
Oral Intake of Chicken Bone Collagen Peptides Anti-Skin Aging in Mice by Regulating Collagen Degradation and Synthesis, Inhibiting Inflammation and Activating Lysosomes

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials, Chemicals, and Animals
2.2. Collagen Preparation and Amino Acid Composition
2.3. Preparation and Molecular Weight Distribution of Collagen Peptides (CPs)
2.4. Animal Test
- (i).
- Normal group (N): UV unexposed; oral administration of 0.4 mL saline daily.
- (ii).
- Model group (M): UV exposed + D-galactose (0.2 mL); oral administration of 0.4 mL saline daily.
- (iii).
- Low dose collagen peptides group (LCPs): UV exposed + D-galactose (0.2 mL); oral administration of 0.4 mL CPs (dose: 200 mg·kg−1 body weight) daily.
- (iv).
- Medium dose collagen peptides group (MCPs): UV exposed + D-galactose; oral administration of 0.4 mL CPs (dose: 500 mg·kg−1 body weight) daily.
- (v).
- High dose collagen peptides group (HCPs): UV exposed + D-galactose; oral administration of 0.4 mL CPs (dose: 1000 mg·kg−1 body weight) daily.
2.5. Skin Moisture, Visceral Index, and Body Weight Gain
2.6. Oxidative Stress, HA, and HYP Content of the Skin
2.7. Skin Histological
2.8. Skin Transcriptome Sequencing
2.8.1. RNA Extraction, Library Construction, and Transcriptome Sequencing
2.8.2. Bioinformatics Analyses of RNA-Sequencing Data
2.8.3. Reverse Transcriptase-Polymerase Chain Reaction (qRT-PCR)
2.9. Western Blot
2.10. ELISA
2.11. Statistical Analyses
3. Results and Discussion
3.1. Amino Acid Composition of Collagen
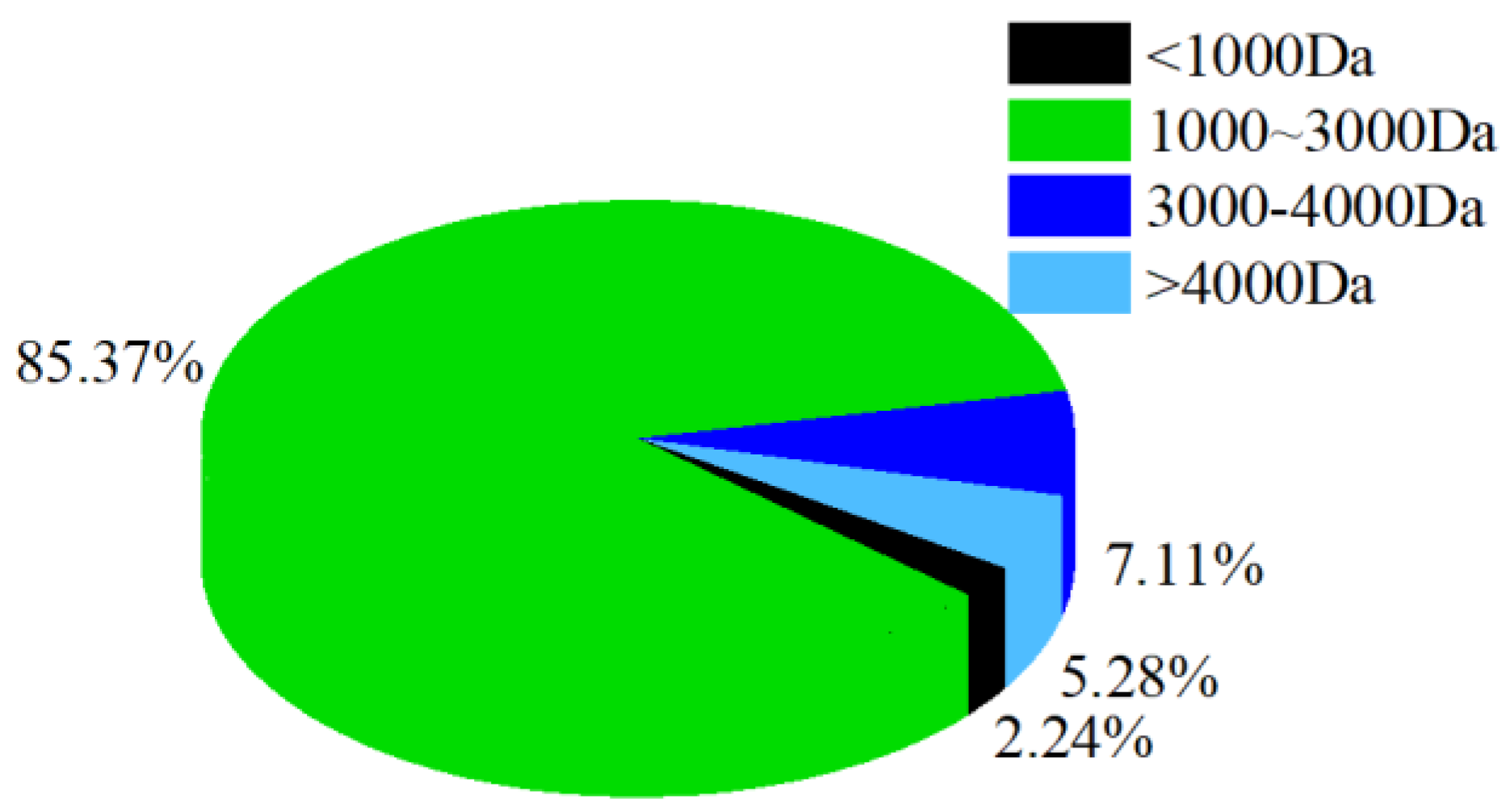
3.2. Molecular Weight Distribution of CPs
3.3. Effect of Oral CPs on Alleviating Skin Aging
3.3.1. Body Weight and Organ Index
3.3.2. Skin Composition
3.3.3. Skin Histological Changes
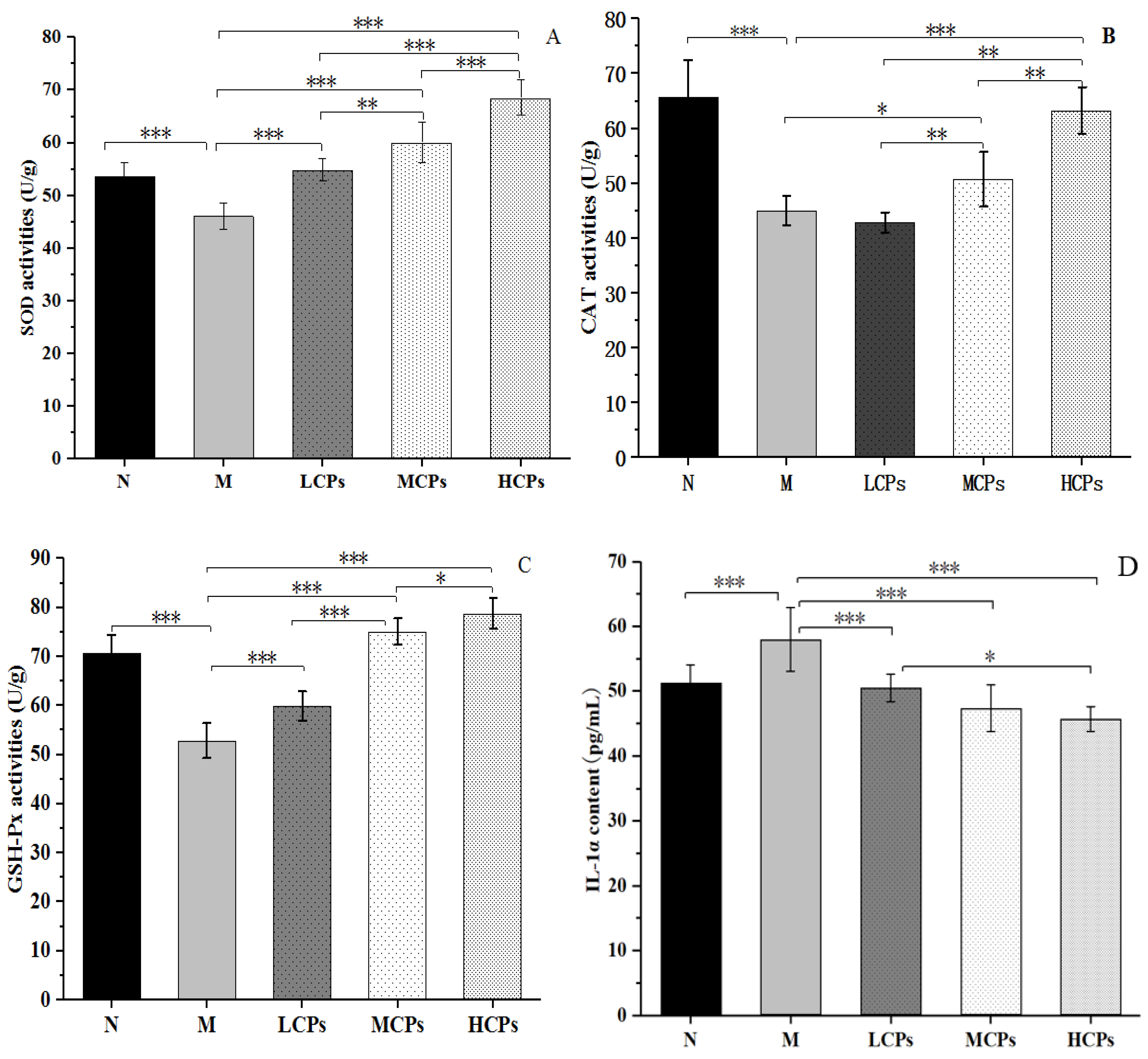
3.3.4. Skin Antioxidant and Inflammatory Levels
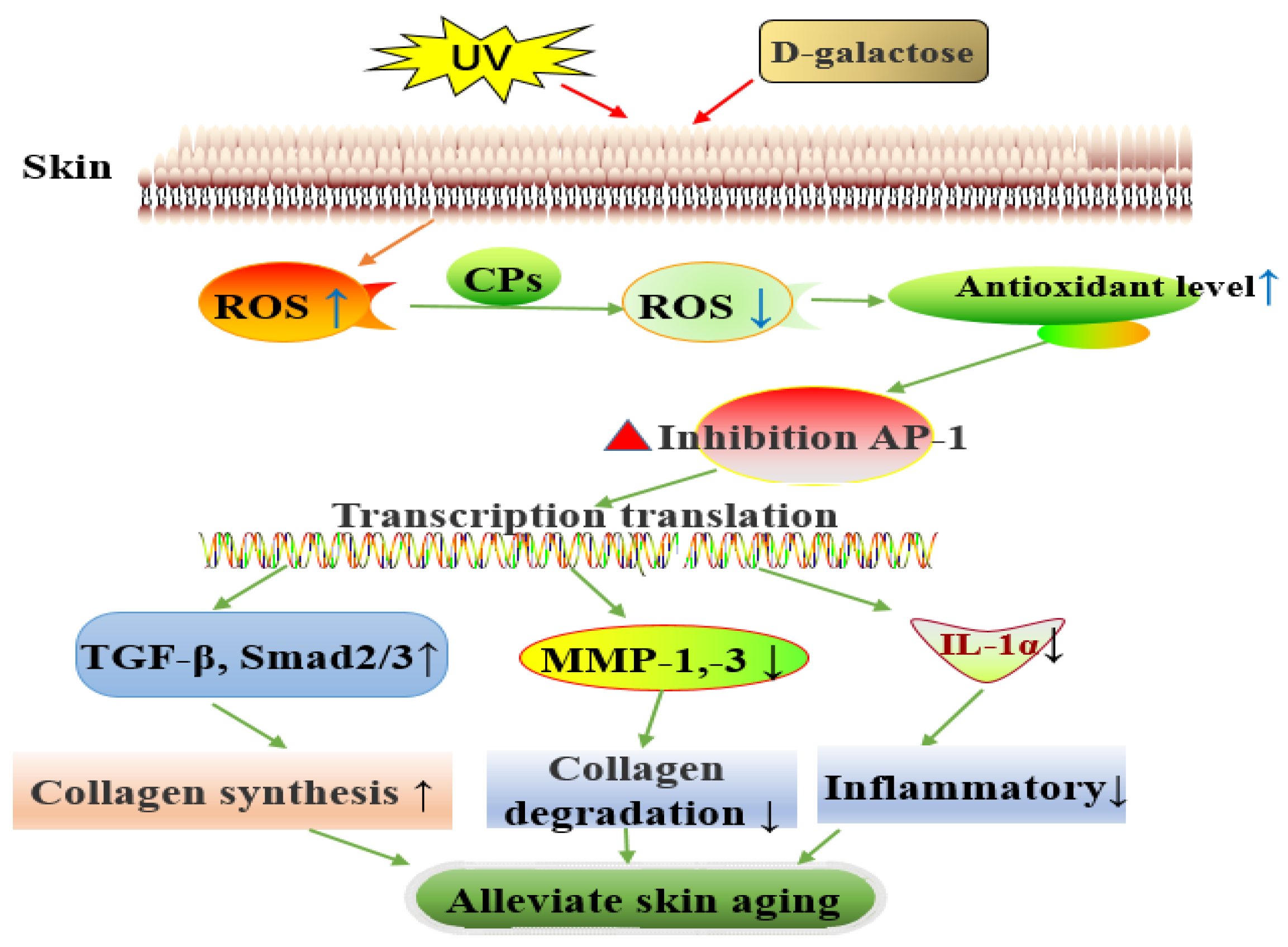
3.4. Mechanism of Action of Dietary CPs in Alleviating Skin Aging
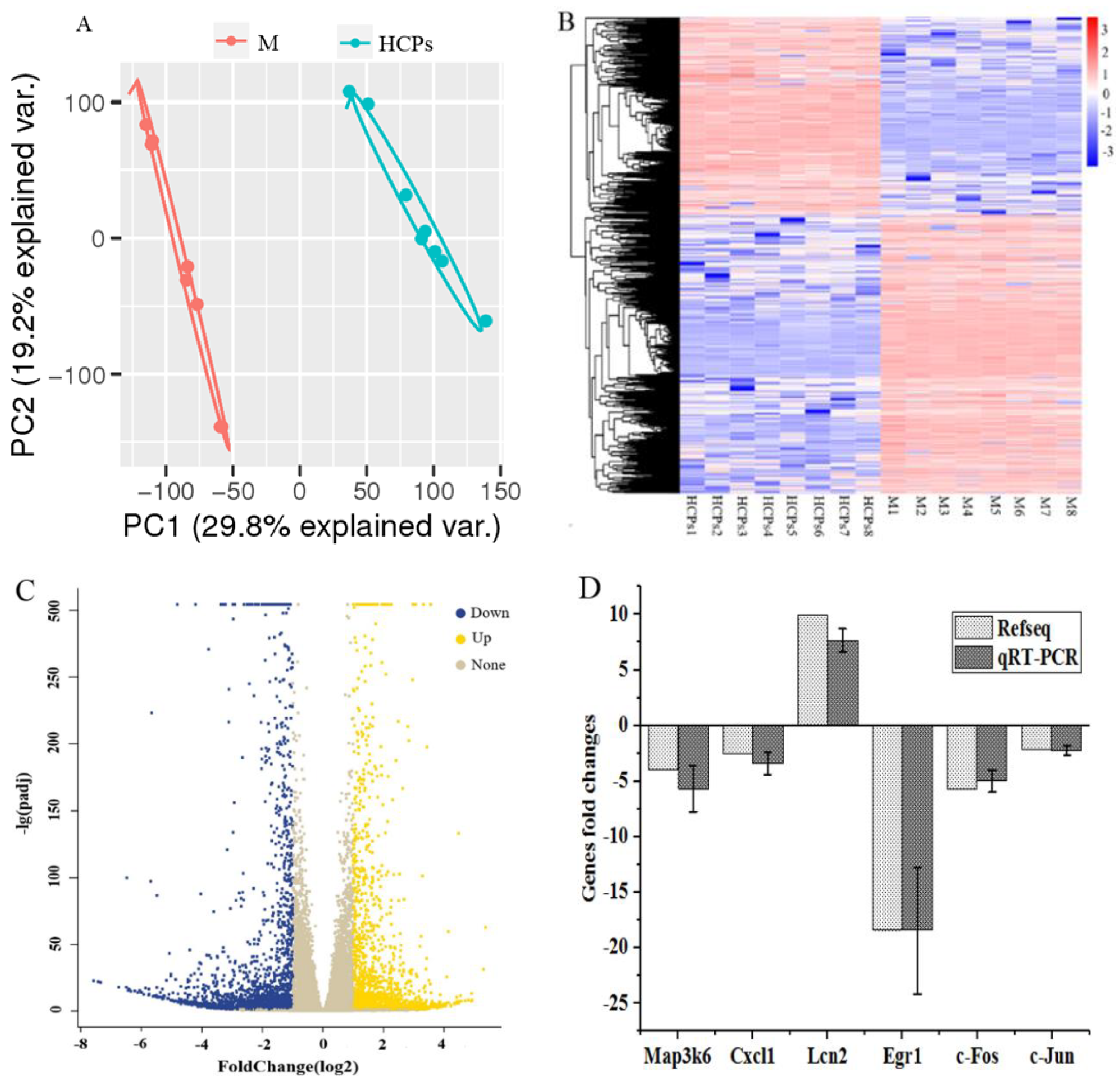
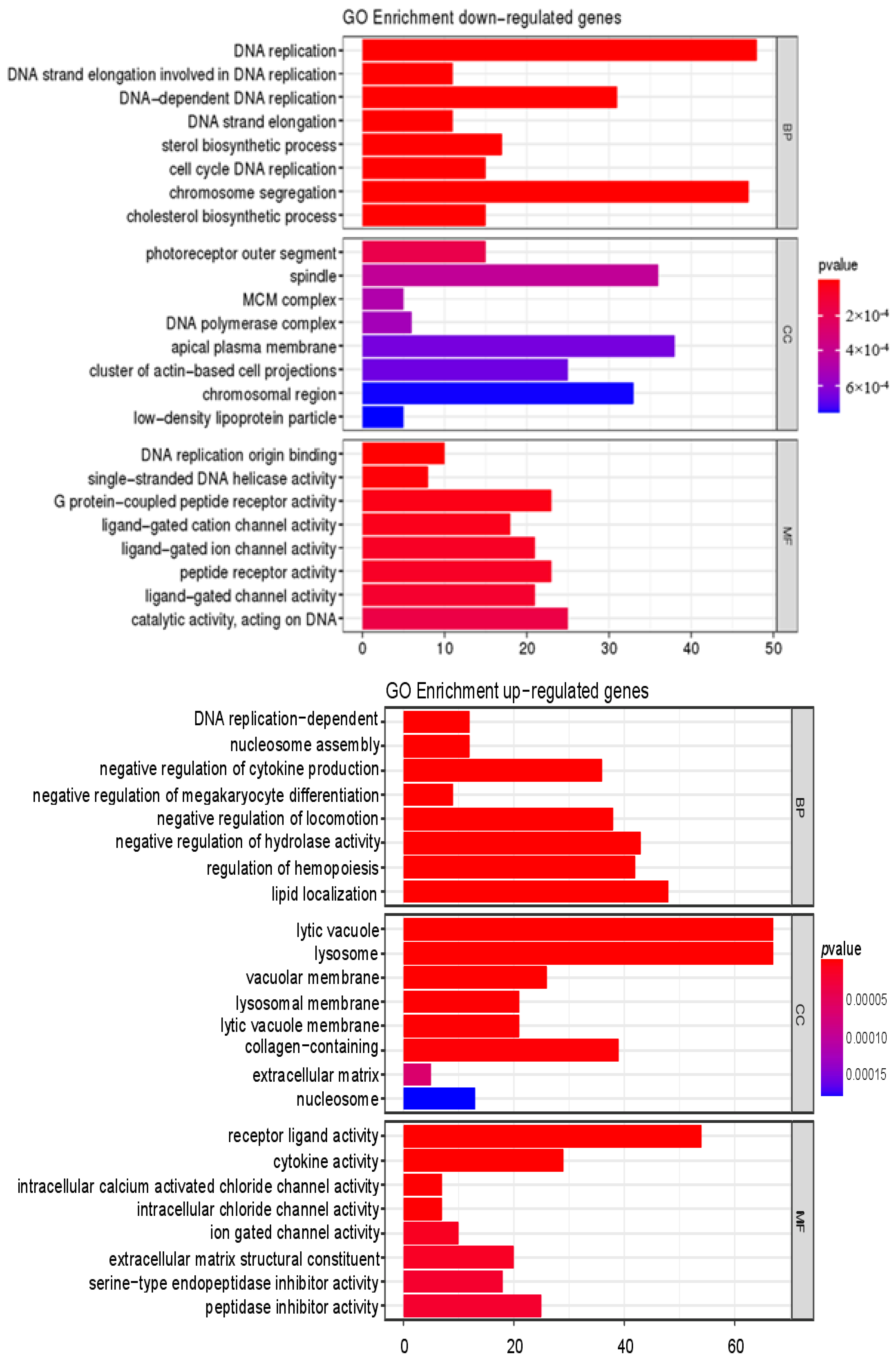
3.4.1. Analysis and Validation of RNA-Seq Data
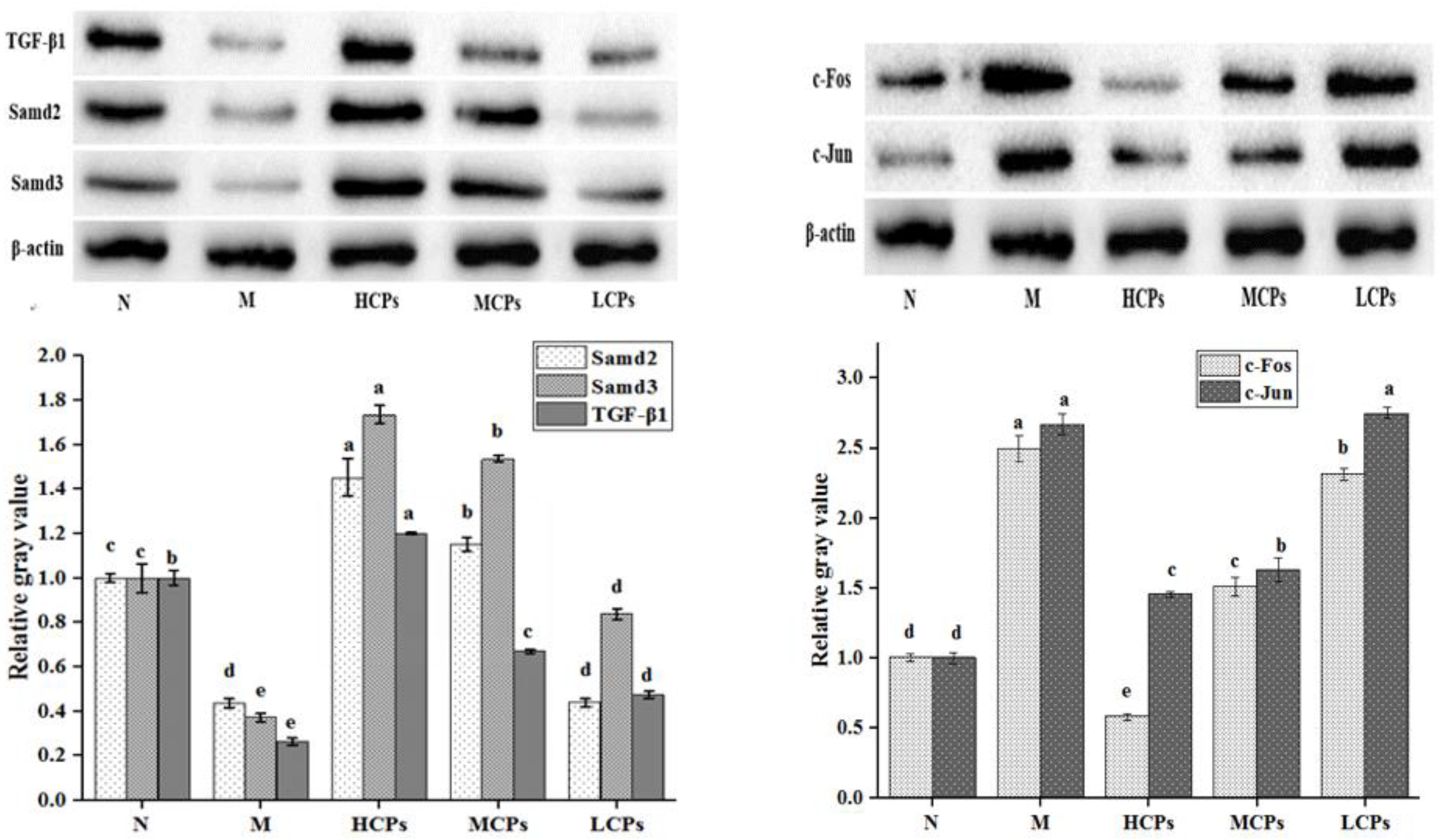
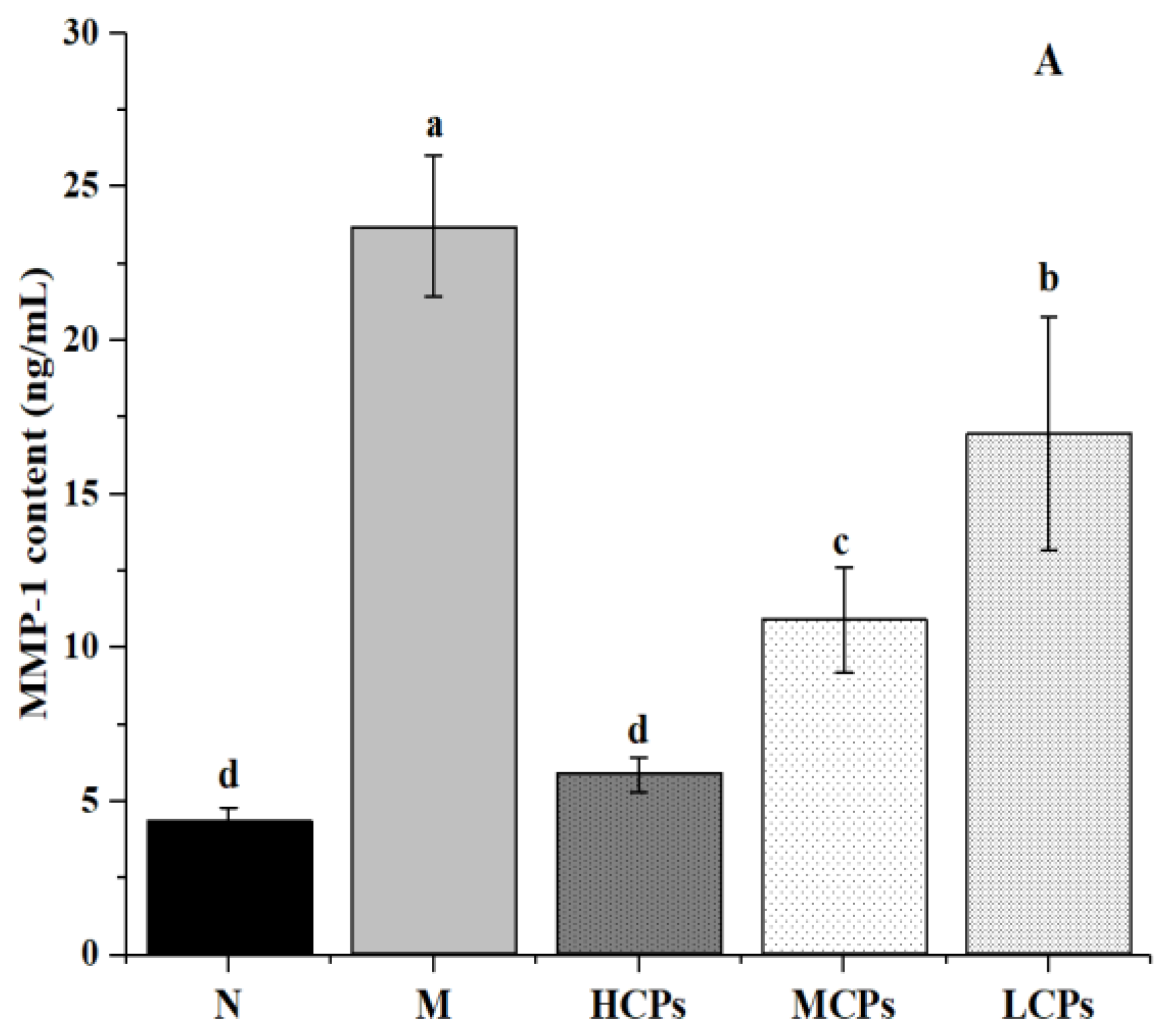
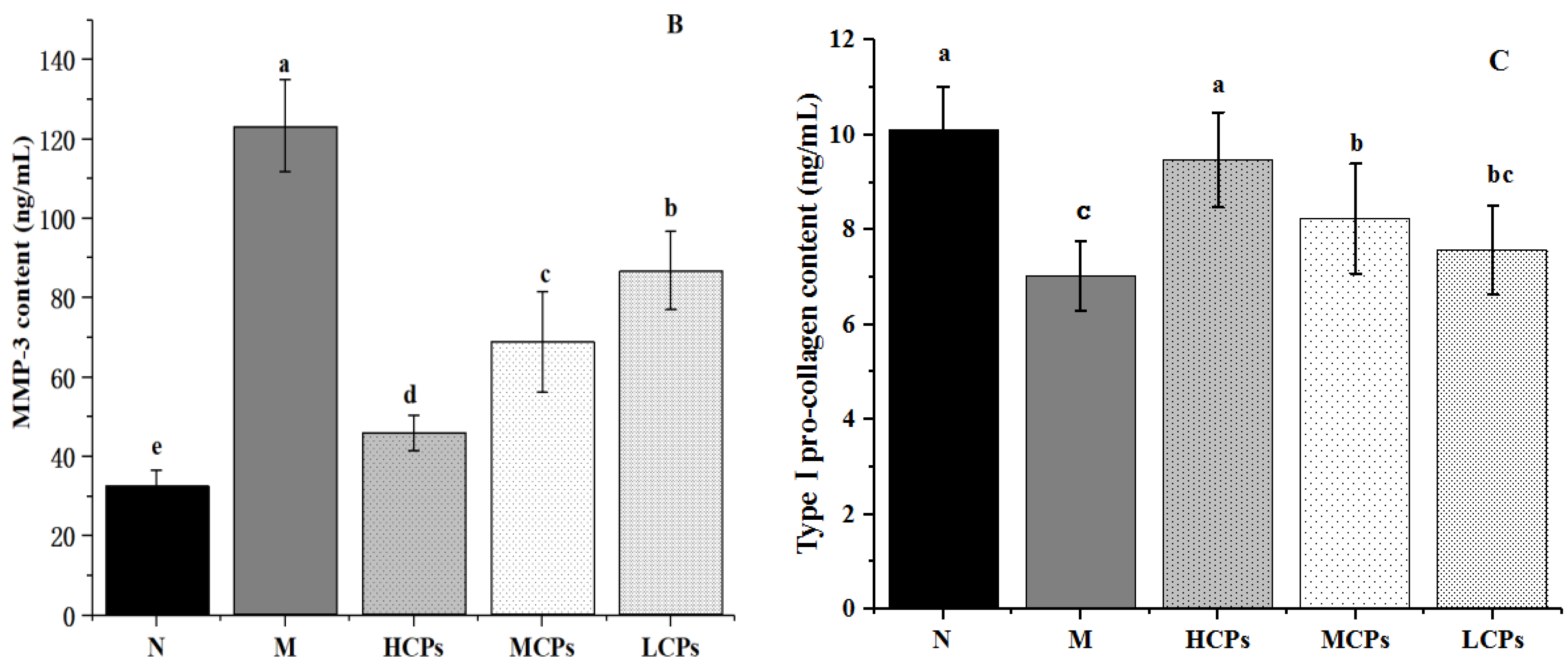
3.4.2. Verification of the Mechanism of Action of CPs in Alleviating Skin Aging
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blanpain, C.; Fuchs, E. Epidermal stem cells of the skin. Annu. Rev. Cell Dev. Biol. 2006, 22, 339–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orioli, D.; Dellambra, E. Epigenetic Regulation of Skin Cells in Natural Aging and Premature Aging Diseases. Cells 2018, 7, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Duan, E. Fighting against Skin Aging: The Way from Bench to Bedside. Cell Transplant. 2018, 27, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Su, G.; Zhou, F.; Zhang, J.; Zheng, L.; Zhao, M. Protective Effect of Bovine Elastin Peptides against Photoaging in Mice and Identification of Novel Antiphotoaging Peptides. J. Agric. Food Chem. 2018, 66, 10760–10768. [Google Scholar] [CrossRef]
- Cao, C.; Xiao, Z.; Wu, Y.; Ge, C. Diet and Skin Aging—From the Perspective of Food Nutrition. Nutrients 2020, 12, 870. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, Q.; Qian, J.; Liang, Q.; Wang, Z.; Xu, J.; He, S.; Ma, H. Bioavailability and bioavailable forms of collagen after oral administration to rats. J. Agric. Food Chem. 2015, 63, 3752–3756. [Google Scholar] [CrossRef]
- Halim, N.R.; Yusof, H.M.; Sarbon, N.M. Functional and bioactive properties of fish protein hydolysates and peptides: A comprehensive review. Trends Food Sci. Technol. 2016, 51, 24–33. [Google Scholar] [CrossRef]
- Wenlong, W.; Mengshi, C.; Jinhong, W.; Shaoyun, W. Hypothermia protection effect of antifreeze peptides from pigskin collagen on freeze-dried Streptococcus thermophiles and its possible action mechanism. LWT Food Sci. Technol. 2015, 63, 878–885. [Google Scholar]
- Cao, C.; Xiao, Z.; Ge, C.; Wu, Y. Animal by-products collagen and derived peptide, as important components of innovative sustainable food systems—A comprehensive review. Crit. Rev. Food Sci. 2021, 1–25. [Google Scholar] [CrossRef]
- Ohara, H.; Matsumoto, H.; Ito, K.; Iwai, K.; Sato, K. Comparison of quantity and structures of hydroxyproline-containing peptides in human blood after oral ingestion of gelatin hydrolysates from different sources. J. Agric. Food Chem. 2007, 55, 1532–1535. [Google Scholar] [CrossRef]
- Yamamoto, S.; Hayasaka, F.; Deguchi, K.; Okudera, T.; Furusawa, T.; Sakai, Y. Absorption and plasma kinetics of collagen tripeptide after peroral or intraperitoneal administration in rats. Biosci. Biotechnol. Biochem. 2015, 79, 2026–2033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe-Kamiyama, M.; Shimizu, M.; Kamiyama, S.; Taguchi, Y.; Sone, H.; Morimatsu, F.; Shirakawa, H.; Furukawa, Y.; Komai, M. Absorption and effectiveness of orally administered low molecular weight collagen hydrolysate in rats. J. Agric. Food Chem. 2010, 58, 835–841. [Google Scholar] [CrossRef] [PubMed]
- Yazaki, M.; Ito, Y.; Yamada, M.; Goulas, S.; Teramoto, S.; Nakaya, M.A.; Ohno, S.; Yamaguchi, K. Oral Ingestion of Collagen Hydrolysate Leads to the Transportation of Highly Concentrated Gly-Pro-Hyp and Its Hydrolyzed Form of Pro-Hyp into the Bloodstream and Skin. J. Agric. Food Chem. 2017, 65, 2315–2322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Li, Y.; Song, H.; He, J.; Li, G.; Zheng, Y.; Li, B. Collagen peptides promote photoaging skin cell repair by activating the TGF-beta/Smad pathway and depressing collagen degradation. Food Funct. 2019, 10, 6121–6134. [Google Scholar] [CrossRef]
- Song, H.; Zhang, S.; Zhang, L.; Li, B. Effect of Orally Administered Collagen Peptides from Bovine Bone on Skin Aging in Chronologically Aged Mice. Nutrients 2017, 9, 1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiaofeng, W.; Hui, H.; Jianping, W. Hen collagen hydrolysate alleviates UVA-induced damage in human dermal fibroblasts. J. Funct. Foods 2019, 63, 103574. [Google Scholar]
- Kuwahara, J. Extraction of Type I Collagen from Tilapia Scales Using Acetic Acid and Ultrafine Bubbles. Processes 2021, 9, 288. [Google Scholar] [CrossRef]
- Liu, D.; Zhang, X.; Li, T.; Yang, H.; Zhang, H.; Regenstein, J.M.; Zhou, P. Extraction and characterization of acid- and pepsin-soluble collagens from the scales, skins and swim-bladders of grass carp (Ctenopharyngodon idella). Food Biosci. 2015, 9, 68–74. [Google Scholar] [CrossRef]
- Zheng, Y.; Xu, Q.; Chen, H.; Chen, Q.; Gong, Z.; Lai, W. Transcriptome analysis of ultraviolet A-induced photoaging cells with deep sequencing. J. Dermatol. 2018, 45, 175–181. [Google Scholar] [CrossRef]
- Park, B.; Hwang, E.; Seo, S.A.; Zhang, M.; Park, S.; Yi, T. Dietary Rosa damascena protects against UVB-induced skin aging by improving collagen synthesis via MMPs reduction through alterations of c-Jun and c-Fos and TGF-β1 stimulation mediated smad2/3 and smad7. J. Funct. Foods 2017, 36, 480–489. [Google Scholar] [CrossRef]
- Cole, M.A.; Quan, T.; Voorhees, J.J.; Fisher, G.J. Extracellular matrix regulation of fibroblast function: Redefining our perspective on skin aging. J. Cell Commun. Signal. 2018, 12, 35–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akram, A.N.; Zhang, C. Effect of ultrasonication on the yield, functional and physicochemical characteristics of collagen-II from chicken sternal cartilage. Food Chem. 2020, 307, 125544. [Google Scholar] [CrossRef]
- Hong, H.; Fan, H.; Chalamaiah, M.; Wu, J. Preparation of low-molecular-weight, collagen hydrolysates (peptides): Current progress, challenges, and future perspectives. Food Chem. 2019, 301, 125222. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Meng, M.; Cheng, X.; Li, B.; Wang, C. The effect of collagen hydrolysates from silver carp (Hypophthalmichthys molitrix) skin on UV-induced photoaging in mice: Molecular weight affects skin repair. Food Funct. 2017, 8, 1538–1546. [Google Scholar] [CrossRef] [PubMed]
- Proksch, E.; Segger, D.; Degwert, J.; Schunck, M.; Zague, V.; Oesser, S. Oral supplementation of specific collagen peptides has beneficial effects on human skin physiology: A double-blind, placebo-controlled study. Skin Pharmacol. Physiol. 2014, 27, 47–55. [Google Scholar] [CrossRef]
- Proksch, E.; Schunck, M.; Zague, V.; Segger, D.; Degwert, J.; Oesser, S. Oral intake of specific bioactive collagen peptides reduces skin wrinkles and increases dermal matrix synthesis. Skin Pharmacol. Physiol. 2014, 27, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Torrent, A.; Ruhí, R.; Arañó, A.; Pérez, L.; van El, B. 546 Chondroprotective activity of a formulation that includes mannosamine. Osteoarthr. Cartil. 2010, 18, S245. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Jiang, Y.; Wang, X.; Zhou, J.; Haiying, C.; Weidong, X.; Yuanqing, H.; Haile, M.; Ruichang, G. Effect of oral administration of collagen hydrolysates from Nile tilapia on the chronologically aged skin. J. Funct. Foods 2018, 44, 112–117. [Google Scholar] [CrossRef]
- Min, C.K.; Silvia, Y.; Sun, Y.K. Oral Intake of Collagen Peptide Attenuates Ultraviolet B Irradiation-Induced Skin Dehydration In Vivo by Regulating Hyaluronic Acid Synthesis. Int. J. Mol. Sci. 2018, 19, 3551. [Google Scholar]
- Chen, T.; Hou, H.; Lu, J.; Zhang, K.; Li, B. Protective effect of gelatin and gelatin hydrolysate from salmon skin on UV irradiation-induced photoaging of mice skin. J. Ocean Univ. China 2016, 15, 711–718. [Google Scholar] [CrossRef]
- Zhang, L.; Zheng, Y.; Cheng, X.; Meng, M.; Luo, Y.; Li, B. The anti-photoaging effect of antioxidant collagen peptides from silver carp (Hypophthalmichthys molitrix) skin is preferable to tea polyphenols and casein peptides. Food Funct. 2017, 8, 1698–1707. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhu, H.; Zheng, Y.; Zhang, L.; Wang, X.; Luo, Z.; Tang, J.; Lin, L.; Du, Z.; Dong, C. The effects and mechanism of collagen peptide and elastin peptide on skin aging induced by D-galactose combined with ultraviolet radiation. J. Photochem. Photobiol. B Biol. 2020, 210, 111964. [Google Scholar] [CrossRef] [PubMed]
- Masaki, H. Role of antioxidants in the skin: Anti-aging effects. J. Dermatol. Sci. 2010, 58, 85–90. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, R.; Xue, Y.; Xiao, H.; Sheng, Y.; Zhao, D.; He, J.; Huang, H.; Wang, Q.; Wang, H. RNA-seq profiling reveals differentially expressed genes as potential markers for vital reaction in skin contusion: A pilot study. Forensic Sci. Res. 2018, 3, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Yeo, H.; Lee, Y.H.; Ahn, S.S.; Jung, E.; Lim, Y.; Shin, S.Y. Chrysin Inhibits TNFalpha-Induced TSLP Expression through Downregulation of EGR1 Expression in Keratinocytes. Int. J. Mol. Sci. 2021, 22, 4350. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Li, Q.; Liao, T.; Yin, X.; Chen, Q.; Wang, Z.; Dai, M.; Yi, L.; Ge, S.; Miao, C.; et al. Metabolomic profiling of single enlarged lysosomes. Nat. Methods 2021, 18, 788–798. [Google Scholar] [CrossRef] [PubMed]
- Shefler, I.; Pasmanik-Chor, M.; Kidron, D.; Mekori, Y.A.; Hershko, A.Y. T cell–derived microvesicles induce mast cell production of IL-24: Relevance to inflammatory skin diseases. J. Allergy Clin. Immunol. 2014, 133, 217–224. [Google Scholar] [CrossRef]
- Gerecke, D.R.; Chen, M.; Isukapalli, S.S.; Gordon, M.K.; Chang, Y.; Tong, W.; Androulakis, I.P.; Georgopoulos, P.G. Differential gene expression profiling of mouse skin after sulfur mustard exposure: Extended time response and inhibitor effect. Toxicol. Appl. Pharm. 2009, 234, 156–165. [Google Scholar] [CrossRef] [Green Version]
- Hough, C.M.; Purschke, D.N.; Huang, C.; Titova, L.V.; Kovalchuk, O.V.; Warkentin, B.J.; Hegmann, F.A. Intense terahertz pulses inhibit Ras signaling and other cancer-associated signaling pathways in human skin tissue models. J. Phys. Photonics 2021, 3, 34004. [Google Scholar] [CrossRef]
- Schonthaler, H.B.; Guinea-Viniegra, J.; Wagner, E.F. Targeting inflammation by modulating the Jun/AP-1 pathway. Ann. Rheum. Dis. 2011, 70 (Suppl. S1), i109–i112. [Google Scholar] [CrossRef]
- Rittié, L.; Fisher, G.J. UV-light-induced signal cascades and skin aging. Ageing Res. Rev. 2002, 1, 705–720. [Google Scholar] [CrossRef]
- Uluckan, O.; Guinea-Viniegra, J.; Jimenez, M.; Wagner, E.F. Signalling in inflammatory skin disease by AP-1 (Fos/Jun). Clin. Exp. Rheumatol. 2015, 33, S44–S49. [Google Scholar] [PubMed]
- Kammeyer, A.; Luiten, R.M. Oxidation events and skin aging. Ageing Res. Rev. 2015, 21, 16–29. [Google Scholar] [CrossRef] [PubMed]
- Fabi, S.; Sundaram, H. The Potential of Topical and Injectable Growth Factors and Cytokines for Skin Rejuvenation. Facial Plast. Surg. 2014, 30, 157–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leeman, D.S.; Hebestreit, K.; Ruetz, T.; Webb, A.E.; Mckay, A.; Pollina, E.A.; Dulken, B.W.; Zhao, X.; Yeo, R.W.; Ho, T.T.; et al. Lysosome activation clears aggregates and enhances quiescent neural stem cell activation during aging. Science 2018, 359, 1277–1283. [Google Scholar] [CrossRef] [Green Version]
- Kotaro, F.; Ruoyan, L.; Hengyu, C.; Kimiko, D.C.; Helen, Z.H.; Jianhua, X.; Fan, B.; Guang, Y. Graded regulation of cellular quiescence depth between proliferation and senescence by a lysosomal dimmer switch. Proc. Natl. Acad. Sci. USA 2019, 116, 22624–22634. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid Types | Content (%) | Amino Acid Types | Content (%) |
|---|---|---|---|
| Asp | 4.45 | Leu | 2.54 |
| Thr | 2.37 | Tyr | 0.49 |
| Ser | 3.72 | Phe | 2.03 |
| Glu | 9.14 | Lys | 3.15 |
| Gly | 27.86 | His | 0.57 |
| Ala | 9.61 | Arg | 5.22 |
| Val | 1.45 | Pro | 11.60 |
| Met | 2.11 | Hyp | 9.83 |
| Ile | 1.48 | Trp | 0 |
| Items | N | M | LCPs | MCPs | HCPs |
|---|---|---|---|---|---|
| Weight gain (g) | 2.53 | 2.38 | 2.44 | 4.11 | 3.98 |
| Moisture (%) | 71.32 ± 1.61 ab | 61.79 ± 3.94 d | 68.78 ± 3.44 bc | 66.48 ± 4.40 c | 73.83 ± 1.88 a |
| liver index (g/kg) | 46.97 ± 3.03 | 43.09 ± 2.79 | 48.83 ± 3.64 | 49.91 ± 4.90 | 49.34 ± 5.48 |
| Spleen index (g/kg) | 4.86 ± 0.31 | 4.62 ± 0.52 | 5.53 ± 0.61 | 5.32 ± 0.68 | 5.55 ± 0.40 |
| HA (pg/mg) | 40.98 ± 2.18 b | 31.11 ± 2.70 d | 35.27 ± 1.54 c | 42.03 ± 2.26 ab | 44.47 ± 2.74 a |
| Hyp (ug/g) | 1531.19 ± 157.64 b | 1289.15 ± 153.15 d | 1399.37 ± 116.59 cd | 1639.26 ± 155.18 ab | 1749.79 ± 147.32 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, C.; Xiao, Z.; Tong, H.; Liu, Y.; Wu, Y.; Ge, C. Oral Intake of Chicken Bone Collagen Peptides Anti-Skin Aging in Mice by Regulating Collagen Degradation and Synthesis, Inhibiting Inflammation and Activating Lysosomes. Nutrients 2022, 14, 1622. https://doi.org/10.3390/nu14081622
Cao C, Xiao Z, Tong H, Liu Y, Wu Y, Ge C. Oral Intake of Chicken Bone Collagen Peptides Anti-Skin Aging in Mice by Regulating Collagen Degradation and Synthesis, Inhibiting Inflammation and Activating Lysosomes. Nutrients. 2022; 14(8):1622. https://doi.org/10.3390/nu14081622
Chicago/Turabian StyleCao, Changwei, Zhichao Xiao, Huiquan Tong, Yuntao Liu, Yinglong Wu, and Changrong Ge. 2022. "Oral Intake of Chicken Bone Collagen Peptides Anti-Skin Aging in Mice by Regulating Collagen Degradation and Synthesis, Inhibiting Inflammation and Activating Lysosomes" Nutrients 14, no. 8: 1622. https://doi.org/10.3390/nu14081622