
β-hydroxybutyrate as an Anti-Aging Metabolite


Shanghai Frontiers Science Research Base of Exercise and Metabolic Health, Shanghai University of Sport, Shanghai 200438, China
*
Authors to whom correspondence should be addressed.
Nutrients 2021, 13(10), 3420; https://doi.org/10.3390/nu13103420
Submission received: 8 August 2021
/
Revised: 25 September 2021
/
Accepted: 26 September 2021
/
Published: 28 September 2021
(This article belongs to the Section Nutrition and Public Health)
Abstract
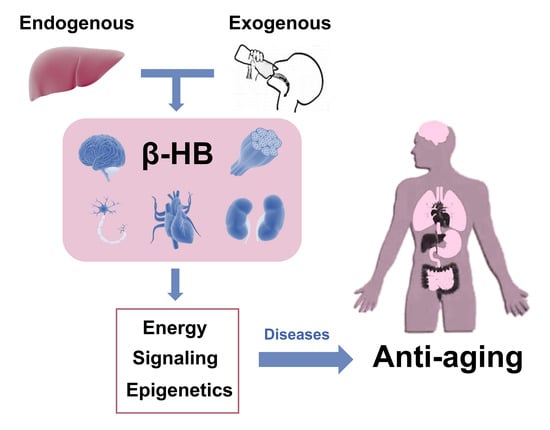
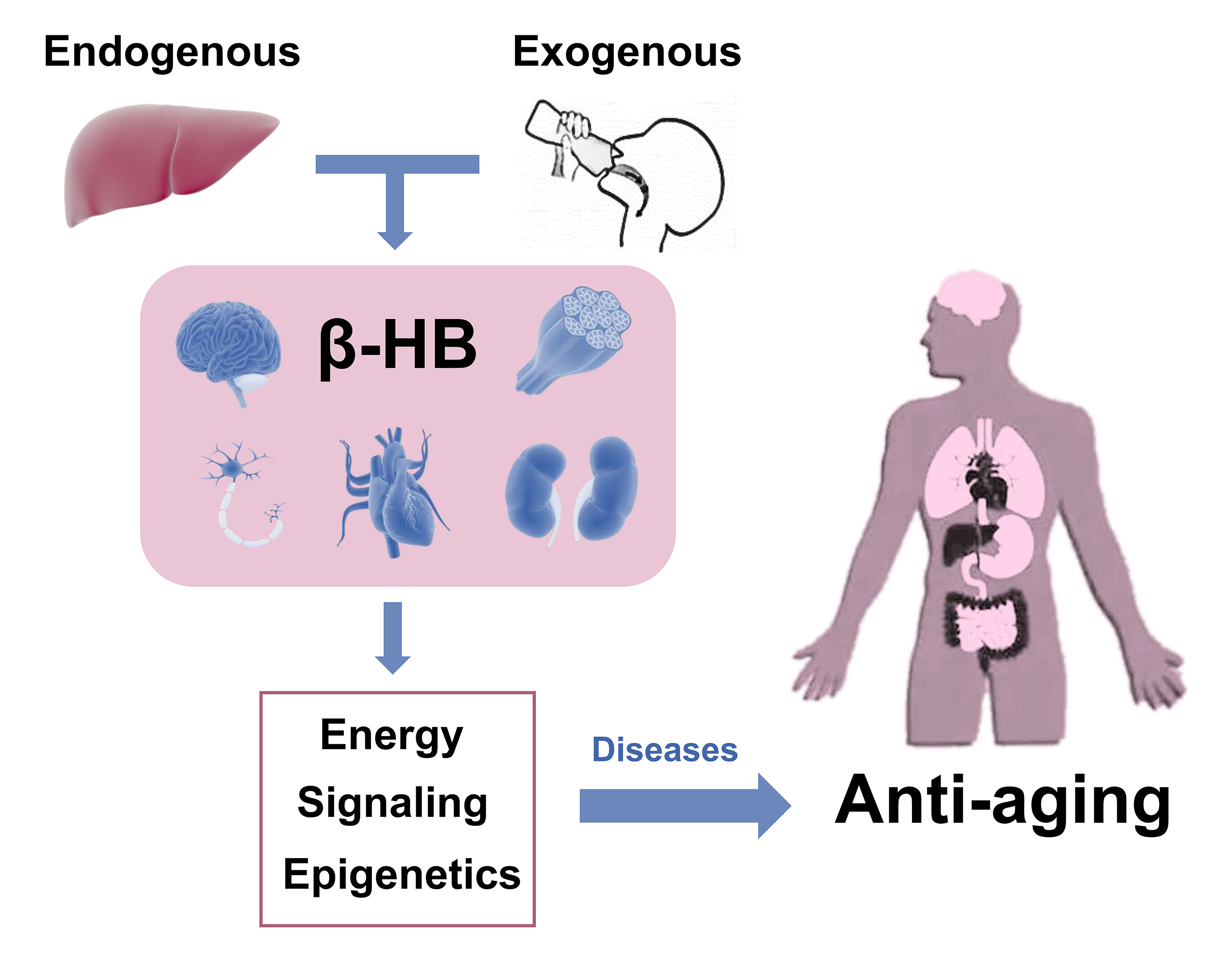
:The ketone bodies, especially β-hydroxybutyrate (β-HB), derive from fatty acid oxidation and alternatively serve as a fuel source for peripheral tissues including the brain, heart, and skeletal muscle. β-HB is currently considered not solely an energy substrate for maintaining metabolic homeostasis but also acts as a signaling molecule of modulating lipolysis, oxidative stress, and neuroprotection. Besides, it serves as an epigenetic regulator in terms of histone methylation, acetylation, β-hydroxybutyrylation to delay various age-related diseases. In addition, studies support endogenous β-HB administration or exogenous supplementation as effective strategies to induce a metabolic state of nutritional ketosis. The purpose of this review article is to provide an overview of β-HB metabolism and its relationship and application in age-related diseases. Future studies are needed to reveal whether β-HB has the potential to serve as adjunctive nutritional therapy for aging.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
Ketone bodies (KB) refer to β-hydroxybutyrate (C4H8O3, β-HB; also known as 3-hydroxybutyric acid), acetoacetate (AcAc), and acetone (Figure 1). They are small molecules primarily synthesized in the liver, produced from fatty acid oxidation-derived acetyl-coenzyme A (Ac-CoA), and transported to extrahepatic tissues for terminal oxidation. β-HB, the most abundant KB, is a water-soluble principal reversibly formed from the reduction of AcAc in the mitochondria [1]. It is present as R-/S-β-HB enantiomers (or termed D-/L-β-HB, respectively), with the R-enantiomer being the predominant circulating KB. Nutritional ketosis (NK) is defined as a nutritionally induced metabolic state of plasma β-HB concentration ≥0.5 mM, which can be achieved endogenously through caloric and dietary administration, mainly by ketogenic diet, or exogenously with oral supplementation, including β-HB and its related forms, resulting in a therapeutic effect in several medical conditions [2,3]. Apart from a role as an alternative energy source for extrahepatic tissues such as the brain, heart, or skeletal muscle, β-HB also acts as a signaling mediator involved in many regulations of cellular functions and adaptive responses [4,5]. It as well plays an epigenetic role in metabolic diseases through alterations in gene expression, cell surface receptor activation, as well as histone modifications.
Although the clinical efficacy of KB therapy is gradually widely recognized, there are still considerations about its potential mechanisms which are not fully clarified yet. In the past two decades, an explosive knowledge of the genetic and metabolic factors that affect aging has been witnessed. And a recent study has reported that the supplementation of D-β-HB to C. elegans (Caenorhabditis elegans, a kind of roundworm which is usually used for aging research) could improve an extension of lifespan, demonstrating the role of β-HB as “an anti-aging ketone body” [6]. This review will describe β-HB metabolism, outline the role of β-HB in age-related diseases, as well as summarize the therapeutic application of endogenous and exogenous β-HB. Hence, we discussed the underlying mechanisms of the metabolic effects of β-HB and explored whether β-HB had the potential to serve as adjunctive nutritional therapy for age-related diseases.
2. Overview of β-HB Metabolism
2.1. Ketone Body Production and Utilization
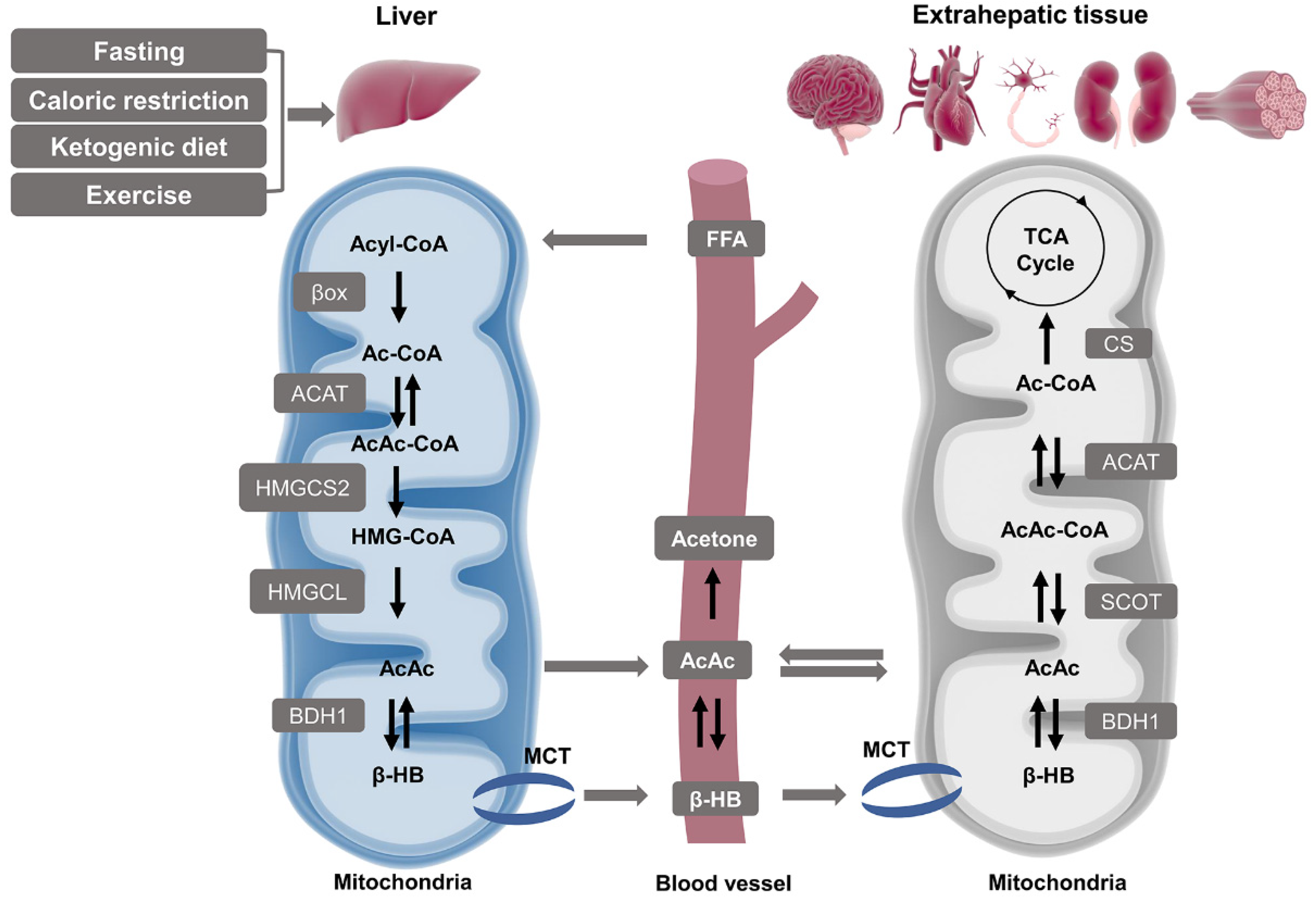
2.1.1. Ketogenesis in Liver
KB synthesis occurs mainly in the liver mitochondrial matrix which is called hepatic ketogenesis governed by the physiological and biochemical conversions of fat (Figure 2). Ketogenic amino acids may also undertake ketogenesis, but the production of circulating KB is likely less than 5% [7]. In the fasting state, low insulin combining with high cortisol and glucagon promotes adipocytes to release non-esterified fatty acids, which will enter the bloodstream [8]. Perivenous hepatocytes take up fatty acids in the mitochondria [9,10,11,12]. There is a series of reactions sequentially starting from Ac-CoA as well as acetoacetyl CoA (AcAc-CoA), and then transforming to 3-hydroxymethylglutaryl-CoA (HMG-CoA), finally ending with the releasing of AcAc [13]. Some of the AcAc is exported from the hepatocytes, yet the majority is reduced to β-HB catalyzed by 3-hydroxybutyrate dehydrogenase 1 (BDH1) in an NAD+/NADH-coupled equilibrium reaction [14,15].
2.1.2. Ketolysis in Extra-Hepatic Tissues
Circulating KB are transported through the plasma membrane via the monocarboxylate transporter (MCT) proteins (Figure 2). The expression of MCT is tissue-specific, MCT1 is ubiquitously expressed, MCT2 is specifically expressed in the neurons and kidney, and MCT4 is expressed in skeletal muscle, lung, heart, and glia [16,17,18]. Ketolysis occurs in these extrahepatic organs, including the brain, heart, neurons, kidney cortex, and skeletal muscle, using β-HB for terminal oxidation. This is the metabolic response to some definite physiological situations such as limited carbohydrate (CHO) content, post-exercise, or abnormal insulin signaling [19]. In these extrahepatic tissues, the oxidation of β-HB to AcAc is catalyzed by BDH in the mitochondria. Next, in the rate-limiting reaction of ketolysis, AcAc is activated into AcAc-CoA and succinate by succinyl-CoA:3-oxoacid CoA transferase (SCOT), which is also called 3-oxoacid-CoA transferase 1 (OXCT1). This enzyme is not expressed in the liver, and thus hepatic tissues are incapable of utilizing the β-HB they produce [20,21]. Two molecules of Ac-CoA subsequently transform from AcAc-CoA and enter the TCA cycle for energy requirement.
2.2. β-HB as an Energy Substrate
Under the circumstance of CHO restriction, prolonged fasting as well as exercise, fatty acids break down to β-HB, which turns out to be a compensatory energy fuel in the brain, heart, kidneys, and muscles (Figure 2) [22,23,24]. Thus, a ketosis-like condition is generated under the circumstance of lower blood glucose or glycogen depletion in which liver particularly provides fatty acid-derived β-HB [25]. Under physiological situations, the blood KB concentration ranges from 0.05 to 0.1 mM in humans, and when rising over 0.5 mM, it achieves a metabolic state of NK. Moreover, its level may reach 5–7 mM during prolonged fasting [26], starvation, caloric restriction (CR), ketogenic diet (KD) [27,28], exercise [29], insulin deficiency, pregnancy, or neonatal period [17], and even reach 10–25 mM in a particular circumstance of diabetic ketoacidosis [30,31]. 300 g of KB are produced daily by the human liver, and ketones could provide approximately 5% of the energy requirements in the fed state, rising to 20% in long-term fasting [32,33].
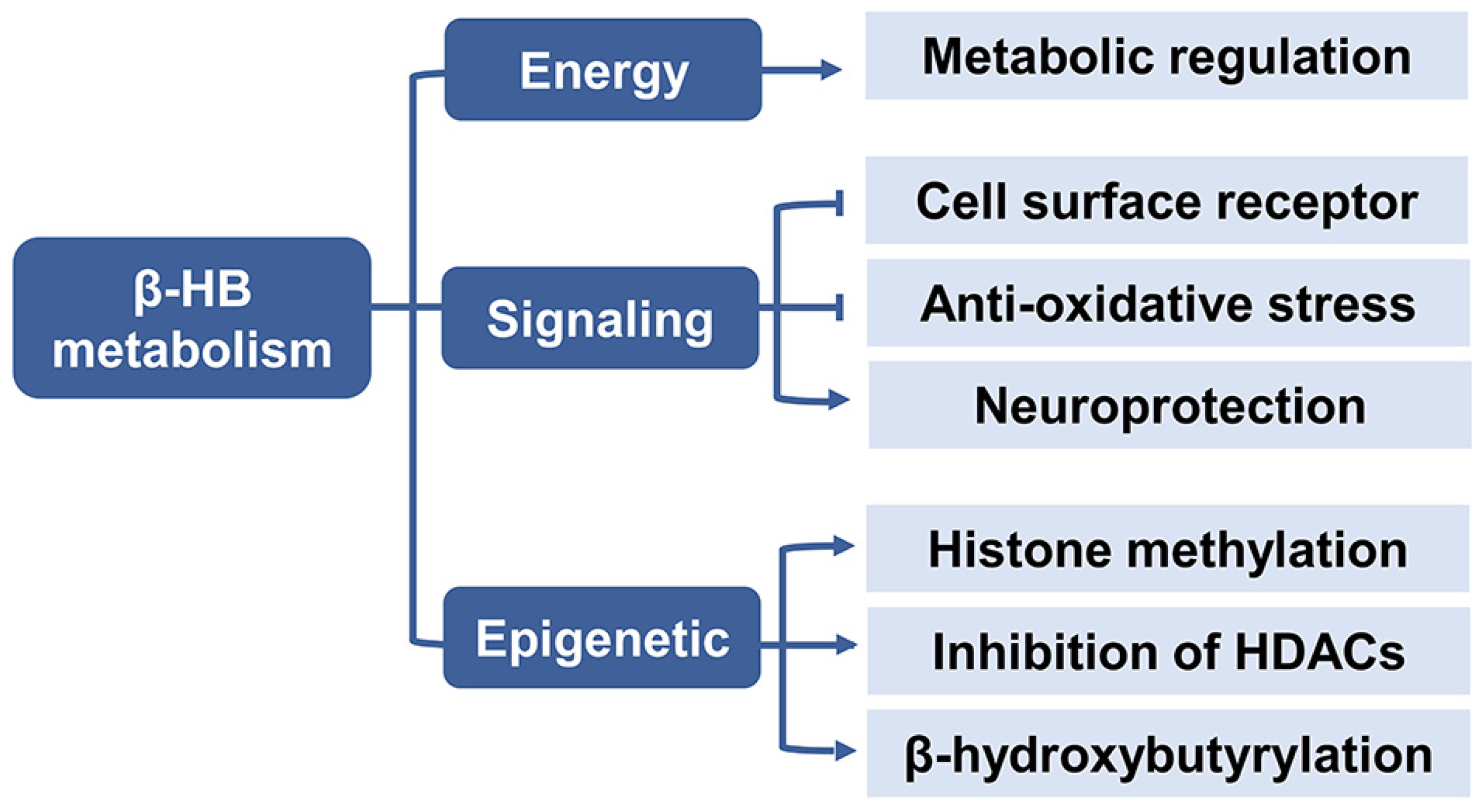
2.3. β-HB as a Signaling Mediator
β-HB is not only defined as a fat-derived energetic substrate for the brain, heart, and skeletal muscles under prolonged fasting and exercise but also acts as a metabolic signal modulating a wide variety of cellular functions (Figure 3) [34,35].
2.3.1. Cell Surface Receptor
β-HB has been described as a signaling molecule as it is a ligand for G protein-coupled receptors (GPCRs) including GPR41 and GPR109A (HM74A in humans, PUMA-G in mice), inducing the activation or inactivation of various signaling pathways relevant to lipid metabolism or cellular growth [36,37,38]. β-HB can modulate body energy expenditure and metabolic homeostasis through the mechanism which GPR41, known as free fatty acid receptor 3 (FFAR3), suppresses sympathetic nervous system activity via the Gβγ-PLCβ-MAPK signaling pathway [37,39,40]. GPR109A, which is regarded as hydroxy-carboxylic acid receptor 2 (HCAR2), is located on multiple cells such as neutrophils, macrophages, adipocytes, mediating anti-lipolytic activity, inflammation, and atherogenesis [41,42]. Thus, β-HB exerts numerous effects on neurodegeneration, inflammatory processes, and dyslipidemia as an endogenous ligand via the GPR109A signaling pathway [43].
2.3.2. Anti-Oxidative Stress
Oxidative stress is commonly defined as a condition in which the reactive oxygen species (ROS) are in excess, and this might be owing to excessive production or impaired elimination [44,45]. β-HB is associated with the attenuation of oxidative stress, due to its inhibition of ROS production, prevention of lipid peroxidation and protein oxidation as well as increasing the levels of antioxidant proteins [46]. β-HB plays an anti-oxidative effect in vivo and in vitro studies, especially in the aspect of neuroprotection. It is reported that in the year 2000, Kashiwaya and his colleagues found that β-HB could protect neurons from oxidative damage [47]. Treating cells with β-HB could reduce the cytosolic NADP+/NADPH ratio and increase reduced glutathione, which is one of the major cellular low molecular weight antioxidants. It was reported that treatment of lipopolysaccharides (LPS) in cells induced a proinflammatory stimulation, and the signaling pathway of nuclear factor-κB (NF-κB) was inhibited by β-HB via translocation and degradation of NF-κB inhibitor α. The proinflammatory response to LPS was alleviated by the management of β-HB to cells due to NF-κB regulating various proinflammatory genes expression, including interleukin (IL)-1β, IL-6, tumor necrosis factor-α, inducible nitric oxide synthase, and cyclooxygenase-2 [48]. β-HB metabolism may reduce the generation of ROS and enhance antioxidant defenses through altering redox couples ratios in cytoplasm and mitochondria, in which ROS is mainly produced [49].
2.3.3. Neuroprotection
β-HB could inhibit abnormal glycolysis and reduce glucose uptake through altering metabolic activity in neurodegeneration [50]. β-HB plays a role in neuroprotection by ATP production and regulates NADP+/NADPH ratio, glutathione activity, mitochondrial permeability, glycolytic flux, apoptosis, and neuroinflammation, triggering normal synaptic neurotransmission. β-HB supplementation and CR also mitigate oxidative stress, neuronal apoptosis, neuroinflammation, and intensify neurotrophins generation, including brain-derived neurotrophic factor (BDNF), neurotrophin-3 as well as glial cell line-derived neurotrophic factor, which associates with altered neuronal functions, especially in the senescent brain. It has been demonstrated that β-HB not only enhances neuronal respiration, ATP generation, oxidative functions, ATP-sensitive potassium channel activity, but also improves dopamine volume in the mesencephalon, and regulates motor functions as well [51,52].
2.4. β-HB as an Epigenetic Regulator
Epigenetics is the study of inherited changes in the gene function or phenotype caused by molecular mechanisms other than changes in DNA sequence [53]. The three major epigenetic mechanisms include DNA methylation, histone post-translational modifications (PTM), and microRNAs (miRNAs) transcriptional regulation. PTM consist of histone methylation, acetylation, phosphorylation, and ubiquitination. Of note, β-HB is particularly regarded as an endogenous inhibitor of class I histone deacetylase (HDAC) as an epigenetic regulator [54]. Besides, it has recently been shown that high levels of β-HB could induce lysine β-hydroxybutyrylation (Kbhb) in histones of cells [55]. β-HB has been shown to play various signaling roles of linking gene transcription and cellular function with the outside environment in epigenetic modifications which could explain its pleiotropic effects on aging (Figure 4).
2.4.1. Histone Deacetylases Inhibition
β-HB has been demonstrated to naturally inhibit HDACs in a dose-dependent manner, including HDAC 1, 3, and 4, which increases histone acetylation, decreases α-synuclein toxicity and prevents dopaminergic neurons from cell death [56,57]. β-HB may regulate HDACs and alter gene expression through these underlying mechanisms. For instance, β-HB-mediated inhibition of HDACs in mice raises BDNF expression, reduces the NAD+/NADH ratio, and increases ATP, which is closely related to adult neurogenesis [58,59]. In rodent models, BDNF also protects the central nervous system from NF-κB-mediated neuroinflammation and apoptosis [60,61]. Moreover, in the basal ganglia of Parkinson’s disease (PD) rats, the activity of various antioxidant enzymes increased and oxidative damage was restrained [62,63]. Therefore, β-HB may protect against energy depletion, oxidative stress, inflammation, autophagy, and apoptosis by inducing BDNF expression through the inhibition of HDACs.
2.4.2. Histone Lysine β-hydroxybutyrylation
Kbhb is recently considered as an adaptation to changes in cellular energy levels, promoting the transcription level of circadian rhythms and energy stress-response genes [64]. Kbhb as a newly identified epigenetic mark integrates the classic DNA methylation with PTM, offering a novel way to explore chromatin regulation and various functions of β-HB under the circumstance of important human pathophysiological states [65,66,67]. It has been reported that β-HB-mediated p53 Kbhb, which is an essential tumor suppression factor, is dramatically increased in cultured cells treated with β-HB, explaining the association between Kbhb and tumor as well as providing a promising therapeutic target for cancer treatment [68]. A recent study has demonstrated that the p300-dependent Kbhb pathway could directly mediate in vitro transcription through catalyzing the enzymatic addition, representing an adapted epigenetic mechanism for various cellular processes and physiopathological conditions [69]. Consequently, the actions of Kbhb may play vital roles in the pathogenesis and treatment of diverse human diseases.
3. Role of β-HB in Age-Related Diseases
3.1. Relationship between β-HB and Aging
It has been demonstrated that CR plays an important role in prolonging life and delaying the onset of age-related diseases in multiple species, such as mice, rats, fish, worms, flies, and yeast [70]. KD is considered to promote a longer lifespan the same as CR [71]. Circulating β-HB is seen as an antiaging metabolite, as it is increased significantly during CR and KD [72]. Moreover, the supplementation of β-HB could extend the lifespan of C. elegans and regulate aging and longevity [73]. β-HB diminishes senescence-associated secretory phenotype as well as senescent vascular cells in mammals [74]. Although a variety of studies verify the therapeutic effects of KD in regenerative medicine including aging and neurodegenerative diseases, the exact molecular mechanism of β-HB has not been fully explored. In addition, exogenous β-HB contributed to stem cell homeostasis and intestinal stem cell function via Notch signaling activation for tissue regeneration [75]. Therefore, β-HB could be regarded as a potential regenerative mediator as well as alleviate age-related diseases (Figure 5).
3.2. Age-Related Diseases
3.2.1. Cancers
Aging is a critical risk factor in the development of cancer, which is a primary cause of human death [76]. Plenty of energy is needed for cancer cells to enhance their proliferation rate. Cancer cells generate energy mainly by aerobic glycolysis, which is known as the Warburg effect [77]. This is regarded as an adaptive response of permitting carbons to be delivered via anabolic pathways and getting rid of the mitochondrial dysfunction induced by increased ROS levels. Thus, inhibiting glucose availability in cancer cells is considered an effective option. Recent animal studies showed that KD provides a therapeutic approach to tumor cells through selective metabolic oxidative stress, while simultaneously inhibiting primary tumorigenesis and systemic metastasis [78,79,80,81]. Thus, the published data indicated that KD may not significantly affect advanced and terminal cancers progression, yet it is safely used as an adjuvant therapy with other verified anticancer therapies and has the potential to improve the quality of cancer patients’ life [82,83]. It is suggested that this therapy should be further investigated to figure out the exact connections between β-HB and various types of cancer.
3.2.2. Neurological Disorders
There are multifactorial complex underlying pathophysiological mechanisms in neurodegenerative diseases. Ketogenic nutrition or exogenous β-HB is reported to have the capability to weaken the progression of pathological conditions such as Alzheimer’s disease (AD) and PD. Neurodegenerative diseases have the characteristic of defects in mitochondrial metabolism, including attenuated ATP generation, increased ROS production, complex IV dysfunction in AD, and complex I dysfunction in PD [84,85,86]. β-HB may promote mitochondrial metabolism via multiple mechanisms such as inducing mitochondrial turnover, reducing oxidative stress and impairment of mitochondria, as well as serving as an alternative energy substrate [87,88]. Moreover, β-HB supplementation may resist neuroinflammation through inhibiting pathologic microglial activation and regulating NOD-like receptor family pyrin domain containing 3 (NLRP3) inflammasome pathways [89,90]. Finally, β-HB could prevent the toxicity of neurotoxins of AD and PD [91]. It is obvious that not only by single intervention or lifestyle change such as drug, diet, exercise, and sleep, but combining ketone supplements along with these changes, may improve the neurological debilitating diseases.
3.2.3. Cardiovascular Diseases
Both morbidity and mortality of cardiovascular diseases (CVDs) increase with age, which is the central risk factor of many forms of major diseases. Different modalities of CR are regarded as effective interventions of extending lifespan on the aging process [92]. CR has been proven to produce positive defense responses in a variety of stress states, of which cardiovascular protective signaling is the core response, including mammalian target of rapamycin (mTOR), AMP-activated protein kinase (AMPK), sirtuins, and endothelial nitric oxide synthase signaling pathway [93]. KD is one kind of CR, which is closely associated with anti-aging impacts, during which high levels of β-HB are produced. Studies in mice have already demonstrated the role of β-HB in terms of extending healthspan, regulating HDACs, the enzymatic activity of epigenetic regulators, and thereby activating the expression of age-related genes [94]. β-HB therefore may be a powerful factor in attenuating cardiovascular aging which requires future research.
3.2.4. Muscle Dysfunction
Age-related muscle dysfunction is characterized by progressive sarcopenia and atrophy of skeletal muscles, which is related to frailty, muscle weakness, and disability in the elderly [95]. The mechanisms underlying this process including some dominant features, such as inhibited anabolic pathways, enhanced catabolic pathways, loss of muscle fiber, and muscle denervation in both older human and rodents’ studies [96]. Recently, the anticatabolic, anabolic, and regenerative potential of β-HB has been demonstrated by therapeutic ketosis in human skeletal muscle atrophy under an inflammatory microenvironment. [97,98]. β-HB has also been proved to potentially slow muscle loss with myopathies by maintaining mitochondrial respiration and morphology within muscle tissue [99]. Cell and animal studies have particularly suggested that β-HB induced by exercise or provided by exogenous supplementation might improve skeletal muscle and cognitive function [100,101]. HDACs have been reported to be associated with age-related muscle dysfunction, playing crucial roles in regulating metabolic processes in skeletal muscle [102]. β-HB, which is one kind of HDAC inhibitor, could be a promising target to treat sarcopenia clinically.
3.2.5. Inflammation
It is generally acknowledged that inflammatory mechanisms are closely associated with age-related diseases, such as arthritis, atherosclerosis, metabolic syndrome, fatty liver disease, type 2 diabetes, frailty, cachexia, cancer, and neurodegenerative disorders [103,104,105,106]. Long-term stimulation of immune response may lead to progressive degeneration of both innate and adaptive immune systems, causing decreased immunological competence, which is considered as “immunosenescence”. Chronic low-grade inflammation occurs throughout this immunological process which is named “inflammaging” [107]. β-HB is now considered a modulator of inflammation and immune cell function, yet various underlying mechanisms remain controversial [17]. β-HB administration could promote anti-inflammatory actions, involving the regulation of NLRP3 inflammasome in neutrophils and macrophages and the declining production of inflammatory molecules. [108]. Inflammation is an important component of many age-related diseases; thus, an anti-inflammatory response can be attributed to β-HB protection as recently reported [109].
3.2.6. Metabolic Syndrome
Due to multiple physiological mechanisms, aging is becoming one of the most critical risk factors for the occurrence and progression of metabolic syndrome, which is an age-related disease consisting of obesity, glucose intolerance, insulin resistance, dyslipidemia, and hypertension [110]. Endoplasmic reticulum (ER) stress is disturbed homeostasis leading to an impaired protein synthesis process such as the accumulation of unfolded and misfolded proteins, which is especially associated with the onset of metabolic disorders such as diabetes mellitus and disorderly hepatic lipid metabolism [111]. Nonalcoholic fatty liver disease has been regarded as a novel component of metabolic syndrome, with attenuated hepatic ketogenesis, insulin sensitivity, and abnormal fat accumulation [112,113]. NK has been found to improve metabolic and inflammatory markers, as well as lower insulin levels and promote β-HB production particularly during KD [114]. Of note, β-HB administration has already been reported to inhibit inflammasome formation, lipid accumulation, and oxidative stress by binding to specific HCARs, and inhibiting HDACs, FFARs, and NLRP3, to suppress ER stress, suggesting the beneficial effects of β-HB supplementation on liver steatosis and restoration of liver functions in aging progression [115,116,117].
4. Therapeutic Application of β-HB in Aging
4.1. Endogenous Ketosis
Endogenous ketosis can be achieved by using various durations of fasting, CR, or CHO restriction and particularly by a KD [118] (Figure 5).
4.1.1. Intermittent Fasting and Caloric Restriction
Energy-restricted metabolic states such as intermittent fasting (IF) or CR, have obvious characteristics of increased ketosis and could extend lifespan in animals [119]. Ketogenesis is believed to increase with prolonged starvation. IF is relatively easy to practice for a long time, with alternative periods of feeding or fasting which can last 24 h from one to four days per week, such as alternate-day fasting, whole-day fasting (periodic fasting), and time-restricted feeding [120,121]. IF has been proven to be safe in monitored patients and the hormonal level of this state is typically marked by a low-insulin, high-glucagon, and increased plasma fatty acids and cortisol environment which heavily promote lipolysis [122,123]. It is currently gaining more popularity and is being considered as a potential non-pharmacological way to promote healthy aging [124]. CR is achieved by reducing energy intake of about 25–30% without lacking essential nutrients, which has also been observed to improve age-related mortality and morbidity, delay aging progression, and result in healthspan in invertebrate and vertebrate species [125,126]. Circulating β-HB level is elevated as a beneficial metabolite and mediator during these two states, which are widely accepted as anti-aging interventions [127]. However, before these regimens, it is hard to maintain long-term ketosis. The mechanism of β-HB as a potential CR mimic to slow aging has yet to be explored further.
4.1.2. Ketogenic Diets
A large amount of data on the effects of β-HB metabolism comes from studies on KD, especially in rodents. KD is not an energy-limiting state, yet the related phenotype replicates some of the biochemical properties of IF and CR which are strongly associated with longevity. KD is composed of high-fat, adequate-protein, and a very low level of CHO (typically about 88%, 10%, and 2%). The KD promotes endogenous ketogenesis without fasting [128]. In terms of lifespan extension, β-HB has been proposed to promote longevity in worms via two different anti-aging pathways, which are inhibiting HDACs, leading to increased DAF-16/FOXO activity, as well as involving the mitochondrial metabolism of β-HB, and activating the SKN-1/Nrf2 antioxidant response pathway [73]. KD has also been demonstrated to improve the longevity and survival of mice, together with increased protein acetylation and decreased activation of tissue-specific mTOR complex 1 [71]. KD has been mechanistically investigated to improve neuroprotection and mitochondrial metabolism, activate autophagy, enhance antioxidative and anti-inflammatory capability, and inhibit insulin/insulin-like growth factor signaling, which contributes to the anti-aging process [129]. Although KD has already been clinically used as a therapy, which is easier to sustain than CR, it is in some ways difficult to rigorously follow and requires specific medical guidance and strong motivation [130].
4.2. Exogenous Ketosis and Supplementation of β-HB
It has been demonstrated that not only KD, but also exogenous ketone supplements (EKSs) can increase and maintain blood KB level, especially β-HB, so as to promote anti-aging effects [131]. β-HB supplements are now being commercially marketed as an alternative to KD. The supplements are commonly present in either a powder form of ketone salts (KS) or a liquid form of ketone ester (KE). In addition, medium-chain triglycerides (MCTs) or their combination with MCTs oil are also usually used to induce and sustain NK to improve ketotic response [132,133]. The production of β-HB from these supplements would not be affected by CHO, thus administration of EKSs may be practical and alternative when maintaining a normal diet to achieve therapeutic ketogenesis (Figure 5) [134].
4.2.1. Ketone Salts
Oral administration of isolated β-HB would be the most direct method of exogenously inducing NK. However, KB in its free acid form can be expensive, unstable, and ineffective at producing sustained ketosis. Thus, ketone acids buffering with sodium, potassium, calcium, or other electrolytes have been explored to enhance efficacy, inhibit overload of any single mineral, and these compounds are commercially available. It is reported that co-ingestion with MCTs may improve the efficiency of increasing β-HB relatively, at least in rats [135]. However, a few undesirable adverse effects exist while consuming large doses of KS, which usually results in gastrointestinal distress, and inappropriate cation overload or acidosis [136].
4.2.2. Ketone Esters
Several existing synthetic KEs prove to be the most effective agents to induce immediate, sustained, and dose-dependent elevation in serum ketones concentration, which provides an alternative way to increase β-HB and is well-tolerated in rodents and humans [137,138]. Ester bonds hold KEs together and are cleaved by gastric esterases to release KB in their free acid form from the backbone molecule, which is often a ketogenic precursor molecule R,S-1,3-butanediol (BD). Two prominent KEs in the recent research studies are (R)-3-hydroxybutyl (R)-3-hydroxybutyrate ketone monoester (KME) and R,S-1,3-butanediol acetoacetate ketone diester (KDE), the former appears safer and superior at appropriate doses in healthy adults, whatever acutely or daily sustained up to 28 days [139,140,141,142]. NK produced by KEs is therefore achieved without prolonged fasting or KD yet it is currently the most potent method of EKSs.
4.2.3. Medium Chain Triglycerides
MCTs have a much greater ketogenic potential than long-chain fatty acids since they are rapidly absorbed, energy-dense, water-miscible, and tasteless. 6 to 12 carbons of fatty acids are contained in MCTs in length. MCTs can be hydrolyzed to medium-chain fatty acids by lipases in the gastrointestinal tract, and then rapidly metabolized to Ac-CoA, finally to KB in the liver [143]. MCTs are consequently regarded as ketogenic fats due to their ability of ketogenesis without the restriction of dietary CHO intake [144]. Unfortunately, high MCTs consumption are often not well adopted because of their gastrointestinal side effects, including diarrhea, dyspepsia, and flatulence, which could be alleviated through a progressive 1 or 2-week period [145]. In addition, the generation of β-HB by supplementation of MCTs is at a low level in the blood [146].
4.2.4. R, S-1,3-Butanediol
BD is an organic butyl alcohol approved by the Food and Drug Administration, which is metabolized to produce two isoforms of β-HB, D- and L-β-HB or R- and S-β-HB via hepatic conversion, even though it is not a fatty acid or MCT [147]. Oral administration of BD could achieve ketosis and approach a KD state in dogs [148]. It was demonstrated that a dose-dependent elevation of KB in a ratio of 6:1 of R-β-HB to AcAc could be produced by BD in rodents [149]. BD is often utilized as a backbone in the synthesis of KE. Gut or tissue esterases could easily break the ester bond and release KB and BD without the involvement of salts or acid [150]. A variety of preclinical toxicology studies have found that BD is safe and tolerable [151].
4.2.5. β-HB Enantiomers
β-HB is a chiral molecule, with two enantiomers, R/D and S/L, which is an important characteristic in terms of its signaling activities as well as possible therapeutic applications [152]. Currently, a racemic mixture enantiomer of β-HB is the most commercially available ingestible EKSs apart from KS and KME, as its synthesis is more affordable than pure enantiomers. The chiral specificity is introduced by BDH1, determining that only R-β-HB is the normal product of human metabolism and could be readily catabolized into ATP and Ac-CoA [153]. IF, CR, KD, exercise, or any other situation which leads to endogenous β-HB would produce only R-β-HB. It is reported that ingestion of the same amount of racemic EKSs may produce higher and more sustained S-β-HB in blood circulation due to its slower metabolization compared to R-β-HB [154,155]. Despite divergent metabolic effects, these two enantiomers have similar molecular interactions as well as intracellular signal transduction cascades, which remains a hotly debated topic.
4.3. Comparisons between Endogenous and Exogenous Ketosis Induced by NK
In essence, the metabolic conditions of chronic endogenous dietary ketosis are in obvious contrast to the rapid exogenous ketosis delivered by ketone bodies. The following are the differentiations. Firstly, KD elevates blood β-HB level to a range of 0.5 mM to 3.0 mM, whereas EKSs approximately elevate to 0.3 mM to 1.0 mM [156]. Secondly, KD requires a couple of days to achieve sustained NK state, while EKSs elevate β-HB concentration acutely. Thirdly, KD need to follow strict CHO intake while EKSs require no direct CHO restriction, which determines EKSs has higher compliance, especially in short-term supplementation [142]. Fourthly, circulating glucose concentrations are therefore divergent because of the diverse CHO requirement. Fifthly, their similar anabolic and anticatabolic effects have as well been demonstrated [157,158]. Finally, KD and EKSs have been reported to reduce the substrate utilization of CHO during exercise, yet under distinct metabolic states [159,160]. NK is defined as a metabolic state, exerting physiological changes at both systemic and cellular level wherein β-HB concentration is over 0.5 mM regardless if induced by endogenous or exogenous ketones. These effects can be similar or different and can be universal or tissue-specific. NK is believed to be a potential state for performance-enhancing or therapeutic benefits under endogenous or exogenous ketosis, which need further evaluation.
5. Future Perspectives
Future studies are necessary to further elucidate the following practical issues: activating endogenous NK in a normal dietary context to sustain a steady state of metabolism; improving the targeted delivery of β-HB prodrugs or precursors to avoid excessive salt load or acidosis; bringing β-HB to the sites of action by using existing endogenous transporters and metabolite gradients to explore specific downstream signaling events; confirming whether the synthetic KE compounds need been strictly pure due to its enormous financial burden for the majority of patients and health systems; investigating if S-β-HB has a better pharmacokinetic than R-β-HB, which might help reduce cost and discover another signaling function; establishing methodologies to quantify β-HB flux rates and differentiate these two enantiomers in terms of concentrations and impacts. Besides, the recommended dose and timing of β-HB supplements, the short half-life and bitter taste of KME as well as the interaction with other substrates in various nutritional surroundings are also needed to be specified. β-HB is emerging as vitally important regulators of metabolic health and longevity, alleviating aging phenotypes via multiple and yet unknown molecular mechanisms. By modulating lipolysis, energy expenditure, metabolic rate, insulin resistance, autophagy, feeding behavior, as well as exercise performance, β-HB might serve as a signaling biomolecule to affect cellular function and human healthspan. The evaluation of β-HB may be a crucial approach for the treatment of the aging population.
6. Conclusions
In conclusion, β-HB is the most abundant KB and plays a vital role as an energetic metabolite, a signaling molecule, as well as an epigenetic regulator, which could be used as a therapeutic agent in a range of cancers, neurodegeneration, traumatic brain disorders, cardiac diseases, muscle dysfunction, metabolic syndrome, and inflammation. Endogenous ketosis and an exogenous supplement may be promising strategies for numerous diseases. Further research is needed to investigate whether ketotherapeutics can promote healthy aging, and to figure out the specific relationship and underlying mechanisms between β-HB and the aging process, which may offer a novel way in delaying the onset and development of age-associated dysfunctions.
Author Contributions
The authors’ contributions were as follows—L.W. performed the literature searches and drafted the manuscript; L.W., P.C. and W.X. were involved in the design, review, and editing of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China, grant No. 31271273 and 31300975; Shanghai Key Laboratory of Human Movement Development and Protection (Shanghai University of Sport), Grant No. 11DZ2261100; “Shuguang Program” (20SG50) funded by Shanghai Education Development Foundation and Shanghai Municipal Education Commission; and The Research Program of Exercise and Public Health (No. 0831).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wakeman, A.J.; Dakin, H.D. On the decomposition of β-oxybutyric acid and aceto-acetic acid by enzymes of the liver. J. Biol. Chem. 1909, 6, 373–389. [Google Scholar] [CrossRef]
- Poff, A.M.; Koutnik, A.P.; Egan, B. Nutritional Ketosis with Ketogenic Diets or Exogenous Ketones: Features, Convergence, and Divergence. Curr. Sports Med. Rep. 2020, 19, 251–259. [Google Scholar] [CrossRef]
- Kovács, Z.; Brunner, B.; Ari, C. Beneficial Effects of Exogenous Ketogenic Supplements on Aging Processes and Age-Related Neurodegenerative. Dis. Nutr. 2021, 13, 2197. [Google Scholar]
- Bhattacharya, K.; Matar, W.; Tolun, A.A.; Devanapalli, B.; Thompson, S.; Dalkeith, T.; Lichkus, K.; Tchan, M. The use of sodium DL-3-Hydroxybutyrate in severe acute neuro-metabolic compromise in patients with inherited ketone body synthetic disorders. Orphanet J. Rare Dis. 2020, 15, 53. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Ramanathan, A. The Aging Metabolome-Biomarkers to Hub Metabolites. Proteomics 2020, 20, e1800407. [Google Scholar] [CrossRef] [Green Version]
- Edwards, C.; Copes, N.; Bradshaw, P.C. D-ß-hydroxybutyrate: An anti-aging ketone body. Oncotarget 2015, 6, 3477–3478. [Google Scholar] [CrossRef]
- Thomas, L.K.; Ittmann, M.; Cooper, C. The role of leucine in ketogenesis in starved rats. Biochem. J. 1982, 204, 399–403. [Google Scholar] [CrossRef] [Green Version]
- Soto-Mota, A.; Norwitz, N.G.; Clarke, K. Why a d-β-hydroxybutyrate monoester? Biochem. Soc. Trans. 2020, 48, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Cahill, G.F., Jr.; Herrera, M.G.; Morgan, A.P.; Soeldner, J.S.; Steinke, J.; Levy, P.L.; Reichard, G.A., Jr.; Kipnis, D.M. Hormone-fuel interrelationships during fasting. J. Clin. Investig. 1966, 45, 1751–1769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flatt, J.P. On the maximal possible rate of ketogenesis. Diabetes 1972, 21, 50–53. [Google Scholar] [CrossRef]
- Garber, A.J.; Menzel, P.H.; Boden, G.; Owen, O.E. Hepatic ketogenesis and gluconeogenesis in humans. J. Clin. Investig. 1974, 54, 981–989. [Google Scholar] [CrossRef] [Green Version]
- Reichard, G.A., Jr.; Owen, O.E.; Haff, A.C.; Paul, P.; Bortz, W.M. Ketone-body production and oxidation in fasting obese humans. J. Clin. Investig. 1974, 53, 508–515. [Google Scholar] [CrossRef]
- Newman, J.C.; Verdin, E. β-hydroxybutyrate: Much more than a metabolite. Diabetes Res. Clin. Pr. 2014, 106, 173–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bock, H.; Fleischer, S. Preparation of a homogeneous soluble D-beta-hydroxybutyrate apodehydrogenase from mitochondria. J. Biol. Chem. 1975, 250, 5774–5781. [Google Scholar] [CrossRef]
- Lehninger, A.L.; Sudduth, H.C.; Wise, J.B. D-beta-Hydroxybutyric dehydrogenase of muitochondria. J. Biol. Chem. 1960, 235, 2450–2455. [Google Scholar] [CrossRef]
- Halestrap, A.P.; Wilson, M.C. The monocarboxylate transporter family--role and regulation. IUBMB Life 2012, 64, 109–119. [Google Scholar] [CrossRef]
- Puchalska, P.; Crawford, P.A. Multi-dimensional roles of ketone bodies in fuel metabolism, signaling, and therapeutics. Cell Metab. 2017, 25, 262–284. [Google Scholar] [CrossRef] [Green Version]
- Halestrap, A.P. The SLC16 gene family—Structure, role and regulation in health and disease. Mol. Asp. Med. 2013, 34, 337–349. [Google Scholar] [CrossRef]
- Fukao, T.; Lopaschuk, G.D.; Mitchell, G.A. Pathways and control of ketone body metabolism: On the fringe of lipid biochemistry. Prostaglandins Leukot. Essent. Fat. Acids 2004, 70, 243–251. [Google Scholar] [CrossRef]
- Fukao, T.; Song, X.Q.; Mitchell, G.A.; Yamaguchi, S.; Sukegawa, K.; Orii, T.; Kondo, N. Enzymes of ketone body utilization in human tissues: Protein and messenger RNA levels of succinyl-coenzyme A (CoA):3-ketoacid CoA transferase and mitochondrial and cytosolic acetoacetyl-CoA thiolases. Pediatr. Res. 1997, 42, 498–502. [Google Scholar] [CrossRef] [Green Version]
- Orii, K.E.; Fukao, T.; Song, X.Q.; Mitchell, G.A.; Kondo, N. Liver-specific silencing of the human gene encoding succinyl-CoA: 3-ketoacid CoA transferase. Tohoku J. Exp. Med. 2008, 215, 227–236. [Google Scholar] [CrossRef] [Green Version]
- Owen, O.E.; Morgan, A.P.; Kemp, H.G.; Sullivan, J.M.; Herrera, M.G.; Cahill, G.F., Jr. Brain metabolism during fasting. J. Clin. Investig. 1967, 46, 1589–1595. [Google Scholar] [CrossRef]
- Sultan, A.M. D-3-hydroxybutyrate metabolism in the perfused rat heart. Mol. Cell Biochem. 1988, 79, 113–118. [Google Scholar] [CrossRef]
- Abbasi, J. Ketone Body Supplementation-A Potential New Approach for Heart Disease. JAMA 2021, 326, 17–18. [Google Scholar] [CrossRef]
- Paoli, A.; Bosco, G.; Camporesi, E.M.; Mangar, D. Ketosis, ketogenic diet and food intake control: A complex relationship. Front. Psychol. 2015, 6, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krebs, H. Biochemical aspects of ketosis. Proc. R. Soc. Med. 1960, 53, 71–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, A.M.; Williamson, D.H. Physiological roles of ketone bodies as substrates and signals in mammalian tissues. Physiol. Rev. 1980, 60, 143–187. [Google Scholar] [CrossRef] [PubMed]
- Laffel, L. Ketone bodies: A review of physiology, pathophysiology and application of monitoring to diabetes. Diabetes Metab. Res. Rev. 1999, 15, 412–426. [Google Scholar] [CrossRef]
- Johnson, R.H.; Walton, J.L.; Krebs, H.A.; Williamson, D.H. Metabolic fuels during and after severe exercise in athletes and non-athletes. Lancet 1969, 2, 452–455. [Google Scholar] [CrossRef]
- Gershuni, V.M.; Yan, S.L.; Medici, V. Nutritional ketosis for weight management and reversal of metabolic syndrome. Curr. Nutr. Rep. 2018, 7, 97–106. [Google Scholar] [CrossRef]
- Cahill, G.F., Jr. Fuel metabolism in starvation. Annu. Rev. Nutr. 2006, 26, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Balasse, E.O.; Féry, F. Ketone body production and disposal: Effects of fasting, diabetes, and exercise. Diabetes Metab. Rev. 1989, 5, 247–270. [Google Scholar] [CrossRef]
- Balasse, E.O.; Fery, F.; Neef, M.A. Changes induced by exercise in rates of turnover and oxidation of ketone bodies in fasting man. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1978, 44, 5–11. [Google Scholar] [CrossRef]
- Rojas-Morales, P.; Tapia, E.; Pedraza-Chaverri, J. β-Hydroxybutyrate: A signaling metabolite in starvation response? Cell. Signal. 2016, 28, 917–923. [Google Scholar] [CrossRef] [PubMed]
- Cotter, D.G.; Schugar, R.C.; Crawford, P.A. Ketone body metabolism and cardiovascular disease. Am. J. Physiol. Heart Circ. Physiol. 2013, 304, H1060–H1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taggart, A.K.; Kero, J.; Gan, X.; Cai, T.Q.; Cheng, K.; Ippolito, M.; Ren, N.; Kaplan, R.; Wu, K.; Wu, T.J.; et al. (D)-beta-Hydroxybutyrate inhibits adipocyte lipolysis via the nicotinic acid receptor PUMA-G. J. Biol. Chem. 2005, 280, 26649–26652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, I.; Inoue, D.; Maeda, T.; Hara, T.; Ichimura, A.; Miyauchi, S.; Kobayashi, M.; Hirasawa, A.; Tsujimoto, G. Short-chain fatty acids and ketones directly regulate sympathetic nervous system via G protein-coupled receptor 41 (GPR41). Proc. Natl. Acad. Sci. USA 2011, 108, 8030–8035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Offermanns, S. The nicotinic acid receptor GPR109A (HM74A or PUMA-G) as a new therapeutic target. Trends Pharm. Sci. 2006, 27, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Stilling, R.M.; van de Wouw, M.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. The neuropharmacology of butyrate: The bread and butter of the microbiota-gut-brain axis? Neurochem. Int. 2016, 99, 110–132. [Google Scholar] [CrossRef] [PubMed]
- Won, Y.J.; Lu, V.B.; Puhl, H.L., 3rd; Ikeda, S.R. β-Hydroxybutyrate modulates N-type calcium channels in rat sympathetic neurons by acting as an agonist for the G-protein-coupled receptor FFA3. J. Neurosci. 2013, 33, 19314–19325. [Google Scholar] [CrossRef]
- Tunaru, S.; Kero, J.; Schaub, A.; Wufka, C.; Blaukat, A.; Pfeffer, K.; Offermanns, S. PUMA-G and HM74 are receptors for nicotinic acid and mediate its anti-lipolytic effect. Nat. Med. 2003, 9, 352–355. [Google Scholar] [CrossRef] [PubMed]
- Lukasova, M.; Malaval, C.; Gille, A.; Kero, J.; Offermanns, S. Nicotinic acid inhibits progression of atherosclerosis in mice through its receptor GPR109A expressed by immune cells. J. Clin. Investig. 2011, 121, 1163–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovács, Z.; D’Agostino, D.P.; Diamond, D.; Kindy, M.S.; Rogers, C.; Ari, C. Therapeutic Potential of Exogenous Ketone Supplement Induced Ketosis in the Treatment of Psychiatric Disorders: Review of Current Literature. Front. Psychiatry 2019, 10, 363. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.T. Oxidative stress and mitochondrial dysfunction-linked neurodegenerative disorders. Neurol. Res. 2017, 39, 73–82. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Shan, W.; Zhu, F.; Wu, J.; Wang, Q. Ketone bodies in neurological diseases: Focus on neuroprotection and underlying mechanisms. Front. Neurol. 2019, 10, 585. [Google Scholar] [CrossRef] [Green Version]
- Kashiwaya, Y.; Takeshima, T.; Mori, N.; Nakashima, K.; Clarke, K.; Veech, R.L. D-beta-hydroxybutyrate protects neurons in models of Alzheimer’s and Parkinson’s disease. Proc. Natl. Acad. Sci. USA 2000, 97, 5440–5444. [Google Scholar] [CrossRef] [Green Version]
- Fu, S.P.; Li, S.N.; Wang, J.F.; Li, Y.; Xie, S.S.; Xue, W.J.; Liu, H.M.; Huang, B.X.; Lv, Q.K.; Lei, L.C.; et al. BHBA suppresses LPS-induced inflammation in BV-2 cells by inhibiting NF-κB activation. Mediat. Inflamm. 2014, 2014, 983401. [Google Scholar] [CrossRef] [Green Version]
- Norwitz, N.G.; Hu, M.T.; Clarke, K. The mechanisms by which the ketone body D-β-Hydroxybutyrate may improve the multiple cellular pathologies of Parkinson’s disease. Front. Nutr. 2019, 6, 63. [Google Scholar] [CrossRef]
- Włodarek, D. Role of ketogenic diets in neurodegenerative diseases (Alzheimer’s disease and Parkinson’s disease). Nutrients 2019, 11, 169. [Google Scholar] [CrossRef] [Green Version]
- Shaafi, S.; Najmi, S.; Aliasgharpour, H.; Mahmoudi, J.; Sadigh-Etemad, S.; Farhoudi, M.; Baniasadi, N. The efficacy of the ketogenic diet on motor functions in Parkinson’s disease: A rat model. Iran. J. Neurol. 2016, 15, 63–69. [Google Scholar] [PubMed]
- Tieu, K.; Perier, C.; Caspersen, C.; Teismann, P.; Wu, D.C.; Yan, S.D.; Naini, A.; Vila, M.; Jackson-Lewis, V.; Ramasamy, R.; et al. D-beta-hydroxybutyrate rescues mitochondrial respiration and mitigates features of Parkinson disease. J. Clin. Investig. 2003, 112, 892–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bird, A. Perceptions of epigenetics. Nature 2007, 447, 396–398. [Google Scholar] [CrossRef] [PubMed]
- Newman, J.C.; Verdin, E. Ketone bodies as signaling metabolites. Trends Endocrinol. Metab. 2014, 25, 42–52. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Zhang, D.; Chung, D.; Tang, Z.; Huang, H.; Dai, L.; Qi, S.; Li, J.; Colak, G.; Chen, Y.; et al. Metabolic regulation of gene expression by histone lysine β-Hydroxybutyrylation. Mol. Cell 2016, 62, 194–206. [Google Scholar] [CrossRef] [Green Version]
- Kontopoulos, E.; Parvin, J.D.; Feany, M.B. Alpha-synuclein acts in the nucleus to inhibit histone acetylation and promote neurotoxicity. Hum. Mol. Genet. 2006, 15, 3012–3023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, C.; Campbell, K.R.; Ryan, B.J.; Carling, P.; Attar, M.; Vowles, J.; Perestenko, O.V.; Bowden, R.; Baig, F.; Kasten, M.; et al. Single-cell sequencing of iPSC-dopamine neurons reconstructs disease progression and identifies HDAC4 as a regulator of Parkinson cell phenotypes. Cell Stem Cell 2019, 24, 93–106. [Google Scholar] [CrossRef] [Green Version]
- Sleiman, S.F.; Henry, J.; Al-Haddad, R.; El Hayek, L.; Abou Haidar, E.; Stringer, T.; Ulja, D.; Karuppagounder, S.S.; Holson, E.B.; Ratan, R.R.; et al. Exercise promotes the expression of brain derived neurotrophic factor (BDNF) through the action of the ketone body β-hydroxybutyrate. eLife 2016, 5, e15092. [Google Scholar] [CrossRef]
- Marosi, K.; Kim, S.W.; Moehl, K.; Scheibye-Knudsen, M.; Cheng, A.; Cutler, R.; Camandola, S.; Mattson, M.P. 3-Hydroxybutyrate regulates energy metabolism and induces BDNF expression in cerebral cortical neurons. J. Neurochem. 2016, 139, 769–781. [Google Scholar] [CrossRef]
- Xu, D.; Lian, D.; Wu, J.; Liu, Y.; Zhu, M.; Sun, J.; He, D.; Li, L. Brain-derived neurotrophic factor reduces inflammation and hippocampal apoptosis in experimental Streptococcus pneumoniae meningitis. J. Neuroinflamm. 2017, 14, 156. [Google Scholar] [CrossRef] [Green Version]
- Makar, T.K.; Trisler, D.; Sura, K.T.; Sultana, S.; Patel, N.; Bever, C.T. Brain derived neurotrophic factor treatment reduces inflammation and apoptosis in experimental allergic encephalomyelitis. J. Neurol. Sci. 2008, 270, 70–76. [Google Scholar] [CrossRef]
- Mattson, M.P.; Lovell, M.A.; Furukawa, K.; Markesbery, W.R. Neurotrophic factors attenuate glutamate-induced accumulation of peroxides, elevation of intracellular Ca2+ concentration, and neurotoxicity and increase antioxidant enzyme activities in hippocampal neurons. J. Neurochem. 1995, 65, 1740–1751. [Google Scholar] [CrossRef]
- Kirschner, P.B.; Jenkins, B.G.; Schulz, J.B.; Finkelstein, S.P.; Matthews, R.T.; Rosen, B.R.; Beal, M.F. NGF, BDNF and NT-5, but not NT-3 protect against MPP+ toxicity and oxidative stress in neonatal animals. Brain Res. 1996, 713, 178–185. [Google Scholar] [CrossRef]
- Zhang, X.; Cao, R.; Niu, J.; Yang, S.; Ma, H.; Zhao, S.; Li, H. Molecular basis for hierarchical histone de-β-hydroxybutyrylation by SIRT3. Cell Discov. 2019, 5, 35. [Google Scholar] [CrossRef]
- Dąbek, A.; Wojtala, M.; Pirola, L.; Balcerczyk, A. Modulation of cellular biochemistry, epigenetics and metabolomics by ketone bodies. Implications of the ketogenic diet in the physiology of the organism and pathological states. Nutrients 2020, 12, 788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boison, D. New insights into the mechanisms of the ketogenic diet. Curr. Opin. Neurol. 2017, 30, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Ruan, H.B.; Crawford, P.A. Ketone bodies as epigenetic modifiers. Curr. Opin. Clin. Nutr. Metab. Care 2018, 21, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Li, F.; Sun, Q.; Lin, N.; Han, H.; You, K.; Tian, F.; Mao, Z.; Li, T.; Tong, T.; et al. p53 β-hydroxybutyrylation attenuates p53 activity. Cell Death Dis. 2019, 10, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Zhang, D.; Weng, Y.; Delaney, K.; Tang, Z.; Yan, C.; Qi, S.; Peng, C.; Cole, P.A.; Roeder, R.G.; et al. The regulatory enzymes and protein substrates for the lysine β-hydroxybutyrylation pathway. Sci. Adv. 2021, 7, eabe2771. [Google Scholar] [CrossRef] [PubMed]
- Witte, A.V.; Fobker, M.; Gellner, R.; Knecht, S.; Flöel, A. Caloric restriction improves memory in elderly humans. Proc. Natl. Acad. Sci. USA 2009, 106, 1255–1260. [Google Scholar] [CrossRef] [Green Version]
- Roberts, M.N.; Wallace, M.A.; Tomilov, A.A.; Zhou, Z.; Marcotte, G.R.; Tran, D.; Perez, G.; Gutierrez-Casado, E.; Koike, S.; Knotts, T.A.; et al. A ketogenic diet extends longevity and healthspan in adult mice. Cell Metab. 2017, 26, 539–546. [Google Scholar] [CrossRef] [Green Version]
- Veech, R.L.; Bradshaw, P.C.; Clarke, K.; Curtis, W.; Pawlosky, R.; King, M.T. Ketone bodies mimic the life span extending properties of caloric restriction. IUBMB Life 2017, 69, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Edwards, C.; Canfield, J.; Copes, N.; Rehan, M.; Lipps, D.; Bradshaw, P.C. D-beta-hydroxybutyrate extends lifespan in C. elegans. Aging 2014, 6, 621–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.M.; Bedarida, T.; Ding, Y.; Somba, B.K.; Lu, Q.; Wang, Q.; Song, P.; Zou, M.H. β-Hydroxybutyrate prevents vascular senescence through hnRNP A1-mediated upregulation of Oct4. Mol. Cell 2018, 71, 1064–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, C.W.; Biton, M.; Haber, A.L.; Gunduz, N.; Eng, G.; Gaynor, L.T.; Tripathi, S.; Calibasi-Kocal, G.; Rickelt, S.; Butty, V.L.; et al. Ketone body signaling mediates intestinal stem cell homeostasis and adaptation to diet. Cell 2019, 178, 1115–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aunan, J.R.; Cho, W.C.; Søreide, K. The biology of aging and cancer: A brief overview of shared and divergent molecular hallmarks. Aging Dis. 2017, 8, 628–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumari, S.; Badana, A.K.; Malla, R. Reactive oxygen species: A key constituent in cancer survival. Biomark Insights 2018, 13, 1177271918755391. [Google Scholar] [CrossRef] [Green Version]
- Klement, R.J.; Champ, C.E.; Otto, C.; Kämmerer, U. Anti-tumor effects of ketogenic diets in mice: A meta-analysis. PLoS ONE 2016, 11, e0155050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poff, A.M.; Ari, C.; Seyfried, T.N.; D’Agostino, D.P. The ketogenic diet and hyperbaric oxygen therapy prolong survival in mice with systemic metastatic cancer. PLoS ONE 2013, 8, e65522. [Google Scholar] [CrossRef] [Green Version]
- Otto, C.; Kaemmerer, U.; Illert, B.; Muehling, B.; Pfetzer, N.; Wittig, R.; Voelker, H.U.; Thiede, A.; Coy, J.F. Growth of human gastric cancer cells in nude mice is delayed by a ketogenic diet supplemented with omega-3 fatty acids and medium-chain triglycerides. BMC Cancer 2008, 8, 122. [Google Scholar] [CrossRef] [Green Version]
- Allen, B.G.; Bhatia, S.K.; Anderson, C.M.; Eichenberger-Gilmore, J.M.; Sibenaller, Z.A.; Mapuskar, K.A.; Schoenfeld, J.D.; Buatti, J.M.; Spitz, D.R.; Fath, M.A. Ketogenic diets as an adjuvant cancer therapy: History and potential mechanism. Redox Biol. 2014, 2, 963–970. [Google Scholar] [CrossRef] [Green Version]
- Klement, R.J.; Sweeney, R.A. Impact of a ketogenic diet intervention during radiotherapy on body composition: I. Initial clinical experience with six prospectively studied patients. BMC Res. Notes 2016, 9, 143. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.; Pfetzer, N.; Schwab, M.; Strauss, I.; Kämmerer, U. Effects of a ketogenic diet on the quality of life in 16 patients with advanced cancer: A pilot trial. Nutr. Metab. 2011, 8, 54. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Rissman, R.A.; Feng, J. Characterization of ATP alternations in an Alzheimer’s disease transgenic mouse model. J. Alzheimers Dis. 2015, 44, 375–378. [Google Scholar] [CrossRef] [Green Version]
- Greenamyre, J.T.; Sherer, T.B.; Betarbet, R.; Panov, A.V. Complex I and Parkinson’s disease. IUBMB Life 2001, 52, 135–141. [Google Scholar] [CrossRef]
- Mosconi, L.; de Leon, M.; Murray, J.; E., L.; Lu, J.; Javier, E.; McHugh, P.; Swerdlow, R.H. Reduced mitochondria cytochrome oxidase activity in adult children of mothers with Alzheimer’s disease. J. Alzheimers Dis. 2011, 27, 483–490. [Google Scholar] [CrossRef]
- Fortier, M.; Castellano, C.A.; Croteau, E.; Langlois, F.; Bocti, C.; St-Pierre, V.; Vandenberghe, C.; Bernier, M.; Roy, M.; Descoteaux, M.; et al. A ketogenic drink improves brain energy and some measures of cognition in mild cognitive impairment. Alzheimers Dement. 2019, 15, 625–634. [Google Scholar] [CrossRef]
- Srivastava, S.; Baxa, U.; Niu, G.; Chen, X.; Veech, R.L. A ketogenic diet increases brown adipose tissue mitochondrial proteins and UCP1 levels in mice. IUBMB Life 2013, 65, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhang, Q.; Tu, J.; Ren, Z. D-β-hydroxybutyrate inhibits microglial activation in a cell activation model in vitro. J. Med. Coll. PLA 2011, 26, 117–127. [Google Scholar] [CrossRef]
- Youm, Y.H.; Nguyen, K.Y.; Grant, R.W.; Goldberg, E.L.; Bodogai, M.; Kim, D.; D’Agostino, D.; Planavsky, N.; Lupfer, C.; Kanneganti, T.D.; et al. The ketone metabolite β-hydroxybutyrate blocks NLRP3 inflammasome-mediated inflammatory disease. Nat. Med. 2015, 21, 263–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.M.; Ramprasath, T.; Zou, M.H. β-hydroxybutyrate and its metabolic effects on age-associated pathology. Exp. Mol. Med. 2020, 52, 548–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campisi, J.; Kapahi, P.; Lithgow, G.J.; Melov, S.; Newman, J.C.; Verdin, E. From discoveries in ageing research to therapeutics for healthy ageing. Nature 2019, 571, 183–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abiri, B.; Vafa, M. Dietary Restriction, Cardiovascular Aging and Age-Related Cardiovascular Diseases: A Review of the Evidence. Adv. Exp. Med. Biol. 2019, 1178, 113–127. [Google Scholar] [PubMed]
- Newman, J.C.; Covarrubias, A.J.; Zhao, M.; Yu, X.; Gut, P.; Ng, C.P.; Huang, Y.; Haldar, S.; Verdin, E. Ketogenic Diet Reduces Midlife Mortality and Improves Memory in Aging Mice. Cell Metab. 2017, 26, 547–557.e548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shou, J.; Chen, P.J.; Xiao, W.H. Mechanism of increased risk of insulin resistance in aging skeletal muscle. Diabetol. Metab. Syndr. 2020, 12, 14. [Google Scholar] [CrossRef] [Green Version]
- Bonaldo, P.; Sandri, M. Cellular and molecular mechanisms of muscle atrophy. Dis. Model Mech. 2013, 6, 25–39. [Google Scholar] [CrossRef] [Green Version]
- Stubbs, B.J.; Koutnik, A.P.; Volek, J.S.; Newman, J.C. From bedside to battlefield: Intersection of ketone body mechanisms in geroscience with military resilience. GeroScience 2021, 43, 1071–1081. [Google Scholar] [CrossRef] [PubMed]
- Koutnik, A.P.; D’Agostino, D.P.; Egan, B. Anticatabolic Effects of Ketone Bodies in Skeletal Muscle. Trends Endocrinol. Metab. 2019, 30, 227–229. [Google Scholar] [CrossRef] [PubMed]
- Ahola-Erkkilä, S.; Carroll, C.J.; Peltola-Mjösund, K.; Tulkki, V.; Mattila, I.; Seppänen-Laakso, T.; Oresic, M.; Tyynismaa, H.; Suomalainen, A. Ketogenic diet slows down mitochondrial myopathy progression in mice. Hum. Mol. Genet. 2010, 19, 1974–1984. [Google Scholar] [CrossRef] [Green Version]
- Kwak, S.E.; Bae, J.H.; Lee, J.H.; Shin, H.E.; Zhang, D.; Cho, S.C.; Song, W. Effects of exercise-induced beta-hydroxybutyrate on muscle function and cognitive function. Physiol. Rep. 2021, 9, e14497. [Google Scholar] [CrossRef]
- Munroe, M.; Pincu, Y.; Merritt, J.; Cobert, A.; Brander, R.; Jensen, T.; Rhodes, J.; Boppart, M.D. Impact of β-hydroxy β-methylbutyrate (HMB) on age-related functional deficits in mice. Exp. Gerontol. 2017, 87, 57–66. [Google Scholar] [CrossRef]
- Walsh, M.E.; Bhattacharya, A.; Sataranatarajan, K.; Qaisar, R.; Sloane, L.; Rahman, M.M.; Kinter, M.; Van Remmen, H. The histone deacetylase inhibitor butyrate improves metabolism and reduces muscle atrophy during aging. Aging Cell 2015, 14, 957–970. [Google Scholar] [CrossRef]
- Angiolilli, C.; Baeten, D.L.; Radstake, T.R.; Reedquist, K.A. The acetyl code in rheumatoid arthritis and other rheumatic diseases. Epigenomics 2017, 9, 447–461. [Google Scholar] [CrossRef]
- Pawelec, G.; Goldeck, D.; Derhovanessian, E. Inflammation, ageing and chronic disease. Curr. Opin. Immunol. 2014, 29, 23–28. [Google Scholar] [CrossRef]
- Chamorro, A.; Hallenbeck, J. The harms and benefits of inflammatory and immune responses in vascular disease. Stroke 2006, 37, 291–293. [Google Scholar] [CrossRef] [PubMed]
- Praticò, D.; Trojanowski, J.Q. Inflammatory hypotheses: Novel mechanisms of Alzheimer’s neurodegeneration and new therapeutic targets? Neurobiol. Aging 2000, 21, 441–453. [Google Scholar] [CrossRef]
- Pasyukova, E.G.; Vaiserman, A.M. HDAC inhibitors: A new promising drug class in anti-aging research. Mech. Ageing Dev. 2017, 166, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Camberos-Luna, L.; Massieu, L. Therapeutic strategies for ketosis induction and their potential efficacy for the treatment of acute brain injury and neurodegenerative diseases. Neurochem. Int. 2020, 133, 104614. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Hao, J.; Liu, R.; Turner, G.; Shi, F.D.; Rho, J.M. Inflammation-mediated memory dysfunction and effects of a ketogenic diet in a murine model of multiple sclerosis. PLoS ONE 2012, 7, e35476. [Google Scholar] [CrossRef] [Green Version]
- Alberti, K.G.; Zimmet, P.; Shaw, J. The metabolic syndrome--a new worldwide definition. Lancet 2005, 366, 1059–1062. [Google Scholar] [CrossRef]
- Ron, D.; Walter, P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat. Rev. Mol. Cell Biol. 2007, 8, 519–529. [Google Scholar] [CrossRef]
- Nunes-Souza, V.; César-Gomes, C.J.; Da Fonseca, L.J.; Guedes Gda, S.; Smaniotto, S.; Rabelo, L.A. Aging Increases Susceptibility to High Fat Diet-Induced Metabolic Syndrome in C57BL/6 Mice: Improvement in Glycemic and Lipid Profile after Antioxidant Therapy. Oxid. Med. Cell Longev. 2016, 2016, 1987960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mey, J.T.; Erickson, M.L.; Axelrod, C.L.; King, W.T.; Flask, C.A.; McCullough, A.J.; Kirwan, J.P. β-Hydroxybutyrate is reduced in humans with obesity-related NAFLD and displays a dose-dependent effect on skeletal muscle mitochondrial respiration in vitro. Am. J. Physiol. Endocrinol. Metab. 2020, 319, e187–e195. [Google Scholar] [CrossRef] [PubMed]
- Cavaleri, F.; Bashar, E. Potential Synergies of β-Hydroxybutyrate and Butyrate on the Modulation of Metabolism, Inflammation, Cognition, and General Health. J. Nutr. Metab. 2018, 2018, 7195760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.K.; Kim, D.H.; Bang, E.; Choi, Y.J.; Chung, H.Y. β-Hydroxybutyrate Suppresses Lipid Accumulation in Aged Liver through GPR109A-mediated Signaling. Aging Dis. 2020, 11, 777–790. [Google Scholar] [CrossRef]
- Bae, H.R.; Kim, D.H.; Park, M.H.; Lee, B.; Kim, M.J.; Lee, E.K.; Chung, K.W.; Kim, S.M.; Im, D.S.; Chung, H.Y. β-Hydroxybutyrate suppresses inflammasome formation by ameliorating endoplasmic reticulum stress via AMPK activation. Oncotarget 2016, 7, 66444–66454. [Google Scholar] [CrossRef] [Green Version]
- Møller, N. Ketone Body, 3-Hydroxybutyrate: Minor Metabolite—Major Medical Manifestations. J. Clin. Endocrinol. Metab. 2020, 105, dgaa370. [Google Scholar] [CrossRef]
- Paoli, A.; Rubini, A.; Volek, J.S.; Grimaldi, K.A. Beyond weight loss: A review of the therapeutic uses of very-low-carbohydrate (ketogenic) diets. Eur. J. Clin. Nutr. 2013, 67, 789–796. [Google Scholar] [CrossRef] [Green Version]
- Fontana, L. Excessive adiposity, calorie restriction, and aging. JAMA 2006, 295, 1577–1578. [Google Scholar] [CrossRef]
- Kim, B.H.; Joo, Y.; Kim, M.-S.; Choe, H.K.; Tong, Q.; Kwon, O. Effects of Intermittent Fasting on the Circulating Levels and Circadian Rhythms of Hormones. Endocrinol. Metab. 2021, 36, 745–756. [Google Scholar] [CrossRef]
- Hutchison, A.T.; Liu, B.; Wood, R.E.; Vincent, A.D.; Thompson, C.H.; O’Callaghan, N.J.; Wittert, G.A.; Heilbronn, L.K. Effects of Intermittent Versus Continuous Energy Intakes on Insulin Sensitivity and Metabolic Risk in Women with Overweight. Obesity 2019, 27, 50–58. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Arbelaez, D.; Bellido, D.; Castro, A.I.; Ordoñez-Mayan, L.; Carreira, J.; Galban, C.; Martinez-Olmos, M.A.; Crujeiras, A.B.; Sajoux, I.; Casanueva, F.F. Body composition changes after very-low-calorie ketogenic diet in obesity evaluated by 3 standardized methods. J. Clin. Endocrinol. Metab. 2017, 102, 488–498. [Google Scholar] [CrossRef]
- Stewart, W.K.; Fleming, L.W. Features of a successful therapeutic fast of 382 days’ duration. Postgrad. Med. J. 1973, 49, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Brandhorst, S.; Choi, I.Y.; Wei, M.; Cheng, C.W.; Sedrakyan, S.; Navarrete, G.; Dubeau, L.; Yap, L.P.; Park, R.; Vinciguerra, M.; et al. A Periodic Diet that Mimics Fasting Promotes Multi-System Regeneration, Enhanced Cognitive Performance, and Healthspan. Cell Metab. 2015, 22, 86–99. [Google Scholar] [CrossRef] [Green Version]
- Bordone, L.; Guarente, L. Calorie restriction, SIRT1 and metabolism: Understanding longevity. Nat. Rev. Mol. Cell Biol. 2005, 6, 298–305. [Google Scholar] [CrossRef]
- Pignatti, C.; D’Adamo, S.; Stefanelli, C.; Flamigni, F.; Cetrullo, S. Nutrients and Pathways that Regulate Health Span and Life Span. Geriatrics 2020, 5, 95. [Google Scholar] [CrossRef]
- Fontana, L.; Partridge, L.; Longo, V.D. Extending healthy life span--from yeast to humans. Science 2010, 328, 321–326. [Google Scholar] [CrossRef] [Green Version]
- Abdul Kadir, A.; Clarke, K.; Evans, R.D. Cardiac ketone body metabolism. Biochim. Biophys. Acta. Mol. Basis Dis. 2020, 1866, 165739. [Google Scholar] [CrossRef]
- Wallace, M.A.; Aguirre, N.W.; Marcotte, G.R.; Marshall, A.G.; Baehr, L.M.; Hughes, D.C.; Hamilton, K.L.; Roberts, M.N.; Lopez-Dominguez, J.A.; Miller, B.F.; et al. The ketogenic diet preserves skeletal muscle with aging in mice. Aging Cell 2021, 20, e13322. [Google Scholar] [CrossRef]
- O’Neill, B.; Raggi, P. The ketogenic diet: Pros and cons. Atherosclerosis 2020, 292, 119–126. [Google Scholar] [CrossRef] [Green Version]
- Ari, C.; Kovács, Z.; Juhasz, G.; Murdun, C.; Goldhagen, C.R.; Koutnik, A.P.; Poff, A.M.; Kesl, S.L.; D’Agostino, D.P. Exogenous Ketone Supplements Reduce Anxiety-Related Behavior in Sprague-Dawley and Wistar Albino Glaxo/Rijswijk Rats. Front. Mol. Neurosci. 2016, 9, 137. [Google Scholar] [CrossRef] [Green Version]
- CJ, D.C.H.; Schofield, G.M.; Williden, M.; McQuillan, J.A. The effect of medium chain triglycerides on time to nutritional ketosis and symptoms of keto-induction in healthy adults: A randomised controlled clinical trial. J. Nutr. Metab. 2018, 2018, 2630565. [Google Scholar]
- Kesl, S.L.; Poff, A.M.; Ward, N.P.; Fiorelli, T.N.; Ari, C.; Van Putten, A.J.; Sherwood, J.W.; Arnold, P.; D’Agostino, D.P. Effects of exogenous ketone supplementation on blood ketone, glucose, triglyceride, and lipoprotein levels in Sprague-Dawley rats. Nutr. Metab. 2016, 13, 9. [Google Scholar] [CrossRef] [Green Version]
- Brunengraber, H. Potential of ketone body esters for parenteral and oral nutrition. Nutrition 1997, 13, 233–235. [Google Scholar] [CrossRef]
- Ari, C.; Murdun, C.; Koutnik, A.P.; Goldhagen, C.R.; Rogers, C.; Park, C.; Bharwani, S.; Diamond, D.M.; Kindy, M.S.; D’Agostino, D.P.; et al. Exogenous Ketones Lower Blood Glucose Level in Rested and Exercised Rodent Models. Nutrients 2019, 11, 2330. [Google Scholar] [CrossRef] [Green Version]
- Veech, R.L. The therapeutic implications of ketone bodies: The effects of ketone bodies in pathological conditions: Ketosis, ketogenic diet, redox states, insulin resistance, and mitochondrial metabolism. Prostaglandins Leukot. Essent. Fat. Acids 2004, 70, 309–319. [Google Scholar] [CrossRef]
- Stubbs, B.J.; Blade, T.; Mills, S.; Thomas, J.; Yufei, X.; Nelson, F.R.; Higley, N.; Nikiforov, A.I.; Rhiner, M.O.; Verdin, E.; et al. In vitro stability and in vivo pharmacokinetics of the novel ketogenic ester, bis hexanoyl (R)-1,3-butanediol. Food Chem. Toxicol. 2021, 147, 111859. [Google Scholar] [CrossRef]
- Clarke, K.; Tchabanenko, K.; Pawlosky, R.; Carter, E.; Todd King, M.; Musa-Veloso, K.; Ho, M.; Roberts, A.; Robertson, J.; Vanitallie, T.B.; et al. Kinetics, safety and tolerability of (R)-3-hydroxybutyl (R)-3-hydroxybutyrate in healthy adult subjects. Regul. Toxicol. Pharm. 2012, 63, 401–408. [Google Scholar] [CrossRef] [Green Version]
- Stubbs, B.J.; Cox, P.J.; Evans, R.D.; Santer, P.; Miller, J.J.; Faull, O.K.; Magor-Elliott, S.; Hiyama, S.; Stirling, M.; Clarke, K. On the metabolism of exogenous ketones in humans. Front. Physiol. 2017, 8, 848. [Google Scholar] [CrossRef]
- Stubbs, B.J.; Cox, P.J.; Evans, R.D.; Cyranka, M.; Clarke, K.; de Wet, H. A ketone ester drink lowers human ghrelin and appetite. Obesity 2018, 26, 269–273. [Google Scholar] [CrossRef]
- Myette-Côté, É.; Neudorf, H.; Rafiei, H.; Clarke, K.; Little, J.P. Prior ingestion of exogenous ketone monoester attenuates the glycaemic response to an oral glucose tolerance test in healthy young individuals. J. Physiol. 2018, 596, 1385–1395. [Google Scholar] [CrossRef]
- Soto-Mota, A.; Vansant, H.; Evans, R.D.; Clarke, K. Safety and tolerability of sustained exogenous ketosis using ketone monoester drinks for 28 days in healthy adults. Regul. Toxicol. Pharm. 2019, 109, 104506. [Google Scholar] [CrossRef]
- Schönfeld, P.; Wojtczak, L. Short- and medium-chain fatty acids in energy metabolism: The cellular perspective. J. Lipid Res. 2016, 57, 943–954. [Google Scholar] [CrossRef] [Green Version]
- Huttenlocher, P.R.; Wilbourn, A.J.; Signore, J.M. Medium-chain triglycerides as a therapy for intractable childhood epilepsy. Neurology 1971, 21, 1097–1103. [Google Scholar] [CrossRef]
- Thorburn, M.S.; Vistisen, B.; Thorp, R.M.; Rockell, M.J.; Jeukendrup, A.E.; Xu, X.; Rowlands, D.S. Attenuated gastric distress but no benefit to performance with adaptation to octanoate-rich esterified oils in well-trained male cyclists. J. Appl. Physiol. (1985) 2006, 101, 1733–1743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, S.T. Ketone bodies as a therapeutic for Alzheimer’s disease. Neurotherapeutics 2008, 5, 470–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dymsza, H.A. Nutritional application and implication of 1,3-butanediol. Fed. Proc. 1975, 34, 2167–2170. [Google Scholar] [PubMed]
- Puchowicz, M.A.; Smith, C.L.; Bomont, C.; Koshy, J.; David, F.; Brunengraber, H. Dog model of therapeutic ketosis Induced by oral administration of R,S-1,3-butanediol diacetoacetate. J. Nutr. Biochem. 2000, 11, 281–287. [Google Scholar] [CrossRef]
- D’Agostino, D.P.; Pilla, R.; Held, H.E.; Landon, C.S.; Puchowicz, M.; Brunengraber, H.; Ari, C.; Arnold, P.; Dean, J.B. Therapeutic ketosis with ketone ester delays central nervous system oxygen toxicity seizures in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R829–R836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashim, S.A.; VanItallie, T.B. Ketone body therapy: From the ketogenic diet to the oral administration of ketone ester. J. Lipid Res. 2014, 55, 1818–1826. [Google Scholar] [CrossRef] [Green Version]
- Scala, R.A.; Paynter, O.E. Chronic oral toxicity of 1,3-butanediol. Toxicol. Appl. Pharmacol. 1967, 10, 160–164. [Google Scholar] [CrossRef]
- Desrochers, S.; David, F.; Garneau, M.; Jetté, M.; Brunengraber, H. Metabolism of R- and S-1,3-butanediol in perfused livers from meal-fed and starved rats. Biochem. J. 1992, 285 Pt 2, 647–653. [Google Scholar] [CrossRef] [Green Version]
- Newman, J.C.; Verdin, E. β-Hydroxybutyrate: A signaling metabolite. Annu. Rev. Nutr. 2017, 37, 51–76. [Google Scholar] [CrossRef]
- Desrochers, S.; Dubreuil, P.; Brunet, J.; Jetté, M.; David, F.; Landau, B.R.; Brunengraber, H. Metabolism of (R,S)-1,3-butanediol acetoacetate esters, potential parenteral and enteral nutrients in conscious pigs. Am. J. Physiol. 1995, 268, E660–E667. [Google Scholar] [CrossRef]
- Webber, R.J.; Edmond, J. Utilization of L(+)-3-hydroxybutyrate, D(-)-3-hydroxybutyrate, acetoacetate, and glucose for respiration and lipid synthesis in the 18-day-old rat. J. Biol. Chem. 1977, 252, 5222–5226. [Google Scholar] [CrossRef]
- Valenzuela, P.L.; Morales, J.S.; Castillo-García, A.; Lucia, A. Acute Ketone Supplementation and Exercise Performance: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Int. J. Sports Physiol. Perform. 2020, 15, 298–308. [Google Scholar] [CrossRef]
- Thomsen, H.H.; Rittig, N.; Johannsen, M.; Møller, A.B.; Jørgensen, J.O.; Jessen, N.; Møller, N. Effects of 3-hydroxybutyrate and free fatty acids on muscle protein kinetics and signaling during LPS-induced inflammation in humans: Anticatabolic impact of ketone bodies. Am. J. Clin. Nutr. 2018, 108, 857–867. [Google Scholar] [CrossRef] [Green Version]
- Phinney, S.D.; Bistrian, B.R.; Wolfe, R.R.; Blackburn, G.L. The human metabolic response to chronic ketosis without caloric restriction: Physical and biochemical adaptation. Metabolism 1983, 32, 757–768. [Google Scholar] [CrossRef]
- Burke, L.M.; Ross, M.L.; Garvican-Lewis, L.A.; Welvaert, M.; Heikura, I.A.; Forbes, S.G.; Mirtschin, J.G.; Cato, L.E.; Strobel, N.; Sharma, A.P.; et al. Low carbohydrate, high fat diet impairs exercise economy and negates the performance benefit from intensified training in elite race walkers. J. Physiol. 2017, 595, 2785–2807. [Google Scholar] [CrossRef] [Green Version]
- Cox, P.J.; Kirk, T.; Ashmore, T.; Willerton, K.; Evans, R.; Smith, A.; Murray, A.J.; Stubbs, B.; West, J.; McLure, S.W.; et al. Nutritional Ketosis Alters Fuel Preference and Thereby Endurance Performance in Athletes. Cell Metab. 2016, 24, 256–268. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Structure of ketone bodies.

Figure 2.
Pathways of ketogenesis in liver and ketolysis in extrahepatic tissues. Hepatic mitochondria act as the primary site for the synthesis of blood ketone bodies by using fatty acids-derived Ac-CoA which is generated by β oxidation. The following process requires four enzymes: ACAT, HMGCS2, HMGCL, and BDH1, and the intermediate product covers AcAc-CoA, HMG-CoA, and AcAc. β-HB is finally produced and released to the bloodstream and uptaken by extrahepatic tissues such as the brain, heart, neurons, kidneys, and muscles through MCT. Both β-HB and AcAc can be oxidized and converted to Ac-CoA and produce ATP via the TCA cycle as an alternative energy source. Abbreviations: AcAc, acetoacetate; AcAc-CoA, acetoacetyl CoA; ACAT, acetyl-CoA A acetyltransferase; Ac-CoA, acetyl CoA; BDH1, β-hydroxybutyrate dehydrogenase 1; β-HB, β-hydroxybutyrate; βox, β oxidation; CS, citrate synthase; FFA, free fatty acid; HMGCL, HMG-CoA lyase; HMGCS2, 3-hydroxymethylglutaryl-CoA synthase 2; HMG-CoA, 3-hydroxymethylglutaryl-CoA; MCT, monocarboxylate transporter; SCOT, succinyl-CoA:3-oxoacid CoA transferase; TCA, tricarboxylic acid.
Figure 2.
Pathways of ketogenesis in liver and ketolysis in extrahepatic tissues. Hepatic mitochondria act as the primary site for the synthesis of blood ketone bodies by using fatty acids-derived Ac-CoA which is generated by β oxidation. The following process requires four enzymes: ACAT, HMGCS2, HMGCL, and BDH1, and the intermediate product covers AcAc-CoA, HMG-CoA, and AcAc. β-HB is finally produced and released to the bloodstream and uptaken by extrahepatic tissues such as the brain, heart, neurons, kidneys, and muscles through MCT. Both β-HB and AcAc can be oxidized and converted to Ac-CoA and produce ATP via the TCA cycle as an alternative energy source. Abbreviations: AcAc, acetoacetate; AcAc-CoA, acetoacetyl CoA; ACAT, acetyl-CoA A acetyltransferase; Ac-CoA, acetyl CoA; BDH1, β-hydroxybutyrate dehydrogenase 1; β-HB, β-hydroxybutyrate; βox, β oxidation; CS, citrate synthase; FFA, free fatty acid; HMGCL, HMG-CoA lyase; HMGCS2, 3-hydroxymethylglutaryl-CoA synthase 2; HMG-CoA, 3-hydroxymethylglutaryl-CoA; MCT, monocarboxylate transporter; SCOT, succinyl-CoA:3-oxoacid CoA transferase; TCA, tricarboxylic acid.

Figure 3.
The metabolic role of β-HB. β-HB acts as an energy substrate to regulate metabolic response, and serves as a signaling molecule to inhibit lipolysis, oxidative stress and improve neuroprotection, it also plays a role in epigenetic regulation including histone methylation, inhibition of HDACs, and histone lysine β-hydroxybutyrylation. Abbreviations: β-HB, β-hydroxybutyrate; HDAC, histone deacetylases.
Figure 3.
The metabolic role of β-HB. β-HB acts as an energy substrate to regulate metabolic response, and serves as a signaling molecule to inhibit lipolysis, oxidative stress and improve neuroprotection, it also plays a role in epigenetic regulation including histone methylation, inhibition of HDACs, and histone lysine β-hydroxybutyrylation. Abbreviations: β-HB, β-hydroxybutyrate; HDAC, histone deacetylases.

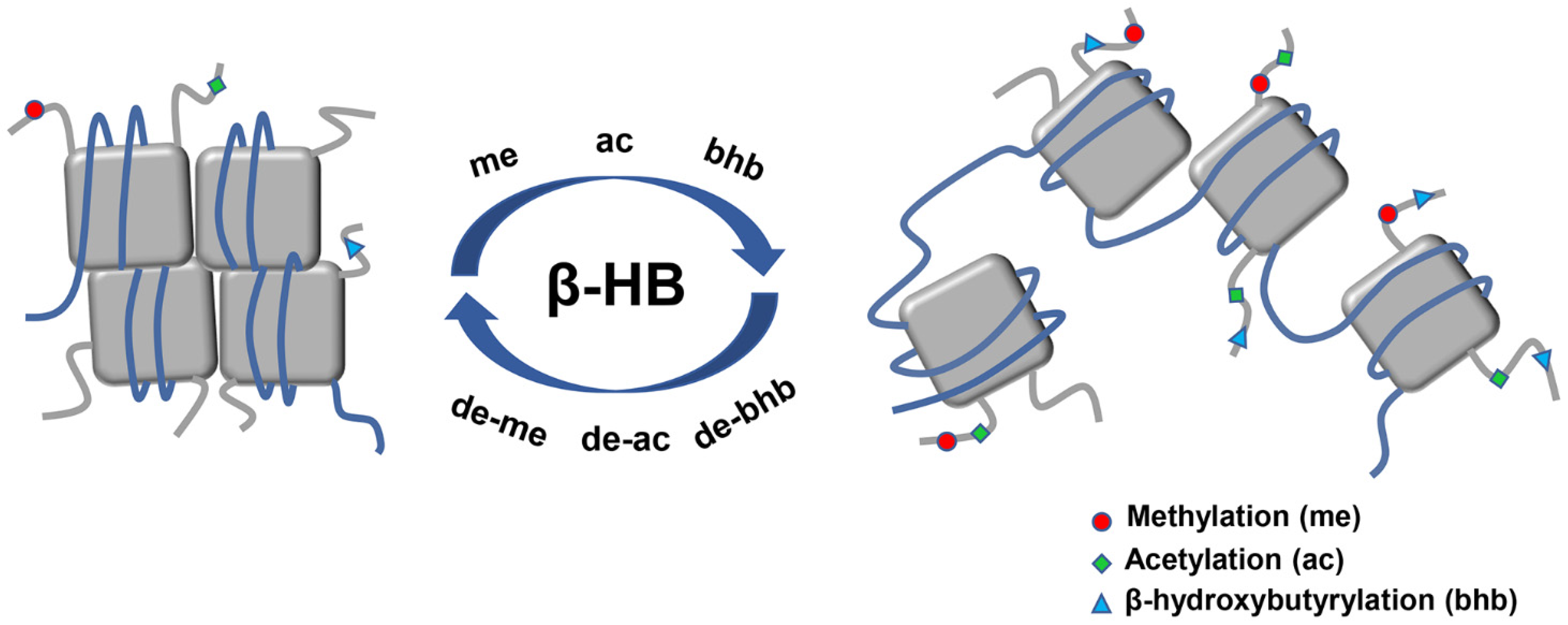
Figure 4.
Main epigenetic alterations of histone posttranslational modifications induced by β-HB. Histone posttranslational modifications which relate to the epigenetic regulation of β-HB mainly consist of histone methylation, acetylation, as well as a new type of histone lysine bhb. β-HB administration alters the modification of chromatin conformation through the epigenetic pathway. Chromatin structure is represented by DNA (blue line) organized in nucleosomes formed by core histones (grey square). Abbreviations: ac, acetylation; bhb, β-hydroxybutyrylation; β-HB, β-hydroxybutyrate; me, methylation.
Figure 4.
Main epigenetic alterations of histone posttranslational modifications induced by β-HB. Histone posttranslational modifications which relate to the epigenetic regulation of β-HB mainly consist of histone methylation, acetylation, as well as a new type of histone lysine bhb. β-HB administration alters the modification of chromatin conformation through the epigenetic pathway. Chromatin structure is represented by DNA (blue line) organized in nucleosomes formed by core histones (grey square). Abbreviations: ac, acetylation; bhb, β-hydroxybutyrylation; β-HB, β-hydroxybutyrate; me, methylation.

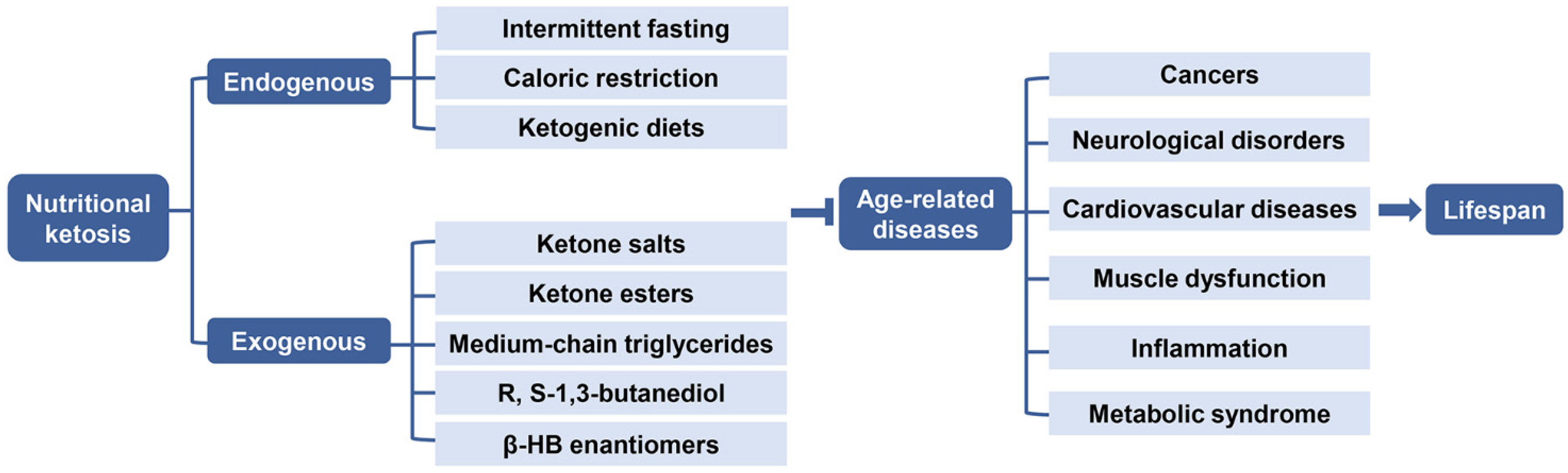
Figure 5.
Role of endogenous and exogenous nutritional ketosis in age-related diseases. Nutritional ketosis can be achieved endogenously through intermittent fasting, caloric restriction, ketogenic diets or exogenously with oral supplementation via ketone salts, ketone esters, medium-chain triglycerides, R, S-1,3- butanediol, and β-HB enantiomers, resulting in a therapeutic effect in several medical conditions, thereby targeting the underlying mechanisms of age-related diseases, such as cancers, neurological disorders, cardiovascular diseases, muscle dysfunction, inflammation, and metabolic syndrome, which may extend the healthy life expectancy. Abbreviations: β-HB, β-hydroxybutyrate.
Figure 5.
Role of endogenous and exogenous nutritional ketosis in age-related diseases. Nutritional ketosis can be achieved endogenously through intermittent fasting, caloric restriction, ketogenic diets or exogenously with oral supplementation via ketone salts, ketone esters, medium-chain triglycerides, R, S-1,3- butanediol, and β-HB enantiomers, resulting in a therapeutic effect in several medical conditions, thereby targeting the underlying mechanisms of age-related diseases, such as cancers, neurological disorders, cardiovascular diseases, muscle dysfunction, inflammation, and metabolic syndrome, which may extend the healthy life expectancy. Abbreviations: β-HB, β-hydroxybutyrate.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, L.; Chen, P.; Xiao, W. β-hydroxybutyrate as an Anti-Aging Metabolite. Nutrients 2021, 13, 3420. https://doi.org/10.3390/nu13103420
AMA Style
Wang L, Chen P, Xiao W. β-hydroxybutyrate as an Anti-Aging Metabolite. Nutrients. 2021; 13(10):3420. https://doi.org/10.3390/nu13103420
Chicago/Turabian StyleWang, Lian, Peijie Chen, and Weihua Xiao. 2021. "β-hydroxybutyrate as an Anti-Aging Metabolite" Nutrients 13, no. 10: 3420. https://doi.org/10.3390/nu13103420
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.