Macrophages from Mice Administered Rhus verniciflua Stokes Extract Show Selective Anti-Inflammatory Activity

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Qualitative and Quantitative Analyses of RVS Extract
2.3. Animals
2.4. In Vivo Experiment for Peritoneal Exudate Cell Preparation
2.5. Peritoneal Macrophage Culture
2.6. RAW264.7 Cell Culture
2.7. Flow Cytometry
2.8. In Vivo Experiment for Serum Cytokine Response
2.9. ELISA
2.10. Statistical Analysis
3. Results
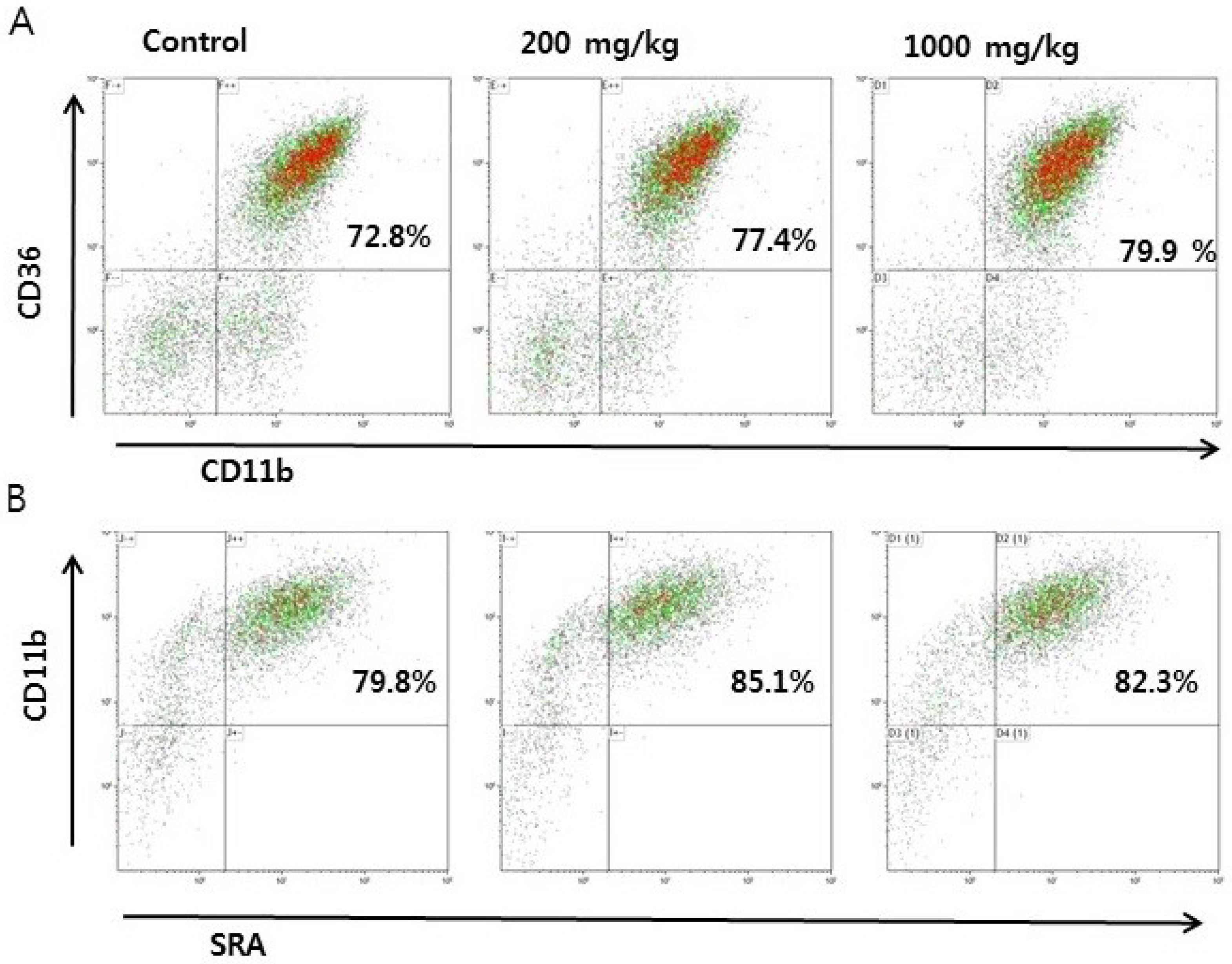
3.1. Effects of RVS Extract on Thioglycollate-Induced Monocyte Differentiation in Peritoneal Exudate Cells
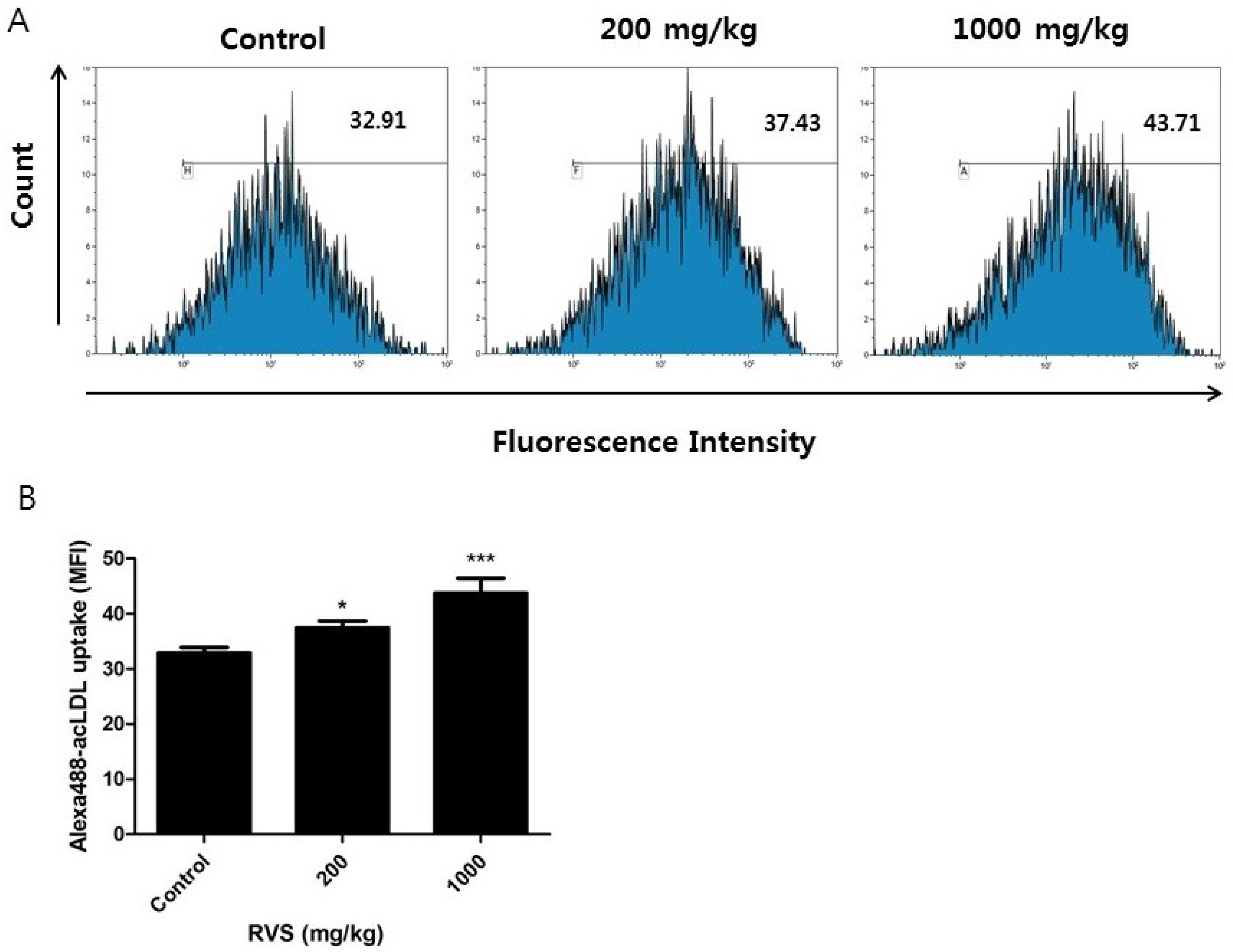
3.2. Effect of RVS Extract on Scavenger Receptor Activity in Peritoneal Macrophages
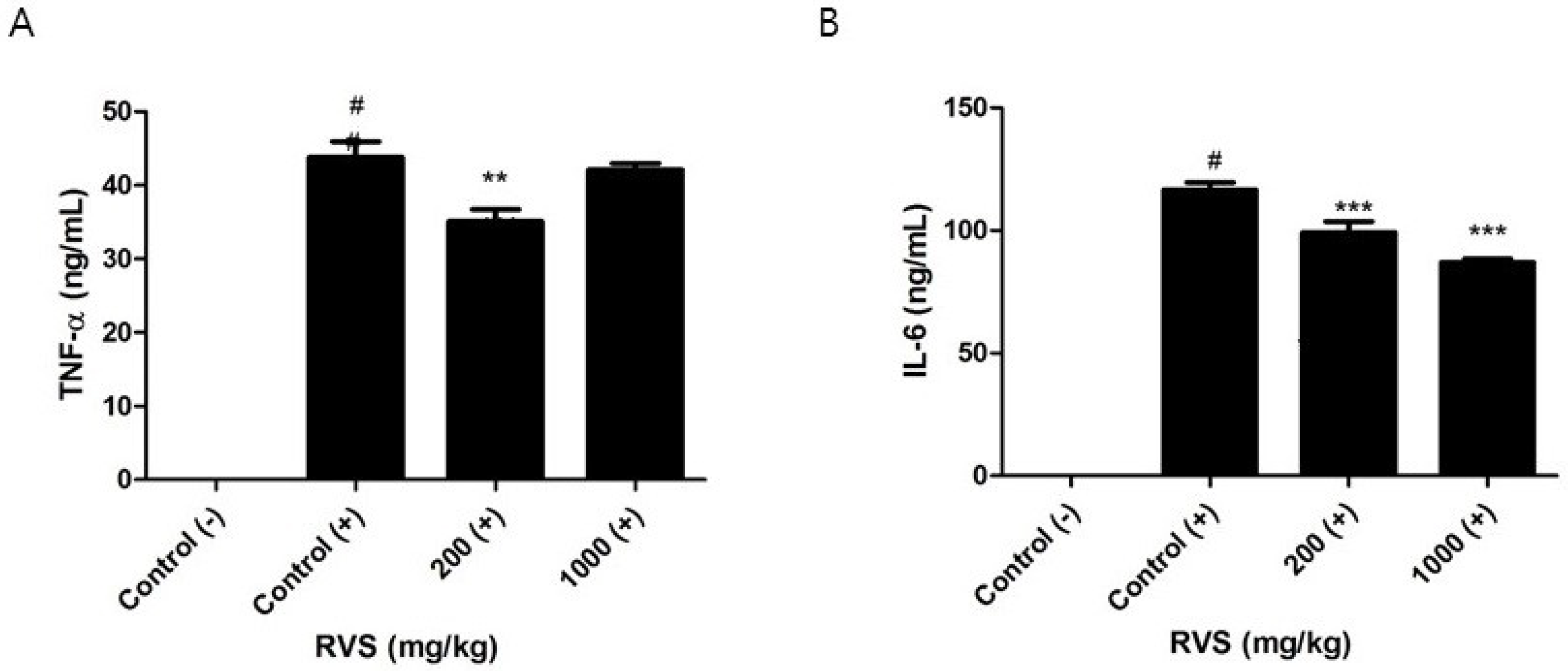
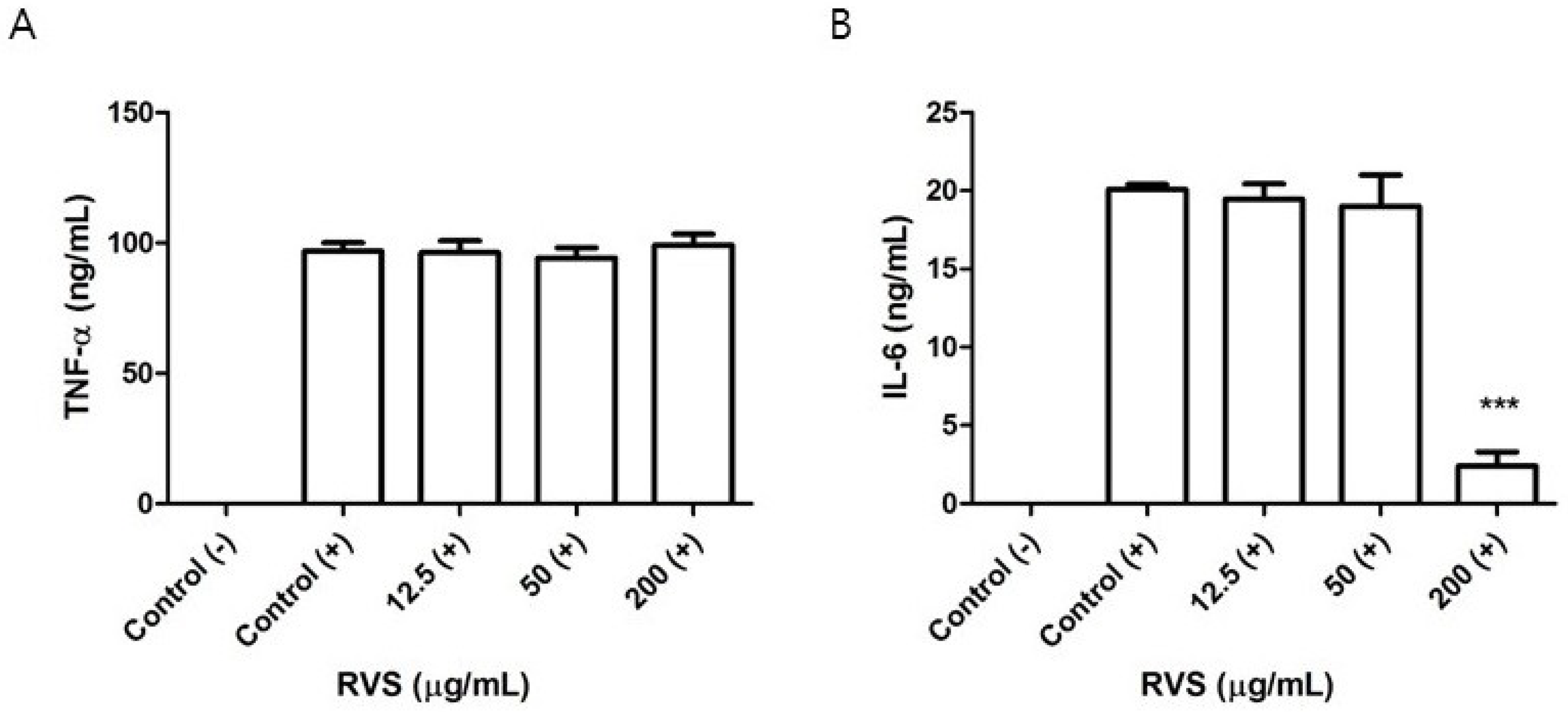
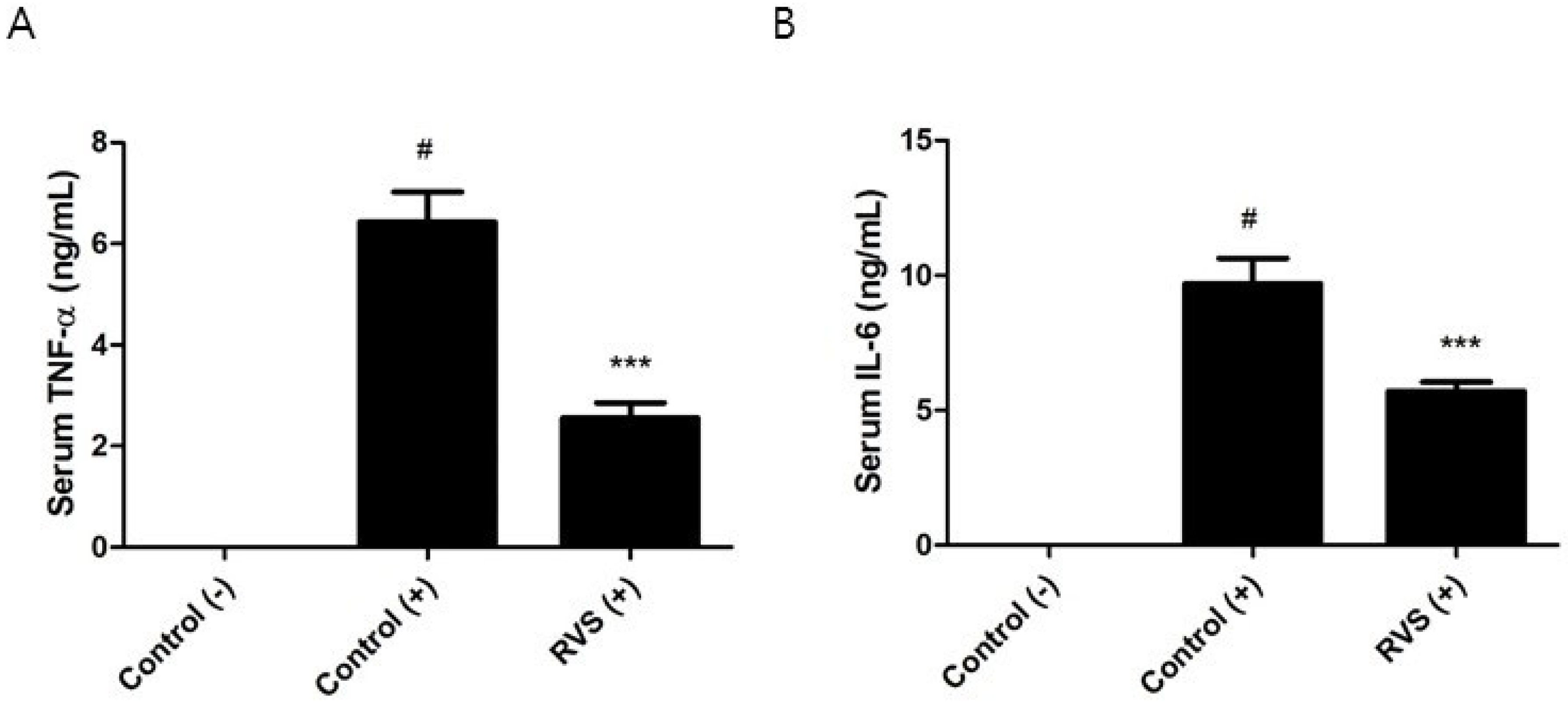
3.3. Effects of RVS Extract on TNF-α and IL-6 Production in Peritoneal Macrophages Stimulated with LPS
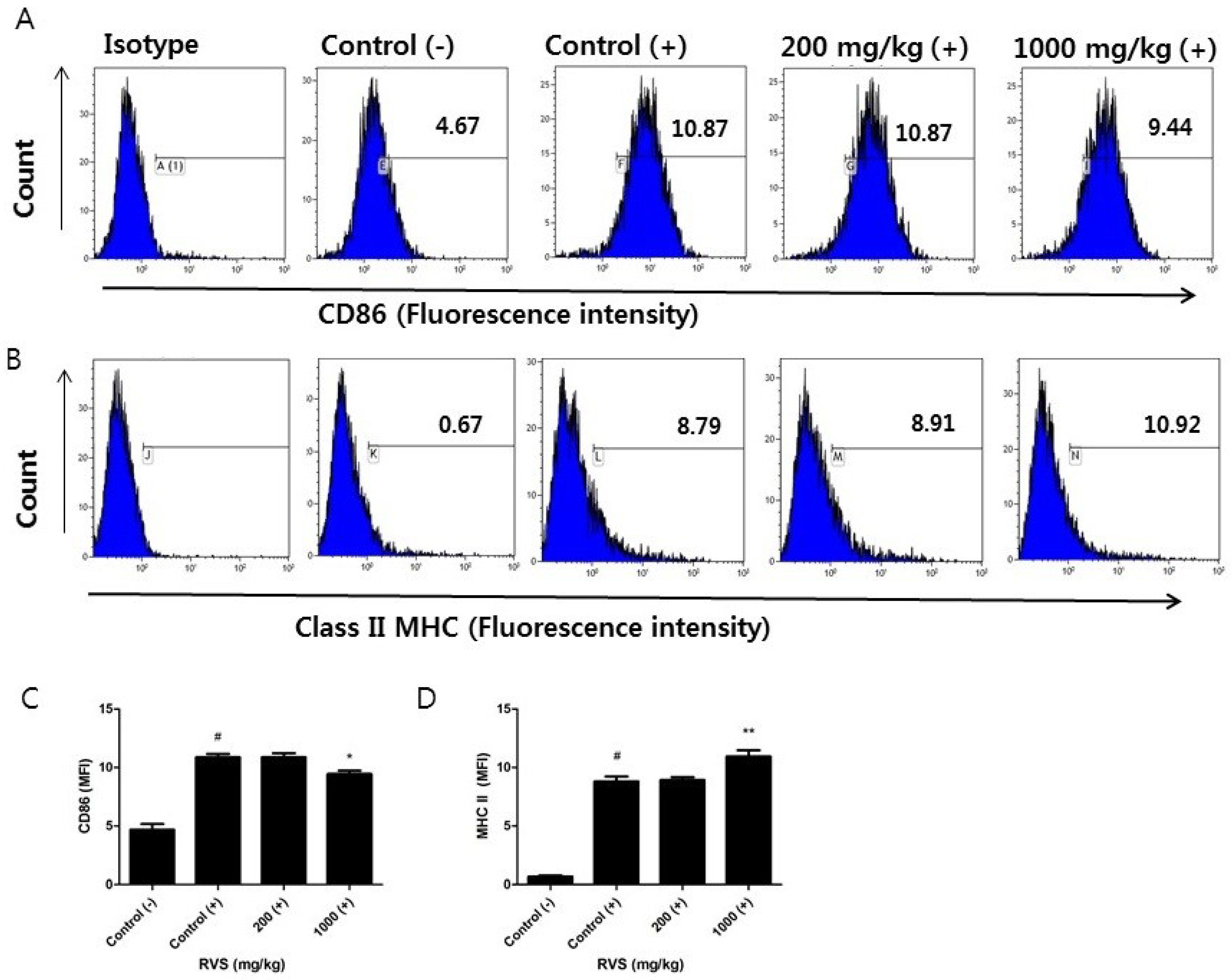
3.4. Effects of RVS Extract on CD86 and Class II MHC Surface Expression in Peritoneal Macrophages Stimulated with LPS
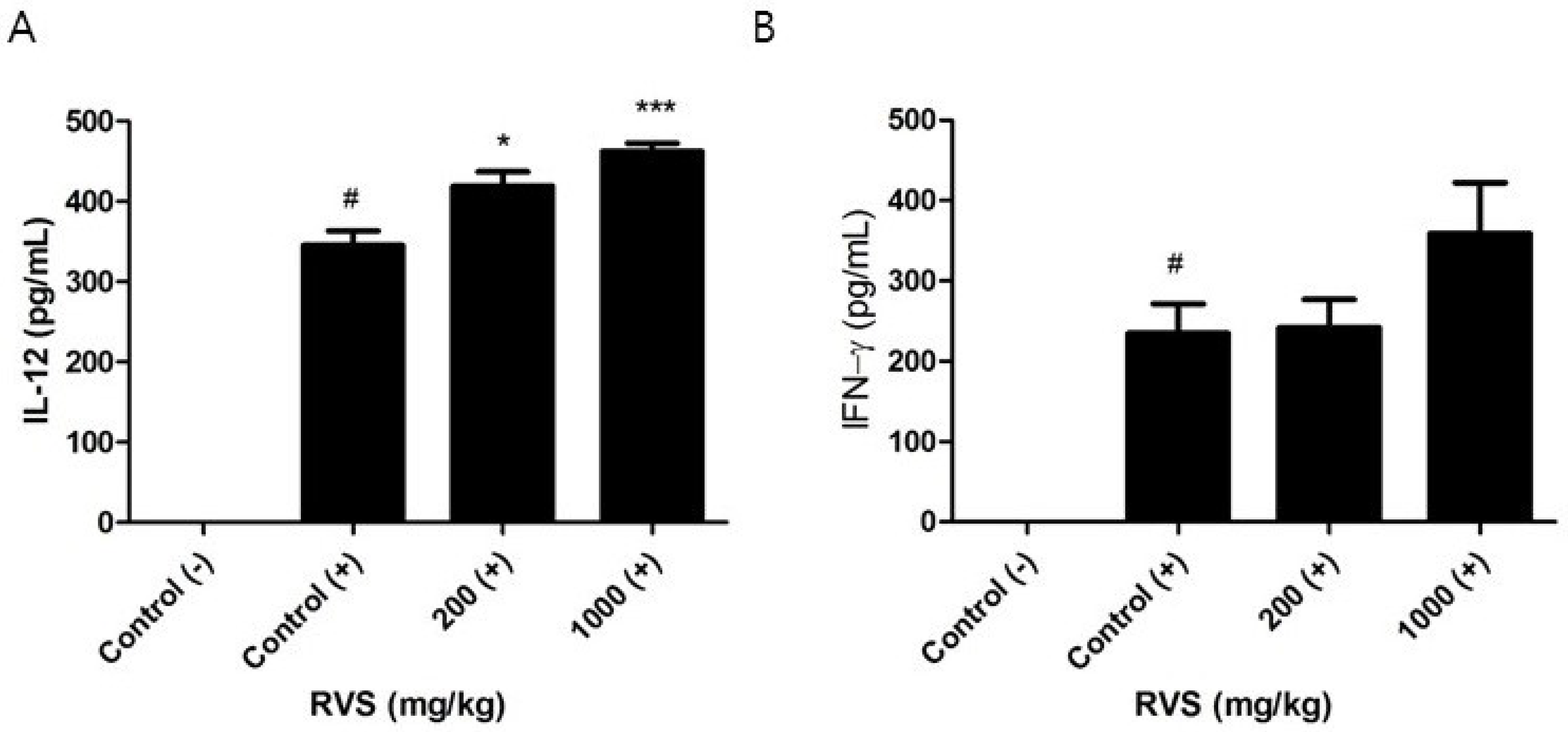
3.5. Effects of RVS Extract on IL-12 and IFN-γ Production in Peritoneal Macrophages Stimulated with LPS
3.6. Effects of RVS Extract on the Systemic Response to LPS
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ginhoux, F.; Jung, S. Monocytes and macrophages: Developmental pathways and tissue homeostasis. Nat. Rev. Immunol. 2014, 14, 392–404. [Google Scholar] [CrossRef] [PubMed]
- Sprangers, S.; de Vries, T.J.; Everts, V. Monocyte heterogeneity: Consequences for monocyte-derived immune cells. J. Immunol. Res. 2016, 2016, 1475435. [Google Scholar] [CrossRef] [PubMed]
- Jakubzick, C.V.; Randolph, G.J.; Henson, P.M. Monocyte differentiation and antigen-presenting functions. Nat. Rev. Immunol. 2017, 17, 349–362. [Google Scholar] [CrossRef] [PubMed]
- Moretti, J.; Blander, J.M. Insights into phagocytosis-coupled activation of pattern recognition receptors and inflammasomes. Curr. Opin. Immunol. 2014, 26, 100–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogdan, C. Nitric oxide synthase in innate and adaptive immunity: An update. Trends Immunol. 2015, 36, 161–178. [Google Scholar] [CrossRef]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef]
- Zundler, S.; Neurath, M.F. Interleukin-12: Functional activities and implications for disease. Cytokine Growth Factor Rev. 2015, 26, 559–568. [Google Scholar] [CrossRef]
- Chaplin, D.D. Overview of the immune response. J. Allergy Clin. Immunol. 2010, 125, S3–S23. [Google Scholar] [CrossRef]
- Cook, A.D.; Braine, E.L.; Hamilton, J.A. The phenotype of inflammatory macrophages is stimulus dependent: Implications for the nature of the inflammatory response. J. Immunol. 2003, 171, 4816–4823. [Google Scholar] [CrossRef]
- Lam, D.; Harris, D.; Qin, Z. Inflammatory mediator profiling reveals immune properties of chemotactic gradients and macrophage mediator production inhibition during thioglycollate elicited peritoneal inflammation. Mediators Inflamm. 2013, 2013, 931562. [Google Scholar] [CrossRef]
- Ghosn, E.E.; Cassado, A.A.; Govoni, G.R.; Fukuhara, T.; Yang, Y.; Monack, D.M.; Bortoluci, K.R.; Almeida, S.R.; Herzenberg, L.A.; Herzenberg, L.A. Two physically, functionally, and developmentally distinct peritoneal macrophage subsets. Proc. Natl. Acad. Sci. USA 2010, 107, 2568–2573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, Y.; Kodama, T.; Hansson, G.K. Differential expression of scavenger receptor isoforms during monocyte-macrophage differentiation and foam cell formation. Arterioscler. Thromb. 1994, 14, 798–806. [Google Scholar] [CrossRef] [PubMed]
- Huh, H.Y.; Pearce, S.F.; Yesner, L.M.; Schindler, J.L.; Silverstein, R.L. Regulated expression of CD36 during monocyte-to-macrophage differentiation: Potential role of CD36 in foam cell formation. Blood 1996, 87, 2020–2028. [Google Scholar] [PubMed]
- Prudovsky, I.; Popov, K.; Akimov, S.; Serov, S.; Zelenin, A.; Meinhardt, G.; Baier, P.; Sohn, C.; Hass, R. Antisense CD11b integrin inhibits the development of a differentiated monocyte/macrophage phenotype in human leukemia cells. Eur. J. Cell Biol. 2002, 81, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Canton, J.; Neculai, D.; Grinstein, S. Scavenger receptors in homeostasis and immunity. Nat. Rev. Immunol. 2013, 13, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Fuhrman, B.; Shiner, M.; Volkova, N.; Aviram, M. Cell-induced copper ion-mediated low density lipoprotein oxidation increases during in vivo monocyte-to-macrophage differentiation. Free Radic. Biol. Med. 2004, 37, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Lee, H.J.; Lee, H.J.; Choi, W.C.; Yoon, S.W.; Ko, S.G.; Ahn, K.S.; Choi, S.H.; Ahn, K.S.; Lieske, J.C.; et al. Rhus verniciflua stokes prevents cisplatin-induced cytotoxicity and reactive oxygen species production in MDCK-I renal cells and intact mice. Phytomedicine 2009, 16, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.H.; Kim, J.H.; Hong, M.H.; Seog, H.M.; Oh, S.H.; Lee, P.J.; Kim, G.J.; Kim, H.M.; Um, J.Y.; Ko, S.G. Phenolic-rich fraction from rhus verniciflua stokes (RVS) suppress inflammatory response via NF-kappaB and JNK pathway in lipopolysaccharide-induced RAW 264.7 macrophages. J. Ethnopharmacol. 2007, 110, 490–497. [Google Scholar] [CrossRef]
- Jung, C.H.; Kim, J.H.; Kim, J.H.; Chung, J.H.; Choi, H.S.; Seo, J.B.; Shin, Y.C.; Kim, S.H.; Ko, S.G. Anti-inflammatory effect of Rhus verniviflua stokes by suppression of iNOS-mediated Akt and ERK pathways: In-vitro and in-vivo studies. J. Pharm. Pharmacol. 2011, 63, 679–687. [Google Scholar] [CrossRef]
- Im, W.K.; Park, H.J.; Lee, K.S.; Lee, J.H.; Kim, Y.D.; Kim, K.H.; Park, S.J.; Hong, S.; Jeon, S.H. Fisetin-rich extracts of Rhus verniciflua stokes improve blood flow rates in mice fed both normal and high-fat diets. J. Med. Food 2016, 19, 120–126. [Google Scholar] [CrossRef]
- Sapkota, K.; Kim, S.; Park, S.E.; Kim, S.J. Detoxified extract of Rhus verniciflua stokes inhibits rotenone-induced apoptosis in human dopaminergic cells, sh-sy5y. Cell. Mol. Neurobiol. 2011, 31, 213–223. [Google Scholar] [CrossRef]
- Son, Y.O.; Lee, K.Y.; Lee, J.C.; Jang, H.S.; Kim, J.G.; Jeon, Y.M.; Jang, Y.S. Selective antiproliferative and apoptotic effects of flavonoids purified from Rhus verniciflua stokes on normal versus transformed hepatic cell lines. Toxicol. Lett. 2005, 155, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.; Jung, H.; Kim, K.; Lee, S.; Yoon, S.; Park, J.; Kim, S.; Cheon, S.; Eo, W.; Lee, S. Rhus verniciflua stokes against advanced cancer: A perspective from the korean integrative cancer center. J. Biomed. Biotechnol. 2012, 2012, 874276. [Google Scholar] [CrossRef]
- Park, D.K.; Lee, Y.G.; Park, H.J. Extract of Rhus verniciflua bark suppresses 2,4-dinitrofluorobenzene-induced allergic contact dermatitis. Evid. Based Complement. Alternat. Med. 2013, 2013, 879696. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Seo, G.S.; Sohn, D.H. Inhibition of lipopolysaccharide-induced expression of inducible nitric oxide synthase by butein in RAW 264.7 cells. Biochem. Biophys. Res. Commun. 2004, 323, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.S.; Jeong, G.S.; Li, B.; Park, H.; Kim, Y.C. Anti-inflammatory effects of sulfuretin from Rhus verniciflua stokes via the induction of heme oxygenase-1 expression in murine macrophages. Int. Immunopharmacol. 2010, 10, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.C.; Kang, S.H.; Jeong, S.J.; Kim, S.H.; Ko, H.S.; Kim, S.H. Inhibition of c-jun N-terminal kinase and nuclear factor kappa B pathways mediates fisetin-exerted anti-inflammatory activity in lipopolysccharide-treated RAW264.7 cells. Immunopharmacol. Immunotoxicol. 2012, 34, 645–650. [Google Scholar] [CrossRef]
- Jin, M.J.; Kim, I.S.; Park, J.S.; Dong, M.S.; Na, C.S.; Yoo, H.H. Pharmacokinetic profile of eight phenolic compounds and their conjugated metabolites after oral administration of Rhus verniciflua extracts in rats. J. Agric. Food Chem. 2015, 63, 5410–5416. [Google Scholar] [CrossRef]
- Puddu, P.; Fantuzzi, L.; Borghi, P.; Varano, B.; Rainaldi, G.; Guillemard, E.; Malorni, W.; Nicaise, P.; Wolf, S.F.; Belardelli, F.; et al. IL-12 induces ifn-gamma expression and secretion in mouse peritoneal macrophages. J. Immunol. 1997, 159, 3490–3497. [Google Scholar]
- Purswani, M.U.; Eckert, S.J.; Arora, H.K.; Noel, G.J. Effect of ciprofloxacin on lethal and sublethal challenge with endotoxin and on early cytokine responses in a murine in vivo model. J. Antimicrob. Chemother. 2002, 50, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.W.; Yang, G.E.; Kim, Y.B.; Eom, S.H.; Lew, J.H.; Kang, H. Anti-inflammatory activity of cinnamon water extract in vivo and in vitro LPS-induced models. BMC Complement. Altern. Med. 2012, 12, 237. [Google Scholar] [CrossRef] [PubMed]
- Michie, H.R.; Manogue, K.R.; Spriggs, D.R.; Revhaug, A.; O’Dwyer, S.; Dinarello, C.A.; Cerami, A.; Wolff, S.M.; Wilmore, D.W. Detection of circulating tumor necrosis factor after endotoxin administration. N. Engl. J. Med. 1988, 318, 1481–1486. [Google Scholar] [CrossRef]
- Kumins, N.H.; Hunt, J.; Gamelli, R.L.; Filkins, J.P. Partial hepatectomy reduces the endotoxin-induced peak circulating level of tumor necrosis factor in rats. Shock 1996, 5, 385–388. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Kirman, I.; Tschepen, I.; Schwab, R.; Weksler, M.E. Peritoneal lavage reduces lipopolysaccharide-induced elevation of serum tnf-alpha and il-6 mortality in mice. Inflammation 1997, 21, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.; Scott, M.J.; Loughran, P.; Gibson, G.; Sodhi, C.; Watkins, S.; Hackam, D.; Billiar, T.R. Lipopolysaccharide clearance, bacterial clearance, and systemic inflammatory responses are regulated by cell type-specific functions of TLR4 during sepsis. J. Immunol. 2013, 190, 5152–5160. [Google Scholar] [CrossRef]
- Moon, J.E.; Shin, J.H.; Kwon, O.; Kim, J.Y. A standardized extract of rhus verniciflua stokes protects wistar rats against lipopolysaccharide-induced acute inflammation. J. Med. Food 2015, 18, 1223–1230. [Google Scholar] [CrossRef] [PubMed]
- Croft, M.; Dubey, C. Accessory molecule and costimulation requirements for cd4 t cell response. Crit. Rev. Immunol. 1997, 17, 89–118. [Google Scholar] [CrossRef]
- Figueiredo, F.; Koerner, T.J.; Adams, D.O. Molecular mechanisms regulating the expression of class ii histocompatibility molecules on macrophages. Effects of inductive and suppressive signals on gene transcription. J. Immunol. 1989, 143, 3781–3786. [Google Scholar]
- Tugues, S.; Burkhard, S.H.; Ohs, I.; Vrohlings, M.; Nussbaum, K.; Vom Berg, J.; Kulig, P.; Becher, B. New insights into il-12-mediated tumor suppression. Cell Death Differ. 2015, 22, 237–246. [Google Scholar] [CrossRef]
- Sieburth, D.; Jabs, E.W.; Warrington, J.A.; Li, X.; Lasota, J.; LaForgia, S.; Kelleher, K.; Huebner, K.; Wasmuth, J.J.; Wolf, S.F. Assignment of genes encoding a unique cytokine (IL12) composed of two unrelated subunits to chromosomes 3 and 5. Genomics 1992, 14, 59–62. [Google Scholar] [CrossRef]
- Liu, J.; Cao, S.; Herman, L.M.; Ma, X. Differential regulation of interleukin (IL)-12 p35 and p40 gene expression and interferon (IFN)-gamma-primed il-12 production by IFN regulatory factor 1. J. Exp. Med. 2003, 198, 1265–1276. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, C.; Schleicher, U. Production of interferon-gamma by myeloid cells—Fact or fancy? Trends Immunol. 2006, 27, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Adachi, O.; Ogawa, T.; Takeda, K.; Akira, S. Unresponsiveness of MyD88-deficient mice to endotoxin. Immunity 1999, 11, 115–122. [Google Scholar] [CrossRef]
- Hoebe, K.; Janssen, E.M.; Kim, S.O.; Alexopoulou, L.; Flavell, R.A.; Han, J.; Beutler, B. Upregulation of costimulatory molecules induced by lipopolysaccharide and double-stranded RNA occurs by Trif-dependent and Trif-independent pathways. Nat. Immunol. 2003, 4, 1223–1229. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | MS [M + H]+ | Content (%) |
|---|---|---|
| Fisetin | 287.1 | 0.56 |
| Sulfuretin | 271.1 | 0.01 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, B.-G.; Song, Y.; Lee, M.-G.; Ku, J.-M.; Jin, S.-J.; Hong, J.-W.; Lee, S.; Kang, H. Macrophages from Mice Administered Rhus verniciflua Stokes Extract Show Selective Anti-Inflammatory Activity. Nutrients 2018, 10, 1926. https://doi.org/10.3390/nu10121926
Kim B-G, Song Y, Lee M-G, Ku J-M, Jin S-J, Hong J-W, Lee S, Kang H. Macrophages from Mice Administered Rhus verniciflua Stokes Extract Show Selective Anti-Inflammatory Activity. Nutrients. 2018; 10(12):1926. https://doi.org/10.3390/nu10121926
Chicago/Turabian StyleKim, Bo-Geun, Youngju Song, Mi-Gi Lee, Jin-Mo Ku, So-Jung Jin, Joung-Woo Hong, SeungGwan Lee, and Hee Kang. 2018. "Macrophages from Mice Administered Rhus verniciflua Stokes Extract Show Selective Anti-Inflammatory Activity" Nutrients 10, no. 12: 1926. https://doi.org/10.3390/nu10121926