
Associations between Postprandial Gut Hormones and Markers of Bone Remodeling
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Measurements
2.3. Statistical Analyses
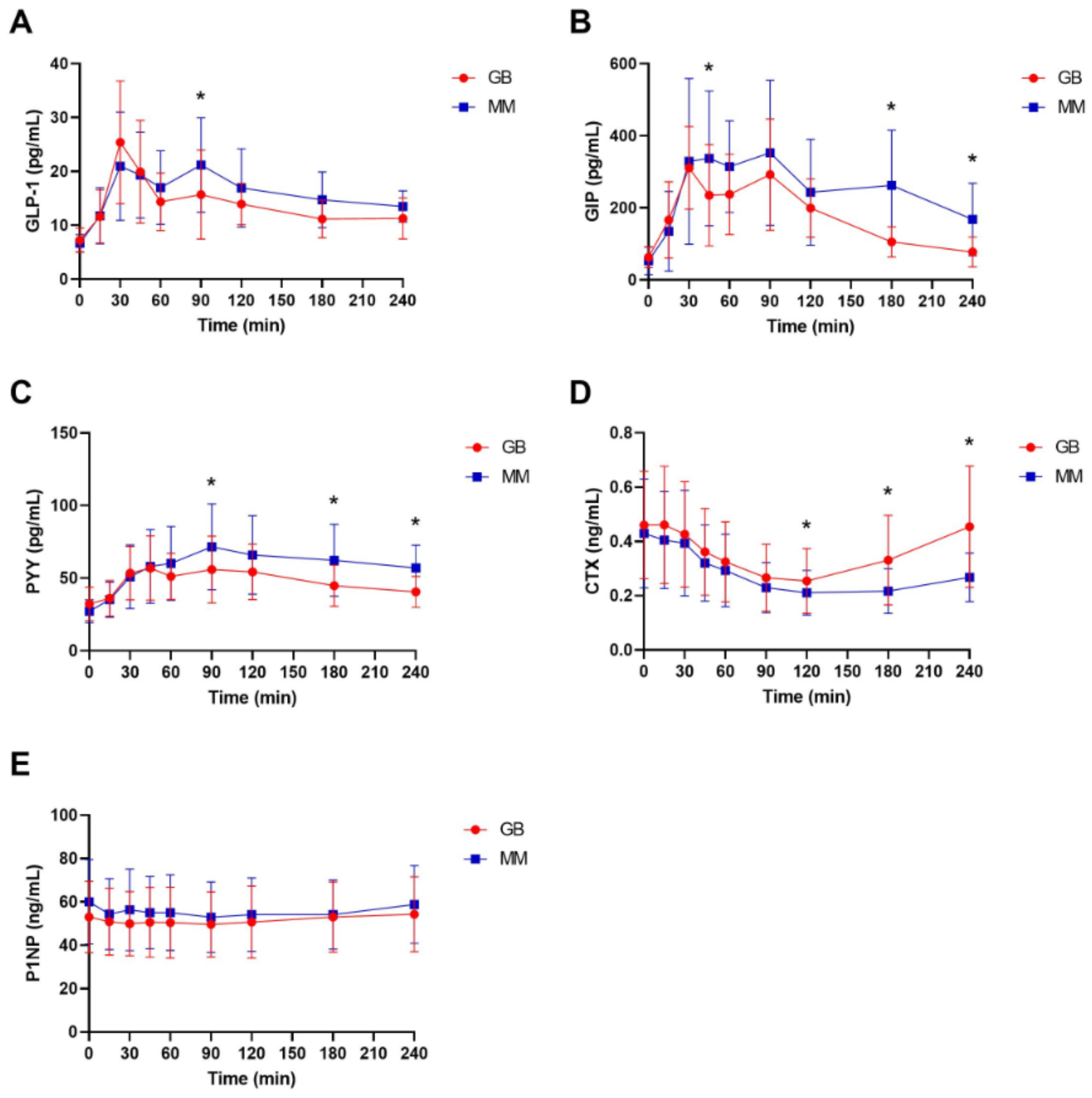
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clarke, B. Normal Bone Anatomy and Physiology. Clin. J. Am. Soc. Nephrol. 2008, 3 (Suppl. 3), S131–S139. [Google Scholar] [CrossRef] [Green Version]
- Greenblatt, M.B.; Tsai, J.N.; Wein, M.N. Bone Turnover Markers in the Diagnosis and Monitoring of Metabolic Bone Disease. Clin. Chem. 2017, 63, 464–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattoa, H.P.; Cavalier, E.; Eastell, R.; Heijboer, A.C.; Jørgensen, N.R.; Makris, K.; Ulmer, C.Z.; Kanis, J.A.; Cooper, C.; Silverman, S.L.; et al. Analytical considerations and plans to standardize or harmonize assays for the reference bone turnover markers PINP and β-CTX in blood. Clin. Chim. Acta 2020, 515, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Vasikaran, S.; Cooper, C.; Eastell, R.; Griesmacher, A.; Morris, H.A.; Trenti, T.; Kanis, J.A. International Osteoporosis Foundation and International Federation of Clinical Chemistry and Laboratory Medicine Position on bone marker standards in osteoporosis. Clin. Chem. Lab. Med. 2011, 49, 1271–1274. [Google Scholar] [CrossRef] [PubMed]
- Clowes, J.; Hannon, R.; Yap, T.; Hoyle, N.; Blumsohn, A.; Eastell, R. Effect of feeding on bone turnover markers and its impact on biological variability of measurements. Bone 2002, 30, 886–890. [Google Scholar] [CrossRef]
- Schiellerup, S.P.; Skov-Jeppesen, K.; Windeløv, J.A.; Svane, M.S.; Holst, J.J.; Hartmann, B.; Rosenkilde, M.M. Gut Hormones and Their Effect on Bone Metabolism. Potential Drug Therapies in Future Osteoporosis Treatment. Front. Endocrinol. 2019, 10, 75. [Google Scholar] [CrossRef]
- Henriksen, D.B.; Alexandersen, P.; Bjarnason, N.H.; Vilsbøll, T.; Hartmann, B.; Henriksen, E.E.G.; Byrjalsen, I.; Krarup, T.; Holst, J.J.; Christiansen, C. Role of Gastrointestinal Hormones in Postprandial Reduction of Bone Resorption. J. Bone Miner. Res. 2003, 18, 2180–2189. [Google Scholar] [CrossRef]
- Wichers, M.; Schmidt, E.; Bidlingmaier, F.; Klingmüller, D. Diurnal Rhythm of CrossLaps in Human Serum. Clin. Chem. 1999, 45, 1858–1860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diakogiannaki, E.; Gribble, F.; Reimann, F. Nutrient detection by incretin hormone secreting cells. Physiol. Behav. 2012, 106, 387–393. [Google Scholar] [CrossRef] [Green Version]
- Nissen, A.; Marstrand, S.; Skov-Jeppesen, K.; Bremholm, L.; Hornum, M.; Andersen, U.B.; Holst, J.J.; Rosenkilde, M.M.; Hartmann, B. A Pilot Study Showing Acute Inhibitory Effect of GLP-1 on the Bone Resorption Marker CTX in Humans. JBMR Plus 2019, 3, e10209. [Google Scholar] [CrossRef]
- Bergmann, N.C.; Lund, A.; Gasbjerg, L.S.; Jørgensen, N.R.; Jessen, L.; Hartmann, B.; Holst, J.J.; Christensen, M.B.; Vilsbøll, T.; Knop, F.K. Separate and Combined Effects of GIP and GLP-1 Infusions on Bone Metabolism in Overweight Men without Diabetes. J. Clin. Endocrinol. Metab. 2019, 104, 2953–2960. [Google Scholar] [CrossRef]
- Christensen, M.; Lund, A.; Calanna, S.; Jørgensen, N.R.; Holst, J.J.; Vilsbøll, T.; Knop, F.K. Glucose-Dependent Insulinotropic Polypeptide (GIP) Inhibits Bone Resorption Independently of Insulin and Glycemia. J. Clin. Endocrinol. Metab. 2017, 103, 288–294. [Google Scholar] [CrossRef] [Green Version]
- Gasbjerg, L.S.; Hartmann, B.; Christensen, M.B.; Lanng, A.R.; Vilsbøll, T.; Jørgensen, N.R.; Holst, J.J.; Rosenkilde, M.M.; Knop, F.K. GIP’s effect on bone metabolism is reduced by the selective GIP receptor antagonist GIP(3–30)NH2. Bone 2020, 130, 115079. [Google Scholar] [CrossRef]
- Iepsen, E.W.; Lundgren, J.R.; Hartmann, B.; Pedersen, O.; Hansen, T.; Jørgensen, N.R.; Jensen, J.-E.B.; Holst, J.J.; Madsbad, S.; Torekov, S. GLP-1 Receptor Agonist Treatment Increases Bone Formation and Prevents Bone Loss in Weight-Reduced Obese Women. J. Clin. Endocrinol. Metab. 2015, 100, 2909–2917. [Google Scholar] [CrossRef]
- Quist, J.S.; Jensen, M.M.; Clemmensen, K.K.B.; Pedersen, H.; Bjerre, N.; Størling, J.; Blond, M.B.; Albrechtsen, N.J.W.; Holst, J.J.; Torekov, S.S.; et al. Protocol for a single-centre, parallel-group, randomised, controlled, superiority trial on the effects of time-restricted eating on body weight, behaviour and metabolism in individuals at high risk of type 2 diabetes: The REStricted Eating Time (RESET) study. BMJ Open 2020, 10, e037166. [Google Scholar] [CrossRef]
- Mottalib, A.; Abrahamson, M.J.; Pober, D.M.; Polak, R.; Eldib, A.H.; Tomah, S.; Ashrafzadeh, S.; Hamdy, O. Effect of diabetes-specific nutrition formulas on satiety and hunger hormones in patients with type 2 diabetes. Nutr. Diabetes 2019, 9, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobas. PreciControl Varia 05618860190 05618860922. 2021;0561886092:1–2. Control Material LOT: 431664, pp. 1–2.
- Deacon, C.F.; Plamboeck, A.; Rosenkilde, M.M.; De Heer, J.; Holst, J.J. GIP-(3–42) does not antagonize insulinotropic effects of GIP at physiological concentrations. Am. J. Physiol. Metab. 2006, 291, E468–E475. [Google Scholar] [CrossRef] [PubMed]
- Cummings, D.E.; Overduin, J. Gastrointestinal regulation of food intake. J. Clin. Investig. 2007, 117, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Christensen, M.B.; Lund, A.B.; Jørgensen, N.R.; Holst, J.J.; Vilsbøll, T.; Knop, F.K. Glucose-Dependent Insulinotropic Polypeptide (GIP) Reduces Bone Resorption in Patients with Type 2 Diabetes. J. Endocr. Soc. 2020, 4, bvaa097. [Google Scholar] [CrossRef]
- Westberg-Rasmussen, S.; Starup-Linde, J.; Hermansen, K.; Holst, J.J.; Hartmann, B.; Vestergaard, P.; Gregersen, S. Differential impact of glucose administered intravenously or orally on bone turnover markers in healthy male subjects. Bone 2017, 97, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Pacheco-Pantoja, E.L.; Ranganath, L.R.; Gallagher, J.A.; Wilson, P.J.M.; Fraser, W.D. Receptors and effects of gut hormones in three osteoblastic cell lines. BMC Physiol. 2011, 11, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bollag, R.J.; Zhong, Q.; Phillips, P.; Min, L.; Zhong, L.; Cameron, R.; Mulloy, A.L.; Rasmussen, H.; Qin, F.; Ding, K.H.; et al. Osteoblast-Derived Cells Express Functional Glucose-Dependent Insulinotropic Peptide Receptors1. Endocrinology 2000, 141, 1228–1235. [Google Scholar] [CrossRef] [PubMed]
- Aoyama, E.; Watari, I.; Podyma-Inoue, K.A.; Yanagishita, M.; Ono, T. Expression of glucagon-like peptide-1 receptor and glucose-dependent insulinotropic polypeptide receptor is regulated by the glucose concentration in mouse osteoblastic MC3T3-E1 cells. Int. J. Mol. Med. 2014, 34, 475–482. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.; Ma, X.; Wang, N.; Jia, M.; Bi, L.; Wang, Y.; Li, M.; Zhang, H.; Xue, X.; Hou, Z.; et al. Activation of GLP-1 Receptor Promotes Bone Marrow Stromal Cell Osteogenic Differentiation through β-Catenin. Stem Cell Rep. 2016, 6, 579–591. [Google Scholar] [CrossRef] [Green Version]
- Fuglsang-Nielsen, R.; Rakvaag, E.; Vestergaard, P.; Hartmann, B.; Holst, J.J.; Hermansen, K.; Gregersen, S.; Starup-Linde, J. Consumption of nutrients and insulin resistance suppress markers of bone turnover in subjects with abdominal obesity. Bone 2020, 133, 115230. [Google Scholar] [CrossRef] [PubMed]
- Bjørnshave, A.; Hermansen, K.; Holst, J.J. Pre-Meal Effect of Whey Proteins on Metabolic Parameters in Subjects with and without Type 2 Diabetes: A Randomized, Crossover Trial. Nutrients 2018, 10, 122. [Google Scholar] [CrossRef] [Green Version]
- Heikura, I.; Burke, L.M.; Hawley, J.; Ross, M.L.; Garvican-Lewis, L.; Sharma, A.P.; McKay, A.K.A.; Leckey, J.J.; Welvaert, M.; McCall, L.; et al. A Short-Term Ketogenic Diet Impairs Markers of Bone Health in Response to Exercise. Front. Endocrinol. 2020, 10, 880. [Google Scholar] [CrossRef] [Green Version]
- Hansen, M.S.; Tencerova, M.; Frølich, J.; Kassem, M.; Frost, M.; Terencova, M. Effects of gastric inhibitory polypeptide, glucagon-like peptide-1 and glucagon-like peptide-1 receptor agonists on Bone Cell Metabolism. Basic Clin. Pharmacol. Toxicol. 2018, 122, 25–37. [Google Scholar] [CrossRef] [Green Version]
- Leitch, V.D.; Brassill, M.J.; Rahman, S.; Butterfield, N.C.; Ma, P.; Logan, J.G.; Boyde, A.; Evans, H.; Croucher, P.I.; Batterham, R.L.; et al. PYY is a negative regulator of bone mass and strength. Bone 2019, 127, 427–435. [Google Scholar] [CrossRef]
- Wong, I.P.L.; Driessler, F.; Khor, E.C.; Shi, Y.-C.; Hörmer, B.; Nguyen, A.D.; Enriquez, R.F.; Eisman, J.A.; Sainsbury, A.; Herzog, H.; et al. Peptide YY Regulates Bone Remodeling in Mice: A Link between Gut and Skeletal Biology. PLoS ONE 2012, 7, e40038. [Google Scholar] [CrossRef]
- Kim, T.Y.; Shoback, D.M.; Black, D.M.; Rogers, S.J.; Stewart, L.; Carter, J.T.; Posselt, A.M.; King, N.J.; Schafer, A.L. Increases in PYY and uncoupling of bone turnover are associated with loss of bone mass after gastric bypass surgery. Bone 2019, 131, 115115. [Google Scholar] [CrossRef]
- Misra, M.; Miller, K.K.; Tsai, P.; Gallagher, K.; Lin, A.; Lee, N.; Herzog, D.B.; Klibanski, A. Elevated Peptide YY Levels in Adolescent Girls with Anorexia Nervosa. J. Clin. Endocrinol. Metab. 2006, 91, 1027–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, E.W.; Wewalka, M.; Ding, S.-A.; Simonson, D.C.; Foster, K.; Holst, J.J.; Vernon, A.; Goldfine, A.B.; Halperin, F. Effects of Gastric Bypass and Gastric Banding on Bone Remodeling in Obese Patients with Type 2 Diabetes. J. Clin. Endocrinol. Metab. 2016, 101, 714–722. [Google Scholar] [CrossRef] [PubMed]
- Remmel, L.; Tillmann, V.; Purge, P.; Saar, M.; Maasalu, K. Associations between Bone Mineral Characteristics and Serum Levels of Ghrelin and Peptide YY in Overweight Adolescent Boys. Horm. Res. Paediatr. 2015, 84, 6–13. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Overall | |
|---|---|
| n | 14 |
| Age (years) | 53.8 (45.8, 64.5) |
| Males n (%) | 3 (21.4) |
| BMI (kg/m2) | 23.1 (21.8, 23.9) |
| Weight (kg) | 63.9 (59.9, 69.7) |
| HbA1c (mmol/mol) | 34.5 (32.5, 36.0) |
| Standardized Mixed Meal | Granola Bar | |
|---|---|---|
| n | 14 | 14 |
| Fasting glucose (mmol/L) | 5.2 (0.4) | 5.2 (0.4) |
| Fasting insulin (pmol/L) | 27.0 (20.5, 33.0) | 29.0 (23.2, 34.8) |
| Fasting P1NP (ng/mL) | 60.8 (44.0, 78.6) | 50.4 (43.4, 62.5) |
| Fasting CTX (ng/mL) | 0.4 (0.3, 0.5) | 0.4 (0.3, 0.6) |
| Fasting GLP-1 (pmol/L) | 6.4 (5.6, 8.1) | 7.5 (5.6, 8.5) |
| Fasting GIP (pg/mL) | 34.0 (26.9, 63.4) | 54.3 (44.1, 75.8) |
| Fasting PYY (pg/mL) | 25.7 (20.7, 33.6) | 29.8 (24.5, 39.2) |
| P1NPiAUC (ng/mL × min) | −947.4 (−1911.1, −221.7) | −270.5 (−561.1, −176.1) |
| CTXiAUC (ng/mL × min) | −34.9 (−44.9, −21.2) | −30.7 (−37.0, −17.6) |
| GLP-1iAUC (ng/mL × min) | 7.0 (4.7, 9.6) | 5.0 (2.8, 6.9) |
| GIPiAUC (ng/mL × min) * | 41.2 (34.2, 56.9) | 25.7 (18.8, 31.1) |
| PYYiAUC (ng/mL × min) * | 7.0 (3.7, 10.4) | 2.4 (1.8, 6.0) |
| CTXiAUC | P1NPiAUC | |||
|---|---|---|---|---|
| Estimate (95% CI) | p | Estimate (95% CI) | p | |
| GLP-1iAUC | 6.97 (3.39; 10.54) | <0.001 | 17.59 (−298.93; 334.12) | 0.913 |
| GIPiAUC | 0.51 (0.24; 0.77) | <0.001 | 0.69 (−23.48; 24.87) | 0.955 |
| PYYiAUC | −8.34 (−11.20; −5.59) | <0.001 | −59.82 (−311.40; 191.76) | 0.641 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jensen, N.W.; Clemmensen, K.K.B.; Jensen, M.M.; Pedersen, H.; Færch, K.; Diaz, L.J.; Quist, J.S.; Størling, J. Associations between Postprandial Gut Hormones and Markers of Bone Remodeling. Nutrients 2021, 13, 3197. https://doi.org/10.3390/nu13093197
Jensen NW, Clemmensen KKB, Jensen MM, Pedersen H, Færch K, Diaz LJ, Quist JS, Størling J. Associations between Postprandial Gut Hormones and Markers of Bone Remodeling. Nutrients. 2021; 13(9):3197. https://doi.org/10.3390/nu13093197
Chicago/Turabian StyleJensen, Nina Wittorff, Kim Katrine Bjerring Clemmensen, Marie Møller Jensen, Hanne Pedersen, Kristine Færch, Lars Jorge Diaz, Jonas Salling Quist, and Joachim Størling. 2021. "Associations between Postprandial Gut Hormones and Markers of Bone Remodeling" Nutrients 13, no. 9: 3197. https://doi.org/10.3390/nu13093197