
Effect of Yakae-Prajamduen-Jamod Traditional Thai Remedy on Cognitive Impairment in an Ovariectomized Mouse Model and Its Mechanism of Action

 , , , , , ,
, , , , , ,  , and
, and
Abstract
:1. Introduction
2. Results
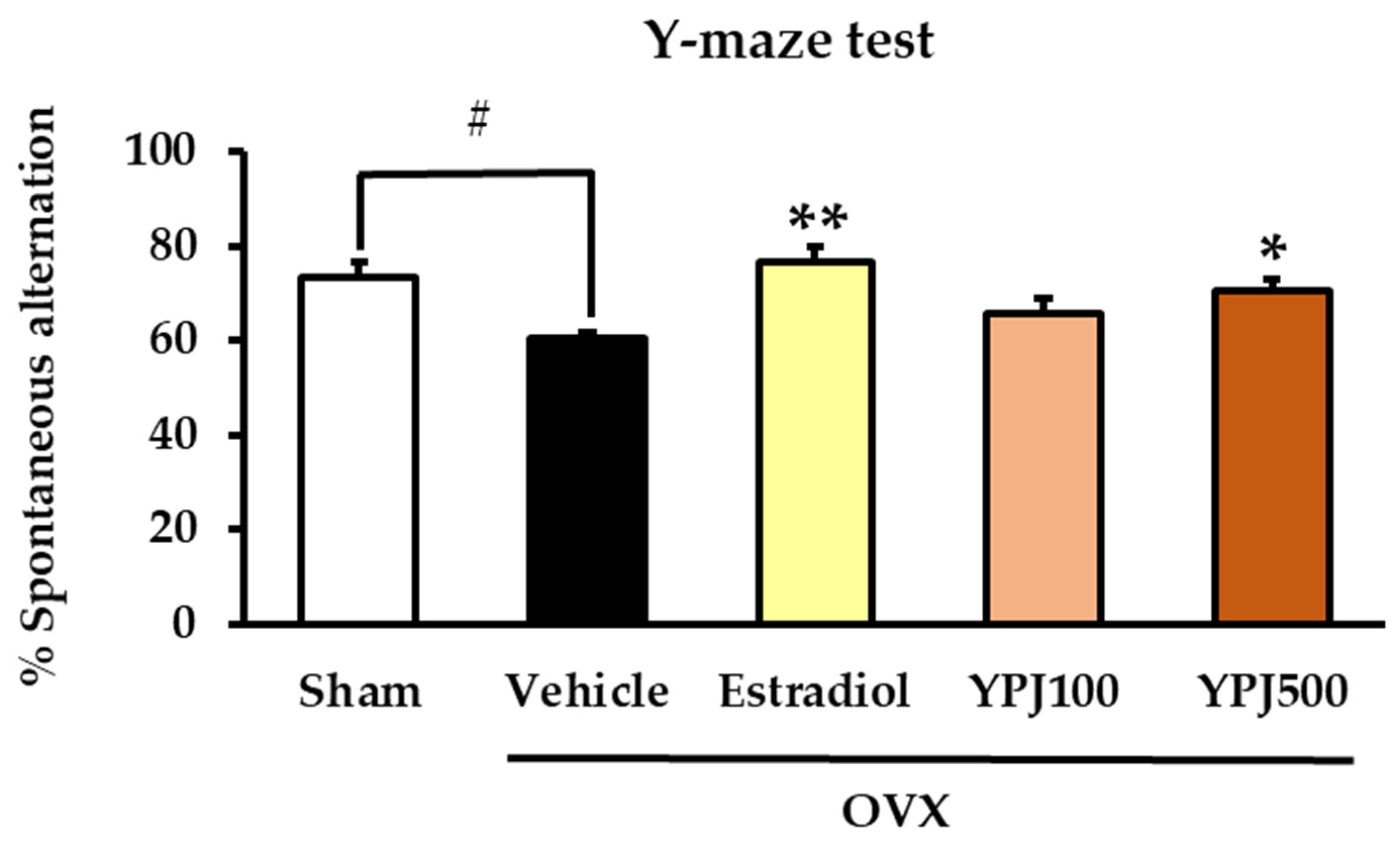
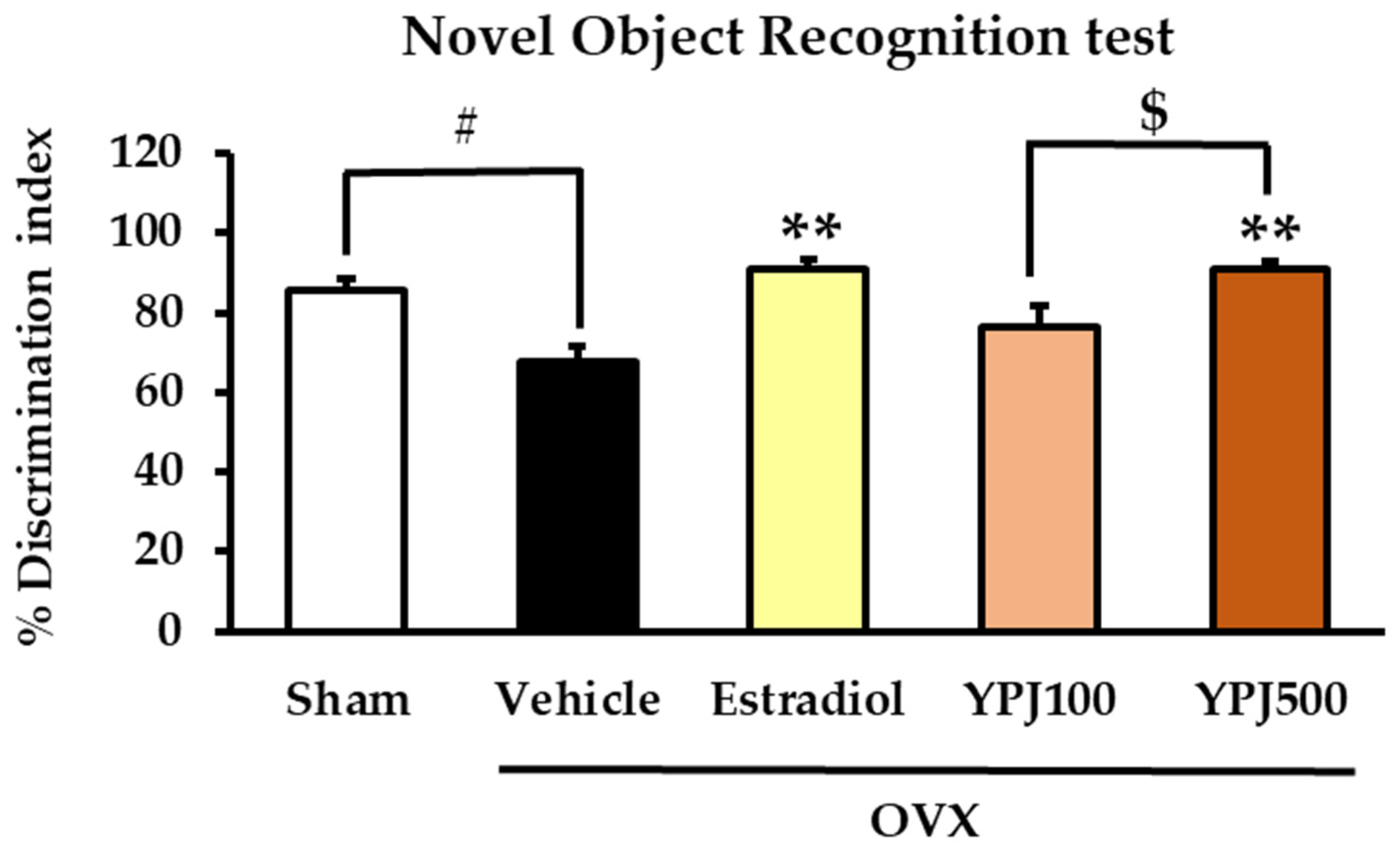
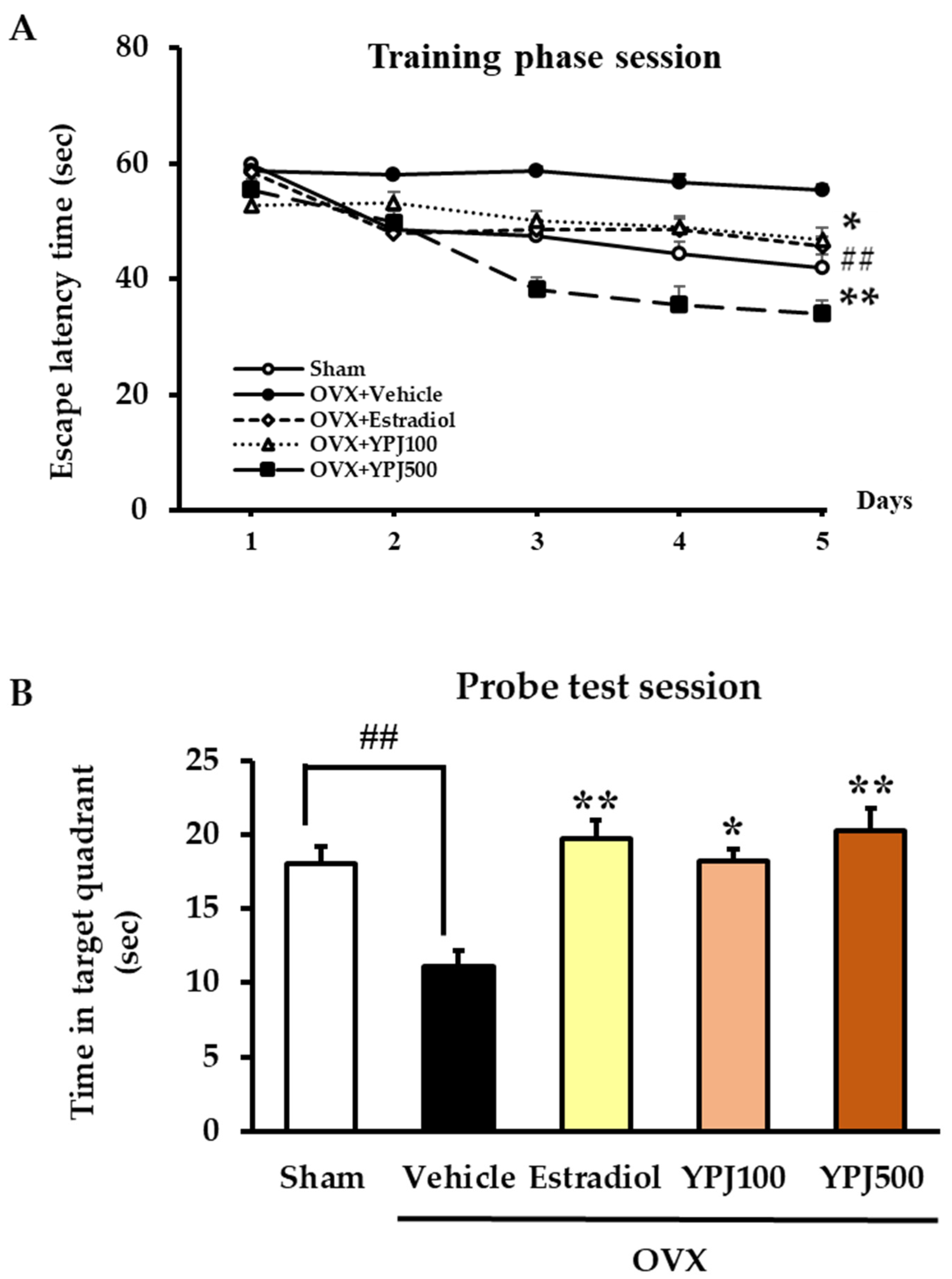
2.1. Effect of YPJ Remedy on OVX-Induced Cognitive Deficit-like Behavior
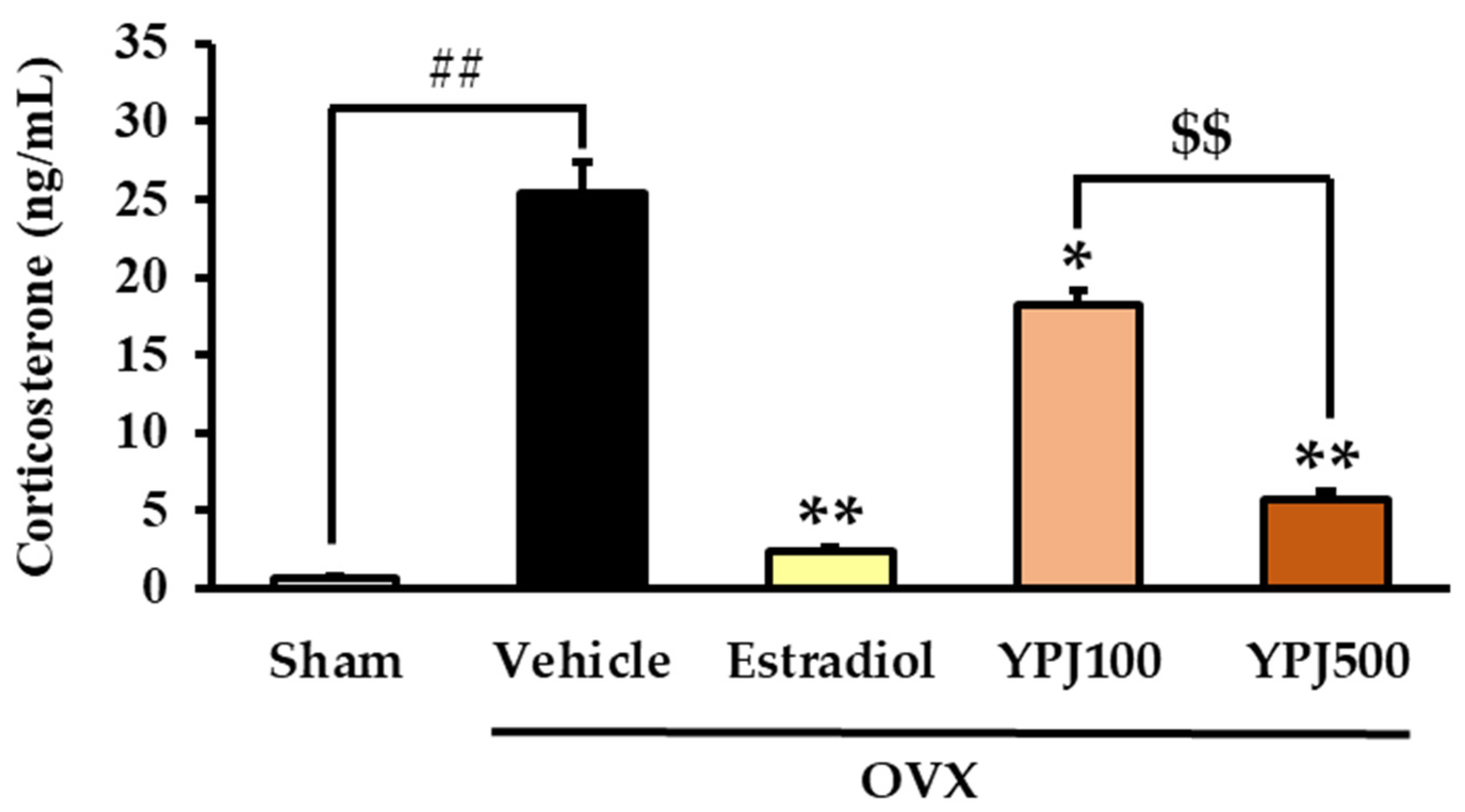
2.2. Effect of YPJ Remedy on Serum CORT Level
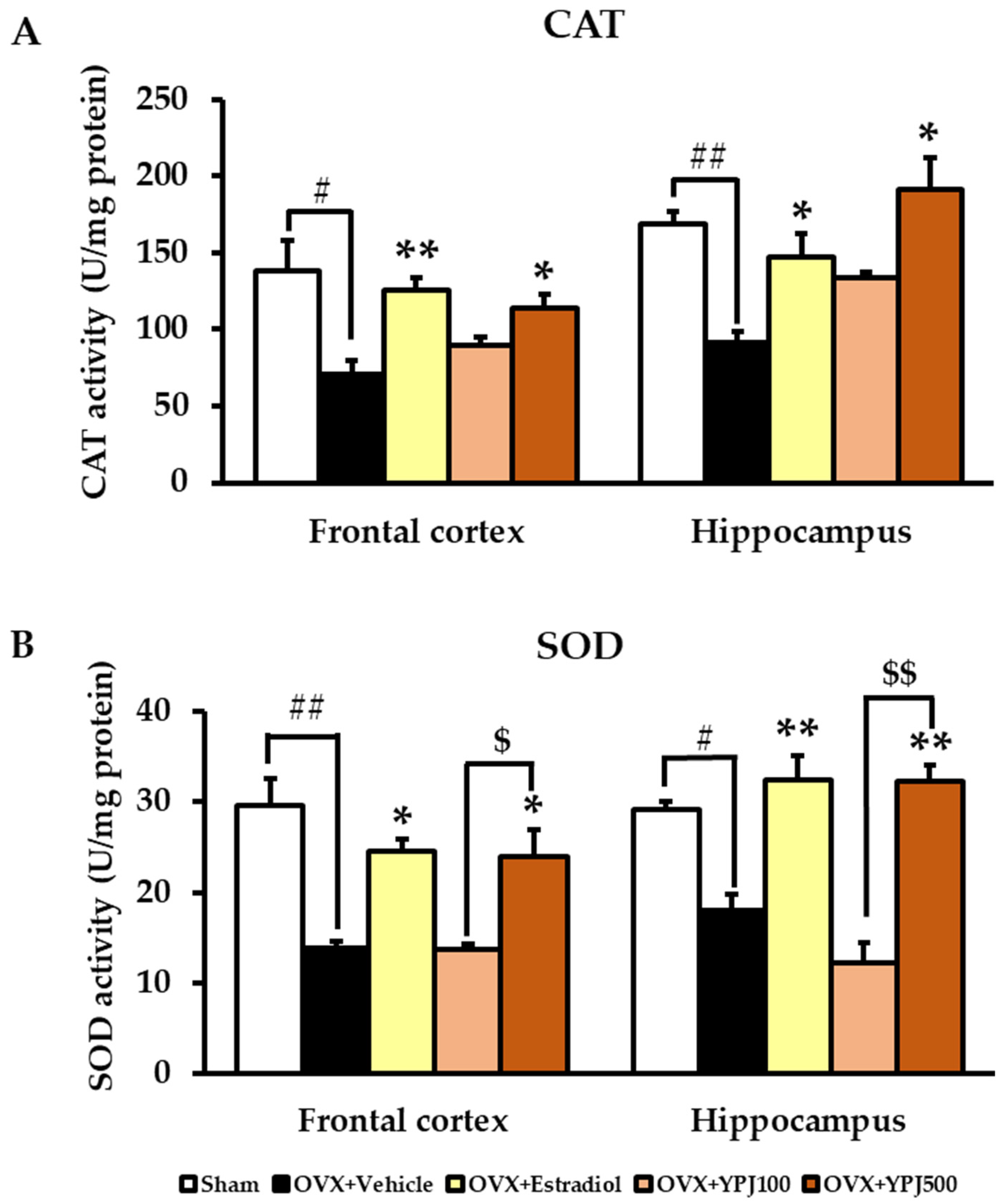
2.3. Effect of YPJ Remedy on Antioxidant Enzyme Activities in the Frontal Cortex and Hippocampus
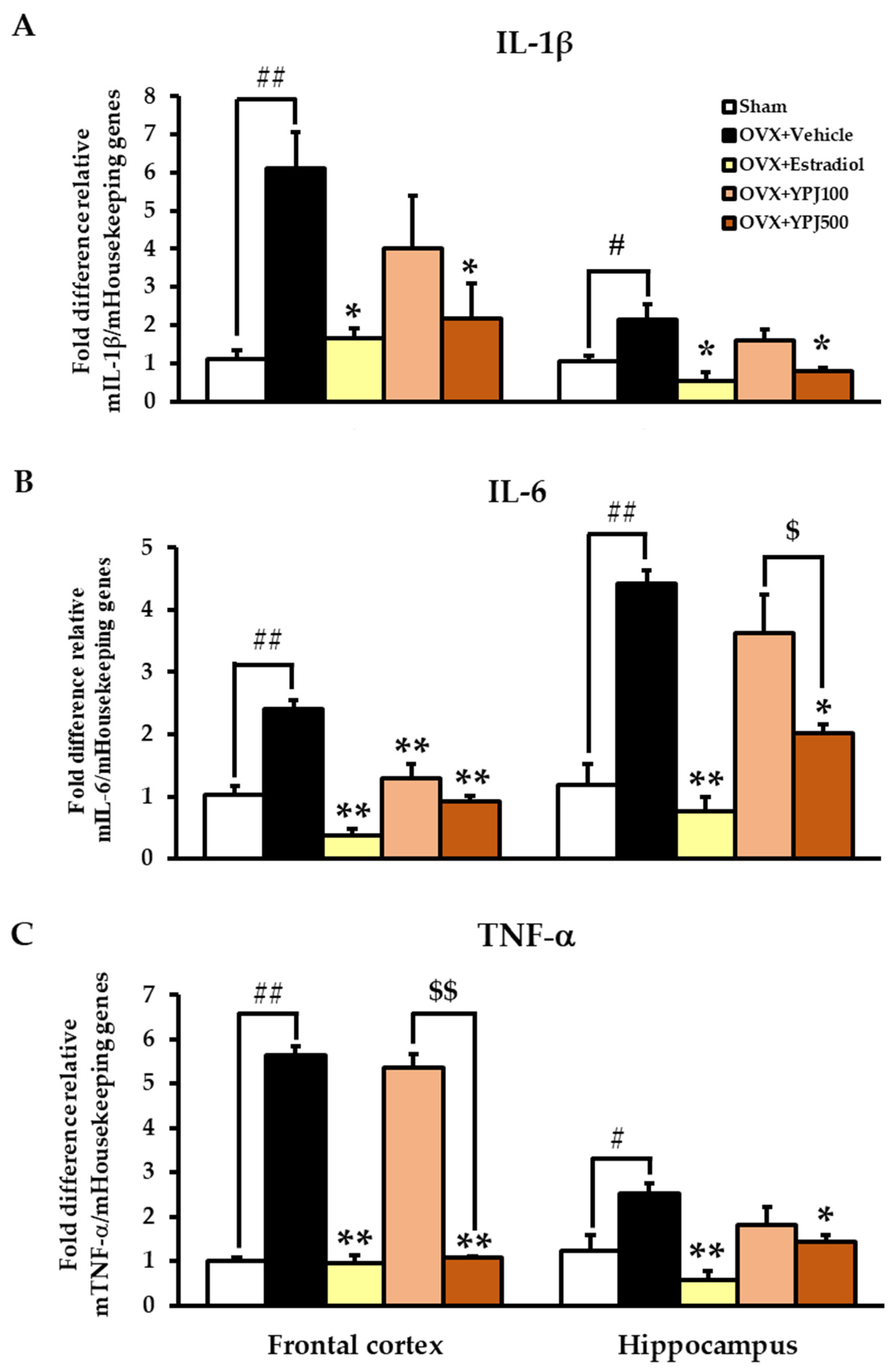
2.4. Effect of YPJ Remedy on OVX-Induced Changes in Inflammatory Cytokine mRNA Expression in the Frontal Cortex and Hippocampus
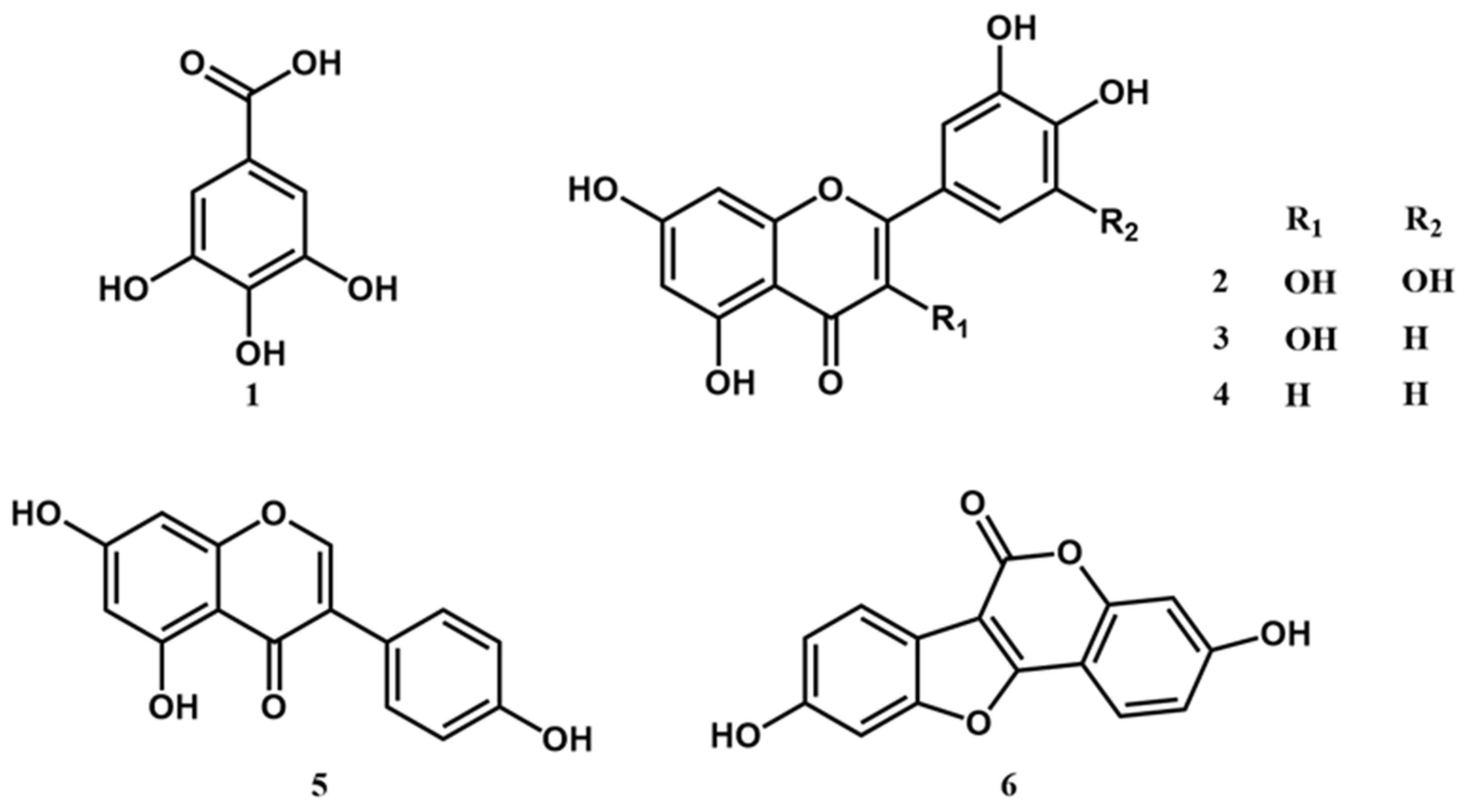
2.5. High Performance Liquid Chromatography (HPLC) Analysis of Constituents of YPJ Extract and Validation of the Method
3. Discussion
4. Materials and Methods
4.1. Plant Materials and YPJ Preparations
4.2. Animals
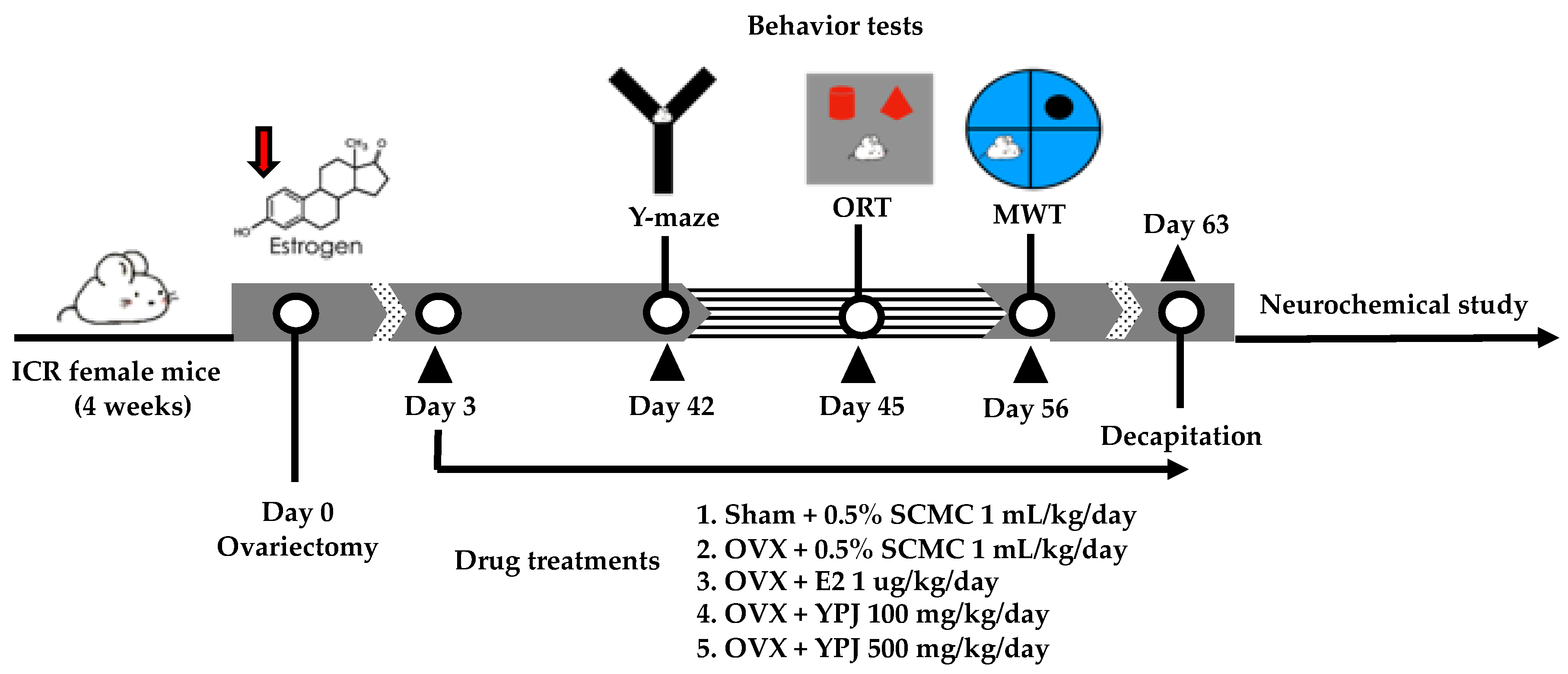
4.3. Surgical Procedure and Drug Administration
4.4. Behavioral Studies
4.4.1. Y-Maze Test
4.4.2. Novel Object Recognition Test (NORT)
4.4.3. Morris Water Maze Test (MWMT)
4.5. Determination of Serum Corticosterone Level
4.6. Determination of Antioxidant Enzyme Activities
4.7. Quantitative Real-Time Polymerase Chain Reaction (Q-PCR)
4.8. HPLC Analysis and Validation
4.8.1. Standards and Reagents
4.8.2. Hydrolysis of the Yakae-Prajamduen-Jamod Extract
4.8.3. HPLC Analysis and Validation
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Nelson, L.M. Clinical practice. Primary ovarian insufficiency. N. Engl. J. Med. 2009, 360, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Irwin, R.W.; Zhao, L.; Nilsen, J.; Hamilton, R.T.; Brinton, R.D. Mitochondrial bioenergetic deficit precedes Alzheimer’s pathology in female mouse model of Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2009, 106, 14670–14675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, Y.; Jia, J.P.; Ji, X.J.; Tian, T. Estrogen associated gene polymorphisms and their interactions in the progress of Alzheimer’s disease. Prog. Neurobiol. 2013, 111, 53–74. [Google Scholar] [CrossRef] [PubMed]
- Albert, K.M.; Newhouse, P.A. Estrogen, stress, and depression: Cognitive and biological interactions. Annu. Rev. Clin. Psychol. 2019, 15, 99–423. [Google Scholar] [CrossRef]
- Echeverria, V.; Echeverria, F.; Barreto, G.E.; Echeverría, J.; Mendoza, C. Estrogenic plants: To prevent neurodegeneration and memory loss and other symptoms in women after menopause. Front. Pharmacol. 2021, 12, 644103. [Google Scholar] [CrossRef]
- Agca, C.; Klakotskaia, D.; Stopa, E.G.; Schachtman, T.R.; Agca, Y. Ovariectomy Influences Cognition and Markers of Alzheimer’s Disease. J. Alzheimer’s Dis. 2020, 73, 529–541. [Google Scholar] [CrossRef]
- Rocca, W.A.; Grossardt, B.R.; Shuster, L.T. Oophorectomy, menopause, estrogen treatment, and cognitive aging: Clinical evidence for a window of opportunity. Brain Res. 2011, 1379, 188–198. [Google Scholar] [CrossRef] [Green Version]
- Patchev, V.K.; Almeida, O.F. Gonadal steroids exert facilitating and “buffering” effects on glucocorticoid-mediated transcriptional regulation of corticotropin-releasing hormone and corticosteroid receptor genes in rat brain. J. Neurosci. 1996, 16, 7077–7084. [Google Scholar] [CrossRef]
- Baxi, D.B.; Singh, P.K.; Vachhrajani, K.D.; Ramachandran, A.V. Melatonin supplementation in rat ameliorates ovariectomy-induced oxidative stress. Climacteric 2013, 16, 274–283. [Google Scholar] [CrossRef]
- Engler-Chiurazzi, E.B.; Brown, C.M.; Povroznik, J.M.; Simpkins, J.W. Estrogens as neuroprotectants: Estrogenic actions in the context of cognitive aging and brain injury. Prog. Neurobiol. 2017, 157, 188–211. [Google Scholar] [CrossRef] [Green Version]
- Benedusi, V.; Meda, C.; Della Torre, S.; Monteleone, G.; Vegeto, E.; Maggi, A. A lack of ovarian function increases neuroinflammation if aged mice. Endocrinology 2012, 153, 2153–2788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marongiu, R. Accelerated ovarian failure as a unique model to study peri-menopause influence on Alzheimer’s disease. Front. Aging Neurosci. 2019, 11, 242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarrel, P.M.; Sullivan, S.D.; Nelson, L.M. Hormone replacement therapy in young women with surgical primary ovarian insufficiency. Fertil. Steril. 2016, 106, 1580–1587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordhandas, S.; Norquist, B.M.; Pennington, K.P.; Yung, R.L.; Laya, M.B.; Swisher, E.M. Hormone replacement therapy after risk reducing salpingo-oophorectomy in patients with BRCA1 or BRCA2 mutations; a systematic review of risks and benefits. Gynecol. Oncol. 2019, 153, 192–200. [Google Scholar] [CrossRef]
- He, K. Traditional Chinese and Thai medicine in a comparative perspective. Complement. Ther. Med. 2015, 23, 821–826. [Google Scholar] [CrossRef]
- Saniotis, A. Changing ethics in medical practice: A Thai perspective. Indian J. Med. Ethics 2007, 4, 24–25. [Google Scholar] [CrossRef]
- Malaivijitnond, S. Medical applications of phytoestrogens from the Thai herb Pueraria mirifica. Front. Med. 2012, 6, 8–21. [Google Scholar] [CrossRef]
- Bopana, N.; Saxena, S. Asparagus racemosus—Ethnopharmacological evaluation and conservation needs. J. Ethnopharmacol. 2007, 110, 1–15. [Google Scholar] [CrossRef]
- Pitiporn, S.; Tengtermwong, N. (Eds.) List of Herbal Medicine Products; Chao Phya Abhaibhubejhr Hospital, A.D.; Poramut Printing: Bangkok, Thailand, 2016; pp. 64–65. [Google Scholar]
- Tantipongpiradet, A.; Monthakantirat, O.; Daodee, S.; Boonyarat, C.; Matsumoto, K.; Pitiporn, S.; Chulikhit, Y. Yakae-prajamduen-jamod recipe reduced anxiety behavior and brain oxidative damage in ovariectomized mice. Songkla J. Sci. Technol. 2020, 42, 172–179. [Google Scholar]
- Monthakantirat, O.; Sukano, W.; Umehara, K.; Noguchi, H.; Chulikhit, Y.; Matsumoto, K. Effect of miroestrol on ovariectomy-induced cognitive impairment and lipid peroxidation in mouse brain. Phytomedicine 2014, 21, 1249–1255. [Google Scholar] [CrossRef]
- Fonseca, C.S.; Gusmão, I.D.; Raslan, A.C.; Monteiro, B.M.; Massensini, A.R.; Moraes, M.F.; Pereira, G.S. Object recognition memory and temporal lobe activation after delayed estrogen replacement therapy. Neurobiol. Learn. Mem. 2013, 101, 19–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkaki, A.; Amani, R.; Badavi, M.; Safahani, M.; Aligholi, H. Effect of ovariectomy on reference memory version of Morris water maze in young adult rats. Iran. Biomed. J. 2008, 12, 123–128. [Google Scholar] [PubMed]
- Lejri, I.; Grimm, A.; Eckert, A. Mitochondria, estrogen and female brain aging. Front. Aging Neurosci. 2018, 10, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, Y.; Waters, E.M.; McEwen, B.S.; Morrison, J.H. Estrogen effects on cognitive and synaptic health over the lifecourse. Physiol. Rev. 2015, 95, 785–807. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Sareddy, G.R.; Wang, J.; Wang, R.; Li, Y.; Dong, Y.; Zhang, Q.; Liu, J.; O’Connor, J.C.; Xu, J.; et al. Neuron-derived estrogen regulates synaptic plasticity and memory. J. Neurosci. 2019, 39, 2792–2809. [Google Scholar] [CrossRef]
- Rettberg, J.R.; Yao, J.; Brinton, R.D. Estrogen: A master regulator of bioenergetic systems in the brain and body. Front. Neuroendocrinol. 2014, 35, 8–30. [Google Scholar] [CrossRef] [Green Version]
- Uddin, M.S.; Rahman, M.M.; Jakaria, M.; Rahman, M.S.; Hossain, M.S.; Islam, A.; Ahmed, M.; Mathew, B.; Omar, U.; Barreto, G.; et al. Estrogen signaling in Alzheimer’s disease: Molecular insights and therapeutic targets for Alzheimer’s dementia. Mol. Neurobiol. 2020, 57, 2654–2670. [Google Scholar] [CrossRef]
- Chulikhit, Y.; Sukhano, W.; Daodee, S.; Putalun, W.; Wongpradit, R.; Khamphukdee, C.; Umehara, K.; Noguchi, H.; Matsumoto, K.; Monthakantirat, O. Effects of Pueraria candollei var mirifica (Airy Shaw and Suvat.) Niyomdham on ovariectomy-induced cognitive impairment and oxidative stress in the mouse brain. Molecules 2021, 26, 3442. [Google Scholar] [CrossRef]
- Kraeuter, A.K.; Guest, P.C.; Sarnyai, Z. The Y-Maze for assessment of spatial working and reference memory in mice. In Pre-Clinical Models. Methods in Molecular Biology; Guest, P.C., Ed.; Humana Press: New York, NY, USA, 2019; Volume 1916, pp. 105–111. [Google Scholar]
- Vorhees, C.V.; Williams, M.T. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef] [Green Version]
- Bouet, V.; Freret, T.; Schumann-Bard, P.; Boulouard, M. Novel object recognition test in rodents: Which roles for serotonin receptors? In Handbook of Object Novelty Recognition; Ennaceur, A., de Souza Silva, M.A., Eds.; Elsevier Academic Press: Cambridge, MA, USA, 2018; Volume 27, pp. 391–402. [Google Scholar]
- Khamphukdee, C.; Turkmani, I.; Chotritthirong, Y.; Chulikhit, Y.; Boonyarat, C.; Sekeroglu, N.; Silva, A.M.S.; Monthakantirat, O.; Kijjoa, A. Effects of the Bark Resin Extract of Garcinia nigrolineata on Chronic Stress-Induced Memory Deficit in Mice Model and the In Vitro Monoamine Oxidases and β-amyloid aggregation inhibitory activities of its prenylated xanthone constituents. Molecules 2022, 27, 3014. [Google Scholar] [CrossRef]
- Ding, F.; Yao, J.; Zhao, L.; Mao, Z.; Chen, S.; Brinton, R.D. Ovariectomy induces a shift in fuel availability and metabolism in the hippocampus of the female transgenic model of familial Alzheimer’s. PLoS ONE 2013, 8, e59825. [Google Scholar] [CrossRef] [PubMed]
- Tecalco-Cruz, A.C.; Zepeda-Cervantes, J.; Ortega-Domínguez, B. Estrogenic hormones receptors in Alzheimer’s disease. Mol. Biol. Rep. 2021, 48, 7517–7526. [Google Scholar] [CrossRef] [PubMed]
- Zárate, S.; Stevnsner, T.; Gredilla, R. Role of Estrogen and other sex hormones in brain aging. Neuroprotection and DNA repair. Front. Aging Neurosci. 2017, 9, 430. [Google Scholar] [CrossRef] [Green Version]
- Ycaza, H.A.; Mather, M. Actions and interactions of estradiol and glucocorticoids in cognition and the brain: Implications for aging women. Neurosci. Biobehav. Rev. 2015, 55, 36–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Q.Q.; Yang, L.; Huang, H.J.; Wang, Y.L.; Yu, R.; Wang, J.; Pilot, A.; Wu, G.C.; Liu, Q.; Yu, J. Differential GR Expression and Translocation in the Hippocampus Mediates Susceptibility vs. Resilience to Chronic Social Defeat Stress. Front. Neurosci. 2017, 11, 287. [Google Scholar] [CrossRef]
- Maneenet, J.; Daodee, S.; Monthakantirat, O.; Boonyarat, C.; Khamphukdee, C.; Kwankhao, P.; Pitiporn, S.; Awale, S.; Chulikhit, Y.; Kijjoa, A. Kleeb Bua Daeng, a Thai traditional herbal formula, ameliorated unpredictable chronic mild stress-induced cognitive impairment in ICR mice. Molecules 2019, 24, 4587. [Google Scholar] [CrossRef] [Green Version]
- Saied, N.M.; Georgy, G.S.; Hussien, R.M.; Hassan, W.A. Neuromodulatory effect of curcumin on catecholamine systems and inflammatory cytokines in ovariectomized female rats. Clin. Exp. Pharmacol. Physiol. 2021, 48, 337–346. [Google Scholar] [CrossRef]
- Khamphukdee, C.; Monthakantirat, O.; Chulikhit, Y.; Boonyarat, C.; Daodee, S.; Aon-Im, P.; Maneenet, J.; Chotritthirong, Y.; Luecha, P.; Sekeroglu, N.; et al. Antidementia Effects of Alternanthera philoxeroides in Ovariectomized Mice Supported by NMR-Based Metabolomic Analysis. Molecules 2021, 26, 2789. [Google Scholar] [CrossRef]
- Villa, A.; Vegeto, E.; Poletti, A.; Maggi, A. Estrogens, Neuroinflammation, and Neurodegeneration. Endocr. Rev. 2016, 37, 372–402. [Google Scholar] [CrossRef] [Green Version]
- Kireev, R.A.; Vara, E.; Viña, J.; Tresguerres, J.A. Melatonin and oestrogen treatments were able to improve neuroinflammation and apoptotic processes in dentate gyrus of old ovariectomized female rats. Age 2014, 36, 9707. [Google Scholar] [CrossRef] [Green Version]
- Dixon, R.A. Phytoestrogens. Annu. Rev. Plant Biol. 2004, 55, 225–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swar, G.; Shailajan, S.; Menon, S. Activity based evaluation of a traditional Ayurvedic medicinal plant: Saraca asoca (Roxb.) de Wilde flowers as estrogenic agents using ovariectomized rat model. J. Ethnopharmacol. 2017, 195, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Fang, F.; Wang, J.; Wong, C.W. Structural activity relationship of flavonoids with estrogen-related receptor gamma. FEBS Lett. 2010, 584, 22–26. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Chang, E.J.; Kim, H.J.; Park, J.H.; Choi, S.W. Antioxidative flavonoids from leaves of Carthamus tinctorius. Arch. Pharm. Res. 2002, 25, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.; Sharma, N.; Oladeji, O.S.; Sourirajan, A.; Dev, K.; Zengin, G.; El-Shazly, M.; Kumar, V. Traditional uses, bioactive composition, pharmacology, and toxicology of Phyllanthus emblica fruits: A comprehensive review. J. Ethnopharmacol. 2022, 282, 114570. [Google Scholar] [CrossRef]
- Ingkaninan, K.; IJzerman, A.P.; Verpoorte, R. Luteolin, a compound with adenosine A(1) receptor-binding activity, and chromone and dihydronaphthalenone constituents from Senna siamea. J. Nat. Prod. 2000, 63, 315–317. [Google Scholar] [CrossRef]
- Huang, G.; Tong, Y.; He, Q.; Wang, J.; Chen, Z. Aucklandia lappa DC. extract enhances gefitinib efficacy in gefitinib-resistance secondary epidermal growth factor receptor mutations. J. Ethnopharmacol. 2017, 206, 353–362. [Google Scholar] [CrossRef]
- Mahleyuddin, N.N.; Moshawih, S.; Ming, L.C.; Zulkifly, H.H.; Kifli, N.; Loy, M.J.; Sarker, M.M.R.; Al-Worafi, Y.M.; Goh, B.H.; Thuraisingam, S.; et al. Coriandrum sativum L.: A Review on Ethnopharmacology, Phytochemistry, and Cardiovascular Benefits. Molecules 2021, 27, 209. [Google Scholar] [CrossRef]
- Taheri, Y.; Herrera-Bravo, J.; Huala, L.; Salazar, L.A.; Sharifi-Rad, J.; Akram, M.; Shahzad, K.; Melgar-Lalanne, G.; Baghalpour, N.; Tamimi, K.; et al. Cyperus spp.: A Review on Phytochemical Composition, Biological Activity, and Health-Promoting Effects. Oxid. Med. Cell. Longev. 2021, 2021, 4014867. [Google Scholar] [CrossRef]
- Hannan, M.A.; Rahman, M.A.; Sohag, A.A.M.; Uddin, M.J.; Dash, R.; Sikder, M.H.; Rahman, M.S.; Timalsina, B.; Munni, Y.A.; Sarker, P.P.; et al. Black Cumin (Nigella sativa L.): A comprehensive review on phytochemistry, health benefits, molecular pharmacology, and safety. Nutrients 2021, 13, 1784. [Google Scholar] [CrossRef]
- Zhao, T.; Li, S.J.; Zhang, Z.X.; Zhang, M.L.; Shi, Q.W.; Gu, Y.C.; Dong, M.; Kiyota, H. Chemical constituents from the genus Saussurea and their biological activities. Heterocycl. Commun. 2017, 23, 331–358. [Google Scholar] [CrossRef]
- Ekiert, H.; Świątkowska, J.; Klin, P.; Rzepiela, A.; Szopa, A. Artemisia annua—Importance in Traditional Medicine and Current state of knowledge on the chemistry, biological activity and possible applications. Planta Med. 2021, 87, 584–599. [Google Scholar] [PubMed]
- Zhou, W.; Hu, M.; Hu, J.; Du, Z.; Su, Q.; Xiang, Z. Luteolin suppresses microglia neuroinflammatory responses and relieves inflammation-induced cognitive impairments. Neurotox. Res. 2021, 39, 1800–1811. [Google Scholar] [CrossRef] [PubMed]
- Zheng, N.; Yuan, P.; Li, C.; Wu, J.; Huang, J. Luteolin reduces BACE1 expression through NF-κB and through estrogen receptor mediated pathways in HEK293 and SH-SY5Y cells. J. Alzheimer’s Dis. 2015, 45, 659–671. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, H.; Cheng, H.; Che, Z. Ameliorating effect of luteolin on memory impairment in an Alzheimer’s disease model. Mol. Med. Rep. 2016, 13, 4215–4220. [Google Scholar] [CrossRef] [Green Version]
- Thu, Z.M.; Myo, K.K.; Aung, H.T.; Armijos, C.; Vidari, G. Flavonoids and stilbenoids of the Genera Dracaena and Sansevieria: Structures and bioactivities. Molecules 2020, 25, 2608. [Google Scholar] [CrossRef]
- Wang, B.; Zhong, Y.; Gao, C.; Li, J. Myricetin ameliorates scopolamine-induced memory impairment in mice via inhibiting acetylcholinesterase and down-regulating brain iron. Biochem. Biophys. Res. Commun. 2017, 490, 336–342. [Google Scholar] [CrossRef]
- Nielsen, B.E.; Bermudez, I.; Bouzat, C. Flavonoids as positive allosteric modulators of α7 nicotinic receptors. Neuropharmacology 2019, 160, 107794. [Google Scholar] [CrossRef]
- Abd El-Fattah, A.I.; Fathy, M.M.; Ali, Z.Y.; El-Garawany, A.E.A.; Mohamed, E.K. Enhanced therapeutic benefit of quercetin-loaded phytosome nanoparticles in ovariectomized rats. Chem. Biol. Interact. 2017, 271, 30–38. [Google Scholar] [CrossRef]
- Aggarwal, A.; Sharma, N.; Khera, A.; Sandhir, R.; Rishi, V. Quercetin alleviates cognitive decline in ovariectomized mice by potentially modulating histone acetylation homeostasis. J. Nutr. Biochem. 2020, 84, 108439. [Google Scholar] [CrossRef]
- Liu, E.Y.L.; Xu, M.L.; Jin, Y.; Wu, Q.; Dong, T.T.X.; Tsim, K.W.K. Genistein, a phytoestrogen in soybean, induces the expression of acetylcholinesterase via G protein-coupled receptor 30 in PC12 cells. Front. Mol. Neurosci. 2018, 11, 59. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, M.; Joghataei, M.T.; Mohseni, S.; Roghani, M. Genistein ameliorates learning and memory deficits in amyloid β(1–40) rat model of Alzheimer’s disease. Neurobiol. Learn. Mem. 2011, 95, 270–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ervin, K.S.; Lymer, J.M.; Matta, R.; Clipperton-Allen, A.E.; Kavaliers, M.; Choleris, E. Estrogen involvement in social behavior in rodents: Rapid and long-term actions. Horm. Behav. 2015, 74, 53–76. [Google Scholar] [CrossRef] [PubMed]
- Goel, B.; Tripathi, N.; Bhardwaj, N.; Sahu, B.; Jain, S.K. Therapeutic Potential of Genus Pongamia and Derris: Phytochemical and Bioactivity. Mini Rev. Med. Chem. 2021, 21, 920–951. [Google Scholar] [CrossRef]
- Wang, C.; Li, C.; Zhou, H.; Huang, J. High-throughput screening assays for estrogen receptor by using coumestrol, a natural fluorescence compound. J. Biomol. Screen. 2014, 19, 253–258. [Google Scholar] [CrossRef]
- Moreira, A.C.; Silva, A.M.; Branco, A.F.; Baldeiras, I.; Pereira, G.C.; Seiça, R.; Santos, M.S.; Sardão, V.A. Phytoestrogen coumestrol improves mitochondrial activity and decreases oxidative stress in the brain of ovariectomized Wistar-Han rats. J. Funct. Foods 2017, 34, 329–339. [Google Scholar] [CrossRef]
- Cornwell, T.; Cohick, W.; Raskin, I. Dietary phytoestrogens and health. Phytochemistry 2004, 65, 995–1016. [Google Scholar] [CrossRef]
- Tantipongpiradet, A.; Monthakantirat, O.; Vipatpakpaiboon, O.; Khampukdee, C.; Umehara, K.; Noguchi, H.; Fujiwara, H.; Matsumoto, K.; Sekeroglu, N.; Kijjoa, A.; et al. Effects of puerarin on the ovariectomy-induced depressive-like behavior in ICR mice and its possible mechanism of action. Molecules 2019, 24, 4569. [Google Scholar] [CrossRef] [Green Version]
- Nuutila, A.M.; Kammiovirta, K.; Oksman-Caldentey, K.-M. Comparison of methods for the hydrolysis of flavonoids and phenolic acids from onion and spinach for HPLC analysis. Food Chem. 2002, 76, 519–525. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species (Family) | Family Name | Voucher Specimen | Part Used | Proportion |
|---|---|---|---|---|
| Dracaena loureiri Gagnep | Asparagaceae | YPJ001 | heartwood | 5.56 |
| Tarenna hoaensis Pitard | Rubiaceae | YPJ002 | heartwood | 5.56 |
| Bridelia ovata Decne. | Phyllanthaceae | YPJ003 | leaves | 5.56 |
| Carthamus tinctorius L. | Asteraceae | YPJ004 | flower | 5.56 |
| Terminalia chebula Retz. var. chebula. | Combretaceae | YPJ005 | fruits | 5.56 |
| Terminalia arjuna Roxb. | Combretaceae | YPJ006 | fruits | 5.56 |
| Phyllanthus emblica L. | Phyllanthaceae | YPJ007 | fruits | 5.56 |
| Aloe vera (L.) Burm.f. | Asphodelaceae | YPJ008 | resin | 5.56 |
| Senna garrettiana (Craib) H.S.Irwin and Barneby | Fabaceae | YPJ009 | heartwood | 3.70 |
| Senna siamea (Lam.) H.S.Irwin and Barneby | Leguminosae | YPJ010 | heartwood | 3.70 |
| Derris scandens (Roxb.) Benth | Fabaceae | YPJ011 | stem | 3.70 |
| Caesalpinia sappan L. | Leguminosae | YPJ012 | heartwood | 3.70 |
| Mesua ferrea L. | Calophyllaceae | YPJ013 | flower | 3.70 |
| Mammea siamensis Kosterm | Calophyllaceae | YPJ014 | flower | 3.70 |
| Coriandrum sativum L. | Apiaceae | YPJ015 | fruits | 3.70 |
| Myristica fragrans Houtt. | Myristicaceae | YPJ016 | fruits | 3.70 |
| Amomum testaceum Ridl. | Zingiberaceae | YPJ017 | fruits | 3.70 |
| Cyperus rotundus Linn | Cyperaceae | YPJ018 | tuber | 3.70 |
| Nigella sativa L. | Ranunculaceae | YPJ019 | seed | 3.70 |
| Piper ribesoides Wall. | Piperaceae | YPJ020 | stem | 3.70 |
| Saussurea lappa Clarke | Asteraceae | YPJ021 | root | 3.70 |
| Artemisia annua L. | Asteraceae | YPJ022 | whole | 3.70 |
| Angelica sinensis | Apiaceae | YPJ023 | root | 3.70 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daodee, S.; Monthakantirat, O.; Tantipongpiradet, A.; Maneenet, J.; Chotritthirong, Y.; Boonyarat, C.; Khamphukdee, C.; Kwankhao, P.; Pitiporn, S.; Awale, S.; et al. Effect of Yakae-Prajamduen-Jamod Traditional Thai Remedy on Cognitive Impairment in an Ovariectomized Mouse Model and Its Mechanism of Action. Molecules 2022, 27, 4310. https://doi.org/10.3390/molecules27134310
Daodee S, Monthakantirat O, Tantipongpiradet A, Maneenet J, Chotritthirong Y, Boonyarat C, Khamphukdee C, Kwankhao P, Pitiporn S, Awale S, et al. Effect of Yakae-Prajamduen-Jamod Traditional Thai Remedy on Cognitive Impairment in an Ovariectomized Mouse Model and Its Mechanism of Action. Molecules. 2022; 27(13):4310. https://doi.org/10.3390/molecules27134310
Chicago/Turabian StyleDaodee, Supawadee, Orawan Monthakantirat, Ariyawan Tantipongpiradet, Juthamart Maneenet, Yutthana Chotritthirong, Chantana Boonyarat, Charinya Khamphukdee, Pakakrong Kwankhao, Supaporn Pitiporn, Suresh Awale, and et al. 2022. "Effect of Yakae-Prajamduen-Jamod Traditional Thai Remedy on Cognitive Impairment in an Ovariectomized Mouse Model and Its Mechanism of Action" Molecules 27, no. 13: 4310. https://doi.org/10.3390/molecules27134310