, W.C.; Lichtenstein, A.H.; Rudel, L.L.; Maki, K.C.; Whelan, J.; Ramsden, C.E.; Block, R.C. Fatty acids in cardiovascular health and disease: A comprehensive update. J. Clin. Lipidol. 2012, 6, 216–234. [Google Scholar] [CrossRef] [PubMed]
- Kuller, L.H. Dietary Fat and Chronic Diseases. J. Am. Diet. Assoc. 1997, 97, S9–S15. [Google Scholar] [CrossRef] [PubMed]
- Visioli, F.; Poli, A. Fatty Acids and Cardiovascular Risk. Evidence, Lack of Evidence, and Diligence. Nutrients 2020, 12, 3782. [Google Scholar] [CrossRef] [PubMed]
- Balta, I.; Stef, L.; Pet, I.; Iancu, T.; Stef, D.; Corcionivoschi, N. Essential Fatty Acids as Biomedicines in Cardiac Health. Biomedicines 2021, 9, 1466. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | 16S rRNA GeneBank Accession | Optimal Growth Temperature |
|---|---|---|
| Lactobacillus delbrueckii | ||
| Lb100 | MN994622 | 37 °C |
| Lb200 | MN994623 | |
| Lacticaseibacillus paracasei | ||
| ABK | MN994625 | 30 °C |
| Lactococcus lactis | ||
| dlA | MN994624 | 30 °C |
| AM1 | MW558124 | |
| MA1 | MW558123 | |
| Streptococcus thermophilus | ||
| 159 | MN994626 | 37 °C |
| 16t | MN994627 | |
| Fatty Acid | Relative Abundance, % | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Name | Abbreviation | Lb. delbrueckii Lb100 | Lb. paracasei ABK | Lc. lactis AM1 | Str. termophilus 159 | ||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | ||
| Saturated Fatty Acids (SFA) | |||||||||
| Pentanoic acid | C5:0 | ND | - | 0.03 | 0.01 | 0.03 | 0.01 | ND | - |
| Hexanoic acid | C6:0 | 5.06 a | 0.32 | 2.99 b | 0.14 | 3.29 b | 0.23 | 3.42 b | 0.13 |
| Heptanoic acid | C7:0 | 0.16 | 0.08 | 0.09 | 0.13 | 0.11 | 0.03 | ND | - |
| Octanoic acid | C8:0 | 8.09 a | 0.16 | 4.84 b | 0.17 | 5.53 b | 0.39 | 6.38 b | 0.09 |
| Nonanoic acid | C9:0 | 0.32 | 0.03 | 0.28 | 0.03 | 0.22 | 0.08 | 0.24 | 0.08 |
| Decanoic acid | C10:0 | 10.36 a | 1.12 | 6.49 b | 1.01 | 7.43 b | 0.91 | 8.11 b | 1.80 |
| Undecanoic acid | C11:0 | ND | - | 0.17 | 0.07 | 0.23 | 0.05 | 0.21 | 0.15 |
| Dodecanoic acid | C12:0 | 4.01 a | 0.95 | 6.06 b | 0.14 | 6.82 b | 0.23 | 6.54 b | 0.43 |
| Tridecanoic acid | C13:0 | 0.30 | 0.22 | 0.27 | 0.02 | 0.23 | 0.03 | ||
| Tetradecanoic acid | C14:0 | 6.15 a | 1.30 | 9.64 b | 0.42 | 8.82 b | 1.01 | 8.90 b | 0.18 |
| Pentadecanoic acid | C15:0 | 1.42 | 0.52 | 2.07 | 0.68 | 2.10 | 0.23 | 2.16 | 0.25 |
| Hexadecanoic acid | C16:0 | 15.96 | 1.80 | 14.77 | 4.23 | 15.52 | 3.20 | 16.24 | 4.01 |
| Heptadecanoic acid | C17:0 | ND | - | 1.21 | 0.11 | 1.07 | 0.08 | 0.91 | 0.12 |
| Octadecanoic acid | C18:0 | 13.29 | 3.12 | 13.18 | 5.20 | 12.75 | 0.74 | 12.69 | 3.25 |
| Eicosanoic acid | C20:0 | 0.61 a | 0.18 | ND | - | 1.17 b | 0.01 | 0.88 a | 0.11 |
| Docosanoic acid | C22:0 | 1.09 a | 0.06 | 0.61 b | 0.14 | 0.64 b | 0.22 | ND | - |
| Tricosanoic acid | C23:0 | 0.78 a | 0.13 | ND | - | 0.44 b | 0.22 | ND | - |
| Tetracosanoic acid | C24:0 | 0.72 a | 0.07 | 0.28 b | 0.08 | 0.30 b | 0.08 | ND | - |
| Total SFA | 68.05 | 4.16 | 63.02 | 6.84 | 66.77 | 3.61 | 66.91 | 5.50 | |
| Monounsaturated fatty acids (MUFA) | |||||||||
| 4-Decenoic acid | C10:1 (n-6) | 2.45 a | 0.33 | 1.48 b | 0.21 | 1.81 b | 0.41 | 1.99 b | 0.22 |
| Dodecenoic acid | C12:1 (n-10) | ND | - | 0.36 | 0.06 | 0.41 | 0.17 | 0.42 | 0.20 |
| 9-Tetradecenoic acid | C14:1 (n-5) | 0.41 a | 0.09 | 1.75 b | 0.73 | 1.13 b | 0.24 | 0.91 b | 0.11 |
| 9-Hexadecenoic acid | C16:1 (n-7) | 1.41 | 0.25 | 3.64 | 1.03 | 2.45 | 0.38 | 2.08 | 0.23 |
| 9-Octadecenoic acid | C18:1 (n-9) | 16.07 | 1.98 | 12.45 | 2.23 | 13.63 | 3.01 | 17.67 | 2.35 |
| 11-Octadecenoic acid | C18:1 (n-7) | 2.91 a | 0.63 | 9.19 b | 1.02 | 5.62 a | 2.01 | 3.59 a | 0.98 |
| 11-Eicosenoic acid | C20:1 (n-9) | ND | - | ND | - | 3.25 a | 1.02 | 0.78 b | 0.09 |
| Total MUFA | 23.25 a | 2.12 | 28.87 b | 2.77 | 28.28 b | 3.81 | 27.44 b | 2.58 | |
| Polyunsaturated fatty acids (PUFA) | |||||||||
| 9,12-Octadecadienoic acid | C18:2 (n-6) | 5.99 a | 1.22 | 4.17 b | 2.03 | 4.14 b | 1.56 | 3.35 b | 0.69 |
| 5,8,11,14-Eicosatetraenoic acid | C20:4 (n-6) | ND | - | ND | - | ND | - | 0.41 | 0.10 |
| 4,7,10,13,16,19-Docosahexaenoic acid | C22:6 (n-3) | ND | - | ND | - | 0.33 | 0.05 | ND | - |
| Total PUFA | 5.99 | 1.22 | 4.17 | 2.03 | 4.48 | 1.56 | 3.77 | 0.70 | |
| Branched chain fatty acids (BCFA) | |||||||||
| Tetradecanoic acid, 9-methyl | 9Me-C14:0 | ND | - | 0.63 | 0.11 | ND | - | 0.76 | 0.11 |
| Hexadecanoic acid, 15-methyl- | 15MeC16:0 (iso-C17:0) | ND | - | ND | - | ND | - | 0.46 | 0.15 |
| Hexadecanoic acid, 14-methyl- | 14MeC16:0 (anteiso-C17:0) | ND | - | ND | - | ND | - | 0.68 | 0.09 |
| Total BCFA | ND | - | 0.63 a | 0.11 | ND | - | 1.89 b | 0.21 | |
| Hydroxy saturated fatty acids (OH-SFA) | |||||||||
| Octanoic acid, 3-hydroxy- | 3OH-C8:0 | ND | - | 0.05 | 0.20 | ND | - | ND | - |
| Decanoic acid, 3-hydroxy- | 3OH-C10:0 | ND | - | 0.07 | 0.22 | ND | - | ND | - |
| Tetradecanoic acid, 3-hydroxy- | 3OH-C14:0 | ND | - | 0.22 | 0.11 | ND | - | ND | - |
| Octadecanoic acid, 10-hydroxy- | 10OH-C18:0 | ND | - | 2.39 | 1.03 | ND | - | ND | - |
| Total OH-SFA | ND | - | 2.73 | 1.08 | ND | - | ND | - | |
| 2-hydroxy branched chain fatty acids (2OH-BCFA) | |||||||||
| Butyric acid, 2-hydroxy-3-methyl- | 2OH-3MeC4:0 (2OH-iso-C5:0) | ND | - | 0.09 | 0.10 | 0.10 | 0.08 | ND | - |
| Pentanoic acid, 2-hydroxy-4-methyl- | 2OH-4MeC5:0 (2OH-iso-C6:0) | 1.80 a | 0.32 | 0.43 b | 0.13 | 0.20 b | 0.26 | ND | - |
| Pentanoic acid, 2-hydroxy-3-methyl- | 2OH-3MeC5:0 (2OH-anteiso-C6:0) | 0.91 a | 0.26 | 0.06 b | 0.02 | 0.18 b | 0.03 | ND | - |
| Total 2OH-BCFA | 2.72 a | 0.41 | 0.58 b | 0.17 | 0.48 b | 0.10 | ND | - | |
| Index | General Interpretation | Lb. delbrueckii Lb100 | Lb. paracasei ABK | Lc. lactis AM1 | Str. termophilus 159 | ||||
|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | ||
| PUFA/SFA | The higher—The better | 0.09 | 0.02 | 0.07 | 0.03 | 0.07 | 0.02 | 0.06 | 0.01 |
| IA | The lower—The better | 1.53 a | 0.15 | 1.80 b | 0.23 | 1.76 b | 0.24 | 1.87 b | 0.21 |
| HPI | The higher—The better | 0.66 a | 0.07 | 0.56 b | 0.07 | 0.57 b | 0.08 | 0.53 b | 0.06 |
| IT | The lower—The better | 2.42 | 0.20 | 2.28 | 0.27 | 2.15 | 0.26 | 2.42 | 0.22 |
| HH | The higher—The better | 0.84 a | 0.12 | 0.55 b | 0.12 | 0.58 b | 0.11 | 0.68 b | 0.09 |
| UI | The higher—The better | 35.22 | 2.45 | 37.21 | 3.43 | 38.56 | 4.12 | 35.79 | 2.67 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glazunova, O.A.; Moiseenko, K.V.; Savinova, O.S.; Fedorova, T.V. In Vitro and In Vivo Antihypertensive Effect of Milk Fermented with Different Strains of Common Starter Lactic Acid Bacteria. Nutrients 2022, 14, 5357. https://doi.org/10.3390/nu14245357
Glazunova OA, Moiseenko KV, Savinova OS, Fedorova TV. In Vitro and In Vivo Antihypertensive Effect of Milk Fermented with Different Strains of Common Starter Lactic Acid Bacteria. Nutrients. 2022; 14(24):5357. https://doi.org/10.3390/nu14245357
Chicago/Turabian StyleGlazunova, Olga A., Konstantin V. Moiseenko, Olga S. Savinova, and Tatyana V. Fedorova. 2022. "In Vitro and In Vivo Antihypertensive Effect of Milk Fermented with Different Strains of Common Starter Lactic Acid Bacteria" Nutrients 14, no. 24: 5357. https://doi.org/10.3390/nu14245357